A Study on the Biodiversity of Pigmented Andean Potatoes: Nutritional Profile and Phenolic Composition

 ,
,  ,
,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Morphological Aspects and Proximate Composition
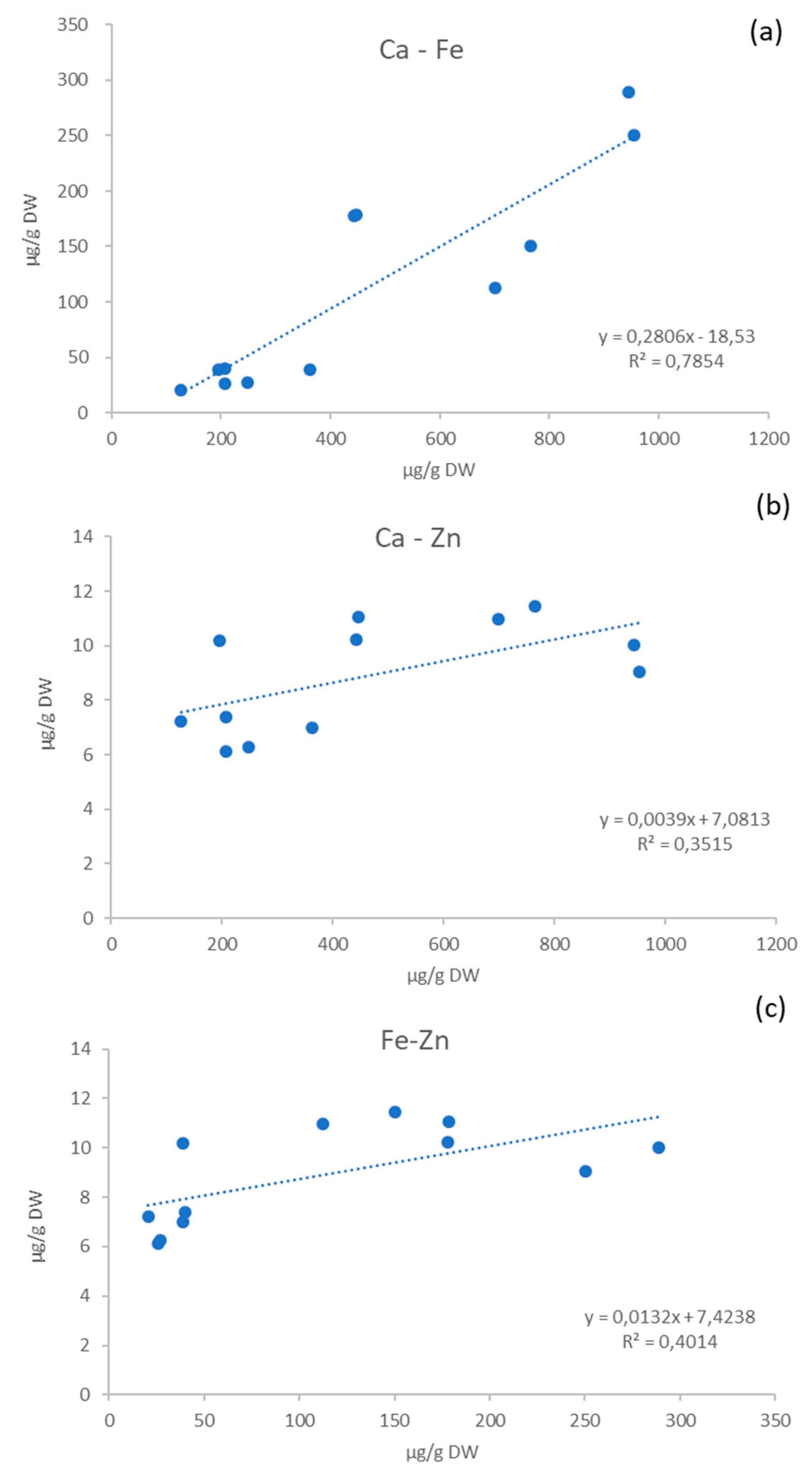
2.2. Mineral Content
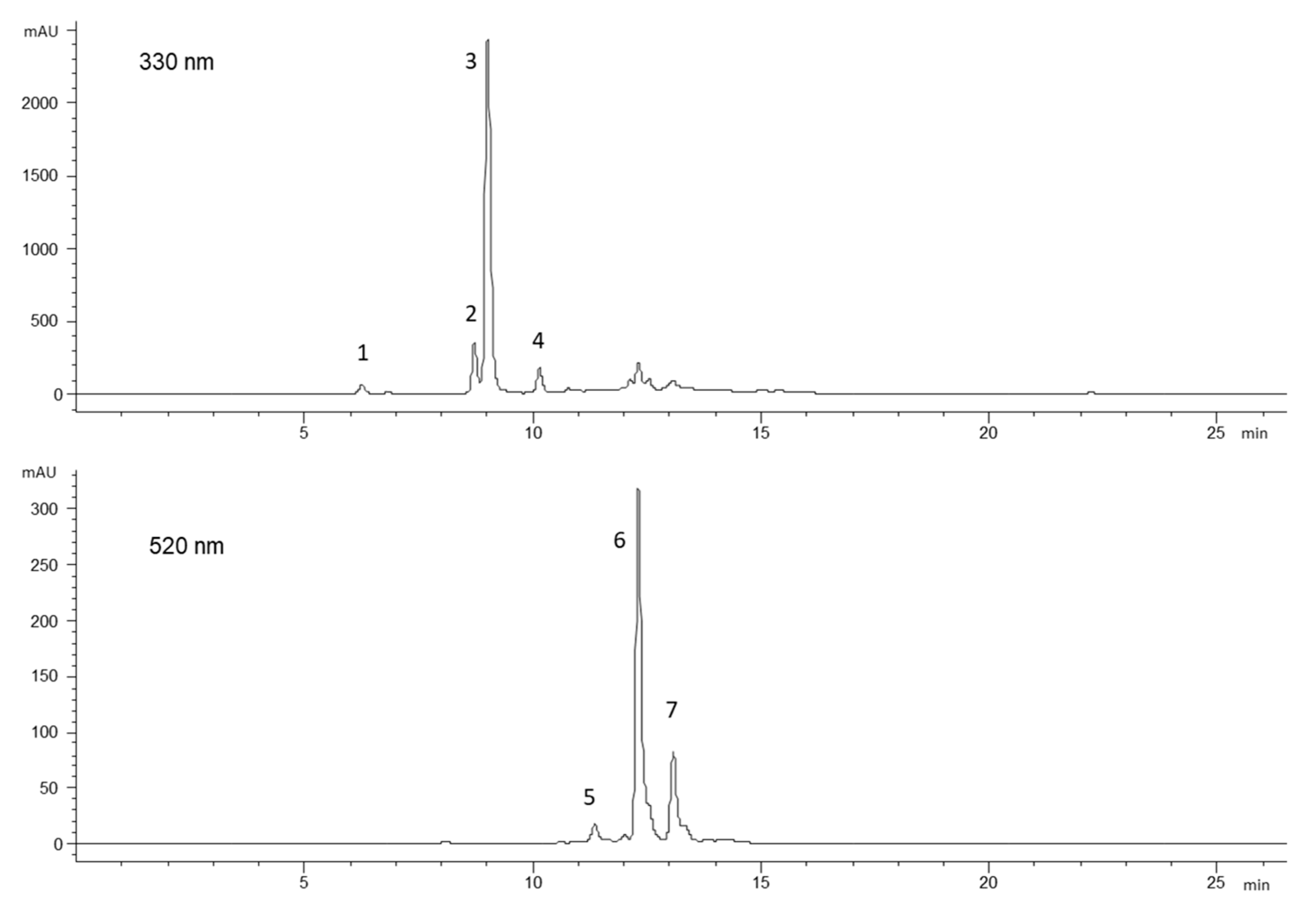
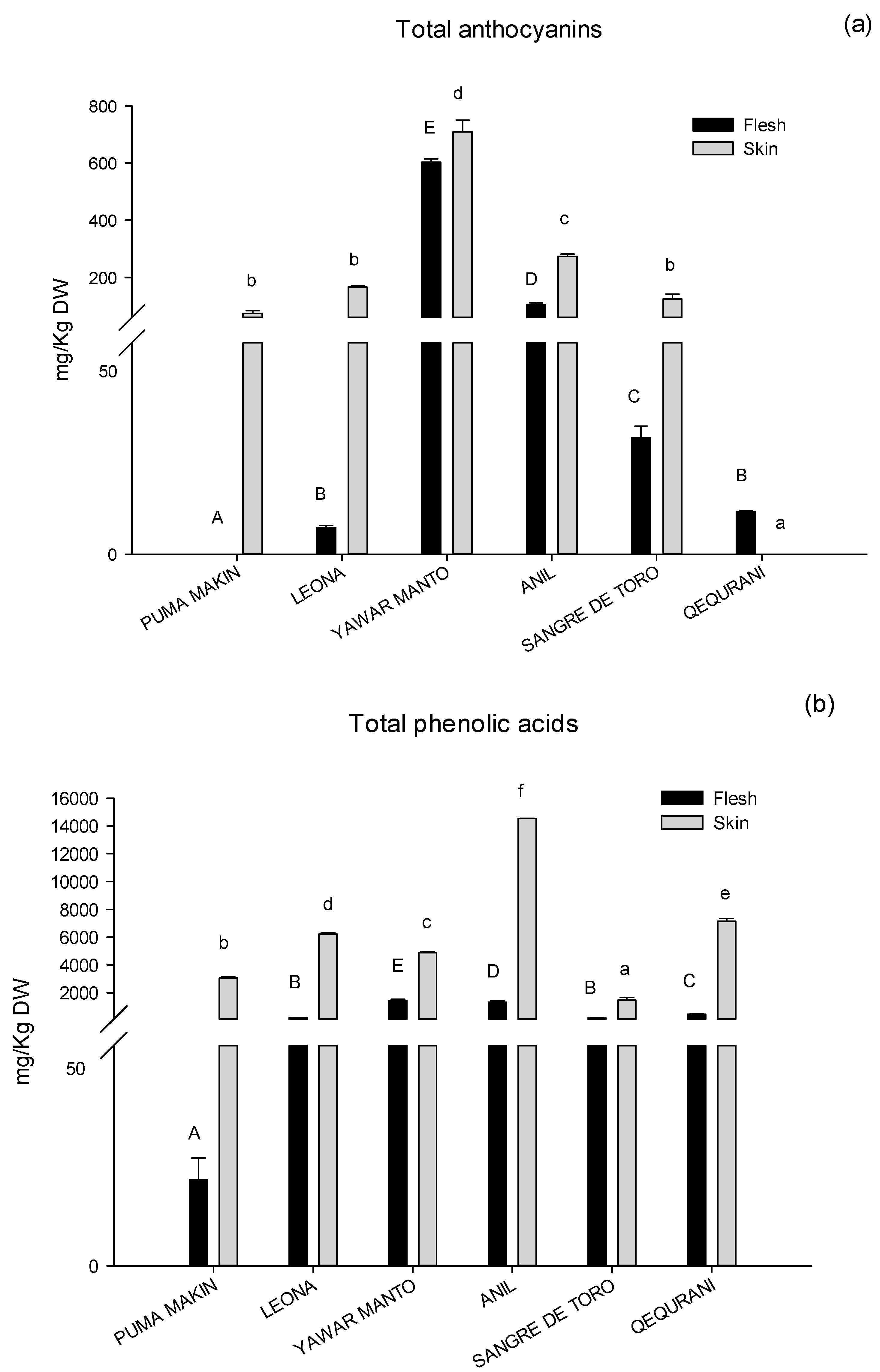
2.3. Phenolic Compounds and Carotenoids
3. Materials and Methods
3.1. Plant Materials and Sample Preparation
3.2. Proximate Analysis
3.3. Determination of the Mineral Content
3.4. Extraction of Phenolic Compounds
3.5. Extraction of Carotenoids
3.6. HPLC–DAD–MS Analysis
3.7. Quantitative Determination of Phenolic Compounds
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ministerio de Agricultura y Riego (MINAGRI); Grupo Yanapai; Instituto Nacional de Innovación Agraria (INIA); Centro Internacional de la Papa (CIP). Catálogo de Variedades de Papa Nativa del Sureste del Departamento de Junín-Perú; Centro Internacional de la Papa: Lima, Peru, 2017; 228p.
- Toledo, A.; Burlingame, B. Biodiversity and nutrition: A common path toward global food security and sustainable development. J. Food Compos. Anal. 2006, 19, 477–483. [Google Scholar] [CrossRef]
- Burlingame, B.; Mouille, B.; Charrondiere, R. Nutrients, bioactive non-nutrients and anti-nutrients in potatoes. J. Food Compos. Anal. 2009, 22, 494–502. [Google Scholar] [CrossRef]
- Galdon, B.R.; Rodriguez, L.H.; Mesa, D.R.; Leon, H.L.; Perez, N.L.; Rodriguez, E.M.R.; Romero, C.D. Differentiation of potato cultivars experimentally cultivated based on their chemical composition and by applying linear discriminant analysis. Food Chem. 2012, 133, 1241–1248. [Google Scholar] [CrossRef]
- Andre, C.M.; Ghislain, M.; Bertin, P.; Oufir, M.; Herrera, M.D.; Hoffmann, L.; Hausman, J.F.; Larondelle, Y.; Evers, D. Andean potato cultivars (Solanum tuberosum L.) as a source of antioxidant and mineral micronutrients. J. Agric. Food Chem. 2007, 55, 366–378. [Google Scholar] [CrossRef] [PubMed]
- Andre, C.M.; Oufir, M.; Hoffmann, L.; Hausman, J.F.; Rogez, H.; Larondelle, Y.; Evers, D. Influence of environment and genotype on polyphenol compounds and in vitro antioxidant capacity of native Andean potatoes (Solanum tuberosum L.). J. Food Compos. Anal. 2009, 22, 517–524. [Google Scholar] [CrossRef]
- Giusti, M.M.; Polit, M.F.; Ayvaz, H.; Tay, D.; Manrique, I. Characterization and Quantitation of Anthocyanins and Other Phenolics in Native Andean Potatoes. J. Agric. Food Chem. 2014, 62, 4408–4416. [Google Scholar] [CrossRef]
- Calliope, S.R.; Lobo, M.O.; Samman, N.C. Biodiversity of Andean potatoes: Morphological, nutritional and functional characterization. Food Chem. 2018, 238, 42–50. [Google Scholar] [CrossRef]
- Ieri, F.; Innocenti, M.; Andrenelli, L.; Vecchio, V.; Mulinacci, N. Rapid HPLC/DAD/MS method to determine phenolic acids, glycoalkaloids and anthocyanins in pigmented potatoes (Solanum tuberosum L) and correlations with variety and geographical origin. Food Chem. 2011, 125, 750–759. [Google Scholar] [CrossRef]
- Ezekiel, R.; Singh, N.; Sharma, S.; Kaur, A. Beneficial phytochemicals in potato—A review. Food Res. Int. 2013, 50, 487–496. [Google Scholar] [CrossRef]
- Bellumori, M.; Innocenti, M.; Michelozzi, M.; Cerretani, L.; Mulinacci, N. Coloured-fleshed potatoes after boiling: Promising sources of known antioxidant compounds. J. Food Compos. Anal. 2017, 59, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, M.E.; Rossi, A.M.; Samman, N.C. Phenotypic, agronomic and nutritional characteristics of seven varieties of Andean potatoes. J. Food Compos. Anal. 2009, 22, 613–616. [Google Scholar] [CrossRef]
- Lefevre, I.; Ziebel, J.; Guignard, C.; Hausman, J.F.; Rosales, R.O.G.; Bonierbale, M.; Hoffmann, L.; Schafleitner, R.; Evers, D. Drought Impacts Mineral Contents in Andean Potato Cultivars. J. Agron. Crop. Sci. 2012, 198, 196–206. [Google Scholar] [CrossRef]
- Camire, M.E.; Kubow, S.; Donnelly, D.J. Potatoes and Human Health. Crit. Rev. Food Sci. 2009, 49, 823–840. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Bradshaw, J.E.; Dale, M.F.B.; Ramsay, G.; Hammond, J.P.; Broadley, M.R. Relationships Between Yield and Mineral Concentrations in Potato Tubers. Hortscience 2009, 44, 6–11. [Google Scholar] [CrossRef]
- Subramanian, N.K.; White, P.J.; Broadley, M.R.; Ramsay, G. The three-dimensional distribution of minerals in potato tubers. Ann. Bot. Lond. 2011, 107, 681–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgos, G.; Amoros, W.; Morote, M.; Stangoulis, J.; Bonierbale, M. Iron and zinc concentration of native Andean potato cultivars from a human nutrition perspective. J. Sci. Food. Agric. 2007, 87, 668–675. [Google Scholar] [CrossRef]
- Anderson, K.A.; Magnuson, B.A.; Tschirgi, M.L.; Smith, B. Determining the geographic origin of potatoes with trace metal analysis using statistical and neural network classifiers. J. Agric. Food Chem. 1999, 47, 1568–1575. [Google Scholar] [CrossRef]
- Manzelli, M.; Romagnoli, S.; Ghiselli, L.; Benedettelli, S.; Palchetti, E.; Andrenelli, L.; Vecchio, V. Typicity in Potato: Characterization of Geographic Origin. Ital. J. Agron. 2010, 5, 61–67. [Google Scholar] [CrossRef]
- Lombardo, S.; Pandino, G.; Mauromicale, G. The influence of growing environment on the antioxidant and mineral content of “early” crop potato. J. Food Compos. Anal. 2013, 32, 28–35. [Google Scholar] [CrossRef]
- Tack, F.M.G. Trace Elements in Potato. Potato Res. 2014, 57, 311–325. [Google Scholar] [CrossRef]
- Lachman, J.; Hamouz, K.; Sulc, M.; Orsak, M.; Pivec, V.; Hejtmankova, A.; Dvorak, P.; Cepl, J. Cultivar differences of total anthocyanins and anthocyanidins in red and purple-fleshed potatoes and their relation to antioxidant activity. Food Chem. 2009, 114, 836–843. [Google Scholar] [CrossRef]
- Mulinacci, N.; Ieri, F.; Giaccherini, C.; Innocenti, M.; Andrenelli, L.; Canova, G.; Saracchi, M.; Casiraghi, M.C. Effect of Cooking on the Anthocyanins, Phenolic Acids, Glycoalkaloids, and Resistant Starch Content in Two Pigmented Cultivars of Solanum tuberosum L. J. Agric. Food Chem. 2008, 56, 11830–11837. [Google Scholar] [CrossRef] [PubMed]
- Valcarcel, J.; Reilly, K.; Gaffney, M.; O’Brien, N.M. Levels of potential bioactive compounds including carotenoids, vitamin C and phenolic compounds, and expression of their cognate biosynthetic genes vary significantly in different varieties of potato (Solanum tuberosum L.) grown under uniform cultural conditions. J. Sci. Food Agric. 2016, 96, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- Valinas, M.A.; Lanteri, M.L.; ten Have, A.; Andreu, A.B. Chlorogenic acid, anthocyanin and flavan-3-ol biosynthesis in flesh and skin of Andean potato tubers (Solanum tuberosum subsp andigena). Food Chem. 2017, 229, 837–846. [Google Scholar] [CrossRef]
- Navarre, D.A.; Pillai, S.S.; Shakya, R.; Holden, M.J. HPLC profiling of phenolics in diverse potato genotypes. Food Chem. 2011, 127, 34–41. [Google Scholar] [CrossRef]
- Deusser, H.; Guignard, C.; Hoffmann, L.; Evers, D. Polyphenol and glycoalkaloid contents in potato cultivars grown in Luxembourg. Food Chem. 2012, 135, 2814–2824. [Google Scholar] [CrossRef]
- Lachman, J.; Hamouz, K.; Orsak, M.; Pivec, V.; Hejtmankova, K.; Pazderu, K.; Dvorak, P.; Cepl, J. Impact of selected factors—Cultivar, storage, cooking and baking on the content of anthocyanins in coloured-flesh potatoes. Food Chem. 2012, 133, 1107–1116. [Google Scholar] [CrossRef]
- Borochov-Neori, H.; Judeinstein, S.; Harari, M.; Bar-Ya’akov, I.; Patil, B.S.; Lurie, S.; Holland, D. Climate Effects on Anthocyanin Accumulation and Composition in the Pomegranate (Punica granatum L.) Fruit Arils. J. Agric. Food Chem. 2011, 59, 5325–5334. [Google Scholar] [CrossRef]
- Hamouz, K.; Lachman, J.; Dvorak, P.; Juzl, M.; Pivec, V. The effect of site conditions, variety and fertilization on the content of polyphenols in potato tubers. Plant Soil Environ. 2006, 52, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Fogelman, E.; Oren-Shamir, M.; Hirschberg, J.; Mandolino, G.; Parisi, B.; Ovadia, R.; Tanami, Z.; Faigenboim, A.; Ginzberg, I. Nutritional value of potato (Solanum tuberosum) in hot climates: Anthocyanins, carotenoids, and steroidal glycoalkaloids. Planta 2019, 249, 1143–1155. [Google Scholar] [CrossRef]
- Figueiredo-Gonzalez, M.; Martinez-Carballo, E.; Cancho-Grande, B.; Santiago, J.L.; Martinez, M.C.; Simal-Gandara, J. Pattern recognition of three Vitis vinifera L. red grapes varieties based on anthocyanin and flavonol profiles, with correlations between their biosynthesis pathways. Food Chem. 2012, 130, 9–19. [Google Scholar] [CrossRef]
- Les, F.; Lopez, V.; Caprioli, G.; Iannarelli, R.; Fiorini, D.; Innocenti, M.; Bellumori, M.; Maggi, F. Chemical constituents, radical scavenging activity and enzyme inhibitory capacity of fruits from Cotoneaster pannosus Franch. Food Funct. 2017, 8, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Burgos, G.; Salas, E.; Amoros, W.; Auqui, M.; Munoa, L.; Kimura, M.; Bonierbale, M. Total and individual carotenoid profiles in Solanum phureja of cultivated potatoes: I. Concentrations and relationships as determined by spectrophotometry and HPLC. J. Food Compos. Anal. 2009, 22, 503–508. [Google Scholar] [CrossRef]
- Taipe Boza, A. Evaluación de la Diversidad Genética de Papa Nativa Cultivada (Solanum sp.) en la Comunidad Campesina de Paccho Molinos del Distrito de Paucará de la Provincia de Acobamba departamento de Huancavelica; Universidad Nacional de Huancavelica: Acobamba, Peru, 2017. [Google Scholar]
- Parejas Ccatamayo, J.C. Evaluación del Ataque del Nemátodo del Quiste (Globodera spp.) en el Cultivo de Papa en Condiciones de Paucará-Acobamba-Huancavelica; Universidad Nacional de Huancavelica: Acobamba, Peru, 2018. [Google Scholar]
- Escobar, B. Evaluación de Parámetros de Rendimiento de Cultivares y Líneas de Cebada (Hordeum Vulgare L.) en Paucará-Acombamba-Huancavelica; Universidad Nacional de Huancavelica: Acobamba, Peru, 2013. [Google Scholar]
Sample Availability: Not available. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Tuber Form | Skin Color | Flesh Color | Predominant Bud Color | Flower Color | Stem Color | ||
|---|---|---|---|---|---|---|---|---|
| Main | Secondary | |||||||
| Puma Makin | S. tuberosum andigenum | Oblong fingered | Blackish | Pale yellow | Violet | Purple | Pale purple | Purple |
| Leona | S. tuberosum andigenum | Round | Blackish | Purple | Yellow (few spots) | Purple | Pale violet | Purple |
| Yawar Manto | S. tuberosum andigenum | Elongate | Blackish | Dark red | - | Red | White with red dots | Red |
| Añil | S. tuberosum andigenum | Oblong | Blackish | Dark purple | - | Purple with white tip | Purple with white dots | Black |
| Sangre De Toro | S. goniocalyx | Round | Brown | Red | Yellow (few spots) | Yellow with red tip | Intense pink | Red |
| Qequrani | S. stenotomum | Oblong | Pale brown | Pale yellow | Violet | Purple | White | Green with spots |
| Puma Makin | Leona | Yawar Manto | Añil | Sangre De Toro | Qequrani | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Flesh | Skin | Flesh | Skin | Flesh | Skin | Flesh | Skin | Flesh | Skin | Flesh | Skin | |
| Whole tuber Kcal/100g | 129.8 | 106.4 | 117.4 | 91.4 | 90.3 | 99.4 | 98.1 | 131.0 | 131.1 | 150.4 | 150.6 | 105.3 |
| % Kcal from carbohydrates | 88.1 | 80.1 | 88.3 | 80.1 | 79.7 | 86.9 | 84.4 | 90.4 | 84.5 | 88.69 | 90.6 | 86.6 |
| Data Expressed as g/100 g | ||||||||||||
| Moisture | 66.6 | 72.7 | 69.8 | 76.8 | 76.5 | 74.4 | 73.9 | 66.2 | 66.8 | 61.9 | 61.4 | 72.4 |
| Carbohydrates | 28.6 | 21.3 | 25.6 | 18.3 | 18.0 | 21.6 | 20.7 | 29.7 | 27.7 | 33.3 | 34.1 | 22.8 |
| Fats | 0.2 | 0.4 | 0.2 | 0.6 | 0.3 | 0.2 | 0.1 | 0.2 | 0.7 | 0.4 | 0.2 | 0.1 |
| Proteins | 3.4 | 4.4 | 3 | 3.2 | 3.9 | 3.4 | 3.6 | 2.7 | 1.3 | 3.4 | 3.1 | 3.3 |
| Ash | 1.2 | 1.2 | 1.1 | 1.1 | 1.3 | 1.0 | 1.7 | 1.3 | 1.3 | 1.0 | 1.2 | 1.4 |
| Puma Makin | Leona | Yawar Manto | ||||
|---|---|---|---|---|---|---|
| Flesh | Skin | Flesh | Skin | Flesh | Skin | |
| Ca | 207.6 ± 18.4 B | 766.1 ± 19.5 c | 206.5 ± 10.7 B | 699.9 ± 14.9 b | 195.3 ± 12.1 B | 446.7 ± 22.4 a |
| K | 7272.9 ± 129.7 A | 13071.8 ± 102.8 b | 8040.3 ± 22.1 C | 14537.4 ± 230.3 d | 13059.9 ± 36.5 F | 17388.6 ± 211.1 e |
| Mg | 351.6 ± 14.8 B | 525.9 ± 12.0 b | 374.2 ± 7.2 C | 627.4 ± 19.9 c | 591.4 ± 8.7 F | 692.4 ± 14.9 d |
| Na | 19.9 ± 8.3 A | 33.1 ± 1.8 a | 17.5 ± 3.6 A | 31.1 ± 2.4 a | 31.1 ± 0.6 BC | 46.1 ± 3.0 b |
| P | 1140.7 ± 9.3 C | 1835.7 ± 6.3 c | 1181.6 ± 26.5 D | 2124.9 ± 24.9 e | 1252.1 ± 21.1 E | 1866.4 ± 33.2 c |
| Cu | 1.5 ± 0.0 B | 3.6 ± 0.1 d | 1.9 ± 0.0 C | 4.0 ± 0.0 e | 2.1 ± 0.0 C | 3.6 ± 0.0 d |
| Fe | 39.9 ± 6.5 C | 150.2 ± 10.7 b | 26.1 ± 1.2 AB | 112.2 ± 6.2 a | 38.9 ± 3.6 C | 178.4 ± 27.0 c |
| Mn | 5.6 ± 0.2 B | 7.6 ± 0.2 a | 5.5 ± 0.1 C | 7.9 ± 0.2 a | 9.8 ± 0.1 E | 18.8 ± 0.3 d |
| Zn | 7.4 ± 0.9 B | 11.4 ± 0.5 c | 6.1 ± 0.8 A | 10.9 ± 0.5 c | 10.2 ± 0.6 C | 11.0 ± 0.5 c |
| Anil | Sangre De Toro | Qequrani | ||||
| Flesh | Skin | Flesh | Skin | Flesh | Skin | |
| Ca | 126.2 ± 13.4 A | 443.4 ± 5.1 a | 361.9 ± 7.8 D | 953.5 ± 10.1 d | 248.4 ± 15.3 C | 943.6 ± 7.9 d |
| K | 10426.4 ± 88.7 E | 17124.2 ± 281.9 e | 7542.7 ± 60.7 B | 12418.0 ± 209.6 a | 8326.9 ± 80.5 D | 13907.8 ± 211.5 c |
| Mg | 528.0 ± 9.6 E | 833.1 ± 12.8 e | 312.1 ± 3.8 A | 414.2 ± 13.7 a | 446.5 ± 6.6 D | 516.2 ± 11.8 b |
| Na | 18.0 ± 0.2 A | 45.4 ± 3.8 b | 37.9 ± 3.3 C | 35.7 ± 6.5 a | 22.3 ± 9.9 AB | 39.9 ± 9.8 ab |
| P | 1074.1 ± 6.2 B | 1973.6 ± 49.5 d | 924.5 ± 11.4 A | 1464.4 ± 26.8 a | 1113.7 ± 21.5 C | 1669.1 ± 19.3 b |
| Cu | 1.0 ± 0.0 A | 2.3 ± 0.0 a | 1.5 ± 0.3 B | 2.8 ± 0.0 b | 1.2 ± 0.0 A | 2.9 ± 0.0 c |
| Fe | 20.5 ± 1.2 A | 178.1 ± 7.9 c | 38.7 ± 0.5 C | 250.2 ± 12.1 d | 26.8 ± 1.4 B | 288.8 ± 5.3 e |
| Mn | 6.7 ± 0.0 D | 18.8 ± 0.4 d | 4.7 ± 0.0 A | 11.4 ± 0.9 b | 5.1 ± 0.0 B | 14.4 ± 0.2 c |
| Zn | 7.2 ± 0.3 AB | 10.2 ± 0.1 b | 7.0 ± 0.5 AB | 9.0 ± 0.2 a | 6.3 ± 0.9 AB | 10.0 ± 0.3 b |
| Variety | Puma Makin | Leona | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Flesh | Skin | Flesh | Skin | |||||||||
| Compound | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | ||||
| pet 3-O-caf-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| pet 3-O-p-coum-rut-5-O-glu | nd | - | A | 43.6 | 7.1 | b | 5.35 | 0.5 | B | 121.4 | 3.6 | c |
| pet 3-O-ferul-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| peo 3-O-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| peo 3-O-p-coum-rut-5-O-glu | nd | - | A | 30.7 | 2.3 | b | 0,97 | 0.0 | B | 45.25 | 1.3 | c |
| peo 3-O-ferul-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| pel 3-O-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| pel 3-O-rut | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| pel 3-O-p-coum-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| pel 3-O-ferul-rut | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| pel 3-O-cis-p-coum-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| Variety | Yawar Manto | Anil | ||||||||||
| Flesh | Skin | Flesh | Skin | |||||||||
| Compound | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | ||||
| pet 3-O-caf-rut-5-O-glu | nd | - | A | nd | - | a | 4.1 | 0.1 | B | nd | - | a |
| pet 3-O-p-coum-rut-5-O-glu | nd | - | A | nd | - | a | 80.5 | 7.2 | C | 221.3 | 7.5 | d |
| pet 3-O-ferul-rut-5-O-glu | nd | - | A | nd | - | a | 6.1 | 0.6 | B | nd | - | a |
| peo 3-O-rut-5-O-glu | nd | - | A | 15.8 | 1.6 | b | nd | - | A | nd | - | a |
| peo 3-O-p-coum-rut-5-O-glu | nd | - | A | nd | - | a | 13.3 | 0.8 | D | 52.6 | 1.7 | d |
| peo 3-O-ferul-rut-5-O-glu | nd | - | A | 66.6 | 0.4 | b | nd | - | A | nd | - | a |
| pel 3-O-rut-5-O-glu | 15.0 | 0.4 | B | 52.9 | 4.2 | b | nd | - | A | nd | - | a |
| pel 3-O-rut | 508.2 | 11.9 | C | 574.1 | 35.2 | c | nd | - | A | nd | - | a |
| pel 3-O-p-coum-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| pel 3-O-ferul-rut | 54.3 | 1.1 | C | nd | - | a | nd | - | A | nd | - | a |
| pel 3-O-cis-p-coum-rut-5-O-glu | 25.4 | 0.1 | B | nd | - | a | nd | - | A | nd | - | a |
| Variety | Sangre De Toro | Qequrani | ||||||||||
| Flesh | Skin | Flesh | Skin | |||||||||
| Compound | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | ||||
| pet 3-O-caf-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| pet 3-O-p-coum-rut-5-O-glu | nd | - | A | nd | - | a | 6.8 | 0.0 | B | nd | - | a |
| pet 3-O-ferul-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| peo 3-O-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| peo 3-O-p-coum-rut-5-O-glu | nd | - | A | nd | - | a | 3.4 | 0.0 | C | nd | - | a |
| peo 3-O-ferul-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| pel 3-O-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| pel 3-O-rut | 23.5 | 2.9 | B | 113.5 | 17.4 | b | nd | - | A | nd | - | a |
| pel 3-O-p-coum-rut-5-O-glu | 1.1 | 0.0 | B | 3.9 | 0.2 | b | nd | - | A | nd | - | a |
| pel 3-O-ferul-rut | 2.9 | 0.2 | B | 6.8 | 0.4 | b | nd | - | A | nd | - | a |
| pel 3-O-cis-p-coum-rut-5-O-glu | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| Variety | Puma Makin | Leona | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Flesh | Skin | Flesh | Skin | |||||||||
| Compound | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | ||||
| cinnamic acid | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| 3-caffeoyquinic acid | nd | - | A | 48.7 | 0.8 | b | nd | - | A | 117.2 | 3.4 | d |
| 5-caffeoyquinic acid | 7.6 | 4.1 | A | 2221.2 | 26.3 | b | 156.4 | 17.7 | AB | 4807.8 | 79.8 | d |
| cinnamic acid | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| 4-caffeoyquinic acid | 11.9 | 1.4 | A | 210.0 | 2.2 | b | 12.0 | 0.9 | A | 865.7 | 10.0 | e |
| caffeic acid | nd | - | A | 500.0 | 33.3 | c | 19.7 | 1.3 | B | 299.6 | 14.9 | b |
| cinnamic acid | nd | - | A | 27.3 | 4.7 | b | 5.8 | 0.4 | B | 140.8 | 2.6 | d |
| cinnamic acid | nd | - | A | 28.6 | 0.6 | c | nd | - | A | nd | - | a |
| cinnamic acid | nd | - | A | 25.6 | 1.0 | b | nd | - | A | nd | - | a |
| Variety | Yawar Manto | Anil | ||||||||||
| Flesh | Skin | Flesh | Skin | |||||||||
| Compound | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | ||||
| cinnamic acid | 54.2 | 2.2 | B | nd | - | a | nd | - | A | nd | - | a |
| 3-caffeoyquinic acid | 58.7 | 4.8 | B | 76.0 | 0.4 | c | nd | - | A | 248.8 | 4.9 | f |
| 5-caffeoyquinic acid | 1046.9 | 88.7 | C | 3893.0 | 69.5 | c | 1172.1 | 78.9 | C | 12173.3 | 37.3 | f |
| cinnamic acid | 27.0 | 3.1 | C | nd | - | a | nd | - | A | nd | - | a |
| 4-caffeoyquinic acid | 56.7 | 4.8 | D | 595.8 | 8.1 | d | 53.8 | 4.4 | C | 1183.4 | 14.2 | f |
| caffeic acid | 87.9 | 0.9 | C | 216.7 | 5.8 | a | 23.2 | 1.8 | B | 579.2 | 56.1 | d |
| cinnamic acid | 25.4 | 2.6 | D | 48.2 | 6.9 | c | 59.4 | 4.5 | E | 348.5 | 17.7 | e |
| cinnamic acid | 35.1 | 2.5 | D | 49.9 | 1.6 | d | nd | - | A | nd | - | a |
| cinnamic acid | 20.4 | 1.1 | C | nd | - | a | nd | - | A | nd | - | a |
| Variety | Sangre De Toro | Qequrani | ||||||||||
| Flesh | Skin | Flesh | Skin | |||||||||
| Compound | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | mg/Kg | SD | ||||
| cinnamic acid | nd | - | A | nd | - | a | nd | - | A | nd | - | a |
| 3-caffeoyquinic acid | nd | - | A | nd | - | a | nd | - | A | 136.0 | 16.8 | e |
| 5-caffeoyquinic acid | 97.2 | 5.7 | A | 1114.3 | 166.3 | a | 363.7 | 19.3 | B | 5681.9 | 70.6 | e |
| cinnamic acid | 12.2 | 0.3 | B | 32.5 | 1.2 | b | nd | - | A | nd | - | a |
| 4-caffeoyquinic acid | 13.2 | 0.9 | A | 105.9 | 19.1 | a | 21.7 | 0.6 | B | 518.3 | 83.8 | c |
| caffeic acid | 17.3 | 0.9 | B | 177.1 | 16.2 | a | 34.5 | 3.3 | B | 809.1 | 40.4 | e |
| cinnamic acid | 10.5 | 0.6 | C | 30.0 | 2.7 | b | 5.5 | 0.0 | B | nd | - | a |
| cinnamic acid | 6.0 | 0.4 | B | 8.2 | 1.5 | b | 10.1 | 0.2 | C | nd | - | a |
| cinnamic acid | 14.9 | 0.3 | B | nd | - | a | nd | - | A | nd | - | a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellumori, M.; Chasquibol Silva, N.A.; Vilca, L.; Andrenelli, L.; Cecchi, L.; Innocenti, M.; Balli, D.; Mulinacci, N. A Study on the Biodiversity of Pigmented Andean Potatoes: Nutritional Profile and Phenolic Composition. Molecules 2020, 25, 3169. https://doi.org/10.3390/molecules25143169
Bellumori M, Chasquibol Silva NA, Vilca L, Andrenelli L, Cecchi L, Innocenti M, Balli D, Mulinacci N. A Study on the Biodiversity of Pigmented Andean Potatoes: Nutritional Profile and Phenolic Composition. Molecules. 2020; 25(14):3169. https://doi.org/10.3390/molecules25143169
Chicago/Turabian StyleBellumori, Maria, Nancy A. Chasquibol Silva, Laida Vilca, Luisa Andrenelli, Lorenzo Cecchi, Marzia Innocenti, Diletta Balli, and Nadia Mulinacci. 2020. "A Study on the Biodiversity of Pigmented Andean Potatoes: Nutritional Profile and Phenolic Composition" Molecules 25, no. 14: 3169. https://doi.org/10.3390/molecules25143169