New Hydrophilic/Lipophilic Tetra-α-(4-carboxyphenoxy) Phthalocyanine Zinc-Mediated Photodynamic Therapy Inhibits the Proliferation of Human Hepatocellular Carcinoma Bel-7402 Cells by Triggering Apoptosis and Arresting Cell Cycle

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
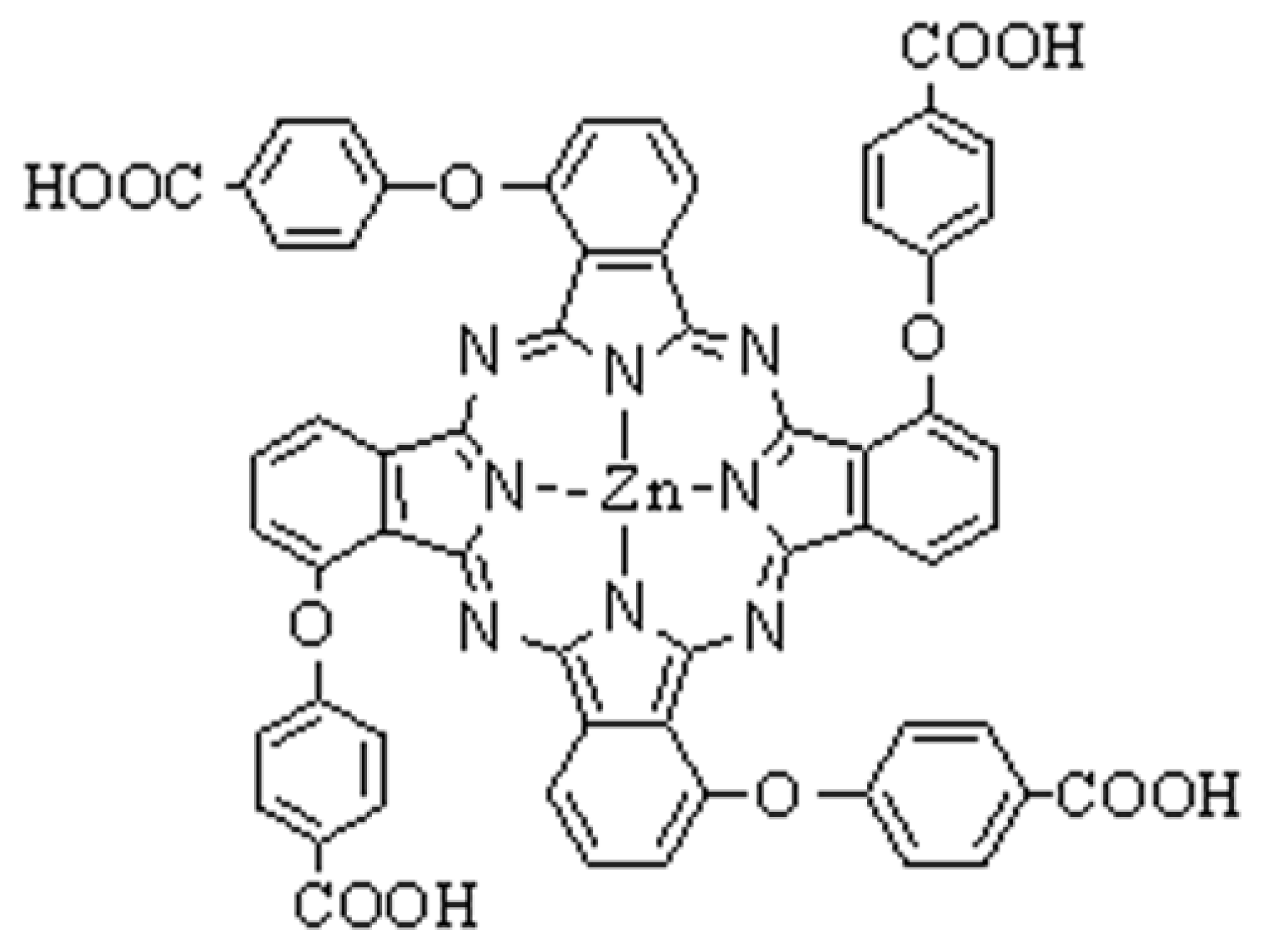
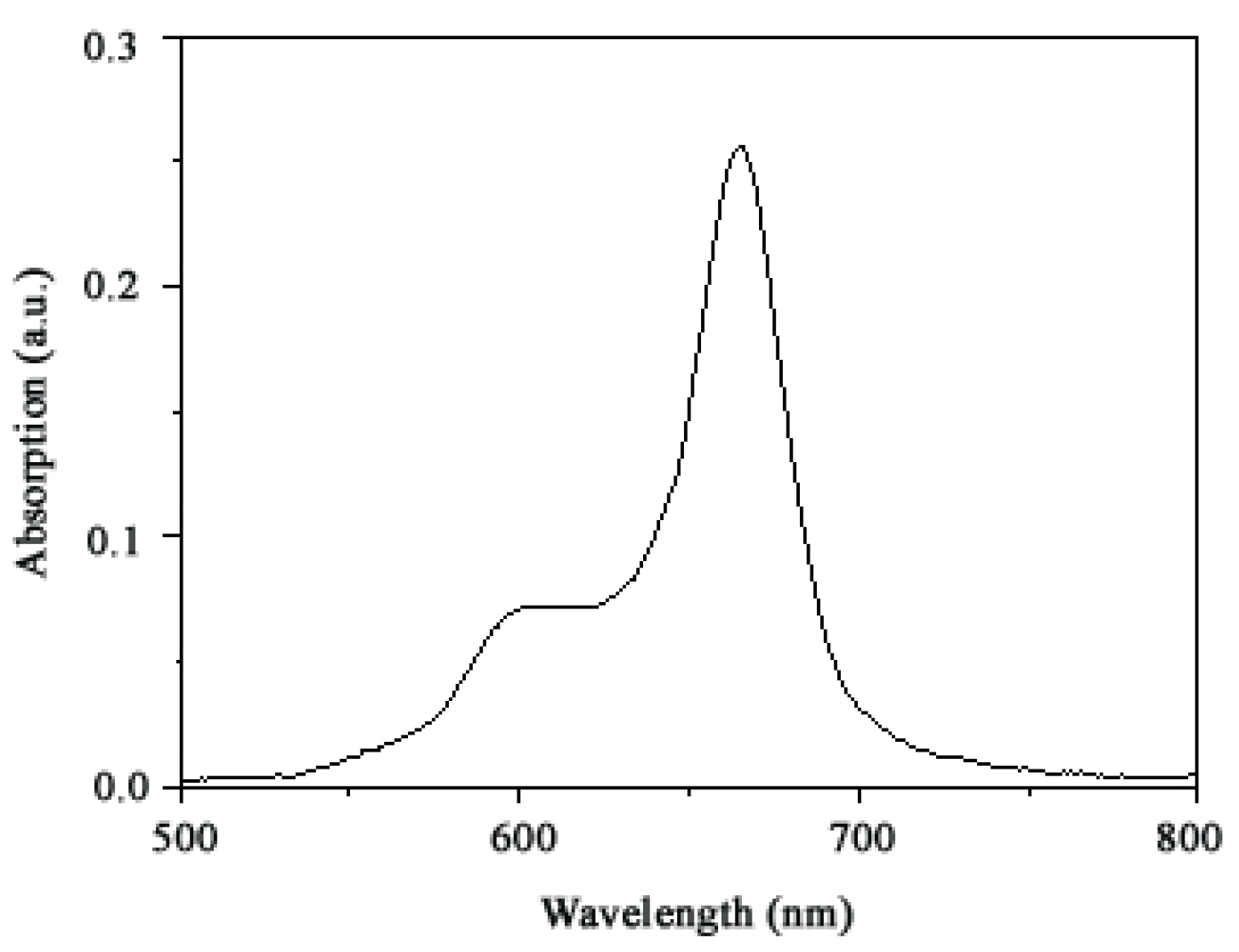
2.1. Ultraviolet-visible absorption spectrum of TαPcZn
2.2. Inhibitory effect of TαPcZn-PDT on the proliferation of Bel-7402 cells and human dermal fibroblasts
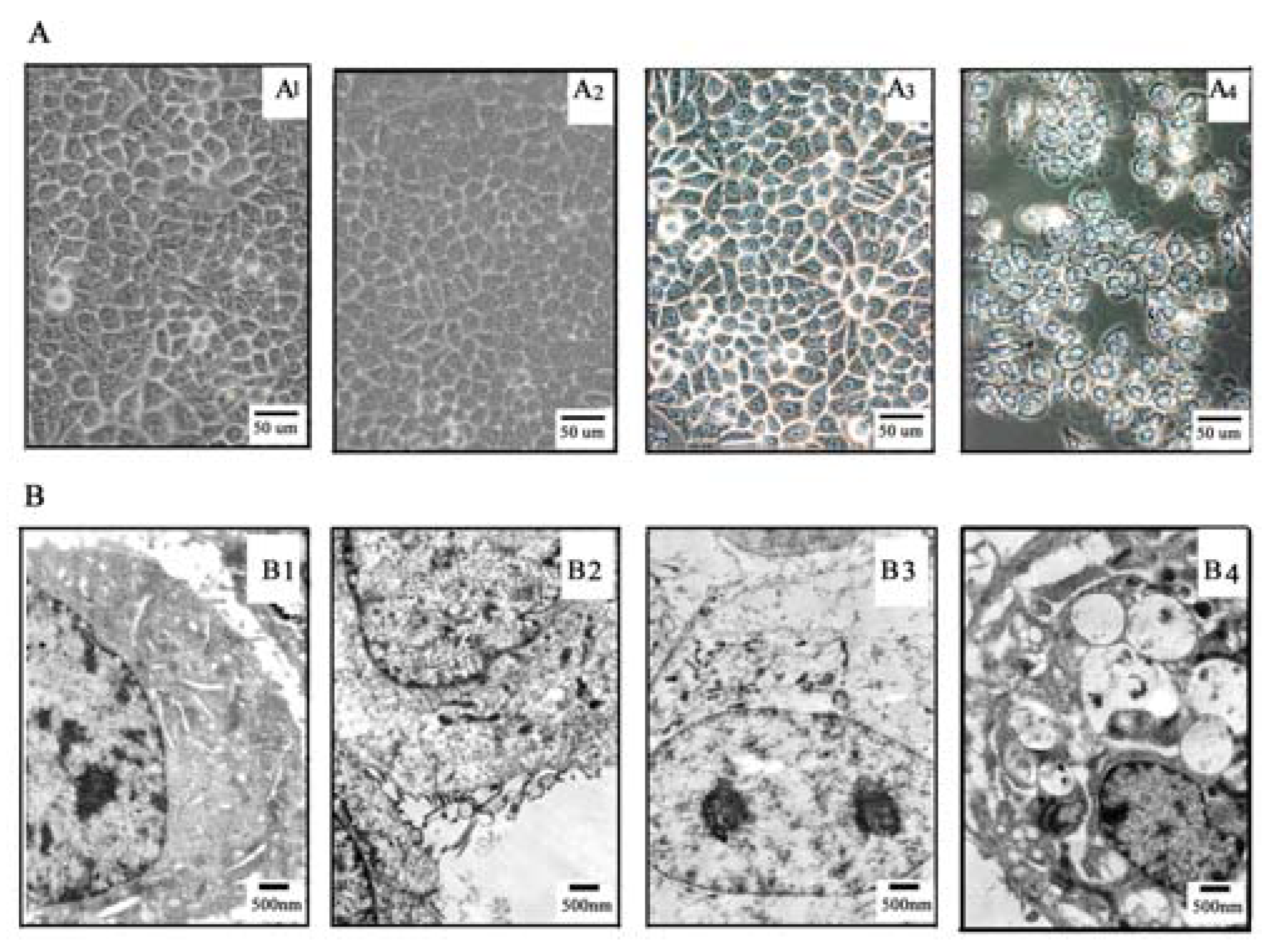
2.3. TαPcZn cellular localization
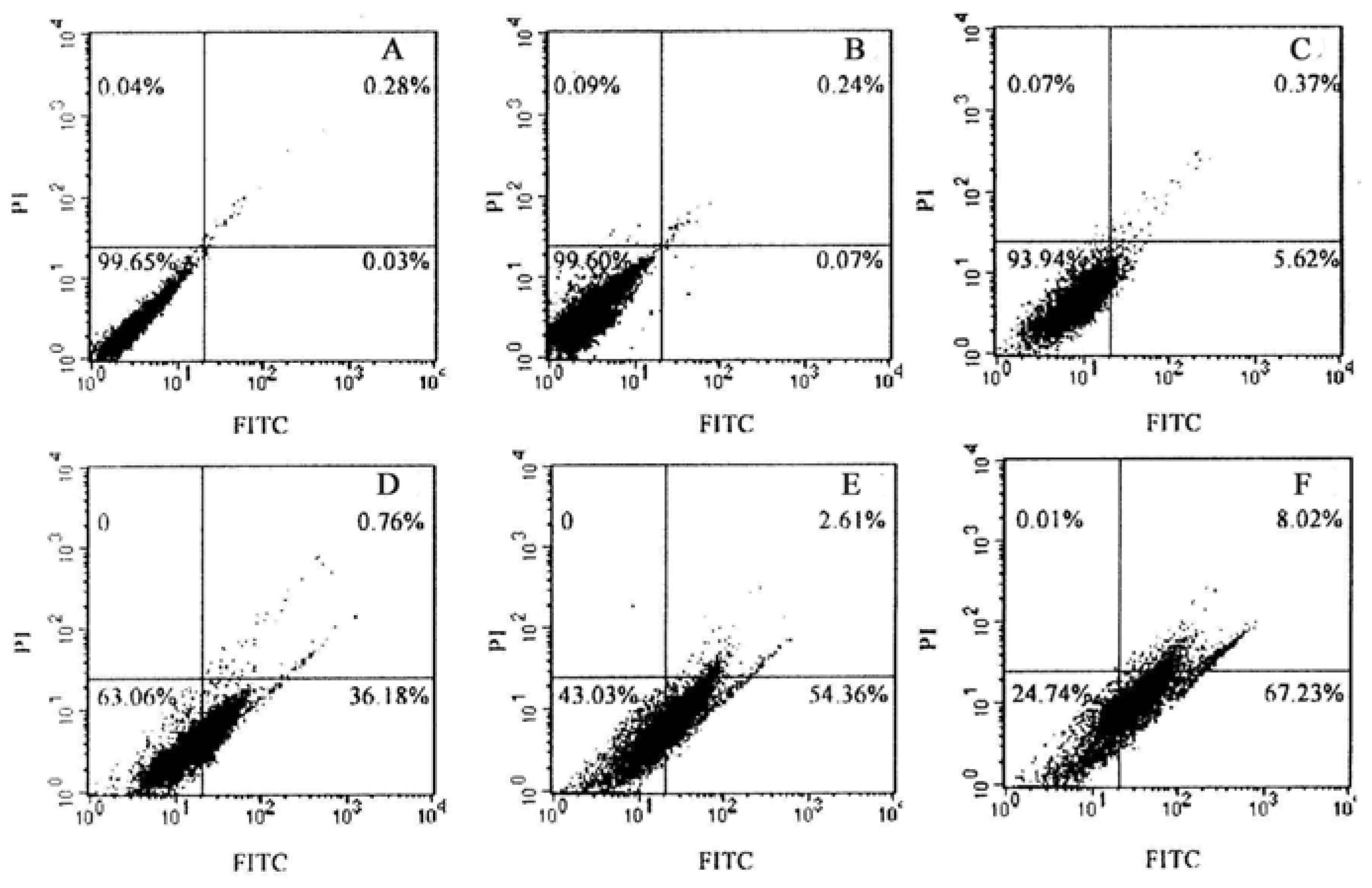
2.4. Effect of TαPcZn-PDT on the apoptosis of Bel-7402 cells
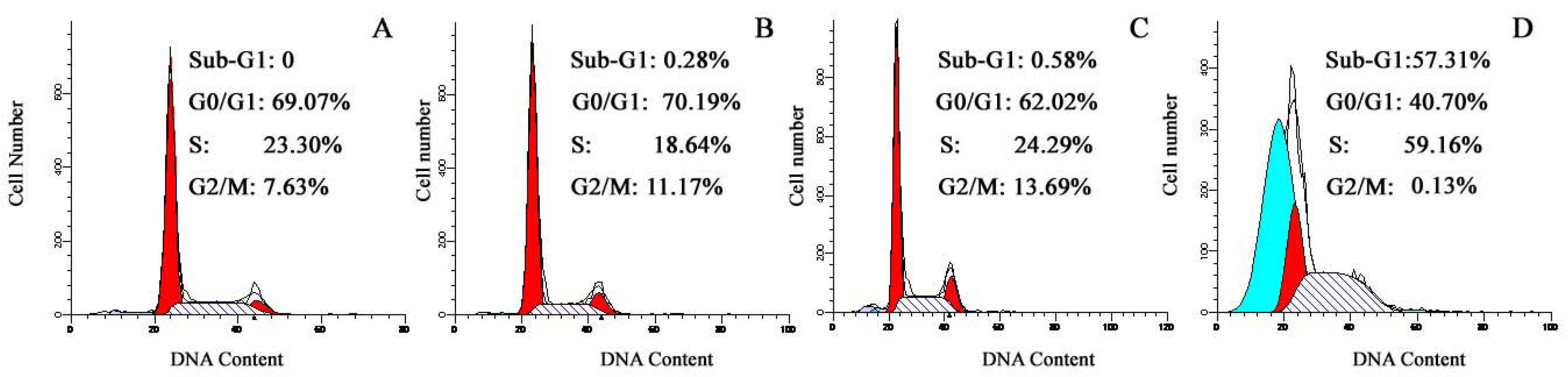
2.5. Effect of TαPcZn-PDT on the cycle of Bel-7402 cells
2.6. Effect of TαPcZn-PDT on Bcl-2 and Fas in TαPcZn-PDT-induced apoptosis of Bel-7402 cells
3. Experimental Section
3.1. Materials
3.2. Ultraviolet-visible absorption spectrum assay
3.3. Cell culture
3.4. TαPcZn-PDT treatment
3.5. Cell viability assay
3.6. Inverted microscope and electron microscopy assay
3.7. Flow cytometry analysis of Annexin V-FITC/PI double stained cells for apoptosis
3.8. DNA flow cytometry analysis for cell cycle and apoptosis
3.9. Immunoblot assay
3.10. Statistical analysis
4. Conclusions
Acknowledgments
References
- Douillard, S.; Olivier, D.; Patrice, T. In vitro and in vivo evaluation of radachlorin(R) sensitizer for photodynamic therapy. Photochem. Photobiol. Sci. 2009, 8, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.H.; Wang, B.Q.; Wang, Y.; Liu, Y.Q.; Xu, G.Y.; Wu, S. Damaging effects of nanosized TiO2 on bel-7402 human liver cancer cell under photoinduce. J. Inorg. Mater. 2006, 21, 1467–1471. [Google Scholar]
- Lim, S.H.; Thivierge, C.; Nowak-Sliwinska, P.; Han, J.; van den Bergh, H.; Wagnières, G.; Burgess, K.; Lee, H.B. In vitro and in vivo photocytotoxicity of boron dipyrromethene derivatives for photodynamic therapy. J. Med. Chem. 2010, 53, 2865–2874. [Google Scholar] [CrossRef] [PubMed]
- Vallinayagam, R.; Weber, J.; Neier, R. Novel bioconjugates of aminolevulinic acid with vitamins. Org. Lett. 2008, 10, 4453–4455. [Google Scholar] [CrossRef] [PubMed]
- Gurba, P.; Vallinayagam, R.; Schmitt, F.; Furrer, J.; Juillerat-Jeanneret, L.; Neier, R. Novel bioconjugates of aminolevulinic acid with nucleosides. Synthesis 2008, 24, 3957–3962. [Google Scholar]
- Longo, J.P.; Lozzi, S.P.; Simioni, A.R.; Morais, P.C.; Tedesco, A.C.; Azevedo, R.B. Photodynamic therapy with aluminum-chloro-phtalocyanine induces necrosis and vascular damage in mice tongue tumors. J. Photochem. Photobiol. B 2009, 94, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Kolarova, H.; Nevrelova, P.; Bajgar, R.; Jirova, D.; Kejlova, K.; Strnad, M. In vitro photodynamic therapy on melanoma cell lines with phthalocyanine. Toxicol. In Vitro 2007, 21, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Lo, P.C.; Leung, S.C.; Chan, E.Y.M.; Fong, W.P.; Ko, W.H.; Ng, D.K. Photodynamic effects of a novel series of silicon(IV) phthalocyanines against human colon adenocarcinoma cells. Photodiagn. Photodyn. Ther. 2007, 4, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Lo, P.C.; Jiang, X.J.; Fong, W.P.; Ng, D.K. Synthesis and in vitro photodynamic activities of di-alpha-substituted zinc(ii) phthalocyanine derivatives. Dalton Trans. 2009, 21, 4129–4135. [Google Scholar] [CrossRef] [PubMed]
- Leung, S.C.; Lo, P.C.; Ng, D.K.; Liu, W.K.; Fung, K.P.; Fong, W.P. Photodynamic activity of BAM-SiPc, an unsymmetrical bisamino silicon (IV) phthalocyanine, in tumour-bearing nude mice. Br. J. Pharmacol. 2008, 154, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Szliszka, E.; Czuba, Z.P.; Domino, M.; Mazur, B.; Zydowicz, G.; Krol, W. Ethanolic extract of propolis (EEP) enhances the apoptosis- inducing potential of TRAIL in cancer cells. Molecules 2009, 14, 738–754. [Google Scholar] [CrossRef] [PubMed]
- Szliszka, E.; Czuba, Z.P.; Mazur, B.; Paradysz, A.; Krol, W. Chalcones and dihydrochalcones augment trail-mediated apoptosis in prostate cancer cells. Molecules 2010, 15, 5336–5353. [Google Scholar] [CrossRef] [PubMed]
- Chiaviello, A.; Paciello, I.; Postiglione, I.; Crescenzi, E.; Palumbo, G. Combination of photodynamic therapy with aspirin in human-derived lung adenocarcinoma cells affects proteasome activity and induces apoptosis. Cell Proliferation 2010, 43, 480–493. [Google Scholar] [CrossRef] [PubMed]
- Otake, E.; Sakuma, S.; Torii, K.; Maeda, A.; Ohi, H.; Yano, S.; Morita, A. Effect and mechanism of a new photodynamic therapy with glycoconjugated fullerene. Photochem. Photobiol. 2010, 86, 1356–1363. [Google Scholar] [CrossRef] [PubMed]
- Awang, K.; Azmi, M.N.; Aun, L.I.; Aziz, A.N.; Ibrahim, H.; Nagoor, N.H. The apoptotic effect of 1’s-1’-acetoxychavicol acetate from alpinia conchigera on human cancer cells. Molecules 2010, 15, 8048–8059. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H. Selenium as an essential micronutrient, roles in cell cycle and apoptosis. Molecules 2009, 14, 1263–1278. [Google Scholar] [CrossRef] [PubMed]
- Svaasand, L.O.; Ellingsen, R. Optical penetration in human intracranial tumors. Photochem. Photobiol. 1985, 41, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Star, W.M. Light dosimetry in vivo. Phys. Med. Biol. 1997, 42, 763–787. [Google Scholar] [CrossRef] [PubMed]
- Gouterman, M. Spectra of porphyrins. J. Mol. Spectrosc. 1961, 6, 138–163. [Google Scholar] [CrossRef]
- Tapajós, E.C.; Longo, J.P.; Simioni, A.R.; Lacava, Z.G.; Santos, M.F.; Morais, P.C.; Tedesco, A.C.; Azevedo, R.B. In vitro photodynamic therapy on human oral keratinocytes using chloroaluminum-phthalocyanine. Oral Oncol. 2008, 44, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Vittar, N.B.; Prucca, C.G.; Strassert, C.; Awruch, J.; Rivarola, V.A. Cellular inactivation and antitumor efficacy of a new zinc phthalocyanine with potential use in photodynamic therapy. Int. J. Biochem. Cell Biol. 2008, 40, 2192–2205. [Google Scholar] [CrossRef] [PubMed]
- Haywood-Small, S.L.; Vernon, D.I.; Griffiths, J.; Schofield, J.; Brown, S.B. Phthalocyanine-mediated photodynamic therapy induces cell death and a G0/G1 cell cycle arrest in cervical cancer cells. Biochem. Biophys. Res. Commun. 2006, 339, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Feyes, D.K.; Agarwal, R.; Mukhtar, H. Photodynamic therapy results in induction of WAF1/CIP1/P21 leading to cell cycle arrest and apoptosis. Proc. Natl. Acad. Sci. 1998, 95, 6977–6982. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Dwarakanath, B.S.; Muralidhar, K.; Koru-Sengul, T.; Jain, V. Non-monotonic changes in clonogenic cell survival induced by disulphonated aluminum phthalocyanine photodynamic treatment in a human glioma cell line. J. Transl. Med. 2010, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Foote, C.S. Definition of type I and type II photosensitized oxidation. Photochem. Photobiol. 1991, 54, 659. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.; Oleinick, N.L.; Nieminen, A.L. Photodynamic therapy-induced apoptosis in epidermoidcarcinoma cells. Reactive oxygen species and mitochondrial inner membrane permeabilization. J. Biol. Chem. 2001, 276, 47379–47386. [Google Scholar] [CrossRef] [PubMed]
- Quiogue, G.; Saggu, S.; Hung, H.I.; Kenney, M.E.; Oleinick, N.L.; Lemasters, J.J.; Nieminen, A.L. Signaling from lysosomes enhances mitochondria-mediated photodynamic therapy in cancer cells. Proc. Soc. Photo. Opt. Instrum. Eng. 2009, 7380, 1–8. [Google Scholar]
- Rello-Varona, S.; Stockert, J.C.; Cañete, M.; Acedo, P.; Villanueva, A. Mitotic catastrophe induced in HeLa cells by photodynamic treatment with Zn(II)-phthalocyanine. Int. J. Oncol. 2008, 32, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Gupta, S.; Feyes, D.K.; Mukhtar, H. Involvement of Fas (APO-1/CD-95) during photodynamic-therapy-mediated apoptosis in human epidermoid carcinoma A431 cells. J. Invest. Dermatol. 2000, 115, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.C.; Lo, P.C.; Ng, D.K.; Ko, W.H.; Leung, S.C.; Fung, K.P.; Fong, W.P. BAM-SiPc, a novel agent for photodynamic therapy, induces apoptosis in human hepatocarcinoma HepG2 cells by a direct mitochondrial action. Cancer Biol. Ther. 2006, 5, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Usuda, J.; Chiu, S.M.; Murphy, E.S.; Lam, M.; Nieminen, A.L.; Oleinick, N.L. Domain-dependent photodamage to Bcl-2. A membrane anchorage region is needed to form the target of phthalocyanine photosensitization. J. Biol. Chem. 2003, 278, 2021–2029. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D.; Baron, E.D.; Scull, H.; Hsia, A.; Berlin, J.C.; McCormick, T.; Colussi, V.; Kenney, M.E.; Cooper, K.D.; Oleinick, N.L. Photodynamic therapy with the phthalocyanine photosensitizer Pc 4: the case experience with preclinical mechanistic and early clinical–translational studies. Toxicol. Appl. Pharmacol. 2007, 224, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Y.; Chen, W.; Li, T.; Wang, Y.; Xia, C.H.; Li, X.L. Study on synthesis and antineoplastic activity of α -tetra-(4-carboxyphenoxy)phthalocyanine zinc. J. Liaoning Norm. Univ. Nat. Sci. 2009, 32, 94–97. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival, application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
Sample Availability: Samples of TαPcZn are available from the authors. |







© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xia, C.; Wang, Y.; Chen, W.; Yu, W.; Wang, B.; Li, T. New Hydrophilic/Lipophilic Tetra-α-(4-carboxyphenoxy) Phthalocyanine Zinc-Mediated Photodynamic Therapy Inhibits the Proliferation of Human Hepatocellular Carcinoma Bel-7402 Cells by Triggering Apoptosis and Arresting Cell Cycle. Molecules 2011, 16, 1389-1401. https://doi.org/10.3390/molecules16021389
Xia C, Wang Y, Chen W, Yu W, Wang B, Li T. New Hydrophilic/Lipophilic Tetra-α-(4-carboxyphenoxy) Phthalocyanine Zinc-Mediated Photodynamic Therapy Inhibits the Proliferation of Human Hepatocellular Carcinoma Bel-7402 Cells by Triggering Apoptosis and Arresting Cell Cycle. Molecules. 2011; 16(2):1389-1401. https://doi.org/10.3390/molecules16021389
Chicago/Turabian StyleXia, Chunhui, Yu Wang, Wei Chen, Wenxue Yu, Baiqi Wang, and Tao Li. 2011. "New Hydrophilic/Lipophilic Tetra-α-(4-carboxyphenoxy) Phthalocyanine Zinc-Mediated Photodynamic Therapy Inhibits the Proliferation of Human Hepatocellular Carcinoma Bel-7402 Cells by Triggering Apoptosis and Arresting Cell Cycle" Molecules 16, no. 2: 1389-1401. https://doi.org/10.3390/molecules16021389