
8-oxoguanine and 8-oxodeoxyguanosine Biomarkers of Oxidative DNA Damage: A Review on HPLC–ECD Determination


1
University of Coimbra, Centre for Mechanical Engineering, Materials and Processes (CEMMPRE), Department of Chemistry, 3004-535 Coimbra, Portugal
2
Instituto Pedro Nunes (IPN), 3030-199 Coimbra, Portugal
Molecules 2022, 27(5), 1620; https://doi.org/10.3390/molecules27051620
Submission received: 18 January 2022
/
Revised: 9 February 2022
/
Accepted: 11 February 2022
/
Published: 1 March 2022
(This article belongs to the Special Issue Theme Issue in Honor of Professor Ana Maria Oliveira Brett: Bioelectrochemical Sensing and DNA Electrochemical Biosensors)
Abstract
:Reactive oxygen species (ROS) are continuously produced in living cells due to metabolic and biochemical reactions and due to exposure to physical, chemical and biological agents. Excessive ROS cause oxidative stress and lead to oxidative DNA damage. Within ROS-mediated DNA lesions, 8-oxoguanine (8-oxoG) and its nucleotide 8-oxo-2′-deoxyguanosine (8-oxodG)—the guanine and deoxyguanosine oxidation products, respectively, are regarded as the most significant biomarkers for oxidative DNA damage. The quantification of 8-oxoG and 8-oxodG in urine, blood, tissue and saliva is essential, being employed to determine the overall effects of oxidative stress and to assess the risk, diagnose, and evaluate the treatment of autoimmune, inflammatory, neurodegenerative and cardiovascular diseases, diabetes, cancer and other age-related diseases. High-performance liquid chromatography with electrochemical detection (HPLC–ECD) is largely employed for 8-oxoG and 8-oxodG determination in biological samples due to its high selectivity and sensitivity, down to the femtomolar range. This review seeks to provide an exhaustive analysis of the most recent reports on the HPLC–ECD determination of 8-oxoG and 8-oxodG in cellular DNA and body fluids, which is relevant for health research.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
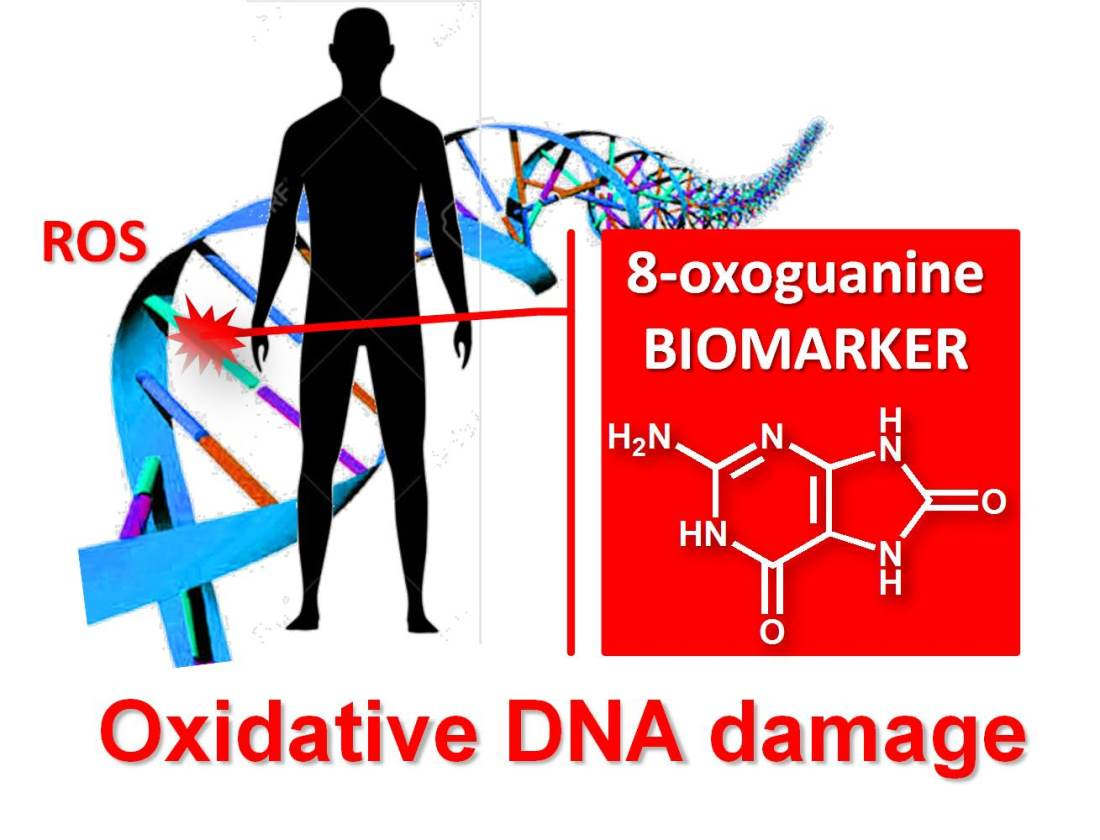
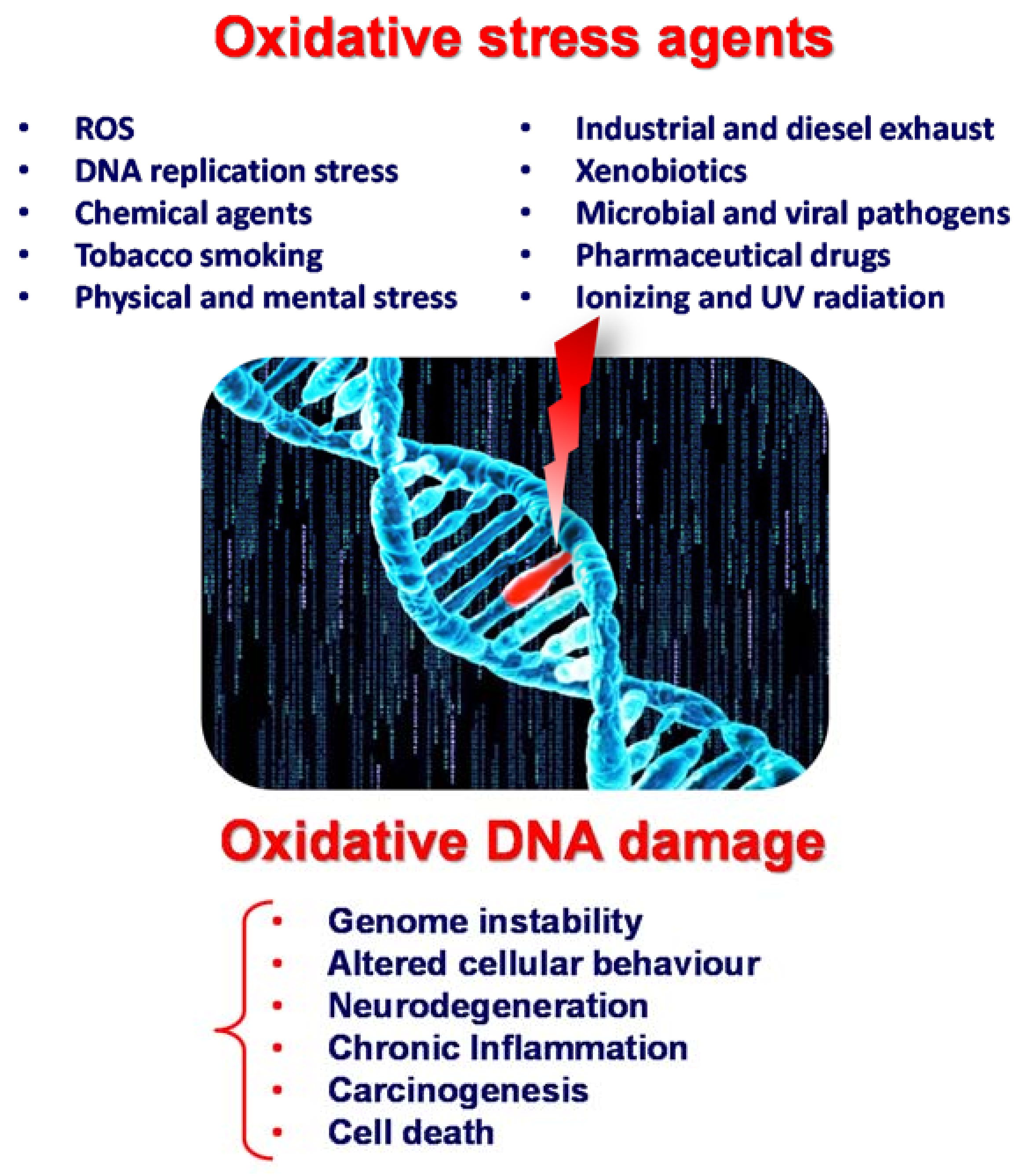
Reactive oxygen species (ROS) are produced endogenously due to cellular metabolism or exogenously due to exposure to physical, chemical, or biological agents such as γ, X-ray and UV radiation, tobacco smoking, pollutants, toxins, xenobiotics, and bacterial and viral pathogens; see Scheme 1. Although ROS exhibit a beneficial role at the cellular level, being involved in inter- and intracellular signalling, excessive ROS cause oxidative stress and lead to more than 100 different types of oxidative DNA lesions in mammals, including DNA base oxidation, double- and single-strand breaks, intra- and interstrand crosslinks, and abasic sites.
Leaving oxidative DNA damage unrepaired results in changes or mutations within the cell genomic material, leading to genomic instability, changes in gene expression, altered cellular behaviour, neurodegeneration, chronic inflammation, carcinogens and (ultimately) cell death; see Scheme 1. Hence, many diseases are correlated with oxidative DNA damage, e.g., neurodegenerative, cardiovascular, and autoimmune inflammatory diseases; diabetes; cancer; and other age-related diseases.
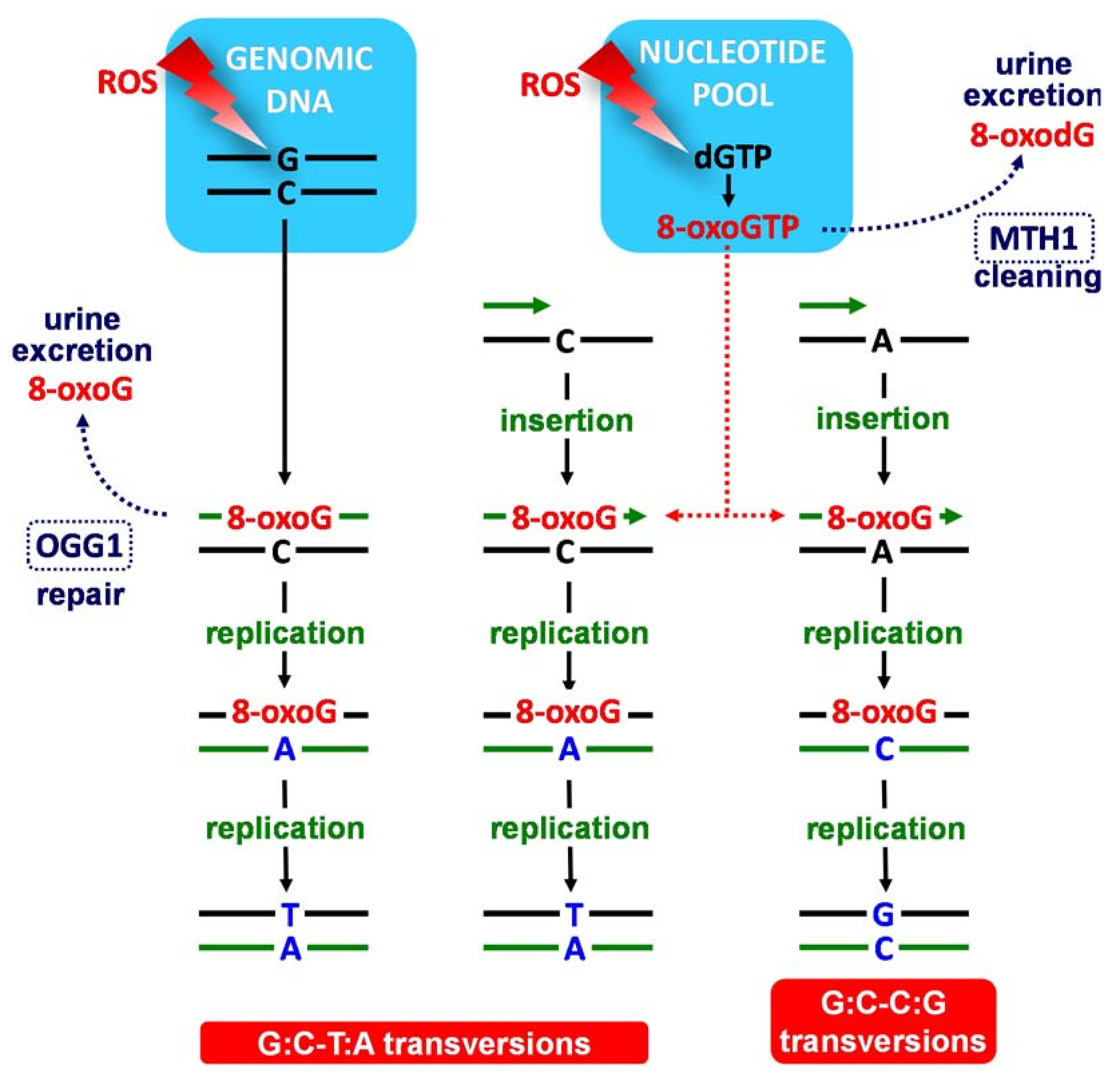
The oxidation of the DNA bases by ROS can occur either directly in the genomic DNA strands or indirectly in the nucleotide pool, from where the modified bases are introduced into the genomic DNA during replication or repair; see Scheme 2 [1,2,3].
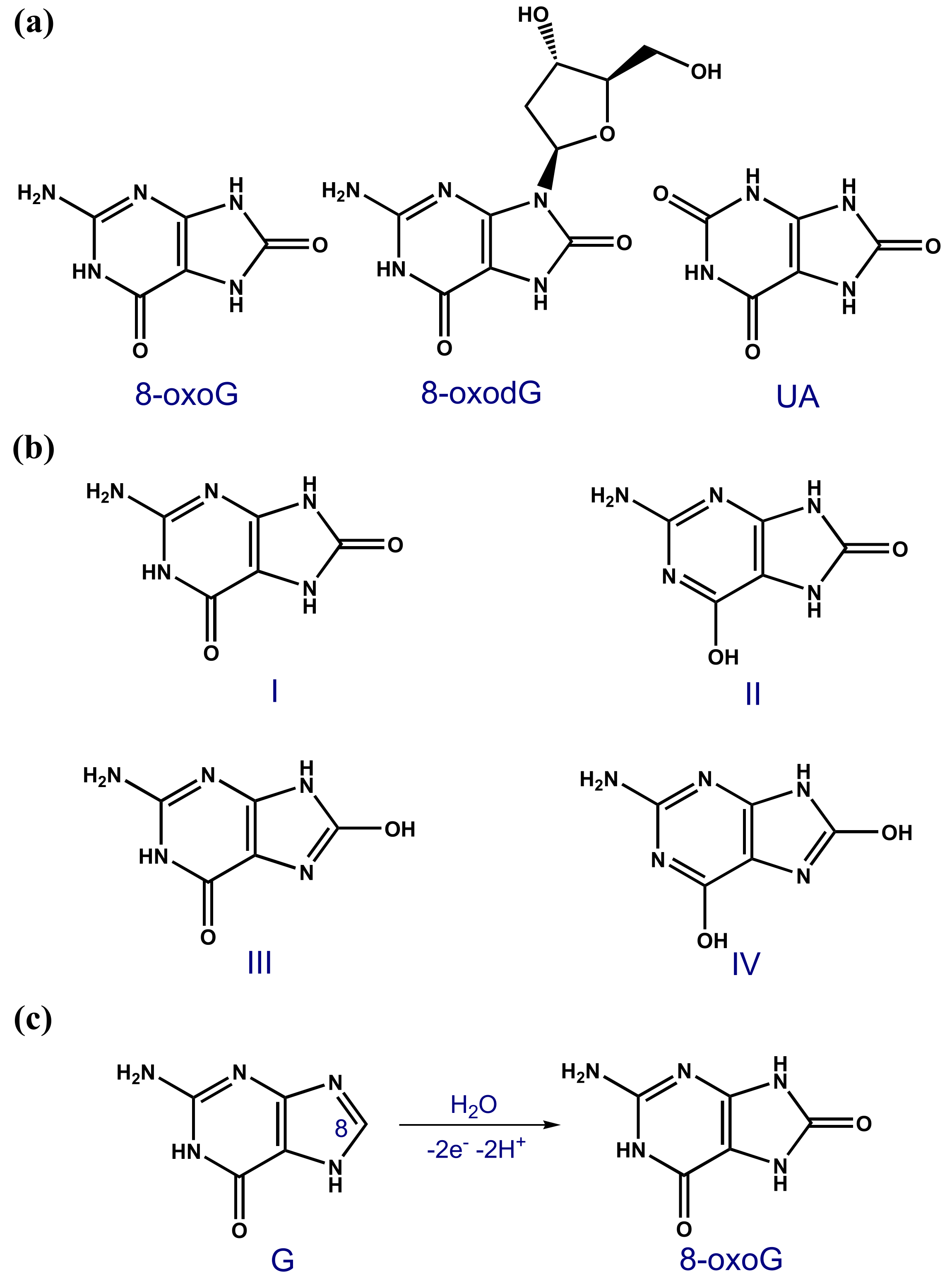
The guanine (G) base is easier oxidised than adenine (A), cytosine (C) and thymine (T) [4], so its oxidation products are the most common forms of oxidative DNA damage [5,6,7]. 8-oxoguanine (8-oxoG or 8-oxoGua), also named 7,8-dihydro-8-oxoguanine, 8-dihydroguanine, or 8-hydroxyguanine (8-OHG); see Scheme 3a [8], the G oxidation product at the C8 position; see Scheme 3c, and its nucleotide 8-oxo-2′-deoxyguanosine (8-oxodG), also named 8-oxo-7,8-dihydro-2′-deoxyguanosine or 8-hydroxy-2′-deoxyguanosine (8-OHdG); see Scheme 3a, are notable biomarkers used for the quantification of oxidation stress and oxidative DNA damage in animals and humans.
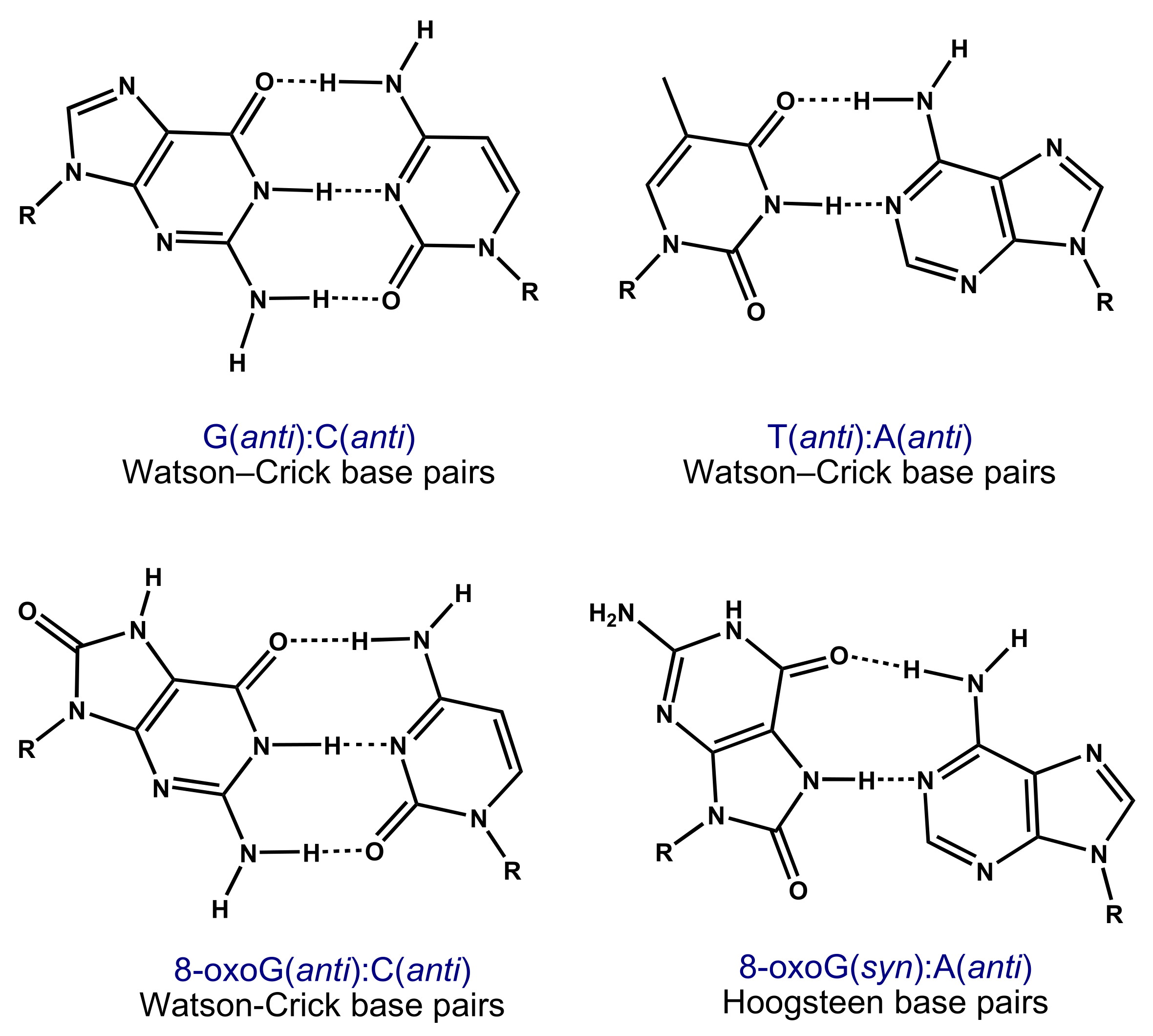
The 6,8-diketo I form of 8-oxoG, which presents an H atom at the N7 position and an oxo group at the C8 position, was predicted to be the most stable tautomer; see Scheme 3b [9,10,11,12]. The high mutagenic capacity of 8-oxoG is related to its ability to form (i) Watson–Crick base pairs with C (Scheme 4) leading to G:C–C:G transversions (Scheme 2) and (ii) Hoogsteen base pairs with A (Scheme 4) leading to G:C–T:A transversions in the course of the DNA replication (Scheme 2) [3,5].
The cellular defence system against 8-oxoG within a DNA strand involves the base excision repair (BER) that corrects small base lesions [11]; it begins with the oxoguanine glycosylase 1 (OGG1) that identifies and eliminates 8-oxoG, which is then excreted in urine; see Scheme 2 [2]. The nucleotide pool is sanitized by the nucleoside diphosphate linked moiety X type motif 1 (MTH1 or NUDT1) that hydrolyses 8-oxodGTP to 8-oxodGMP, leading to 8-oxodG and/or 8-oxodG excretion in urine; see Scheme 2 [2]. Thus, the urinary excretion of 8-oxoG and 8-oxodG from damaged DNA and the nucleotide pool reflects the equilibrium between in vivo oxidative DNA damage and the efficiency of the cellular repair processes. Apart from urinary excreted 8-oxodG/8-oxodG, significantly increased levels of 8-oxodG/8-oxodG in cancerous tissue DNA than in corresponding normal tissues has been observed, especially for individuals with impaired OGG1 repair function that exhibit increased disease aggressiveness.
Therefore, the possibility of quantifying 8-oxoG and/or 8-oxodG in urine, blood, tissue and saliva has received a growing interest; both 8-oxoG and/or 8-oxodG are regarded as the most significant biomarkers for oxidative DNA damage, being used to determine the body oxidative stress and to evaluate the risk, diagnose, and predict the treatment impact of various diseases.
The analytical techniques generally employed for the quantification of 8-oxoG and 8-oxodG levels in biological matrixes are high-performance liquid chromatography (HPLC) with spectrophotometric detection [11] and electrochemical detection (ECD), HPLC–ECD [13,14], HPLC tandem mass spectrometry (HPLC–MS) [15], gas chromatography–mass spectrometry (GC–MS) [14,16], enzyme-linked immunosorbent assay (ELISA) [11,17], and (more recently) electrochemical sensors [18]. Among these methods, HPLC–ECD is highly preferred due to its selectivity and high sensitivity, down to the femtomolar range.
This review seeks to provide an exhaustive analysis of the most recent advances on the determination of 8-oxoG and 8-oxodG biomarkers in cellular DNA and body fluids (urine, blood serum and saliva) using HPLC–ECD, which is relevant for health research.
2. Determination of 8-oxoG and 8-oxodG by HPLC–ECD
Early studies mostly concerned the quantification of 8-oxodG. The first report in 1986 described the use of a gradient reversed-phase HPLC coupled with ECD to quantify 8-oxodG in DNA hydrolysates at the sub-picomole levels [13]. The ECD analytical determination was revealed to be 1000 times more sensitive than the UV detection method [19], which made the quantification of 8-oxodG possible in HeLa cells [20], mouse liver [20], rat kidney [21,22], rat liver [23,24], rat heart [25], and rat urine [21], as well as in human leukocytes [26], blood mononuclear cells [27], embryonic cells [11], lung [28], brain [29], breast [27,30], colorectal tumour tissue [31], and urine [32,33,34,35].
Before 2000, hundreds of studies reported DNA damage in animal organs after the administration of various carcinogenic chemical substances and tumour promoters. The 8-oxodG content in human organs, leucocyte DNA, and urine have been analysed and correlated with various diseases, aging, exposure to ionizing radiation, and lifestyle parameters (e.g., smoking, exercise, and ingestion of antioxidants) [36].
To facilitate the HPLC–ECD analysis of 8-oxoG and 8-oxodG in urine, plasma, cerebrospinal fluid, saliva, leukocytes and tissue samples, different purification methods have been elaborated, e.g., solid-phase extraction (SPE) [33,34,37], and purification in different types of columns: reversed-phase [19,38,39], carbon [40], cation-exchange [38,41], and anion-exchange [39,42,43] columns, coupled-columns (such as anion-exchange coupled with reversed-phase [44]), multifunction columns (such as with gel filtration, reversed-phase and cation-exchange [45]), and columns enriched with polyclonal [46] or monoclonal antibodies [47,48].
Concerning electrochemical conditions, the ECD detection of 8-oxodG initially employed fixed potentials between +0.10 and +0.40 V [48,49,50,51]. Later, the oxidation of other DNA components with similar retention times at ~0.38 V was observed, which led to 8-oxodG overestimation. Therefore, an optimum applied potential of ~ +0.25 V was found to reduce overlapping peaks and resolve the 8-oxodG overestimation problem [52]. The HPLC–ECD detection of 8-oxoG has generally been less employed than 8-oxodG, although 8-oxoG presents a lower oxidation potential than 8-oxodG [53,54,55,56].
2.1. Determination of 8-oxoG and 8-oxodG in DNA
8-oxodG determination in DNA from leukocytes and tissues samples requires a preliminary step of hydrolysis (usually enzymatic) to break down the DNA prior to HPLC separation and electrochemical (amperometric, coulometric, or voltammetric) detection. The results are usually given in terms of the molar ratio of detected 8-oxodG per 105 undamaged deoxyguanosine (8-oxodG/105 dG) [57]. In this context, the HPLC–ECD technique has shown two methodological problems: (1) the occurrence of the artefact oxidation of unmodified G bases during the DNA isolation and hydrolysis steps, especially in the case of enzymatic digestion, leading to the overestimation of 8-oxodG levels [58], and (2) the incomplete hydrolysis that leads to the underestimation of the oxidative damage. Both problems have resulted in a lack of consistency in the results among different laboratories, especially concerning the overestimation of the 8-oxodG background levels in human cellular DNA [59,60].
Different factors influencing the DNA isolation and sample processing for HPLC–ECD have been evaluated, such as (1) the utilization of phenol, (2) the utilisation of chaotropic techniques, (3) the examination of very small DNA concentrations (less than 20 µg), (4) the DNA hydrolysis time, (5) the existence of redox-active metals, (6) the parameters used in the DNA extraction, (7) the chromatographic interference, (8) the adduct recovery during immunoaffinity purification, and (9) the formation of oxidized G derivatives by xanthine oxidase [49]. The use of phenol has been particularly discussed, as it may act as pro-oxidant during DNA extraction [49,61]. Other artefacts could be associated with the prolonged digestion of proteins during DNA extraction and prolonged DNA hydrolysis [49].
To solve the HPLC–ECD issues, especially the artefact oxidation occurring during isolation and purification, The European Standards Committee on Oxidative DNA Damage (ESCODD), with 27 analytical laboratories as members, was established [62]. The 8-oxodG levels in 8-oxodG standard samples, calf thymus DNA, oligonucleotides, HeLa cells and blood lymphocytes have been tested [59,60,62,63,64,65]. Based on controlled methodologies, coulometric and amperometric HPLC–ECD has been used to determine the 8-oxodG/105 dG ratio, and the results have been compared with the HPLC–MS/MS and GC–MS results, and mean values among all techniques were established [65,66,67].
Different methods have been proposed and many protocols have been revised to reduce the above-mentioned problems, including (i) the development of non-phenol DNA extraction methods [11,49,50,51,68] (e.g., chaotropic NaI [49] or DNAzol protocols [68]), (ii) the limitation of the incubation time [69], (iii) the use of cold workup procedures [50,69], and (iv) the use of antioxidants and metal chelators during sample preparation [49,52,70,71]. The use of phenol has been particularly controversial, although different studies showed that phenol extraction may not be responsible for 8-oxodG overestimation, which is instead caused by the sample impurities combined with excessive air exposure [49,69]. The use of desferrioxamine (brand name Desferal®), which chelates traces of redox-active iron, has been found to reduce the problems related with the duration of the hydrolysis process and protein digestion during DNA extraction [49,71], and the use of NaI or guanidine thiocyanate has been found to improve the problems related with DNA precipitation [50,71]. The use of careful HPLC hygiene during analysis helped reducing the artefacts resulting from the analysis of very small quantities of DNA [49].
8-oxodG levels in human leukocytes were used to assess oxidative DNA damage and found to be correlated with smoking, physical exercise, and alcohol consumption [72]. The mean molar 8-oxodG/105 dG ratio was significantly higher in smokers (33.1 ± 10.6) compared to non-smokers (15.3 ± 1.8) and former smokers (17.8 ± 1.5), the highest values being observed for individuals smoking over 10 cigarettes per day (41.8 ± 17.1). The frequency of physical exercise or alcohol drinking did not significantly modify 8-oxodG levels in leukocytes [72]. In a different report, lower mean 8-oxodG/105 dG levels in leukocytes—2.86 ± 1.39 in smokers and 2.72 ± 1.36 in non-smokers—were obtained [73].
Vitamin C’s effects against intracellular ROS production were investigated using HPLC–ECD with a modified pronase/ethanol extraction method [74], and it was shown that the mean 8-oxodG/106 dG ratio in the cellular DNA of peripheral blood lymphocytes decreased from 22.9 to 18.8 following 8 weeks of vitamin C supplementation. The decrease was also observed following vitamin C supplementation in four subgroups: (1) from 17.2 to 14.6 in subjects with less than 500 µg L−1 of ferritin, (2) from 29.1 to 23.3 in subjects with more than 500 µg L−1 of ferritin, (3) from 21.6 to 17.2 in subjects with a transferrin saturation (TSAT) of less than 50% and (4) from 23.8 to 17.8 in subjects with a TSAT of above 50%, but no significant modifications were observed in subjects treated with a placebo [74]. In a different report, using a purification method of genomic DNA from human whole blood via isopropanol-fractionation with concentrated NaI and sodium dodecyl sulphate, it was shown that 80, 200, and 400 mg of vitamin C supplementation had little effect on healthy individuals with diets abounding in vitamin C, with the mean 8-oxodG/106 dG levels remaining unaltered [75].
The 8-oxodG levels in human leukocytes have been used to determine of the role played by oxidative stress in the pathogenesis of Leber’s hereditary optic neuropathy (LHON) [76]. The mean 8-oxodG/105 dG ratio was found to be 1.34 ± 0.99 in patients with LHON with an 11778 mitochondrial DNA mutation, 1.00 ± 0.91 in their asymptomatic maternal relatives, and 0.31 ± 0.20 in normal control subjects, which suggested that the LHON phenotype might be the result of an increase in mitochondrial ROS caused by LHON mutations.
The increase in the oxidative DNA damage in living kidney donors, three years post-donation, was assessed, and a direct association between the pre-donation serum urea and creatinine and post-donation 8-oxodG/dG ratio was observed [77]. The study demonstrated a positive correlation between pre-donation serum urea and creatinine and post-donation oxidative DNA damage (8-oxodG/dG ratio) in kidney donors.
The 8-oxodG/105 dG ratio in different types of cancer tissues has been determined [78], e.g., breast cancer (2.07 ± 0 95 in cancer tissue vs. 1.34 ± 0 46 in control [79] and 10.7 ± 15.5 in cancer tissue vs. 6.3 ± 6.8 in control surrounding tissue [80]) and colorectal cancer (2.53 ± 0.15 in cancer tissue vs. 1.62 ± 0.13 in control [81], 49 in cancer tissue vs. 21 in control [82], 1.34 ± 0.11 in cancer tissue vs. 0.64 ± 0.05 in control [31], and 2.4 ± 1.1 in cancer tissue vs. 1.6 ± 0.6 in control [83]).
Different studies have reported changes in the mean 8-oxodG/105 dG ratios in cellular DNA induced by various environmental factors: (i) ambient particulate air pollution assessed as outdoor concentrations of particulate matter ≤2.5 μm in diameter (PM2.5) (0.55 in autumn, 0.27 in winter, 0.62 in spring, and 0.58 in summer, as determined in lymphocytes) [84], (ii) asbestos (2.61 ± 0.91 in individuals exposed in 1994–1995, 2.96 ± 1.10 in individuals exposed in 1995–1996, 2.55 ± 0.56 in individuals exposed in 1996–1997 vs. 1.52 ± 0.39 in unexposed controls, as determined in leukocytes) [85,86], (iii) silica (2.51 ± 1.36 in silicotics and 3.20 ± 2.25 in exposed workers, as determined in leukocytes) [87], (iv) polycyclic aromatic hydrocarbon (PAH) (1.24 ± 0.70 at the start and 1.57 ± 1.65 at the end of the week in roofers exposed to asphalt fume, 1.03 ± 0.71 at the start and 0.56 ± 0.24 at the end of the week in roofers exposed to coal-tar, 1.96 ± 0.83 at the start and 1.96 ± 0.83 at the end of the week in unexposed controls [88], and 4.31 at bottom, 3.08 at middle, and 3.07 at top PAH exposure in coke-oven workers at an iron-steel factory [89]), (v) KBrO [90], (vi) chloroform [90], (vii) bromodichloromethane [90], (viii) pulmonary carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) found in tobacco [91], (ix) mycotoxin aflatoxin B1 (AFB1) produced by Aspergillus fungi [92], and (x) γ-ray irradiation [93].
2.2. Determination of 8-oxoG and 8-oxodG in Body Fluids
The HPLC–ECD method of 8-oxoG and 8-oxodG determination in urine presents several advantages [94]: (i) the collection of urine samples is less invasive, (ii) the artifacts induced by the DNA isolation and sample processing steps are minimized, and (iii) 8-oxoG and 8-oxodG are very stable in urine, mimicking the equilibrium between oxidative DNA damage and cellular repair processes.
Later on, an European Standards Committee for Urinary (DNA) Lesion Analysis (ESCULA) was established to validate the assays and the value ranges of 8-oxodG in urine [95,96,97]. For urinary 8-oxodG biomarker detection, a correction of urine concentration is made, and the results are generally given as a ratio of 8-oxodG/creatinine concentration [95,98,99].
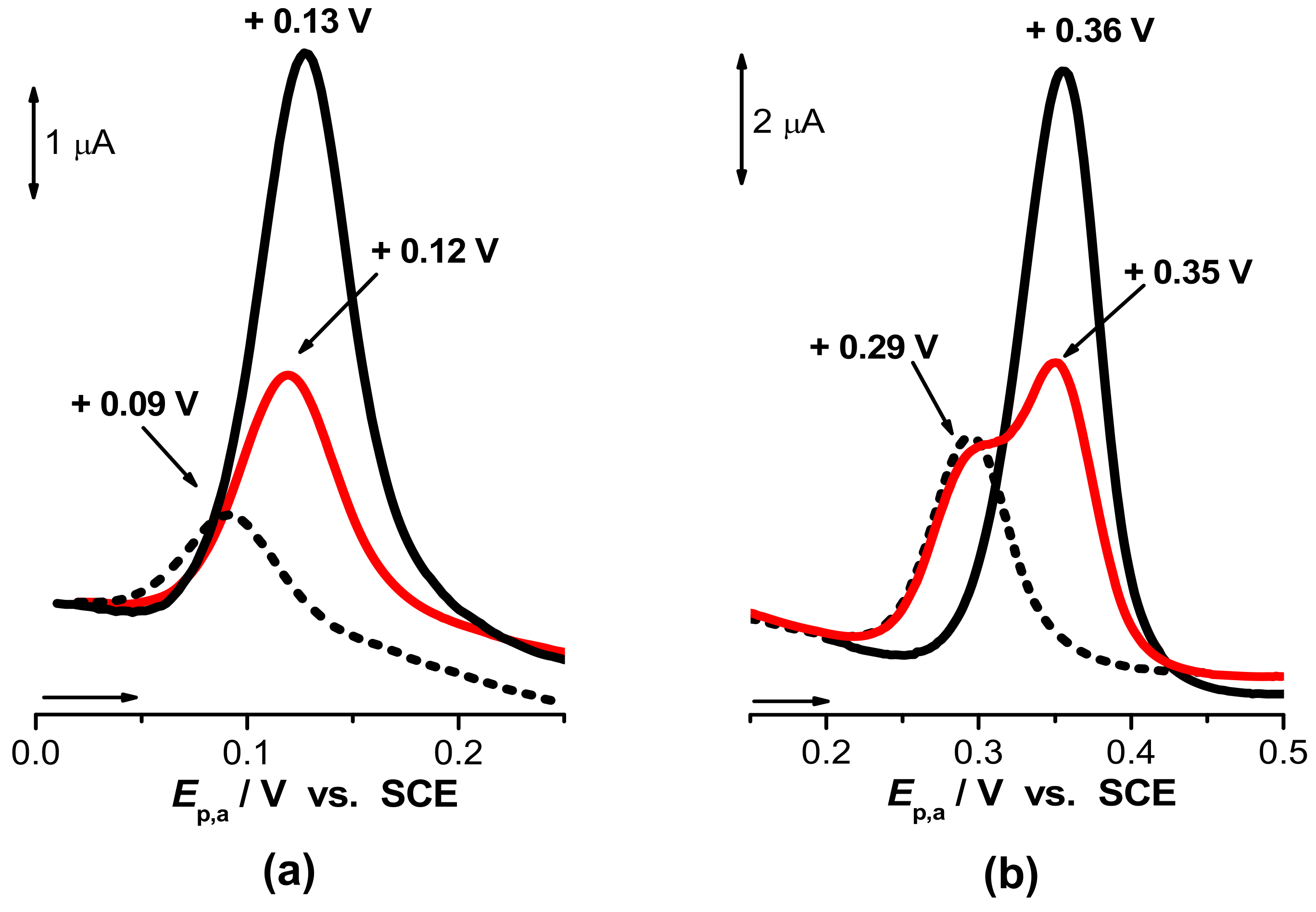
The separation of 8-oxoG and 8-oxodG from constituents of the urine matrix represents the most important challenge in the development of precise analytical determination methods, and different methodologies have been described [54,100,101]. In human urine, the uric acid (UA) concentration is 104-fold higher than the basal levels of 8-oxoG and 8-oxodG. 8-oxoG and UA present comparable chemical structures (Scheme 3), and the difference between their oxidation potentials is less than 0.1 V across the entire pH range; see Figure 1 [102]. In these conditions, 8-oxoG and 8-oxodG detection is especially difficult due to the proximity of their retention times.
Differential pulse voltammograms at a glassy carbon electrode showed that the best separation between the 8-oxoG and UA oxidation peaks was achieved for pH = 6.0; see Figure 1b [102].
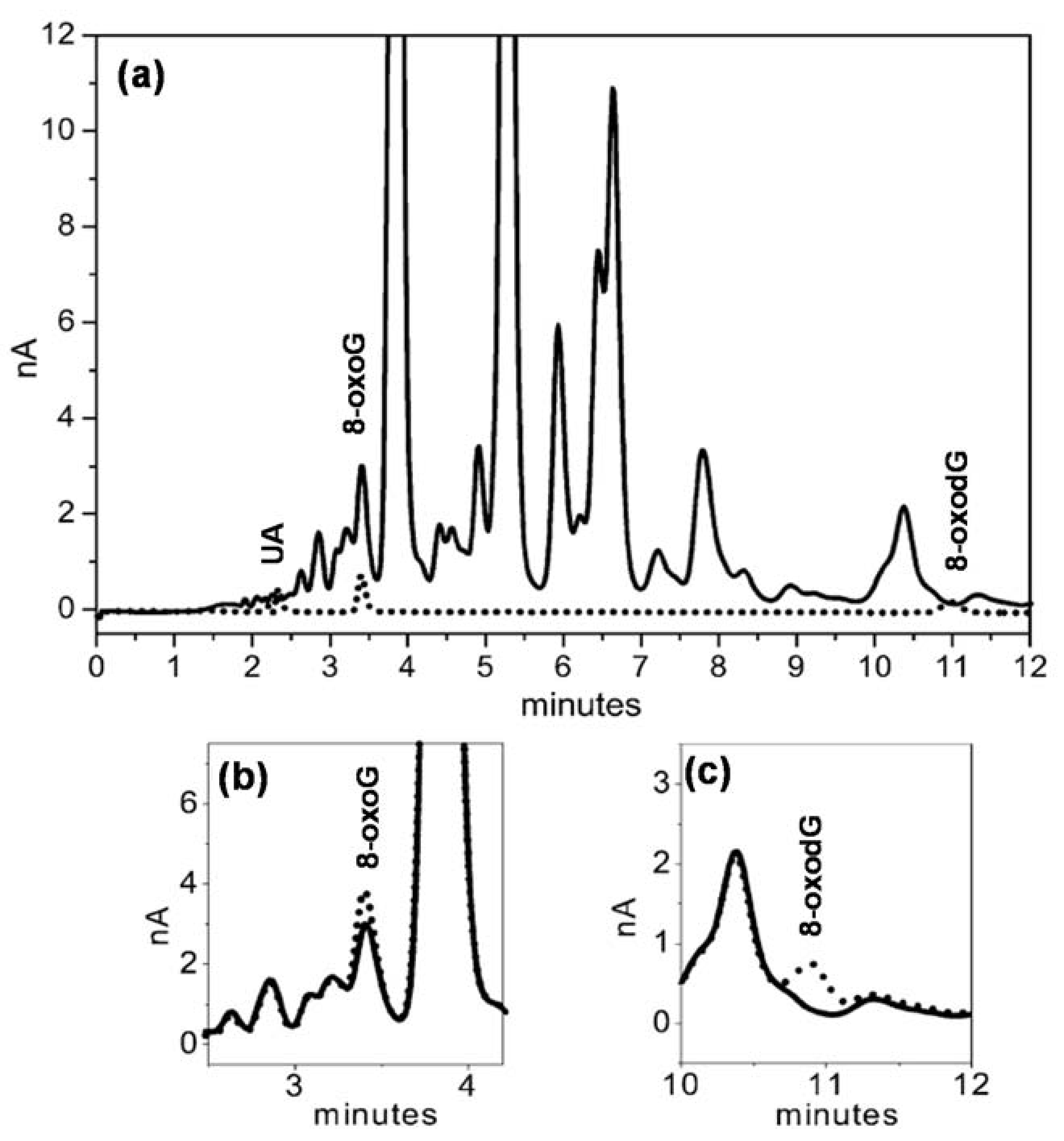
Based on these results, an HPLC–ECD method for the simultaneous detection of 8-oxoG and 8-oxodG with a good separation factor (α = 3.7) in excess UA conditions was developed [54]. This method consists of an HPLC isocratic elution with amperometric detection on a GCE electrode, enabling a limit of detection (LOD) for 8-oxoG and 8-oxodG lower than 1 nM in standard mixtures; see Figure 2. The detection of 25 nM concentrations of 8-oxoG and 8-oxodG in solutions containing 104-fold higher concentration of UA was achieved after one-step SPE and enhanced by uricase digestion. The method was tested in urine samples from children with metabolic disorders (see Figure 2) and adults with cognitive deficits, showing an LOD for 8-oxoG of 80 nM [54].
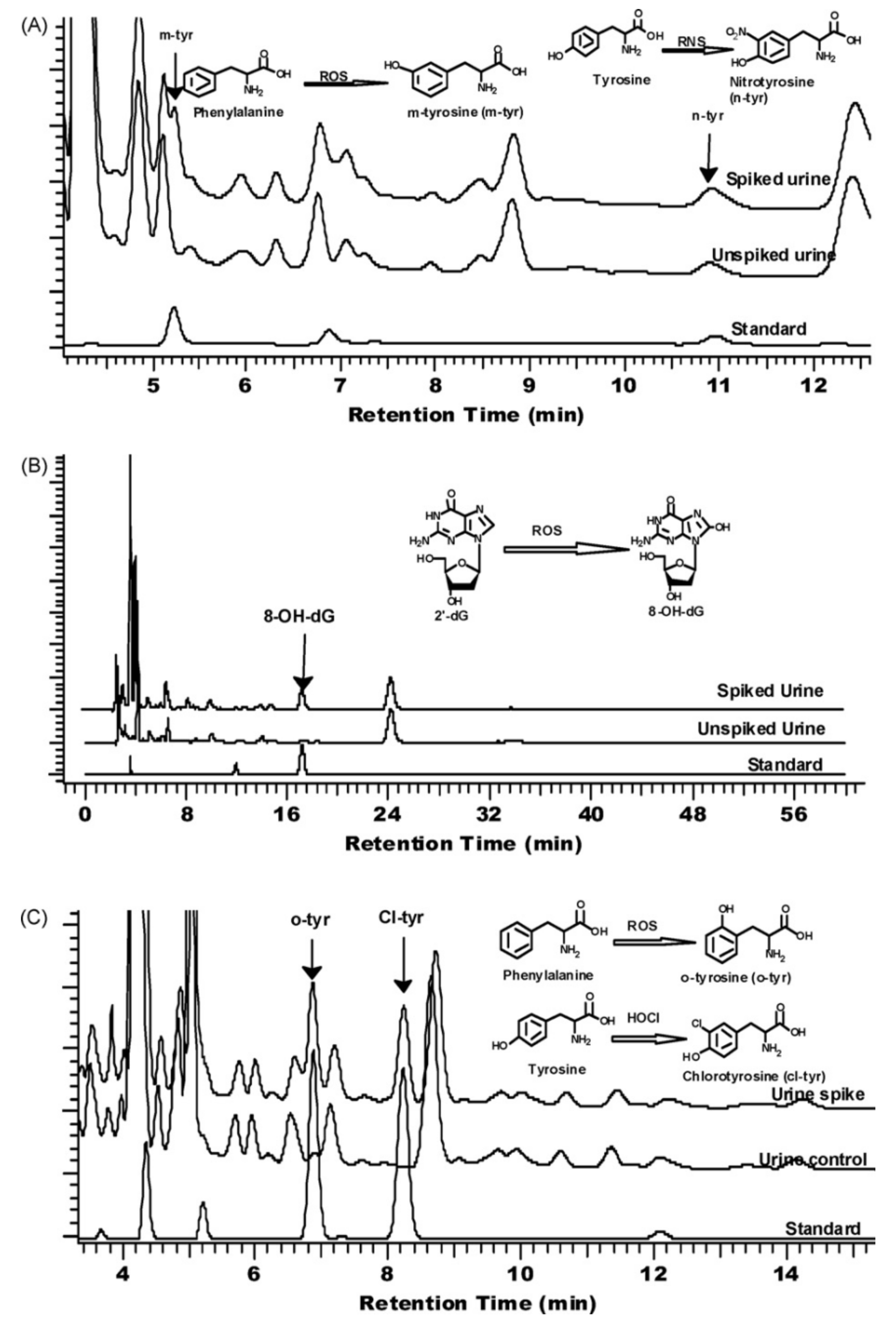
Based on a cleaning step by anion-exchange chromatography and a purification step by reversed-phase HPLC–ECD, the simultaneous determination of urinary 8-oxodG and creatinine [42] or 7-methylguanine [43] has been carried out. Moreover, based on a solid phase extraction method using a reversed-phase polymeric sorbent column prior to HPLC–ECD analysis, the simultaneous analysis of urinary oxidative/nitrative stress biomarkers o-, m-, cl-, 3-nitro-tyrosine, and 8-oxodG in human urine was achieved; see Figure 3 [103].
Different studies have reported baseline urinary 8-oxodG concentrations in healthy adults [94], e.g., 3.16 ± 1.28 µg/g of creatinine in men [104], 2.81 ± 1.07 µg/g of creatinine in men [105], 3.04 ± 1.42 µg/g of creatinine in women [105], 6.3 ± 0.5 µmol/mol of creatinine in men [106], 1.62 ± 0.50 µmol/mol of creatinine in man and women [107], and 4.70 ± 7.1 µmol/mol of creatinine in women [108,109].
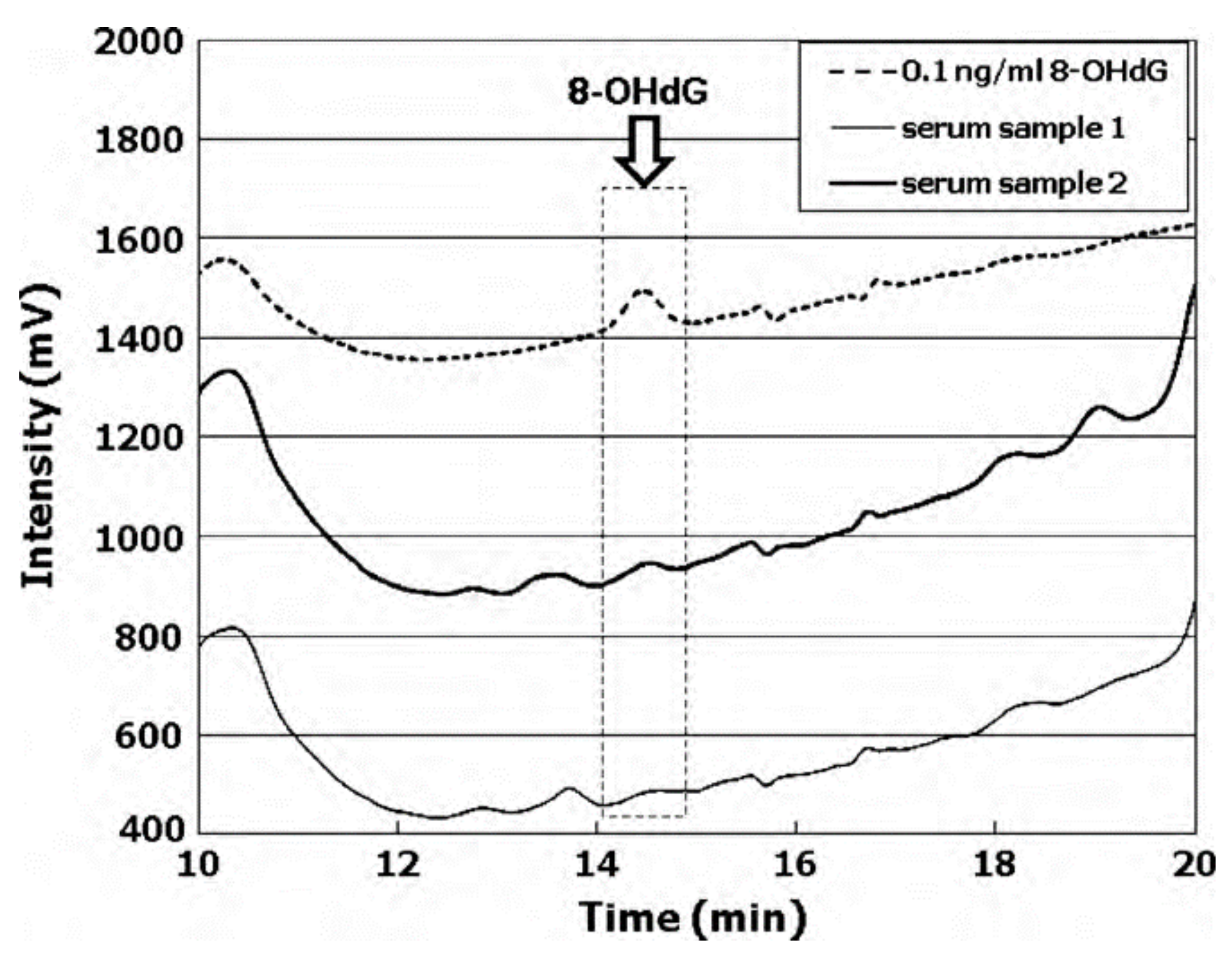
Based on an SPE method consisting on 8-oxodG separation from urine [100] and blood serum [37] in C18 and strong cation-exchange (SCX) columns, the most significant interfering compounds have been eliminated. For 8-oxodG in serum, an LOD in the control group of less than 10 pg mL−1 was achieved, eliminating the necessity for sample evaporation. The 8-oxodG levels in the sera from healthy individuals showed a mean value of 25.5 ± 13.8 pg mL−1; see Figure 4 [37].
8-oxoG levels measured in urine and serum showed a substantial increase in type 2 diabetic db/db mice when compared to control db/m+ mice, but no change in the 8-oxodG levels in the liver and kidney DNA was observed, suggesting that urinary and serum 8-oxodG/8-oxoG are better biomarkers of oxidative stress [98].
Employing the use of monoclonal antibodies [47], the serum 8-oxoG levels in type 2 diabetic patients was assessed and showed significantly higher concentrations (5.03 ± 0.69 nM) compared to control individuals (0.96 ± 0.15 nM) [110]. Moreover, among diabetic patients, those with proliferative retinopathy had significantly higher 8-oxoG levels (8.27 ± 0.31 nM) than those with non-proliferative retinopathy (4.92 ± 0.34 nM) or without retinopathy (3.38 ± 0.22 nM), but no correlation between the 8-oxoG level and duration of diabetes was established.
The influence of lifestyle factors on the urinary levels of 8-oxodG was determined [45,111,112,113]: the 8-oxodG level decreased with moderate physical exercise of less than 5 h/week (3.45 µg/g of creatinine) and high body mass index (BMI) values (3.41 µg/g of creatinine), while its level increased with physical labour (4.32 µg/g of creatinine), smoking (3.94 µg/g of creatinine), and low meat intake (less than once per week; 4.56 µg/g of creatinine) [45].
The urinary 8-oxodG level was also found to be directly correlated with high blood sugar levels (3.40 µg/g of creatinine) and leanness in smokers (3.40 µg/g of creatinine for BMI > 25 kg/m2 vs. 4.35 µg/g of creatinine for BMI < 18 kg/m2) [111]. Moreover, urinary 8-oxodG increased with exposure to organic solvents and hydrochloric acid (3.75 µg/g of creatinine) and long working hours habits (5.05 µg/g of creatinine). The urinary 8-oxodG level was found to be negatively correlated with high LDL cholesterol (3.05 µg/g of creatinine) [111], total cholesterol [114], and anaemia (1.40 µg/g of creatinine) [111]. At the individual level, no differences were detected in the diurnal urinary 8-oxodG levels, but changes have been observed with changes in lifestyle factors, e.g., exercise, sleep duration, drinking habits, diet, and stress [115,116], while depressive symptoms did not present a significant correlation with 8-oxodG [116].
In subjects enrolled in a quit-smoking program, the 8-oxodG levels decreased from 5.21 µg/g of creatinine before to 4.75 µg/g of creatinine after 2 weeks and 5.09 µg/g of creatinine after 8 weeks after the cessation of smoking, following decreases in the smoking exposure markers of nicotine, cotinine and 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanol (NNAL) [117].
Species with short potential life spans usually present higher metabolic rates correlated with higher oxidative DNA damage. The urinary 8-oxoG levels from mammals with different life spans (mice, rats, guinea pigs, cats, chimpanzees, and humans) showed a negative correlation with their potential life spans, while 8-oxodG showed only a slight tendency toward a negative correlation, which suggests that 8-oxoG may be a better overall biomarker for oxidative DNA damage than 8-oxodG [118,119].
The possibility of prediction of cancer risk was evaluated. Using an HPLC–ECD approach based on anion-exchange chromatography, precise fraction collection, and reversed-phase chromatography, it was established that 8-oxodG excretion in urine increased for on-going smokers (1.94 nmol/mmol of creatinine in subjects with lung cancer vs. 1.81 nmol/mmol of creatinine in controls), whilst a high 8-oxodG excretion among never-smokers was correlated with a higher risk of lung cancer development [39]. In a similar study, the urinary 8-oxodG/creatinine ratio was found to be significantly increased in smoking subjects (1.95 ± 0.40 µmol/mol) compared to non-smoking subjects (1.62 ± 0.50 µmol/mol) [107]. Moreover, high levels of the urinary 8-oxoG biomarker were correlated with higher lung cancer risk in men, never-smokers, and former smokers [120].
The urinary 8-oxodG levels of breast cancer patients at each stage of carcinogenesis were determined, and the researchers reported higher values of 16.39 ± 17.3 µmol/mol of creatinine in the breast cancer group compared to the 4.70 ± 7.1 µmol/mol of creatinine in the control group [108,109].
Many studies have reported comparative analyses of the 8-oxodG levels in both urine and cellular DNA. The HPLC–ECD detection of urinary 8-oxodG was used as a predictor for acute radiosensitivity in breast cancer patients [121], and both urinary 8-oxoG and leukocytes 8-oxodG biomarkers were used to predict the clinical success of radiotherapy treatment [122], due to the fact that high levels of urinary 8-oxoG and concomitant stable levels of 8-oxodG in leukocyte DNA were demonstrated to be associated with lower risks of mortality during the course of treatment.
The 8-oxodG levels in urine and in peripheral mononuclear cells (PMNC) from gastric cancer patients and healthy control subjects, before and after tumour resection, were analysed, and it was shown that the 8-oxodG/106 dG ratio was significantly higher in the PMNC of gastric cancer patients (4.16 ± 0.73) compared to healthy control individuals (8.43 ± 1.3), which was also supported by the urinary 8-oxodG levels in gastric cancer patients (22.29 ± 4.79 nmol/mmol of creatinine) and controls (2.49 ± 1.07 nmol/mmol of creatinine) [99]. After gastrectomy, the 8-oxodG levels slowly decreased to values close to those of healthy subjects, suggesting that high 8-oxodG urinary levels could be associated with increased DNA repair activity in tumour tissues.
Patients with monoclonal B lymphocytosis (MBL) and chronic lymphocytic leukaemia (CLL) showed a significant increase in 8-oxodG levels in the lymphocytes (8-oxo-dG/106 dG ratio of 39.5 in MBL and 48.4 in CCL vs. 6.1 in control) and urine (nmol 8-oxodG/mmol of creatinine of 15.1 in MBL and 14.6 in CCL vs. 11.0 in control) [123]. Higher levels of 8-oxodG in lymphocytes than in urine were correlated with decreases in the capacity of DNA repair systems, while no differences in the oxidative statuses of the MBL and CLL patients suggested that the oxidative injuries occurred during a pre-leukemic state of the disease.
Different studies have reported changes in the 8-oxodG levels in urine induced by various environmental factors: (i) ambient PM2.5 (0.26 nmol/kg in autumn, 0.22 nmol/kg in winter, 0.24 nmol/kg in spring and 0.21 nmol/kg in summer) [84,124], (ii) silica (1.99 ± 0.94 µmol/mol of creatinine in silicotics and 2.28 ± 0.92 µmol/mol of creatinine in exposed workers) [87], (iii) chromium (1149.5 ± 759.5 pmol/kg in exposed workers vs. 730.2 ± 377.6 pmol/kg in controls) [125], (iv) cobalt (1.52 ± 1.69 mol/mol creatinine in individuals exposed to Co and 1.63 ± 1.42 mol/mol creatinine in individuals exposed to hard metals vs. 1.46 ± 1.48 mol/mol creatinine in controls) [126], (v) cobalt and chromium from hip-replacement implants (1.15 µmol/mol of creatinine for implants: 3–4 years; 0.75 µmol/mol of creatinine for implants: 1–2 years) [127], (vi) PAHs (2.6 ± 0.8 g/g of creatinine at the start and 2.5 ± 0.8 g/g of creatinine at the end of the week in roofers exposed to asphalt fume, 2.5 ± 1.7 g/g of creatinine at the start and 3.0 ± 1.7 g/g of creatinine at the end of the week in roofers exposed to coal-tar vs. 3.3 ± 1.9 g/g of creatinine at the start and 3.5 ± 1.7 g/g of creatinine at the end of the week in controls [88]; in engine room personnel of ships, the results were 23.3 nmo/L for individuals with skin contaminated by oil and 18.7 nmol/L for individuals without contaminated skin vs. 18.0 nmol/L for control [128]), (vii) welding fumes (1.7 μg L−1/g of creatinine before and 2.1 μg L−1/g of creatinine after 3 h of exposure) [129], and (viii) carcinogenic metals including nickel and cadmium [130].
For oxidative stress measurement, saliva has also gained interest as an effective, non-invasive, and easy-to-collect sample source, but it is difficult to obtain accurate 8-oxodG measurements due to its low salivary levels (several pg/mL). The 8-oxoG content in saliva is several hundred-fold higher than that of 8-oxodG, and an average level of 3.80 ng mL−1 in normal healthy non-smoking subjects has been detected by HPLC–ECD [131,132].
3. Conclusions
The most frequent DNA lesions caused by oxidative stress are 8-oxoG and its nucleotide 8-oxodG (the G and dG oxidation products, respectively), which are regarded as biomarkers for oxidative DNA damage. The analytical quantification of 8-oxoG and 8-oxodG in urine, blood, tissue, and saliva is essential, as it is used to determine body oxidative stress and to evaluate the risk, diagnose in early stages, and predict the consequences of the treatment of various conditions, such as autoimmune, neurodegenerative, cardiovascular, and inflammatory diseases; diabetes; cancer; and other age-related diseases. The present review offers an exhaustive analysis of the most recent developments in HPLC–ECD methodologies for 8-oxoG and 8-oxodG analytical quantification in cellular DNA and in body fluids.
Funding
This research was funded by FEDER funds through the program COMPETE–Programa Operacional Factores de Competitividade and by national funds through FCT–Fundação para a Ciência e a Tecnologia under the project UID/EMS/00285/2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yudkina, A.V.; Shilkin, E.S.; Endutkin, A.V.; Makarova, A.V.; Zharkov, D.O. Reading and misreading 8-oxoguanine, a paradigmatic ambiguous nucleobase. Crystals 2019, 9, 269. [Google Scholar] [CrossRef] [Green Version]
- Loft, S.; Danielsen, P.; Løhr, M.; Jantzen, K.; Hemmingsen, J.G.; Roursgaard, M.; Karotki, D.G.; Møller, P. Urinary excretion of 8-oxo-7,8-dihydroguanine as biomarker of oxidative damage to DNA. Arch. Biochem. Biophys. 2012, 518, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Nakabeppu, Y. Cellular levels of 8-oxoguanine in either DNA or the nucleotide pool play pivotal roles in carcinogenesis and survival of cancer cells. Int. J. Mol. Sci. 2014, 15, 12543–12557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira-Brett, A.M.; Piedade, J.A.P.; Silva, L.A.; Diculescu, V.C. Voltammetric determination of all DNA nucleotides. Anal. Biochem. 2004, 332, 321–329. [Google Scholar] [CrossRef] [Green Version]
- Kino, K.; Hirao-Suzuki, M.; Morikawa, M.; Sakaga, A.; Miyazawa, H. Generation, repair and replication of guanine oxidation products. Genes Environ. 2017, 39, 21. [Google Scholar] [CrossRef]
- Goyal, R.N.; Jain, N.; Garg, D.K. Electrochemical and enzymic oxidation of guanosine and 8-hydroxyguanosine and the effects of oxidation products in mice. Bioelectrochemistry Bioenerg. 1997, 43, 105–114. [Google Scholar] [CrossRef]
- Brett, C.M.A.; Oliveira Brett, A.M.; Serrano, S.H.P. On the adsorption and electrochemical oxidation of DNA at glassy carbon electrodes. J. Electroanal. Chem. 1994, 366, 225–231. [Google Scholar] [CrossRef]
- Goyal, R.N.; Dryhurst, G. Redox chemistry of guanine and 8-oxyguanine and a comparison of the peroxidase-catalyzed and electrochemical oxidation of 8-oxyguanine. J. Electroanal. Chem. 1982, 135, 75–91. [Google Scholar] [CrossRef]
- David, S.S.; O’Shea, V.L.; Kundu, S. Base-excision repair of oxidative DNA damage. Nature 2007, 447, 941–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkateswarlu, D.; Leszczynski, J. Tautomeric equilibria in 8-oxopurines: Implications for mutagenicity. J. Comput. Aided. Mol. Des. 1998, 12, 373. [Google Scholar] [CrossRef]
- Drake, D.M.; Shapiro, A.M.; Wells, P.G. Measurement of the oxidative DNA lesion 8-oxoguanine (8-oxoG) by ELISA or by high-performance liquid chromatography (HPLC) with electrochemical detection. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2019; Volume 1965, Chapter 21; pp. 313–328. [Google Scholar]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-Hydroxy-2′ -deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health Part C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floyd, R.A.; Watson, J.J.; Wong, P.K.; Altmiller, D.H.; Rickard, R.C. Hydroxyl free radical adduct of deoxyguanosine: Sensitive detection and mechanisms of formation. Free Radic. Res. 1986, 1, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, H.E.; Loft, S.; Jensen, B.R.; Sørensen, M.; Hoberg, A.-M.; Weimann, A. HPLC-ECD, HPLC-MS/MS (Urinary Biomarkers). In Critical Reviews of Oxidative Stress and Aging; World Scientific Publishing Co.: Singapore, 2002; Volume 1, Chapter 13; pp. 233–256. [Google Scholar]
- Wang, C.C.; Chen, W.L.; Lin, C.M.; Lai, C.H.; Loh, C.H.; Chen, H.I.; Liou, S.H. The relationship between plasma and urinary 8-hydroxy-2-deoxyguanosine biomarkers measured by liquid chromatography tandem mass spectrometry. Environ. Sci. Pollut. Res. 2016, 23, 17496–17502. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.S.; Jenner, A.M.; Ong, C.N.; Huang, S.H.; Whiteman, M.; Halliwell, B. A high-throughput and sensitive methodology for the quantification of urinary 8-hydroxy-2′-deoxyguanosine: Measurement with gas chromatography-mass spectrometry after single solid-phase extraction. Biochem. J. 2004, 380, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Yamauchi, H.; Hasui, Y.; Kurashige, J.; Ochi, H.; Yoshida, K. Quantitative determination of urinary 8-hydroxydeoxyguanosine (8-OH-dG) by using ELISA. Res. Commun. Mol. Pathol. Pharmacol. 2000, 107, 39–44. [Google Scholar]
- Chiorcea-Paquim, A.M.; Oliveira-Brett, A.M. Nanostructured material–based electrochemical sensing of oxidative DNA damage biomarkers 8-oxoguanine and 8-oxodeoxyguanosine: A comprehensive review. Microchim. Acta 2021, 188, 1–13. [Google Scholar] [CrossRef]
- Tagesson, C.; Källberg, M.; Leanderson, P. Determination of urinary 8-hydroxydeoxyguanosine by coupled-column high-performance liquid chromatography with electrochemical detection: A noninvasive assay for in vivo oxidative DNA damage in humans. Toxicol. Mech. Methods 1991, 1, 242–251. [Google Scholar] [CrossRef]
- Kasai, H.; Crain, P.F.; Kuchino, Y.; Nishimura, S.; Ootsuyama, A.; Tanooka, H. Formation of 8-hydroxyguanine moiety in cellular dna by agents producing oxygen radicals and evidence for its repair. Carcinogenesis 1986, 7, 1849–1851. [Google Scholar] [CrossRef]
- Fraga, C.G.; Shigenaga, M.K.; Park, J.W.; Degan, P.; Ames, B.N. Oxidative damage to DNA during aging: 8-Hydroxy-2′-deoxyguanosine in rat organ DNA and urine. Proc. Natl. Acad. Sci. USA 1990, 87, 4533–4537. [Google Scholar] [CrossRef] [Green Version]
- Kasai, H.; Nishimura, S.; Kurokawa, Y.; Hayashi, Y. Oral administration of the renal carcinogen, potassium bromate, specifically produces 8-hydroxydeoxyguanosine in rat target organ DNA. Carcinogenesis 1987, 8, 1959–1961. [Google Scholar] [CrossRef]
- Richter, C.; Park, J.W.; Ames, B.N. Normal oxidative damage to mitochondrial and nuclear DNA is extensive. Proc. Natl. Acad. Sci. USA 1988, 85, 6465–6467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiala, E.S.; Conaway, C.C.; Mathis, J.E. Oxidative DNA and RNA Damage in the Livers of Sprague-Dawley Rats Treated with the Hepatocarcinogen 2-Nitropropane. Cancer Res. 1989, 49, 5518–5522. [Google Scholar] [PubMed]
- You, H.J.; Kim, G.T.; Kim, Y.H.; Chun, Y.S.; Park, J.W.; Chung, M.H.; Kim, M.S. Increased 8-hydroxyguanine formation and endonuclease activity for its repair in ischemic-reperfused heads of rats. J. Mol. Cell. Cardiol. 2000, 32, 1053–1059. [Google Scholar] [CrossRef]
- Kiyosawa, H.; Suko, M.; Okudaira, H.; Murata, K.; Miyamoto, E.; Chung, M.-H.; Kasai, H.; Nishimura, S. Cigarette Smoking Induces Formation of 8-Hydroxydeoxyguanosine, One of the Oxidative DNA Damages in Human Peripheral Leukocytes. Free Radic. Res. Commun. 1990, 11, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Thusu, K.; Cook, S.; Snyder, B.; Makowski, J.; Armstrong, D.; Nicotera, T. Oxidative damage to DNA in diabetes mellitus. Lancet 1996, 347, 444–445. [Google Scholar] [CrossRef]
- Asami, S.; Manabe, H.; Miyake, J.; Tsurudome, Y.; Hirano, T.; Yamaguchi, R.; Itoh, H.; Kasai, H. Cigarette smoking induces an increase in oxidative DNA damage, 8-hydroxydeoxyguanosine, in a central site of the human lung. Carcinogenesis 1997, 18, 1763–1766. [Google Scholar] [CrossRef] [Green Version]
- Mecocci, P.; MacGarvey, U.; Kaufman, A.E.; Koontz, D.; Shoffner, J.M.; Wallace, D.C.; Beal, M.F. Oxidative damage to mitochondrial DNA shows marked age-dependent increases in human brain. Ann. Neurol. 1993, 34, 609–616. [Google Scholar] [CrossRef]
- Musarrat, J.; Arezina-Wilson, J.; Wani, A.A. Prognostic and aetiological relevance of 8-hydroxyguanosine in human breast carcinogenesis. Eur. J. Cancer Part A 1996, 32, 1209–1214. [Google Scholar] [CrossRef]
- Kondo, S.; Toyokuni, S.; Tanaka, T.; Hiai, H.; Onodera, H.; Kasai, H.; Imamura, M. Overexpression of the hOGG1 gene and high 8-hydroxy-2′-deoxyguanosine (8-OHdG) lyase activity in human colorectal carcinoma: Regulation mechanism of the 8-OHdG level in DNA. Clin. Cancer Res. 2000, 6, 1394–1400. [Google Scholar]
- Loft, S.; Fischer-Nielsen, A.; Jeding, I.B.; Vistisen, K.; Poulsen, H.E. 8-hydroxydeoxyguanosine as a urinary biomarker of oxidative DNA damage. J. Toxicol. Environ. Health 1993, 40, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Shigenaga, M.K.; Gimeno, C.J.; Ames, B.N. Urinary 8-hydroxy-2′-deoxyguanosine as a biological marker of in vivo oxidative DNA damage. Proc. Natl. Acad. Sci. USA 1989, 86, 9697–9701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigenaga, M.K.; Ames, B.N. Assays for 8-hydroxy-2′-deoxyguanosine: A biomarker of in vivo oxidative DNA damage. In Free. Radic. Biol. Med. 1991, 10, 211–216. [Google Scholar] [CrossRef]
- Cundy, K.C.; Kohen, R.; Ames, B.N. Determination of 8-hydroxydeoxyguanosine in human urine: A possible assay for in vivo oxidative DNA damage. Basic Life Sci. 1988, 49, 479–482. [Google Scholar] [CrossRef] [PubMed]
- Kasai, H. Analysis of a form of oxidative DNA damage, 8-hydroxy-2′-deoxyguanosine, as a marker of cellular oxidative stress during carcinogenesis. Mutat. Res. Rev. Mutat. Res. 1997, 387, 147–163. [Google Scholar] [CrossRef]
- Koide, S.; Kinoshita, Y.; Ito, N.; Kimura, J.; Yokoyama, K.; Karube, I. Determination of human serum 8-hydroxy-2′-deoxyguanosine (8-OHdG) by HPLC-ECD combined with solid phase extraction (SPE). J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 2163–2167. [Google Scholar] [CrossRef]
- Loft, S.; Vistisen, K.; Ewertz, M.; Tjønneland, A.; Overvad, K.; Poulsen, H.E. Oxidative DNA damage estimated by 8-hydroxydeoxyguanosine excretion in humans: Influence of smoking, gender and body mass index. Carcinogenesis 1992, 13, 2241–2247. [Google Scholar] [CrossRef]
- Loft, S.; Svoboda, P.; Kasai, H.; Tjønneland, A.; Vogel, U.; Møller, P.; Overvad, K.; Raaschou-Nielsen, O. Prospective study of 8-oxo-7,8-dihydro-2′-deoxyguanosine excretion and the risk of lung cancer. Carcinogenesis 2006, 27, 1245–1250. [Google Scholar] [CrossRef] [Green Version]
- Bogdanov, M.B.; Beal, M.F.; McCabe, D.R.; Griffin, R.M.; Matson, W.R. A carbon column-based liquid chromatography electrochemical approach to routine 8-hydroxy-2′-deoxyguanosine measurements in urine and other biologic matrices: A one-year evaluation of methods. Free Radic. Biol. Med. 1999, 27, 647–666. [Google Scholar] [CrossRef]
- Lengger, C.; Schöch, G.; Topp, H. A high-performance liquid chromatographic method for the determination of 8-oxo-7,8-dihydro-2′-deoxyguanosine in urine from man and rat. Anal. Biochem. 2000, 287, 65–72. [Google Scholar] [CrossRef]
- Kasai, H.; Svoboda, P.; Yamasaki, S.; Kawai, K. Simultaneous determination of 8-hydroxydeoxyguanosine, a marker of oxidative stress, and creatinine, a standardization compound, in urine. Ind. Health 2005, 43, 333–336. [Google Scholar] [CrossRef] [Green Version]
- Svoboda, P.; Kasai, H. Simultaneous HPLC analysis of 8-hydroxydeoxyguanosine and 7-methylguanine in urine from humans and rodents. Anal. Biochem. 2004, 334, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Kasai, H. A new automated method to analyze urinary 8-hydroxydeoxyguanosine by a high-performance liquid chromatography-electrochemical detector system. J. Radiat. Res. 2003, 44, 185–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasai, H.; Iwamoto-Tanaka, N.; Miyamoto, T.; Kawanami, K.; Kawanami, S.; Kido, R.; Ikeda, M. Life style and urinary 8-hydroxydeoxyguanosine, a marker of oxidative DNA damage: Effects of exercise, working conditions, meat intake, body mass index, and smoking. Jpn. J. Cancer Res. 2001, 92, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Degan, P.; Shigenaga, M.K.; Park, E.M.; Alperin, P.E.; Ames, B.N. Immunoaffinity isolation of urinary 8-hydroxy-2′-deoxygunosine and 8-hydroxyguanine and quantitation of 8-hydroxy-2′deoxyguanosine in DNA by polyclonal antibodies. Carcinogenesis 1991, 12, 865–871. [Google Scholar] [CrossRef]
- Park, E.M.; Shigenaga, M.K.; Degan, P.; Korn, T.S.; Kitzler, J.W.; Wehr, C.M.; Kolachana, P.; Ames, B.N. Assay of excised oxidative DNA lesions: Isolation of 8-oxoguanine and its nucleoside derivatives from biological fluids with a monoclonal antibody column. Proc. Natl. Acad. Sci. USA 1992, 89, 3375–3379. [Google Scholar] [CrossRef] [Green Version]
- Shigenaga, M.K.; Aboujaoude, E.N.; Chen, Q.; Ames, B.N. Assays of oxidative DNA damage biomarkers 8-oxo-2′-deoxyguanosine and 8-oxoguanine in nuclear DNA and biological fluids by high-performance liquid chromatography with electrochemical detection. Methods Enzymol. 1994, 234, 16–33. [Google Scholar] [CrossRef]
- Helbock, H.J.; Beckman, K.B.; Shigenaga, M.K.; Walter, P.B.; Woodall, A.A.; Yeo, H.C.; Ames, B.N. DNA oxidation matters: The HPLC-electrochemical detection assay of 8-oxo-deoxyguanosine and 8-oxo-guanine. Proc. Natl. Acad. Sci. USA 1998, 95, 288–293. [Google Scholar] [CrossRef] [Green Version]
- Hofer, T.; Möller, L. Optimization of the workup procedure for the analysis of 8-oxo-7,8-dihydro-2′-deoxyguanosine with electrochemical detection. Chem. Res. Toxicol. 2002, 15, 426–432. [Google Scholar] [CrossRef]
- Hamilton, M.L. A reliable assessment of 8-oxo-2-deoxyguanosine levels in nuclear and mitochondrial DNA using the sodium iodide method to isolate DNA. Nucleic Acids Res. 2001, 29, 2117–2126. [Google Scholar] [CrossRef]
- Bolin, C.; Stedeford, T.; Cardozo-Pelaez, F. Single extraction protocol for the analysis of 8-hydroxy-2′- deoxyguanosine (oxo8dG) and the associated activity of 8-oxoguanine DNA glycosylase. J. Neurosci. Methods 2004, 136, 69–76. [Google Scholar] [CrossRef]
- Oliveira Brett, A.M.; Piedade, J.A.P.; Serrano, S.H.P. Electrochemical oxidation of 8-oxoguanine. Electroanalysis 2000, 12, 969–973. [Google Scholar] [CrossRef]
- Rebelo, I.A.; Piedade, J.A.P.; Oliveira-Brett, A.M. Development of an HPLC method with electrochemical detection of femtomoles of 8-oxo-7,8-dihydroguanine and 8-oxo-7,8-dihydro-2′-deoxyguanosine in the presence of uric acid. Talanta 2004, 63, 323–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diculescu, V.C.; Chiorcea Paquim, A.-M.; Oliveira Brett, A.M. Electrochemical DNA Sensors for Detection of DNA Damage. Sensors 2005, 5, 377–393. [Google Scholar] [CrossRef]
- Chiorcea-Paquim, A.M.; Oliveira-Brett, A.M. Dna electrochemical biosensors for in situ probing of pharmaceutical drug oxidative dna damage. Sensors 2021, 21, 1125. [Google Scholar] [CrossRef]
- 8-Hydroxy-2′-deoxyguanosine as a marker of oxidative DNA damage related to occupational and environmental exposures [BAT Value Documentation, 2010]. In The MAK-Collection for Occupational Health and Safety; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2016; pp. 1–24.
- Collins, A.; Cadet, J.; Epe, B.; Gedik, C. Problems in the measurement of 8-oxoguanine in human DNA—Report of a Workshop, DNA Oxidation, Aberdeen, UK, 19–21 January 1997. Carcinogenesis 1997, 18, 1833–1836. [Google Scholar] [CrossRef]
- Riis, B.; Collins, A.; Gedik, C.; Wood, S.; White, A.A.; Dubois, J.; Duez, P.; Rees, J.F.; Legall, R.; Degand, L.; et al. Comparison of results from different laboratories in measuring 8-oxo-2′-deoxyguanosine in synthetic oligonucleotides. Free Radic. Res. 2002, 36, 649–659. [Google Scholar] [CrossRef]
- Collins, A.R.; Cadet, J.; Moller, L.; Poulsen, H.E.; Viña, J. Are we sure we know how to measure 8-oxo-7,8-dihydroguanine in DNA from human cells? Arch. Biochem. Biophys. 2004, 423, 57–65. [Google Scholar] [CrossRef]
- Harris, G.; Bashir, S.; Winyard, P.G. 7, 8-dihydro-8-oxo-2′-deoxyguanosine present in DNA is not simply an artefact of isolation. Carcinogenesis 1994, 15, 411–413. [Google Scholar] [CrossRef]
- Collins, A.R.; Gedik, C.; Wood, S.; White, A.A.; Dubois, J.; Duez, P.; Rees, J.F.; Legall, R.; Degand, L.; Loft, S.; et al. Inter-laboratory validation of procedures for measuring 8-oxo-7,8-dihydroguanine/8-oxo-7,8-dihydro-2′-deoxyguanosine in DNA. Free Radic. Res. 2002, 36, 239–245. [Google Scholar] [CrossRef]
- Cadet, J.; D’Ham, C.; Douki, T.; Pouget, J.P.; Ravanat, J.L.; Sauvaigo, S. Facts and artifacts in the measurement of oxidative base damage to DNA. Free Radic. Res. 1998, 29, 541–550. [Google Scholar] [CrossRef]
- Collins, A.; Gedik, C.; Vaughan, N.; Wood, S.; White, A.; Dubois, J.; Rees, J.F.; Loft, S.; Møller, P.; Poulsen, H.; et al. Measurement of DNA oxidation in human cells by chromatographic and enzymic methods. Free Radic. Biol. Med. 2003, 34, 1089–1099. [Google Scholar] [CrossRef]
- Collins, A.; Gedik, C.; Vaughan, N.; Wood, S.; White, A.; Dubois, J.; Duez, P.; Dehon, G.; Rees, J.F.; Loft, S.; et al. Comparative analysis of baseline 8-oxo-7,8-dihydroguanine in mammalian cell DNA, by different methods in different laboratories: An approach to consensus. Carcinogenesis 2002, 23, 2129–2133. [Google Scholar]
- Gedik, C.M.; Collins, A. Establishing the background level of base oxidation in human lymphocyte DNA: Results of an interlaboratory validation study. FASEB J. 2005, 19, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Barnett, Y.A.; Barnett, C.R.; Wood, S.G.; Gedik, C.M.; Vaughan, N.J.; Collins, A.R. Measurement of 8-oxo-deoxyguanosine in lymphocytes, cultured cells, and tissue samples by HPLC with electrochemical detection. In Aging Methods and Protocols; Humana Press Inc.: Totowa, NJ, USA, 2000; Volume 38, pp. 171–178. [Google Scholar]
- Chepelev, N.L.; Kennedy, D.A.; Gagné, R.; White, T.; Long, A.S.; Yauk, C.L.; White, P.A. HPLC measurement of the DNA oxidation biomarker, 8-oxo-7,8-dihydro-2′-deoxyguanosine, in cultured cells and animal tissues. J. Vis. Exp. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Hofer, T.; Möller, L. Reduction of oxidation during the preparation of DNA and analysis of 8- hydroxy-2′-deoxyguanosine. Chem. Res. Toxicol. 1998, 11, 882–887. [Google Scholar] [CrossRef]
- Gedik, C.M.; Wood, S.G.; Collins, A.R. Measuring oxidative damage to DNA; HPLC and the comet assay compared. Free Radic. Res. 1998, 29, 609–615. [Google Scholar] [CrossRef]
- Ravanat, J.-L. Cellular background level of 8-oxo-7,8-dihydro-2′-deoxyguanosine: An isotope based method to evaluate artefactual oxidation of DNA during its extraction and subsequent work-up. Carcinogenesis 2002, 23, 1911–1918. [Google Scholar] [CrossRef] [Green Version]
- Lodovici, M.; Casalini, C.; Cariaggi, R.; Michelucci, L.; Dolara, P. Levels of 8-hydroxydeoxyguanosine as a marker of DNA damage in human leukocytes. Free Radic. Biol. Med. 2000, 28, 13–17. [Google Scholar] [CrossRef]
- Kim, Y.D.; Lee, C.H.; Nan, H.M.; Kang, J.W.; Kim, H. Effects of genetic polymorphisms in metabolic enzymes on the relationships between 8-hydroxydeoxyguanosine levels in human leukocytes and urinary 1-hydroxypyrene and 2-naphthol concentrations. J. Occup. Health 2003, 45, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Tarng, D.C.; Liu, T.Y.; Huang, T.P. Protective effect of vitamin C on 8-hydroxy-2′-deoxyguanosine level in peripheral blood lymphocytes of chronic hemodialysis patients. Kidney Int. 2004, 66, 820–831. [Google Scholar] [CrossRef] [Green Version]
- Herbert, K.E.; Fletcher, S.; Chauhan, D.; Ladapo, A.; Nirwan, J.; Munson, S.; Mistry, P. Dietary supplementation with different vitamin C doses: No effect on oxidative DNA damage in healthy people. Eur. J. Nutr. 2006, 45, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Yen, M.Y.; Kao, S.H.; Wang, A.G.; Wei, Y.H. Increased 8-hydroxy-2′-deoxyguanosine in leukocyte DNA in Leber’s hereditary optic neuropathy. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1688–1691. [Google Scholar] [CrossRef] [PubMed]
- Karahan, M.; Yildirim, M.; Kucuk, H.F.; Turunc, V.; Demir, H.; Salturk, C.; Yavuz, A.; Demir, T.; Ari, E. Oxidative DNA Damage Is Increased in Living Kidney Donors. Transplant. Proc. 2019, 51, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Katerji, M.; Filippova, M.; Duerksen-Hughes, P. Approaches and methods to measure oxidative stress in clinical samples: Research applications in the cancer field. Oxid. Med. Cell. Longev. 2019, 1279250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, A.; Ikeda, T.; Enomoto, K.; Hosoda, K.; Nakashima, H.; Omae, K.; Watanabe, M.; Hibi, T.; Kitajima, M. Increased formation of oxidative DNA damage, 8-hydroxy-2′- deoxyguanosine, in human breast cancer tissue and its relationship to GSTP1 and COMT genotypes. Cancer Lett. 2000, 151, 87–95. [Google Scholar] [CrossRef]
- Li, D.; Zhang, W.; Zhu, J.; Chang, P.; Sahin, A.; Singletary, E.; Bondy, M.; Hazra, T.; Mitra, S.; Lau, S.S.; et al. Oxidative DNA damage and 8-hydroxy-2-deoxyguanosine DNA glycosylase/apurinic lyase in human breast cancer. Mol. Carcinog. 2001, 31, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Kondo, S.; Toyokuni, S.; Iwasa, Y.; Tanaka, T.; Onodera, H.; Hiai, H.; Imamura, M. Persistent oxidative stress in human colorectal carcinoma, but not in adenoma. Free Radic. Biol. Med. 1999, 27, 401–410. [Google Scholar] [CrossRef]
- Płachetka, A.; Adamek, B.; Strzelczyk, J.K.; Krakowczyk, Ł.; Migula, P.; Nowak, P.; Wiczkowski, A. 8-hydroxy-2′-deoxyguanosine in colorectal adenocarcinoma—Is it a result of oxidative stress? Med. Sci. Monit. 2013, 19, 690–695. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.J.; Choi, E.Y.; Choi, J.Y.; Park, J.G.; You, H.J.; Chung, M.H. Genetic changes of hOGG1 and the activity of oh8Gua glycosylase in colon cancer. Eur. J. Cancer 2001, 37, 340–346. [Google Scholar] [CrossRef]
- Sørensen, M.; Autrup, H.; Hertel, O.; Wallin, H.; Knudsen, L.E.; Loft, S. Personal exposure to PM2.5 and biomarkers of DNA damage. Cancer Epidemiol. Biomark. Prev. 2003, 12, 191–196. [Google Scholar]
- Marczynski, B.; Kraus, T.; Rozynek, P.; Raithel, H.J.; Baur, X. Association between 8-hydroxy-2′-deoxyguanosine levels in DNA of workers highly exposed to asbestos and their clinical data, occupational and non-occupational confounding factors, and cancer. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2000, 468, 203–212. [Google Scholar] [CrossRef]
- Marczynski, B.; Rozynek, P.; Kraus, T.; Schlösser, S.; Raithel, H.J.; Baur, X. Levels of 8-hydroxy-2′-deoxyguanosine in DNA of white blood cells from workers highly exposed to asbestos in Germany. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2000, 468, 195–202. [Google Scholar] [CrossRef]
- Pilger, A.; Germadnik, D.; Schaffer, A.; Theiler, A.; Pils, P.; Sluka, F.; Winker, N.; Rüdiger, H.W. 8-Hydroxydeoxyguanosine in leukocyte DNA and urine of quartz-exposed workers and patients with silicosis. Int. Arch. Occup. Environ. Health 2000, 73, 305–310. [Google Scholar] [CrossRef]
- Toraason, M.; Hayden, C.; Marlow, D.; Rinehart, R.; Mathias, P.; Werren, D.; DeBord, D.G.; Reid, T.M. DNA strand breaks, oxidative damage, and 1-OH pyrene in roofers with coal-tar pitch dust and/or asphalt fume exposure. Int. Arch. Occup. Environ. Health 2001, 74, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ichiba, M.; Hanaoka, T.; Pan, G.; Yamano, Y.; Hara, K.; Takahashi, K.; Tomokuni, K. Leukocyte 8-hydroxydeoxyguanosine and aromatic DNA adduct in coke-oven workers with polycyclic aromatic hydrocarbon exposure. Int. Arch. Occup. Environ. Health 2003, 76, 499–504. [Google Scholar] [CrossRef] [PubMed]
- McDorman, K.S.; Pachkowski, B.F.; Nakamura, J.; Wolf, D.C.; Swenberg, J.A. Oxidative DNA damage from potassium bromate exposure in Long-Evans rats is not enhanced by a mixture of drinking water disinfection by-products. Chem. Biol. Interact. 2005, 152, 107–117. [Google Scholar] [CrossRef]
- Gupta, N.; Curtis, R.M.; Mulder, J.E.; Massey, T.E. Acute in vivo treatment with 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone does not alter base excision repair activities in murine lung and liver. DNA Repair 2013, 12, 1031–1036. [Google Scholar] [CrossRef]
- Guindon-Kezis, K.A.; Mulder, J.E.; Massey, T.E. In vivo treatment with aflatoxin B1 increases DNA oxidation, base excision repair activity and 8-oxoguanine DNA glycosylase 1 levels in mouse lung. Toxicology 2014, 321, 21–26. [Google Scholar] [CrossRef]
- Orimo, H.; Mei, N.; Boiteux, S.; Tokura, Y.; Kasai, H. Analysis of 8-hydroxyguanine (8-OH-Gua) released from DNA by the formamidopyrimidine DNA glycosylase (Fpg) protein: A reliable method to estimate cellular oxidative stress. J. Radiat. Res. 2004, 45, 455–460. [Google Scholar] [CrossRef] [Green Version]
- Graille, M.; Wild, P.; Sauvain, J.J.; Hemmendinger, M.; Canu, I.G.; Hopf, N.B. Urinary 8-OHdG as a biomarker for oxidative stress: A systematic literature review and meta-analysis. Int. J. Mol. Sci. 2020, 21, 3743. [Google Scholar] [CrossRef]
- ESCULA [European Standards Committee on Urinary (DNA) Lesion Analysis]; Evans, M.D.; Olinski, R.; Loft, S.; Cooke, M.S. Toward consensus in the analysis of urinary 8-oxo-7,8-dihydro-2′-deoxyguanosine as a noninvasive biomarker of oxidative stress. FASEB J. 2010, 24, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Cooke, M.S.; Barregard, L.; Mistry, V.; Potdar, N.; Rozalski, R.; Gackowski, D.; Siomek, A.; Foksinski, M.; Svoboda, P.; Kasai, H.; et al. Interlaboratory comparison of methodologies for the measurement of urinary 8-oxo-7,8-dihydro-2′-deoxyguanosine. Biomarkers 2009, 14, 103–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooke, M.S.; Olinski, R.; Loft, S. Measurement and meaning of oxidatively modified DNA lesions in urine. Cancer Epidemiol. Biomark. Prev. 2008, 17, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.S.; Song, M.F.; Kasai, H.; Kawai, K. 8-Hydroxyguanine in urine and serum as an oxidative stress marker: Effects of diabetes and aging. J. UOEH 2013, 35, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Borrego, S.; Vazquez, A.; Dasí, F.; Cerdá, C.; Iradi, A.; Tormos, C.; Sánchez, J.M.; Bagán, L.; Boix, J.; Zaragoza, C.; et al. Oxidative stress and DNA damage in human gastric carcinoma: 8-Oxo-7′8-dihydro-2′-deoxyguanosine (8-oxo-dG) as a possible tumor marker. Int. J. Mol. Sci. 2013, 14, 3467–3486. [Google Scholar] [CrossRef]
- Inaba, Y.; Koide, S.; Yokoyama, K.; Karube, I. Development of urinary 8-hydroxy-2′-deoxyguanosine (8-OHdG) measurement method combined with SPE. J. Chromatogr. Sci. 2011, 49, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Kato, D.; Komoriya, M.; Nakamoto, K.; Kurita, R.; Hirono, S.; Niwa, O. Electrochemical determination of oxidative damaged DNA with high sensitivity and stability using a nanocarbon film. Anal. Sci. 2011, 27, 703–707. [Google Scholar] [CrossRef] [Green Version]
- Rebelo, I.; Piedade, J.A.P.; Oliveira Brett, A.M. Electrochemical determination of 8-oxoguanine in the presence of uric acid. Bioelectrochemistry 2004, 63, 267–270. [Google Scholar] [CrossRef] [Green Version]
- Saravanabhavan, G.; Blais, E.; Vincent, R.; Kumarathasan, P. A high performance liquid chromatography-electrochemical array method for the measurement of oxidative/nitrative changes in human urine. J. Chromatogr. A 2010, 1217, 3269–3274. [Google Scholar] [CrossRef]
- El-Bayoumy, K.; Richie, J.P.; Boyiri, T.; Komninou, D.; Prokopczyk, B.; Trushin, N.; Kleinman, W.; Cox, J.; Pittman, B.; Colosimo, S. Influence of selenium-enriched yeast supplementation on biomarkers of oxidative damage and hormone status in healthy adult males: A clinical pilot study. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1459–1465. [Google Scholar]
- Miyata, M.; Kasai, H.; Kawai, K.; Yamada, N.; Tokudome, M.; Ichikawa, H.; Goto, C.; Tokudome, Y.; Kuriki, K.; Hoshino, H.; et al. Changes of urinary 8-hydroxydeoxyguanosine levels during a two-day ultramarathon race period in Japanese non-professional runners. Int. J. Sports Med. 2008, 29, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Jia, X.; Chen, C.Y.O.; Blumberg, J.B.; Song, Y.; Zhang, W.; Zhang, X.; Ma, G.; Chen, J. Almond consumption reduces oxidative DNA damage and lipid peroxidation in male smokers. J. Nutr. 2007, 137, 2717–2722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilger, A.; Germadnik, D.; Riedel, K.; Meger-Kossien, I.; Scherer, G.; Rüdiger, H. Longitudinal study of urinary 8-hydroxy-2′-deoxyguanosine excretion in healthy adults. Free Radic. Res. 2001, 35, 273–280. [Google Scholar] [CrossRef]
- Kuo, H.W.; Chou, S.Y.; Hu, T.W.; Wu, F.Y.; Chen, D.J. Urinary 8-hydroxy-2′-deoxyguanosine (8-OHdG) and genetic polymorphisms in breast cancer patients. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2007, 631, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yi, J.; Liu, F. The expression and significance of 8-hydroxydeoxyguanosine in breast cancer patients’ blood, urine and cancer tissue. Trends Immunother. 2021, 5, 36–41. [Google Scholar] [CrossRef]
- Shin, C.S.; Moon, B.S.; Park, K.S.; Kim, S.Y.; Park, S.J.; Chung, M.H.; Lee, H.K. Serum 8-hydroxy-guanine levels are increased in diabetic patients. Diabetes Care 2001, 24, 733–737. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Li, Y.S.; Kawasaki, Y.; Ootsuyama, Y.; Kawai, K. Health examination results and work environment factors affecting urinary 8-hydroxy-2′-deoxyguanosine levels. J. Occup. Health 2021, 63, e12210. [Google Scholar] [CrossRef]
- Irie, M.; Tamae, K.; Iwamoto-Tanaka, N.; Kasai, H. Occupational and lifestyle factors and urinary 8-hydroxydeoxyguanosine. Cancer Sci. 2005, 96, 600–606. [Google Scholar] [CrossRef]
- Hara, M.; Nishida, Y.; Shimanoe, C.; Otsuka, Y.; Nanri, H.; Yasukata, J.; Miyoshi, N.; Yamada, Y.; Horita, M.; Kawai, K.; et al. Intensity-specific effect of physical activity on urinary levels of 8-hydroxydeoxyguanosine in middle-aged Japanese. Cancer Sci. 2016, 107, 1653–1659. [Google Scholar] [CrossRef]
- Kikuchi, H.; Nanri, A.; Hori, A.; Sato, M.; Kawai, K.; Kasai, H.; Mizoue, T. Lower serum levels of total cholesterol are associated with higher urinary levels of 8-hydroxydeoxyguanosine. Nutr. Metab. 2013, 10, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.S.; Kawasaki, Y.; Watanabe, S.; Ootsuyama, Y.; Kasai, H.; Kawai, K. Diurnal and day-to-day variation of urinary oxidative stress marker 8-hydroxy-2′- deoxyguanosine. J. Clin. Biochem. Nutr. 2021, 68, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Shimanoe, C.; Hara, M.; Nishida, Y.; Nanri, H.; Horita, M.; Yamada, Y.; Li, Y.S.; Kasai, H.; Kawai, K.; Higaki, Y.; et al. Perceived Stress, Depressive Symptoms, and Oxidative DNA Damage. Psychosom. Med. 2018, 80, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, Y.; Li, Y.S.; Ootsuyama, Y.; Nagata, K.; Yamato, H.; Kawai, K. Effects of smoking cessation on biological monitoring markers in urine. Genes Environ. 2020, 42, 26. [Google Scholar] [CrossRef] [PubMed]
- Foksinski, M.; Rozalski, R.; Guz, J.; Ruszkowska, B.; Sztukowska, P.; Piwowarski, M.; Klungland, A.; Olinski, R. Urinary excretion of DNA repair products correlates with metabolic rates as well as with maximum life spans of different mammalian species. Free Radic. Biol. Med. 2004, 37, 1449–1454. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, P.; Maekawa, M.; Kawai, K.; Tominaga, T.; Savela, K.; Kasai, H. Urinary 8-hydroxyguanine may be a better marker of oxidative stress than 8-hydroxydeoxyguanosine in relation to the life spans of various species. Antioxidants Redox Signal. 2006, 8, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Loft, S.; Svoboda, P.; Kawai, K.; Kasai, H.; Sørensen, M.; Tjønneland, A.; Vogel, U.; Møller, P.; Overvad, K.; Raaschou-Nielsen, O. Association between 8-oxo-7,8-dihydroguanine excretion and risk of lung cancer in a prospective study. Free Radic. Biol. Med. 2012, 52, 167–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haghdoost, S.; Svoboda, P.; Näslund, I.; Harms-Ringdahl, M.; Tilikides, A.; Skog, S. Can 8-oxo-dG be used as a predictor for individual radiosensitivity? Int. J. Radiat. Oncol. Biol. Phys. 2001, 50, 405–410. [Google Scholar] [CrossRef]
- Roszkowski, K.; Olinski, R. Urinary 8-oxoguanine as a predictor of survival in patients undergoing radiotherapy. Cancer Epidemiol. Biomark. Prev. 2012, 21, 629–634. [Google Scholar] [CrossRef] [Green Version]
- Collado, R.; Oliver, I.; Tormos, C.; Egea, M.; Miguel, A.; Cerdá, C.; Ivars, D.; Borrego, S.; Carbonell, F.; Sáez, G.T. Early ROS-mediated DNA damage and oxidative stress biomarkers in Monoclonal B Lymphocytosis. Cancer Lett. 2012, 317, 144–149. [Google Scholar] [CrossRef]
- Sørensen, M.; Schins, R.P.F.; Hertel, O.; Loft, S. Transition metals in personal samples of PM2.5 and oxidative stress in human volunteers. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1340–1343. [Google Scholar] [CrossRef] [Green Version]
- Kuo, H.W.; Chang, S.F.; Wu, K.Y.; Wu, F.Y. Chromium (VI) induced oxidative damage to DNA: Increase of urinary 8-hydroxydeoxyguanosine concentrations (8-OHdG) among electroplating workers. Occup. Environ. Med. 2003, 60, 590–594. [Google Scholar] [CrossRef] [PubMed]
- De Boeck, M.; Lardau, S.; Buchet, J.; Kirsch-Volders, M.; Lison, D. Absence of significant genotoxicity in lymphocytes and urine from workers exposed to moderate levels of cobalt-containing dust: A cross-sectional study. Environ. Mol. Mutagen. 2000, 36, 151–160. [Google Scholar] [CrossRef]
- Pilger, A.; Schaffer, A.; Rüdiger, H.W.; Osterode, W. Urinary 8-hydroxydeoxyguanosine and sister chromatid exchanges in patients with total hip replacements. J. Toxicol. Environ. Health Part A 2002, 65, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.; Nordlinder, R.; Moen, B.E.; Øvrebø, S.; Bleie, K.; Skorve, A.H.; Hollund, B.E.; Tagesson, C. Increased urinary excretion of 8-hydroxydeoxyquanosine in engine room personnel exposed to polycyclic aromatic hydrocarbons. Occup. Environ. Med. 2004, 61, 692–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graczyk, H.; Lewinski, N.; Zhao, J.; Sauvain, J.J.; Suarez, G.; Wild, P.; Danuser, B.; Riediker, M. Increase in oxidative stress levels following welding fume inhalation: A controlled human exposure study. Part. Fibre Toxicol. 2016, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, N.; Nakashima, H.; Hosoda, K.; Eitaki, Y.; Shimada, N.; Omae, K. Urinary excretion of an oxidative stress marker, 8-hydroxyguanine (8-OH-GUA), among nickel-cadmium battery workers. J. Occup. Health 2008, 50, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Kawai, K.; Kasai, H.; Li, Y.S.; Kawasaki, Y.; Watanabe, S.; Ohta, M.; Honda, T.; Yamato, H. Measurement of 8-hydroxyguanine as an oxidative stress biomarker in saliva by HPLC-ECD. Genes Environ. 2018, 40, 5. [Google Scholar] [CrossRef]
- Watanabe, S.; Kawasaki, Y.; Kawai, K. Salivary 8-hydroxyguanine as a lifestyle-related oxidative stress biomarker in workers. J. Clin. Biochem. Nutr. 2020, 66, 57–61. [Google Scholar] [CrossRef] [Green Version]
Scheme 1.
Schematic representation of oxidative stress agents and oxidative DNA damage.

Scheme 2.
Schematic representation of mutagenesis caused by 8-oxoguanine (8-oxoG), the major base lesion in DNA caused by ROS that leads to G:C–T:A and G:C–C:G or transversions. Black and green lines represent the template DNA strand and the nascent DNA strand, respectively, during replication.
Scheme 2.
Schematic representation of mutagenesis caused by 8-oxoguanine (8-oxoG), the major base lesion in DNA caused by ROS that leads to G:C–T:A and G:C–C:G or transversions. Black and green lines represent the template DNA strand and the nascent DNA strand, respectively, during replication.

Scheme 3.
(a) Chemical structures of 8-oxoguanine (8-oxoG), 8-oxo-2′-deoxyguanosine (8-oxodG), and uric acid (UA); (b) 8-oxoG tautomers; and (c) the oxidation of guanine (G) to 8-oxoG.
Scheme 3.
(a) Chemical structures of 8-oxoguanine (8-oxoG), 8-oxo-2′-deoxyguanosine (8-oxodG), and uric acid (UA); (b) 8-oxoG tautomers; and (c) the oxidation of guanine (G) to 8-oxoG.

Scheme 4.
Structures of normal G(anti):C(anti) and T(anti):A(anti) Watson–Crick, and 8-oxoG(anti):C(anti) Watson–Crick and 8-oxoG(syn):A(anti) Hoogsteen base pairs.
Scheme 4.
Structures of normal G(anti):C(anti) and T(anti):A(anti) Watson–Crick, and 8-oxoG(anti):C(anti) Watson–Crick and 8-oxoG(syn):A(anti) Hoogsteen base pairs.

Figure 1.
Differential pulse voltammograms at a glassy carbon electrode in a 5 µM solution of (black line) 8-oxoG, (black dotted line) UA, and (red line) a 5 µM equimolar mixture of 8-oxoG and UA in (a) pH = 9.4 and (b) pH = 6.2. Reproduced from [102] with permission. Copyright 2004 Elsevier.
Figure 1.
Differential pulse voltammograms at a glassy carbon electrode in a 5 µM solution of (black line) 8-oxoG, (black dotted line) UA, and (red line) a 5 µM equimolar mixture of 8-oxoG and UA in (a) pH = 9.4 and (b) pH = 6.2. Reproduced from [102] with permission. Copyright 2004 Elsevier.

Figure 2.
(a) HPLC chromatograms obtained from (black dotted line) a 10 µL injection of a 100 nM standard mixture of UA, 8-oxoG, and 8-oxodG; (black line) a 10 µL injection of a urine sample from a child with metabolic disorders after being processed by solid phase extraction (SPE). (b,c) Same urine sample (black line) before and (black dotted line) after being spiked with 250 nM standard solutions of (b) 8-oxoG and (c) 8-oxodG. Reproduced from [54] with permission. Copyright 2004 Elsevier.
Figure 2.
(a) HPLC chromatograms obtained from (black dotted line) a 10 µL injection of a 100 nM standard mixture of UA, 8-oxoG, and 8-oxodG; (black line) a 10 µL injection of a urine sample from a child with metabolic disorders after being processed by solid phase extraction (SPE). (b,c) Same urine sample (black line) before and (black dotted line) after being spiked with 250 nM standard solutions of (b) 8-oxoG and (c) 8-oxodG. Reproduced from [54] with permission. Copyright 2004 Elsevier.

Figure 3.
Identification of target analytes in spiked urine samples at different electrochemical channels: (A) m-tyrosine and nitrotyrosine (0.65 V); (B) 8-oxodG (0.25 V); (C) o-tyrosine and cl-tyrosine (0.50 V). Reproduced from [103] with permission. Copyright 2010 Elsevier.
Figure 3.
Identification of target analytes in spiked urine samples at different electrochemical channels: (A) m-tyrosine and nitrotyrosine (0.65 V); (B) 8-oxodG (0.25 V); (C) o-tyrosine and cl-tyrosine (0.50 V). Reproduced from [103] with permission. Copyright 2010 Elsevier.

Figure 4.
HPLC chromatograms of serum samples for healthy people. HPLC conditions were a mobile phase consisting of a 35 mmol L−1 phosphate buffer (pH 7.0) containing 5% acetonitrile and 30 mol L−1 EDTA, a column oven temperature of 27 °C, an injection volume of 100 L, a flow rate of 1.0 mL/min, and an HPLC Develosil C30 prepacked column (5 m particle size). Reproduced from [37] with permission. Copyright 2010 Elsevier.
Figure 4.
HPLC chromatograms of serum samples for healthy people. HPLC conditions were a mobile phase consisting of a 35 mmol L−1 phosphate buffer (pH 7.0) containing 5% acetonitrile and 30 mol L−1 EDTA, a column oven temperature of 27 °C, an injection volume of 100 L, a flow rate of 1.0 mL/min, and an HPLC Develosil C30 prepacked column (5 m particle size). Reproduced from [37] with permission. Copyright 2010 Elsevier.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chiorcea-Paquim, A.-M. 8-oxoguanine and 8-oxodeoxyguanosine Biomarkers of Oxidative DNA Damage: A Review on HPLC–ECD Determination. Molecules 2022, 27, 1620. https://doi.org/10.3390/molecules27051620
AMA Style
Chiorcea-Paquim A-M. 8-oxoguanine and 8-oxodeoxyguanosine Biomarkers of Oxidative DNA Damage: A Review on HPLC–ECD Determination. Molecules. 2022; 27(5):1620. https://doi.org/10.3390/molecules27051620
Chicago/Turabian StyleChiorcea-Paquim, Ana-Maria. 2022. "8-oxoguanine and 8-oxodeoxyguanosine Biomarkers of Oxidative DNA Damage: A Review on HPLC–ECD Determination" Molecules 27, no. 5: 1620. https://doi.org/10.3390/molecules27051620