
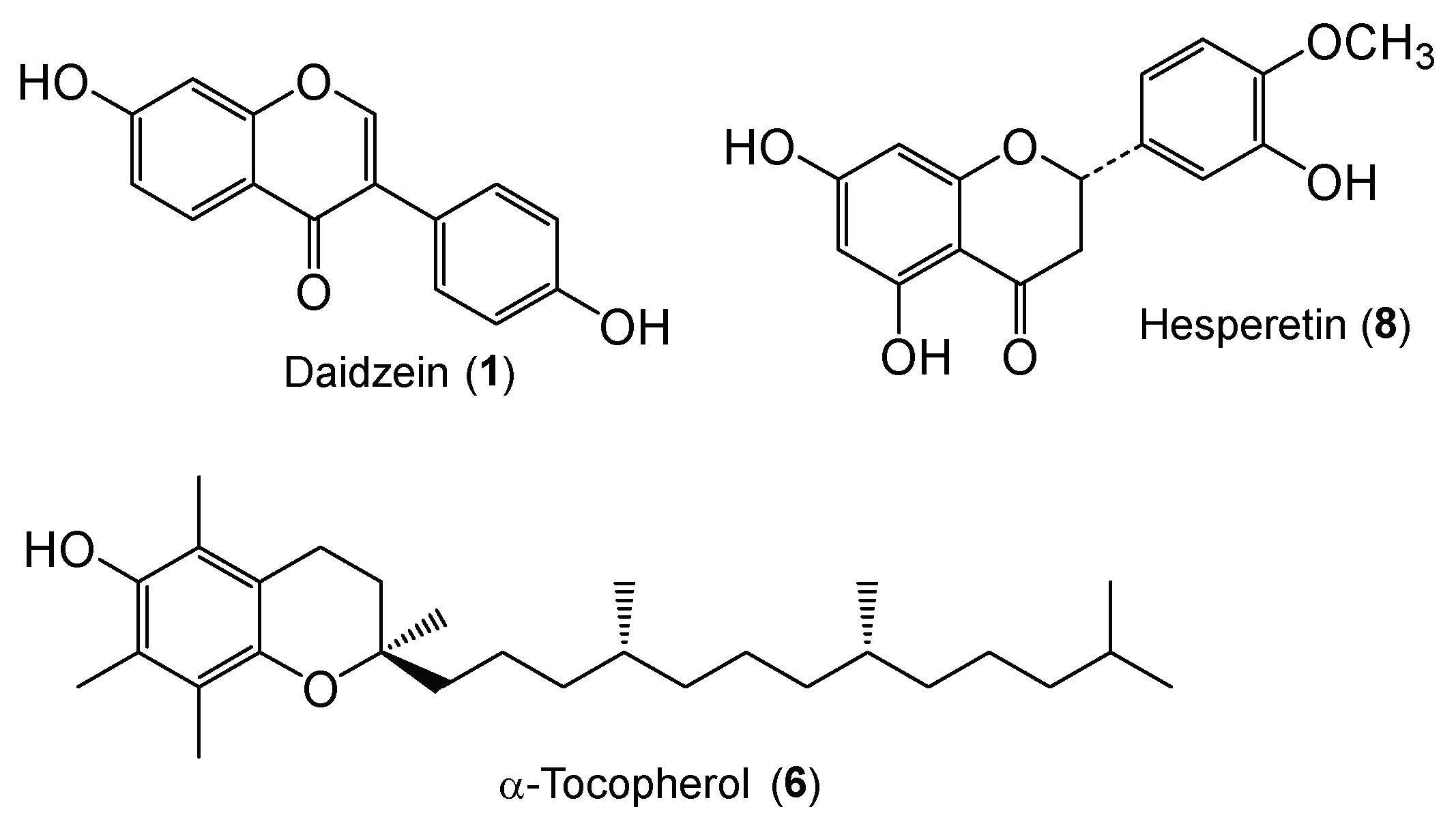
Synthesis of Daidzein Glycosides, α-Tocopherol Glycosides, Hesperetin Glycosides by Bioconversion and Their Potential for Anti-Allergic Functional-Foods and Cosmetics

Abstract
:1. Introduction
2. Results
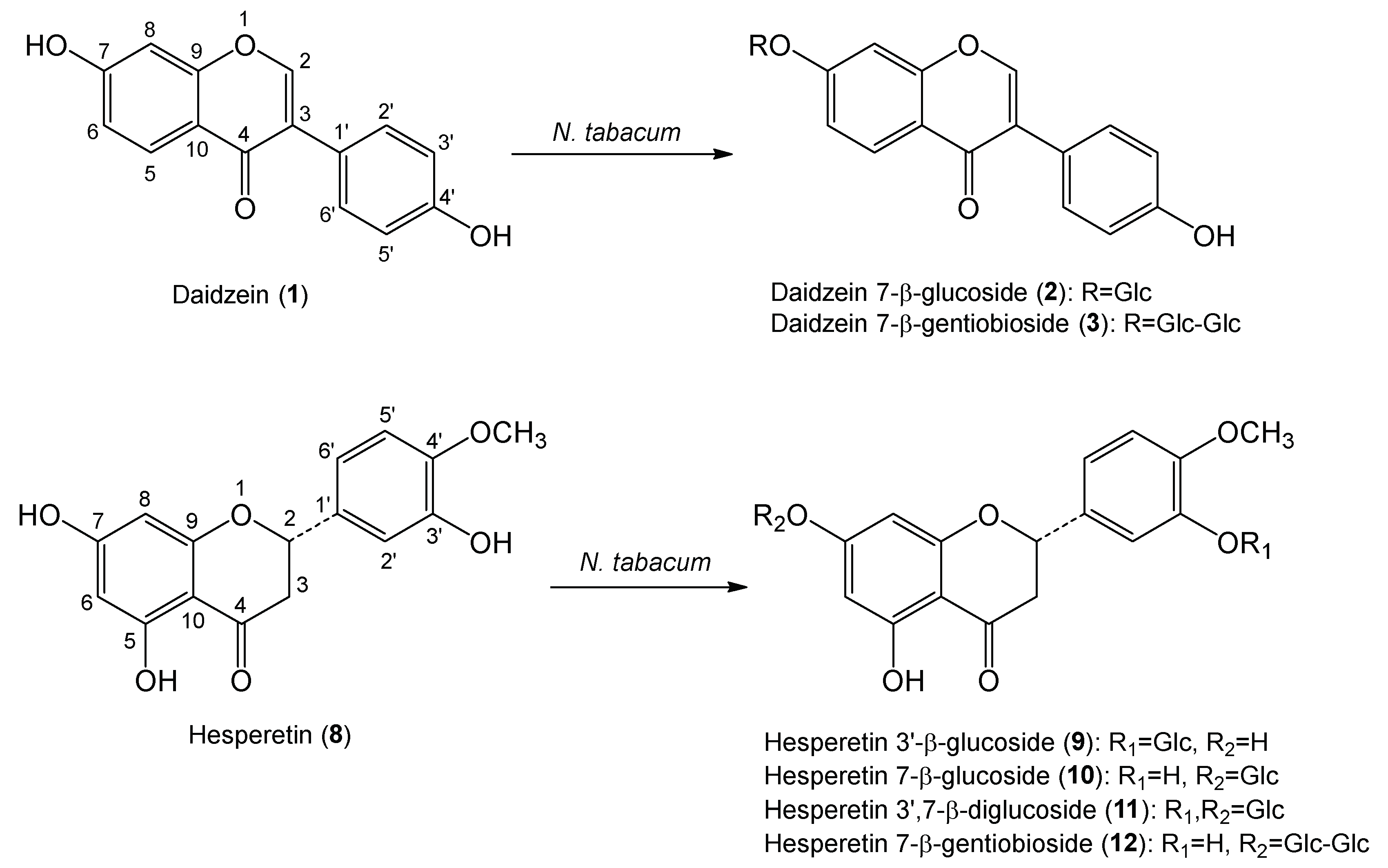
2.1. Synthesis of Daidzein Glycosides and Hesperetin Glycosides
2.2. Enzymatic Galactosylation with β-Glucosidase Isolated from Sweet Almond
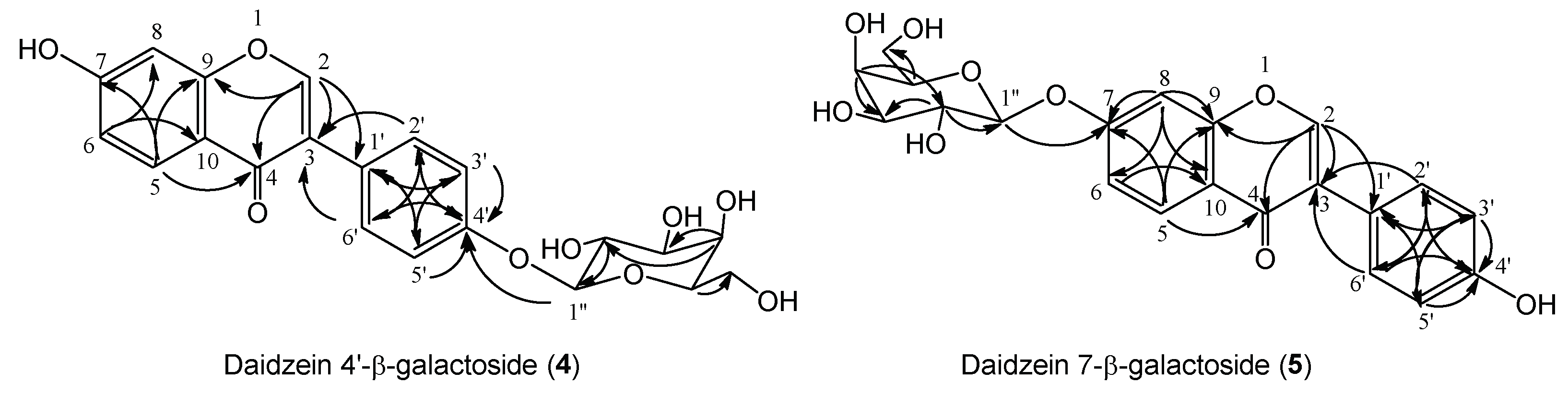
2.2.1. Synthesis of Daidzein Galactosides and α-Tocopherol Galactoside
2.2.2. Determination of Chemical Structure of New Daidzein Galactosides
2.3. Biological Effects of Daidzein Glycosides, α-Tocopherol Glycoside, and Hesperetin Glycosides
2.3.1. Suppression for Histamine Release from Rat Peritoneal Mast Cells
2.3.2. Tyrosinase Inhibitory Activity
3. Discussion
4. Materials and Methods
4.1. Analyses
4.2. Glycosylation by Cultured Plant Cells
4.3. Galactosylation by Enzyme
4.4. Inhibition of Histamine Release from Rat Peritoneal Mast Cells
4.5. Tyrosinase Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sak, K. Current epidemiological knowledge about the role of flavonoids in prostate carcinogenesis. Exp. Oncol. 2017, 39, 98–105. [Google Scholar] [CrossRef]
- Yu, B.; Tang, D.Z.; Li, S.Y.; Wu, Y.; Chen, M. Daidzein promotes proliferation and differentiation in osteoblastic OCT1 cells via activation of the BMP-2/Smads pathway. Pharmazie 2017, 72, 35–40. [Google Scholar] [PubMed]
- Zhang, F.; Ru, N.; Shang, Z.H.; Chen, J.F.; Yan, C.; Li, Y.; Liang, J. Daidzein ameliorates spinal cord ischemia/reperfusion injury-induced neurological function deficits in Sprague-Dawley rats through PI3K/Akt signaling pathway. Exp. Ther. Med. 2017, 14, 4878–4886. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Ejima, K.; Higuchi, T.; Takeshima, M.; Wakimoto, R.; Nakano, S. Equol enhances apoptosis-inducing activity of genistein by increasing Bax/Bcl-xL expression ratio in MCF-7 human breast cancer cells. Nutr. Cancer 2017, 69, 1300–1307. [Google Scholar] [CrossRef] [PubMed]
- Sak, K.; Lust, H.; Kase, M.; Saar, M.; Jaal, J. Suppression of taxanes cytotoxicity by citrus flavonoid hesperetin in PPC-1 human prostate cancer cells. Anticancer Res. 2018, 38, 6209–6215. [Google Scholar] [CrossRef] [PubMed]
- Kühn, G.; Pallauf, K.; Schulz, C.; Rimbach, G. Flavonoids as putative modulators of Δ4-, Δ5-, and Δ6-desaturases: Studies in cultured hepatocytes, myocytes, and adipocytes. Biofactors 2018, 44, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Hua, F.; Li, C.H.; Chen, X.G.; Liu, X.P. Daidzein exerts anticancer activity towards SKOV3 human ovarian cancer cells by inducing apoptosis and cell cycle arrest, and inhibiting the Raf/MEK/ERK cascade. Int. J. Mol. Med. 2018, 41, 3485–3492. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, S.; Shyam, H.; Sharma, R.; Balapure, A.K. Dietary isoflavone daidzein synergizes centchroman action via induction of apoptosis and inhibition of PI3K/Akt pathway in MCF-7/MDA MB-231 human breast cancer cells. Phytomedicine 2018, 40, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Sivoňová, M.K.; Kaplán, P.; Tatarková, Z.; Lichardusová, L.; Dušenka, R.; Jurečeková, J. Androgen receptor and soy isoflavones in prostate cancer. Mol. Clin. Oncol. 2019, 10, 191–204. [Google Scholar] [CrossRef]
- Grainger, E.M.; Moran, N.E.; Francis, D.M.; Schwartz, S.J.; Wan, L.; Thomas-Ahner, J.; Kopec, R.E.; Riedl, K.M.; Young, G.S.; Abaza, R.; et al. A Novel tomato-soy iuice induces a dose-response increase in urinary and plasma phytochemical biomarkers in men with prostate cancer. J. Nutr. 2019, 149, 26–35. [Google Scholar] [CrossRef]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Moretti, R.M.; Marelli, M.M.; Limonta, P. Tocotrienols and cancer: From the state of the art to promising novel patents. Recent Pat. Anticancer Drug Discov. 2019, 14, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Liese, J.; Hinrichs, T.M.; Lange, M.; Fulda, S. Cotreatment with sorafenib and oleanolic acid induces reactive oxygen species-dependent and mitochondrial-mediated apoptotic cell death in hepatocellular carcinoma cells. Anticancer Drugs 2019, 30, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.W.; Yu, Y.L.; Chen, Y.H.; Hung, Y.T.; Yiang, G.T. Anticancer effects of methotrexate in combination with α-tocopherol and α-tocopherol succinate on triple-negative breast cancer. Oncol Rep. 2019, 41, 2060–2066. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.; Lei, Q.; Qi, H.; Zhang, Y.; Cui, N.; Wu, X.; Xie, L.; Yan, X.; Lu, T. Amplification of oxidative stress in MCF-7 cells by a novel pH-responsive amphiphilic micellar system enhances anticancer therapy. Mol. Pharm. 2019, 16, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Cho, H.J. Mitochondria targeting and destabilizing hyaluronic acid derivative-based nanoparticles for the delivery of lapatinib to triple-negative breast cancer. Biomacromolecules 2019, 20, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Formica, J.V.; Regelson, W. Review of the biology of quercetin and related bioflavonoids. Food Chem. Toxicol. 1995, 33, 1061–1080. [Google Scholar] [CrossRef]
- So, F.V.; Guthrie, N.; Chambers, A.F.; Carroll, K.K. Inhibition of proliferation of estrogen receptor-positive MCF-7 human breast cancer cells by flavonoids in the presence and absence of excess estrogen. Cancer Lett. 1997, 112, 127–133. [Google Scholar] [CrossRef]
- Mitsunaga, Y.; Takanaga, H.; Matsuo, H.; Naito, M.; Tsuruo, T.; Ohtani, H.; Sawada, Y. Effect of bioflavonoids on vinicristine transport across blood-brain barrier. Euro. J. Pharm. 2000, 395, 193–201. [Google Scholar] [CrossRef]
- O’Prey, J.; Brown, J.; Fleming, J.; Harrison, P.R. Effects of dietary flavonoids on major signal transduction pathways in human epithelial cells. Biochem. Pharm. 2003, 66, 2075–2088. [Google Scholar] [CrossRef]
- Youdim, K.A.; Dobbie, M.S.; Kuhnle, G.; Proteggente, A.R.; Abbott, N.J.; Rice-Evans, C. Interaction between flavonoids and the blood-brain barrier: In vitro study. J. Neurochem. 2003, 85, 180–192. [Google Scholar] [CrossRef]
- Cooray, H.C.; Janvilisri, T.; Veen, H.W.; Hladky, S.B.; Barrand, M.A. Interaction of the breast cancer resistance protein with plant polyphenols. Biochem. Biophys. Res. Commun. 2004, 317, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, M.; Toyoda, M.; Teshima, R.; Sawada, J.; Hayashi, T.; Arisawa, M.; Shimizu, M.; Morita, N.; Inoue, S.; Saito, Y. In vitro antiallergic activity of flavonoids in histamine release assay using rat basophilic leukemia (RBL-2H3) cells. J. Food Hyg. Soc. Jpn. 1994, 35, 495–503. [Google Scholar] [CrossRef]
- Lee, Y.S.; Park, J.H.; Kim, M.H.; Seo, S.H.; Kim, H.J. Synthesis of tyrosinase inhibitory kojic acid derivative. Arch. Pharm. Chem. Life Sci. 2006, 339, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Han, L. Lipophilisation of caffeic acid through esterification with propanol using water-tolerable acidic ionic liquid as catalyst. J. Oleo Sci. 2015, 64, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Furuya, T.; Ushiyama, M.; Ashida, Y.; Yoshikawa, T. Biotransformation of 2-phenylpropionic acid in root culture of Panax ginseng. Phytochemistry 1989, 28, 483–487. [Google Scholar] [CrossRef]
- Kamel, S.; Brazier, M.; Desmet, G.; Fliniaux, M.-A.; Jacquin-Dubreuil, A.A. Glucosylation of butyric acid by cell suspension culture of Nicotiana plumbaginifolia. Phytochemistry 1992, 31, 1581–1583. [Google Scholar] [CrossRef]
- Morand, C.; Crespy, V.; Manach, C.; Besson, C.; Demigné, C.; Rémésy, C. Plasma metabolites of quercetin and their antioxidant properties. Am. J. Physiol. 1998, 275, R212–R219. [Google Scholar] [CrossRef] [PubMed]
- Moriguchi, Y.; Kita, M.; Hasegawa, S.; Omura, S. Molecular approach to citrus flavonoid and limonoid biosynthesis. J. Food Agri. Environ. 2003, 1, 22–25. [Google Scholar]
- Shimoda, K.; Kondo, Y.; Nishida, T.; Hamada, H.; Nakajima, N.; Hamada, H. Biotransformation of thymol, carvacrol, and eugenol by cultured cells of Eucalyptus perriniana. Phytochemistry 2006, 67, 2256–2261. [Google Scholar] [CrossRef]
- Takenaka, S.; Mulyono; Sasano, Y.; Takahashi, Y.; Murakami, S.; Aoki, K. Microbial transformation of aniline derivatives: Regioselective biotransformation and detoxification of 2-phenylenediamine by Bacillus cereus strain PDa-1. J. Biosci. Bioeng. 2006, 102, 21–27. [Google Scholar] [CrossRef]
- Yang, G.; Zhang, Z.; Bai, H.; Gong, J.; Wang, Y.; Li, B.; Li, J. Biotransformation of beta-amyrin acetate by Rhodobacter sphaeroides. J. Biosci. Bioeng. 2008, 105, 558–561. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Kitagawa, M.; Ishihara, K.; Masuoka, N.; Shimoda, K.; Nakajima, N.; Hamada, H. Glycosylation of trans-resveratrol by plant-cultured cells. Biosci. Biotechnol. Biochem. 2012, 8, 1552–1554. [Google Scholar] [CrossRef] [PubMed]
- Iwakiri, T.; Imai, H.; Hamada, H.; Nakayama, T.; Ozaki, S. Synthesis of 3,5,3′,4′-tetrahydroxy- trans-stilbene-4′-O-beta-d-glucopyranoside by glucosyltransferases from Phytolacca americana. Nat. Prod. Commun. 2013, 8, 119–120. [Google Scholar] [PubMed]
- Shimoda, K.; Sato, N.; Kobayashi, T.; Hamada, H.; Hamada, H. Glycosylation of daidzein by the Eucalyptus cell cultures. Phytochemistry 2008, 69, 2303–2306. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, K.; Hamada, H.; Hamada, H. Glycosylation of hesperetin by plant cell cultures. Phytochemistry 2008, 69, 1135–1140. [Google Scholar] [CrossRef] [PubMed]
- Ponrasu, T.; Charles, R.E.; Sivakumar, R.; Divakar, S. Syntheses of α-tocopheryl glycosides by glucosidases. Biotechnol. Lett. 2008, 30, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Akagi, M.; Katakuse, Y.; Fukuishi, N.; Kan, T.; Akagi, R. Superoxide anion-induced histamine release from rat peritoneal mast cells. Biol. Pharm. Bull. 1994, 17, 732–734. [Google Scholar] [CrossRef] [PubMed]
- Szeja, W.; Grynkiewicz, G.; Rusin, A. Isoflavones, their glycosides and glycoconjugates. Synthesis and biological activity. Curr. Org. Chem. 2017, 21, 218–235. [Google Scholar] [CrossRef]
- Chiang, C.-M.; Wang, T.-Y.; Yang, S.-Y.; Wu, J.-Y.; Chang, T.-S. Production of new isoflavone glucosides from glycosylation of 8-hydroxydaidzein by glycosyltransferase from Bacillus subtilis ATCC 6633. Catalysts 2018, 8, 387. [Google Scholar] [CrossRef]
- Sato, D.; Shimizu, N.; Shimizu, Y.; Akagi, M.; Eshita, Y.; Ozaki, S.; Nakajima, N.; Ishihara, K.; Masuoka, N.; Hamada, H.; et al. Synthesis of glycosides of resveratrol, pterostilbene, and piceatannol, and their anti-oxidant, anti-allergic, and neuroprotective activities. Biosci. Biotechnol. Biochem. 2014, 78, 1123–1128. [Google Scholar] [CrossRef]
- Uesugi, D.; Hamada, H.; Shimoda, K.; Kubota, N.; Ozaki, S.; Nagatani, N. Synthesis, oxygen radical absorbance capacity, and tyrosinase inhibitory activity of glycosides of resveratrol, pterostilbene, and pinostilbene. Biosci. Biotechnol. Biochem. 2017, 81, 226–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Not available. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Histamine Release-Inhibiting Activity / %inhibition 1 |
|---|---|
| Kaempferol | 80 |
| 1 | 58 |
| 2 | 62 |
| 3 | 67 |
| 4 | 70 |
| 5 | 76 |
| 6 | 61 |
| 7 | 82 |
| 8 | 38 |
| 9 | 40 |
| 10 | 45 |
| 11 | 63 |
| 12 | 69 |
| Compound | Tyrosinase Inhibitory Activity IC50 1 / μM |
|---|---|
| Kojic acid | 35 ± 15 |
| 1 | 392 ± 88 |
| 2 | 303 ± 45 |
| 3 | 280 ± 33 |
| 4 | 125 ± 41 |
| 5 | 102 ± 39 |
| 6 | 510 ± 108 |
| 7 | 54 ± 25 |
| 8 | 437 ± 76 |
| 9 | 355 ± 32 |
| 10 | 318 ± 27 |
| 11 | 176 ± 35 |
| 12 | 139 ± 14 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujitaka, Y.; Hamada, H.; Uesugi, D.; Kuboki, A.; Shimoda, K.; Iwaki, T.; Kiriake, Y.; Saikawa, T. Synthesis of Daidzein Glycosides, α-Tocopherol Glycosides, Hesperetin Glycosides by Bioconversion and Their Potential for Anti-Allergic Functional-Foods and Cosmetics. Molecules 2019, 24, 2975. https://doi.org/10.3390/molecules24162975
Fujitaka Y, Hamada H, Uesugi D, Kuboki A, Shimoda K, Iwaki T, Kiriake Y, Saikawa T. Synthesis of Daidzein Glycosides, α-Tocopherol Glycosides, Hesperetin Glycosides by Bioconversion and Their Potential for Anti-Allergic Functional-Foods and Cosmetics. Molecules. 2019; 24(16):2975. https://doi.org/10.3390/molecules24162975
Chicago/Turabian StyleFujitaka, Yuya, Hiroki Hamada, Daisuke Uesugi, Atsuhito Kuboki, Kei Shimoda, Takafumi Iwaki, Yuya Kiriake, and Tomohiro Saikawa. 2019. "Synthesis of Daidzein Glycosides, α-Tocopherol Glycosides, Hesperetin Glycosides by Bioconversion and Their Potential for Anti-Allergic Functional-Foods and Cosmetics" Molecules 24, no. 16: 2975. https://doi.org/10.3390/molecules24162975