Kynurenic Acid and Its Analogs Are Beneficial Physiologic Attenuators in Bdelloid Rotifers

 , , ,
, , ,  ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
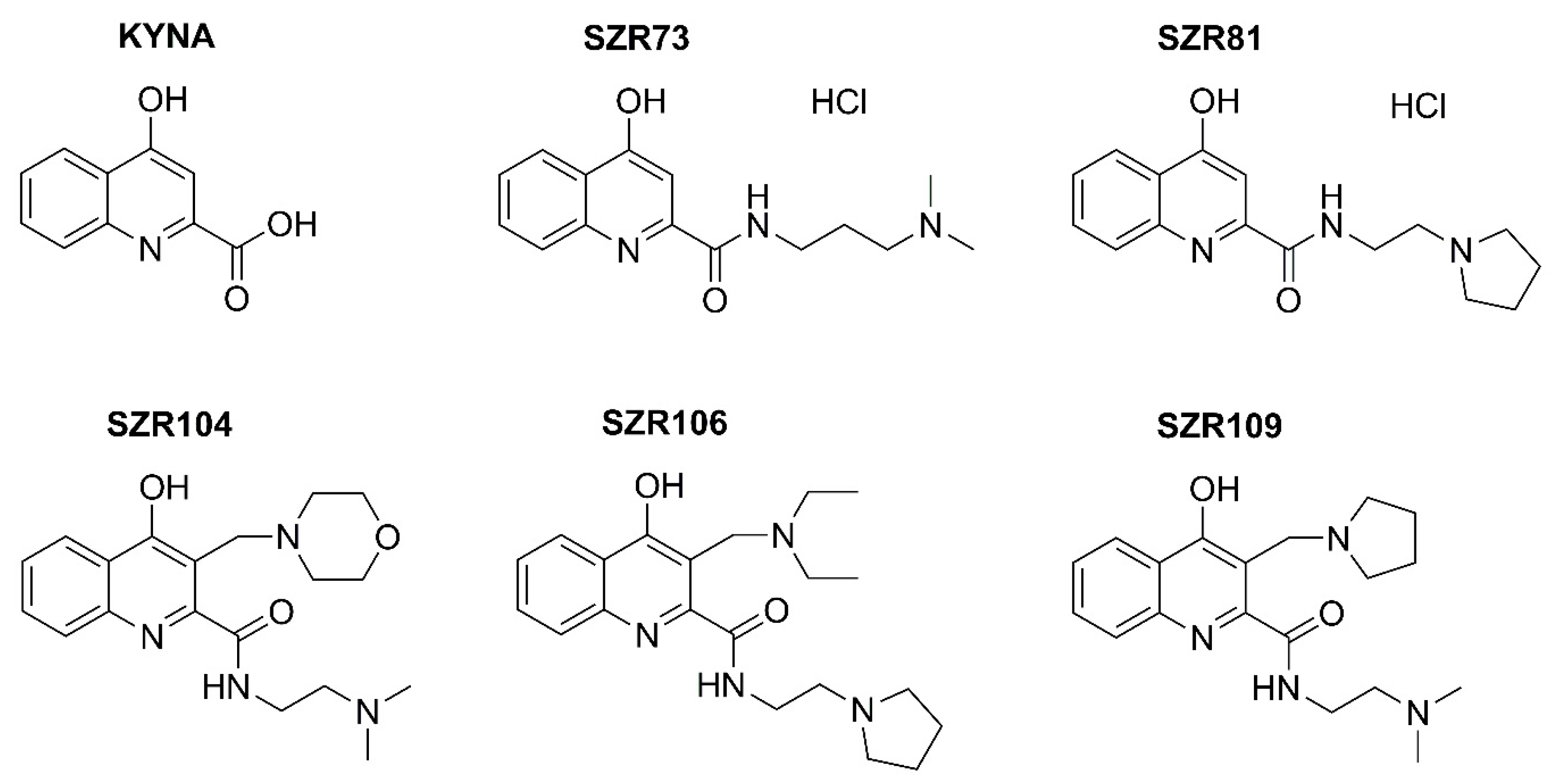
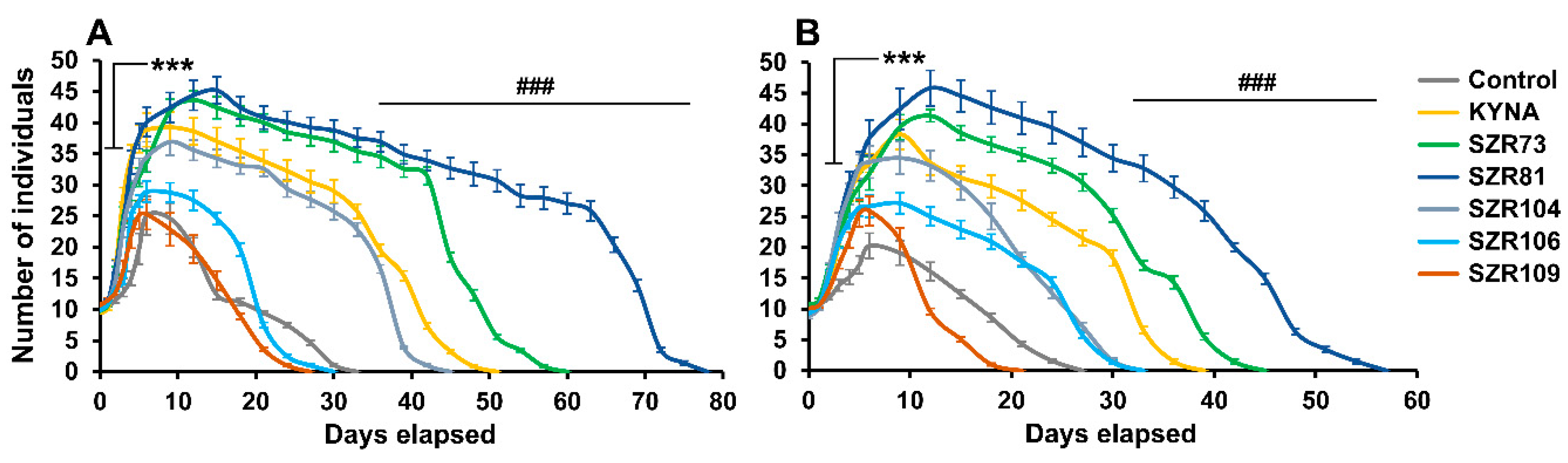
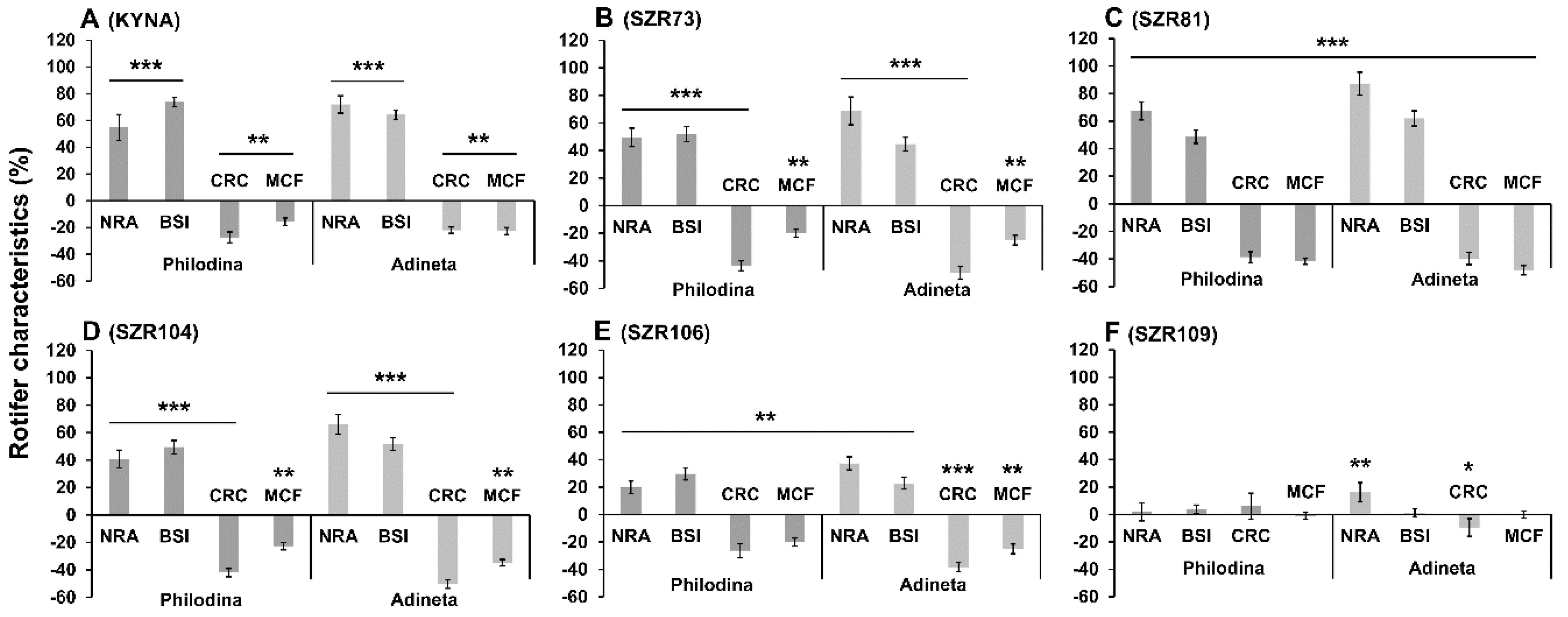
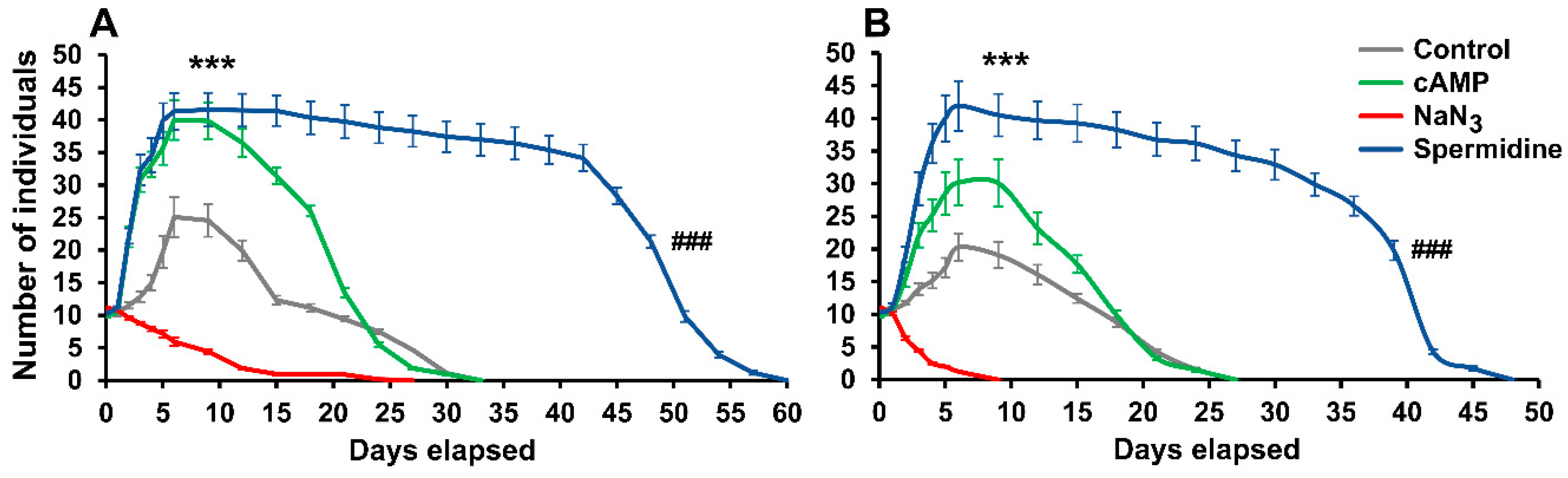
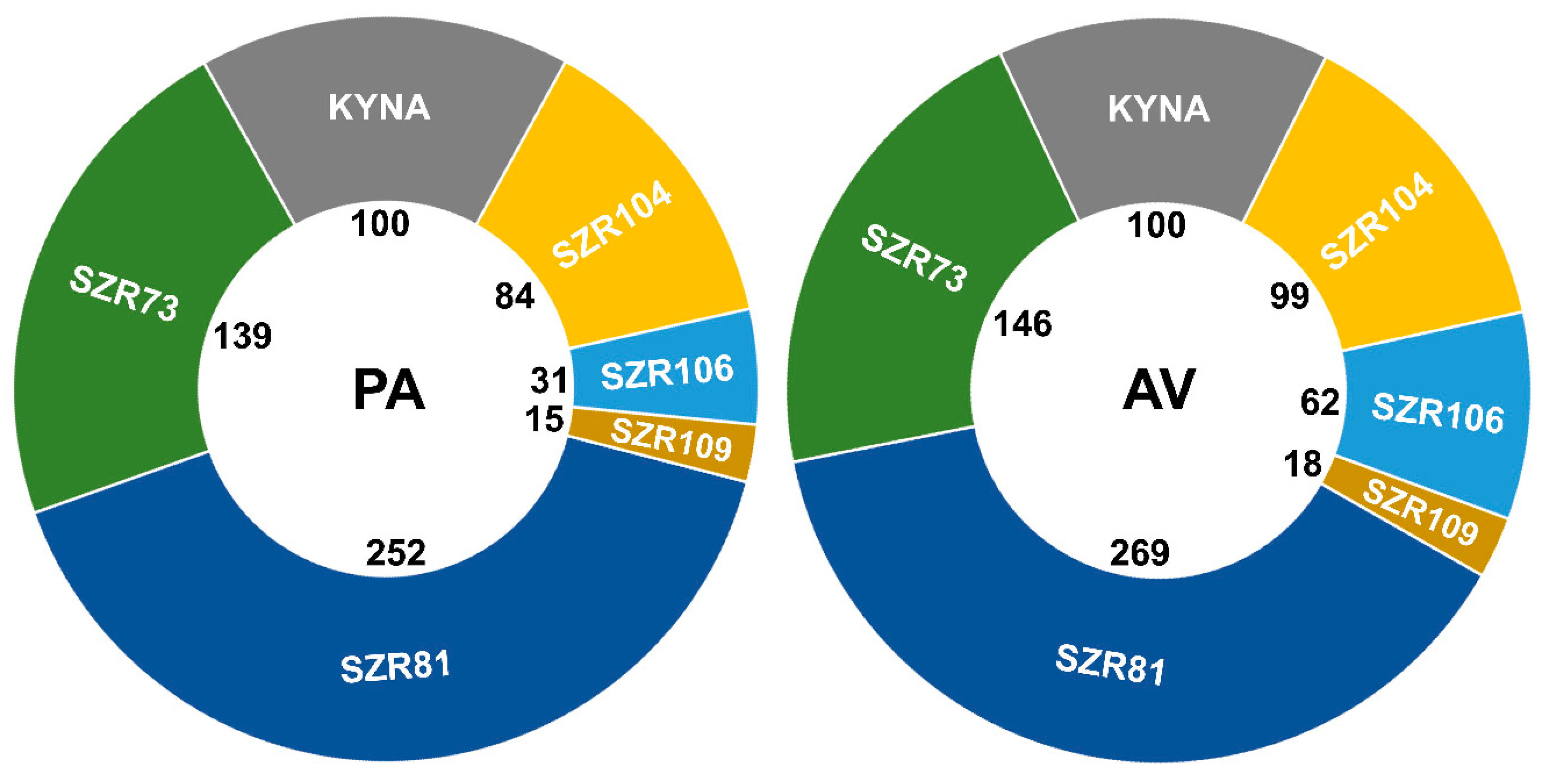
2. Results and Discussion
3. Materials and Methods
3.1. Materials
3.1.1. The Invertebrate Models
3.1.2. Treatment and Monitoring
3.1.3. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sas, K.; Szabó, E.; Vécsei, L. Mitochondria, Oxidative Stress and the Kynurenine System, with a Focus on Ageing and Neuroprotection. Molecules 2018, 23, 191. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.P.; Guillemin, G.J.; Brew, B.J. The kynurenine pathway in stem cell biology. Int. J. Tryptophan. Res. 2013, 6, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Stone, T.W. Kynurenines in the CNS: From endogenous obscurity to therapeutic importance. Prog. Neurobiol. 2001, 64, 185–218. [Google Scholar] [CrossRef]
- Moroni, F.; Cozzi, A.; Sili, M.; Mannaioni, G. Kynurenic acid: A metabolite with multiple actions and multiple targets in brain and periphery. J. Neural. Transm. (Vienna) 2012, 119, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Tuboly, G.; Tar, L.; Bohar, Z.; Safrany-Fark, A.; Petrovszki, Z.; Kekesi, G.; Vecsei, L.; Pardutz, A.; Horvath, G. The inimitable kynurenic acid: The roles of different ionotropic receptors in the action of kynurenic acid at a spinal level. Brain Res. Bull 2015, 112, 52–60. [Google Scholar] [CrossRef]
- Kepplinger, B.; Baran, H.; Kainz, A.; Ferraz-Leite, H.; Newcombe, J.; Kalina, P. Age-related increase of kynurenic acid in human cerebrospinal fluid–IgG and beta2-microglobulin changes. Neurosignals 2005, 14, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Vohra, M.; Lemieux, G.A.; Lin, L.; Ashrafi, K. Kynurenic acid accumulation underlies learning and memory impairment associated with aging. Genes Dev. 2018, 1, 14–19. [Google Scholar] [CrossRef]
- Stone, T.W. Development and therapeutic potential of kynurenic acid and kynurenine derivatives for neuroprotection. Trends Pharmacol. Sci. 2000, 21, 149–154. [Google Scholar] [CrossRef]
- Lemieux, G.A.; Ashrafi, K. Neural Regulatory Pathways of Feeding and Fat in Caenorhabditis elegans. Annu. Rev. Genet 2015, 49, 413–438. [Google Scholar] [CrossRef]
- Cunningham, K.A.; Hua, Z.; Srinivasan, S.; Liu, J.; Lee, B.H.; Edwards, R.H.; Ashrafi, K. AMP-activated kinase links serotonergic signaling to glutamate release for regulation of feeding behavior in C. elegans. Cell Metab. 2012, 16, 113–121. [Google Scholar] [CrossRef]
- Snell, T.W.; Johnston, R.K.; Gribble, K.E.; Mark Welch, D.B. Rotifers as experimental tools for investigating aging. Invertebr. Reprod. Dev. 2015, 59, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Leasi, F.; Pennati, R.; Ricci, C. First description of the serotonergic nervous system in a bdelloid rotifer: Macrotrachela quadricornifera Milne 1886 (Philodinidae). Zoologischer Anzeiger 2009, 248, 47–55. [Google Scholar] [CrossRef]
- Leasi, F.; Ricci, C. The role of serotonin in a bdelloid life cycle. Hydrobiologia 2011, 662, 141–147. [Google Scholar] [CrossRef]
- Snell, T.W.; Johnston, R.K.; Matthews, A.B. Freshwater toxicity testing using rehydrated Philodina sp. (Rotifera) as test animals. Environ. Toxicol. 2017, 32, 2267–2276. [Google Scholar] [CrossRef] [PubMed]
- Macsai, L.; Olah, Z.; Bush, A.I.; Galik, B.; Onody, R.; Kalman, J.; Datki, Z. Redox Modulating Factors Affect Longevity Regulation in Rotifers. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 811–814. [Google Scholar] [CrossRef] [PubMed]
- Olah, Z.; Bush, A.I.; Aleksza, D.; Galik, B.; Ivitz, E.; Macsai, L.; Janka, Z.; Karman, Z.; Kalman, J.; Datki, Z. Novel in vivo experimental viability assays with high sensitivity and throughput capacity using a bdelloid rotifer. Ecotoxicol. Environ. Saf. 2017, 144, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Most, J.; Tosti, V.; Redman, L.M.; Fontana, L. Calorie restriction in humans: An update. Ageing Res. Rev. 2016, 39, 36–45. [Google Scholar] [CrossRef]
- Veres, G.; Fejes-Szabó, A.; Zádori, D.; Nagy-Grócz, G.; László, A.M.; Bajtai, A.; Mándity, I.; Szentirmai, M.; Bohár, Z.; Laborc, K.; et al. A comparative assessment of two kynurenic acid analogs in the formalin model of trigeminal activation: A behavioral, immunohistochemical and pharmacokinetic study. J. Neural. Transm. (Vienna) 2017, 124, 99–122. [Google Scholar] [CrossRef]
- Turski, M.P.; Turska, M.; Zgrajka, W.; Bartnik, M.; Kocki, T.; Turski, W.A. Distribution, synthesis, and absorption of kynurenic acid in plants. Planta Med. 2011, 77, 858–864. [Google Scholar] [CrossRef]
- Turski, M.P.; Zgrajka, W.; Siwicki, A.K.; Paluszkiewicz, P. Presence and content of kynurenic acid in animal feed. J. Anim. Physiol. Anim. Nutr. (Berl) 2015, 99, 73–78. [Google Scholar] [CrossRef]
- Mariano, D.O.; Yamaguchi, L.F.; Jared, C.; Antoniazzi, M.M.; Sciani, J.M.; Kato, M.J.; Pimenta, D.C. Pipa carvalhoi skin secretion profiling: Absence of peptides and identification of kynurenic acid as the major constitutive component. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2015, 167, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Małaczewska, J.; Siwicki, A.K.; Wójcik, R.; Turski, W.A.; Kaczorek, E. The in vitro effect of kynurenic acid on the rainbow trout (Oncorhynchus mykiss) leukocyte and splenocyte activity. Pol. J. Vet. Sci. 2014, 17, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Kaczorek, E.; Szarek, J.; Mikiewicz, M.; Terech-Majewska, E.; Schulz, P.; Małaczewska, J.; Wójcik, R.; Siwicki, A.K. Effect of feed supplementation with kynurenic acid on the morphology of the liver, kidney and gills in rainbow trout (Oncorhynchus mykiss Walbaum, 1792), healthy and experimentally infected with Yersinia ruckeri. J. Fish Dis. 2017, 40, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Snell, T.W.; Johnston, R.K.; Matthews, A.B.; Zhou, H.; Gao, M.; Skolnick, J. Repurposed FDA-approved drugs targeting genes influencing aging can extend lifespan and healthspan in rotifers. Biogerontology 2018, 19, 145–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuluunbaatar, B.; Béni, Z.; Dékány, M.; Kovács, B.; Sárközy, A.; Datki, Z.; Mácsai, L.; Kálmán, J.; Hohmann, J.; Ványolós, A. Triterpenes from the Mushroom Hypholoma lateritium: Isolation, Structure Determination and Investigation in Bdelloid Rotifer Assays. Molecules 2019, 24, 301. [Google Scholar] [CrossRef] [PubMed]
- Datki, Z.; Olah, Z.; Hortobagyi, T.; Macsai, L.; Zsuga, K.; Fulop, L.; Bozso, Z.; Galik, B.; Acs, E.; Foldi, A.; et al. Exceptional in vivo catabolism of neurodegeneration-related aggregates. Acta Neuropathol. Commun. 2018, 6, 6. [Google Scholar] [CrossRef]
- Ma, S.; Gladyshev, V.N. Molecular signatures of longevity: Insights from cross-species comparative studies. Semin Cell Dev. Biol. 2017, 70, 190–203. [Google Scholar] [CrossRef]
- Poeggeler, B.; Sambamurti, K.; Siedlak, S.L.; Perry, G.; Smith, M.A.; Pappolla, M.A. A novel endogenous indole protects rodent mitochondria and extends rotifer lifespan. PLoS ONE 2010, 5, 10206. [Google Scholar] [CrossRef]
- Smith, R.L.; Soeters, M.R.; Wüst, R.C.I.; Houtkooper, R.H. Metabolic Flexibility as an Adaptation to Energy Resources and Requirements in Health and Disease. Endocr. Rev. 2018, 39, 489–517. [Google Scholar] [CrossRef] [Green Version]
- Marotta, R.; Uggetti, A.; Ricci, C.; Leasi, F.; Melone, G. Surviving starvation: Changes accompanying starvation tolerance in a bdelloid rotifer. J. Morphol. 2012, 2731, 1–7. [Google Scholar] [CrossRef]
- Jazwinski, S.M.; Jiang, J.C.; Kim, S. Adaptation to metabolic dysfunction during aging: Making the best of a bad situation. Exp. Gerontol. 2018, 107, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Comley, J.C.W.; Turner, C.H. Potential of a soluble tetrazolium/formazan assay for the evaluation of filarial viability. Int. J. Parasitol. 1990, 20, 251–255. [Google Scholar] [CrossRef]
- López-Lluch, G.; Navas, P. Calorie restriction as an intervention in ageing. J. Physiol. 2016, 594, 2043–2060. [Google Scholar] [CrossRef] [PubMed]
- Bock, M.J.; Jarvis, G.C.; Corey, E.L.; Stone, E.E.; Gribble, K.E. Maternal age alters offspring lifespan, fitness, and lifespan extension under caloric restriction. Sci. Rep. 2019, 9, 3138. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-J.; Ford, E.; Haigis, M.; Liszt, G.; Guarente, L. Calorie restriction extends yeast life span by lowering the level of NADH. Genes Dev. 2004, 18, 12–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, C.; Bogan, K.L.; Song, P.; Burant, C.F.; Kennedy, R.T.; Brenner, C. NAD+ metabolite levels as a function of vitamins and calorie restriction: Evidence for different mechanisms of longevity. BMC Chem. Biol. 2010, 10, 2. [Google Scholar] [CrossRef]
- Trubitsyn, A.G. The Lag of the Proliferative Aging Clock Underlies the Lifespan-Extending Effect of Calorie Restriction. Curr. Aging Sci. 2015, 8, 220–226. [Google Scholar] [CrossRef]
- Latta, L.C.; Tucker, K.N.; Haney, R.A. The relationship between oxidative stress, reproduction, and survival in a bdelloid rotifer. BMC Ecol. 2019, 19, 7. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, L.; Liang, Y.; Zhang, C.; Xu, Z.; Zhang, L.; Fuji, R.; Mu, W.; Li, L.; Jiang, J.; et al. Cyclic AMP Mimics the Anti-ageing Effects of Calorie Restriction by Up-Regulating Sirtuin. Sci. Rep. 2015, 5, 12012. [Google Scholar] [CrossRef]
- Ye, J.; Jiang, Z.; Chen, X.; Liu, M.; Li, J.; Liu, N. Electron transport chain inhibitors induce microglia activation through enhancing mitochondrial reactive oxygen species production. Exp. Cell Res. 2016, 340, 315–326. [Google Scholar] [CrossRef]
- Shintani, H.; Shintani, T.; Ashida, H.; Sato, M. Calorie Restriction Mimetics: Upstream-Type Compounds for Modulating Glucose Metabolism. Nutrients 2018, 10, 1821. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the SZR compounds are available from the authors. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Datki, Z.; Galik-Olah, Z.; Bohar, Z.; Zadori, D.; Fulop, F.; Szatmari, I.; Galik, B.; Kalman, J.; Vecsei, L. Kynurenic Acid and Its Analogs Are Beneficial Physiologic Attenuators in Bdelloid Rotifers. Molecules 2019, 24, 2171. https://doi.org/10.3390/molecules24112171
Datki Z, Galik-Olah Z, Bohar Z, Zadori D, Fulop F, Szatmari I, Galik B, Kalman J, Vecsei L. Kynurenic Acid and Its Analogs Are Beneficial Physiologic Attenuators in Bdelloid Rotifers. Molecules. 2019; 24(11):2171. https://doi.org/10.3390/molecules24112171
Chicago/Turabian StyleDatki, Zsolt, Zita Galik-Olah, Zsuzsanna Bohar, Denes Zadori, Ferenc Fulop, Istvan Szatmari, Bence Galik, Janos Kalman, and Laszlo Vecsei. 2019. "Kynurenic Acid and Its Analogs Are Beneficial Physiologic Attenuators in Bdelloid Rotifers" Molecules 24, no. 11: 2171. https://doi.org/10.3390/molecules24112171