
Pathogenicity and Relative Abundance of Dickeya and Pectobacterium Species in Switzerland: An Epidemiological Dichotomy


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Species Identification
2.2. Inoculation of Tubers for Field Trials
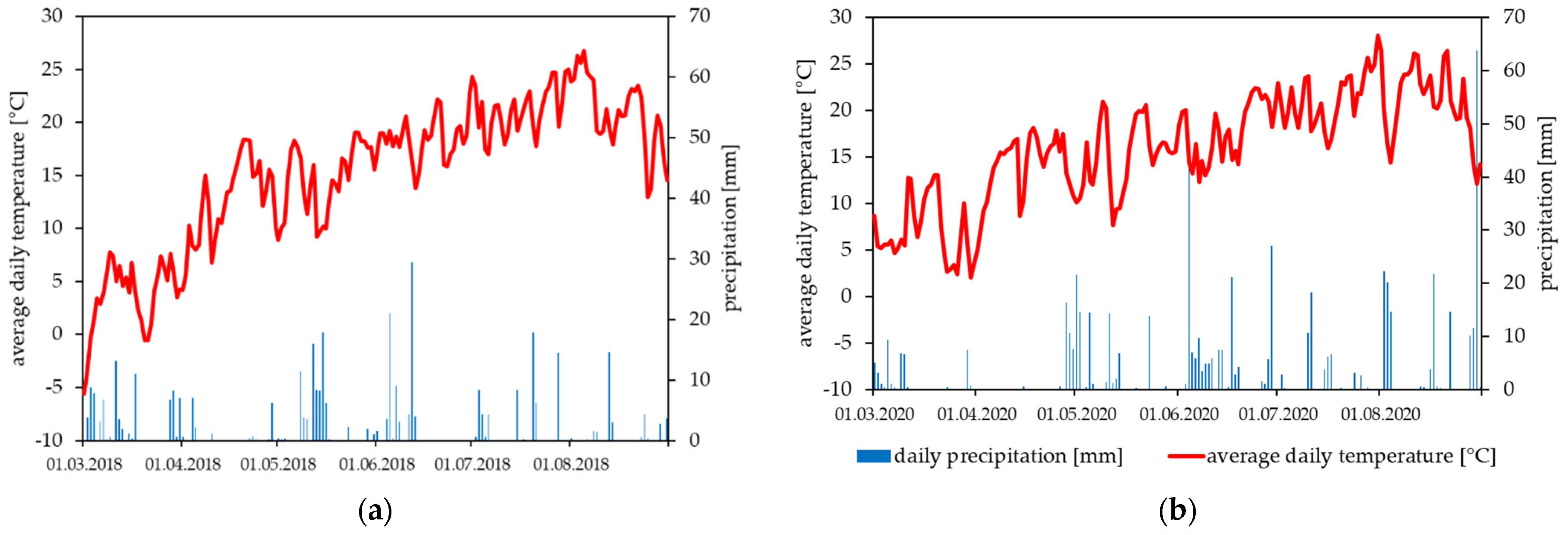
2.3. Field Trials
2.4. Tuber Slice Trial
2.5. Long-Term Strain Survey
2.6. Statistical Analysis
3. Results
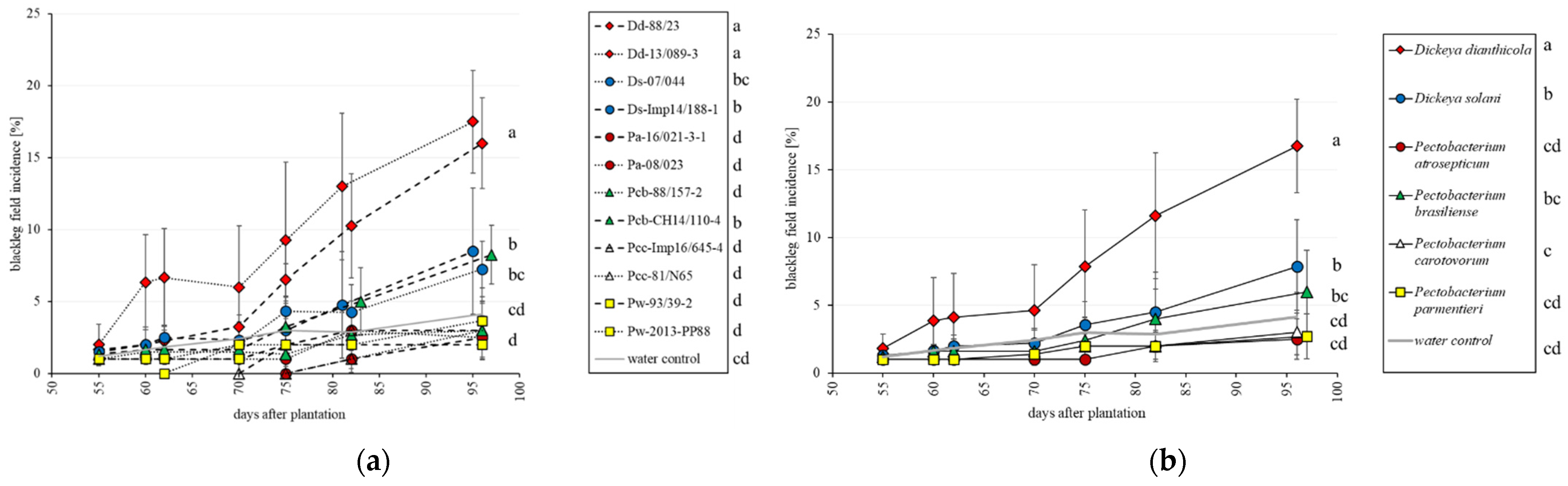
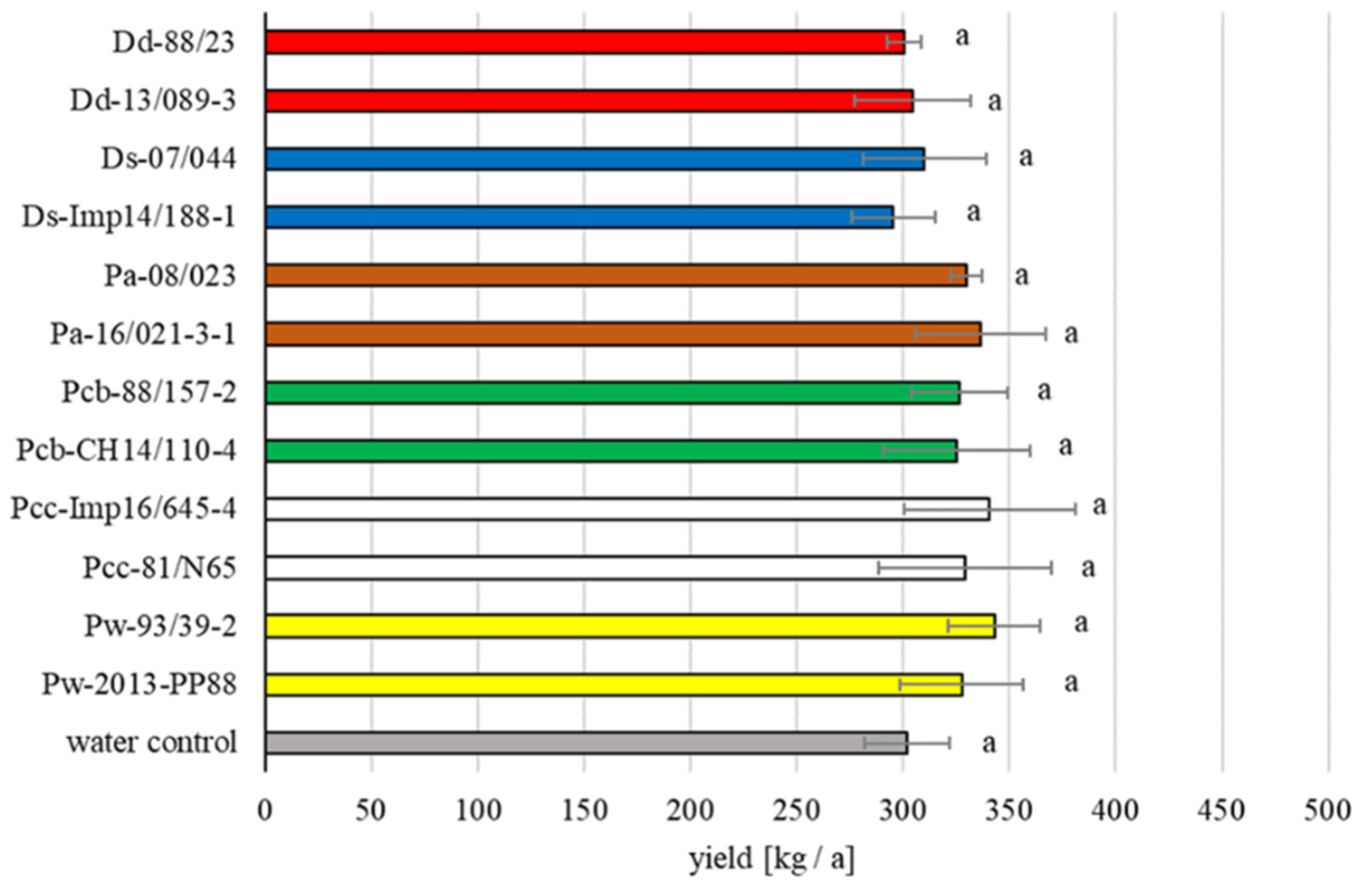
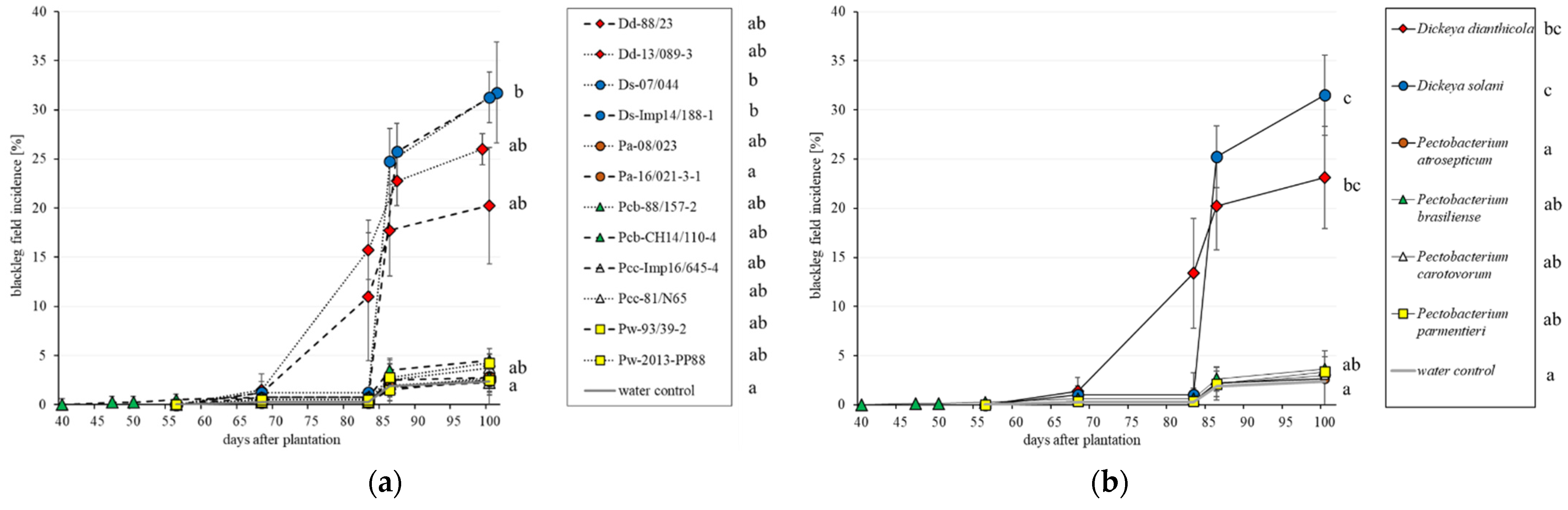
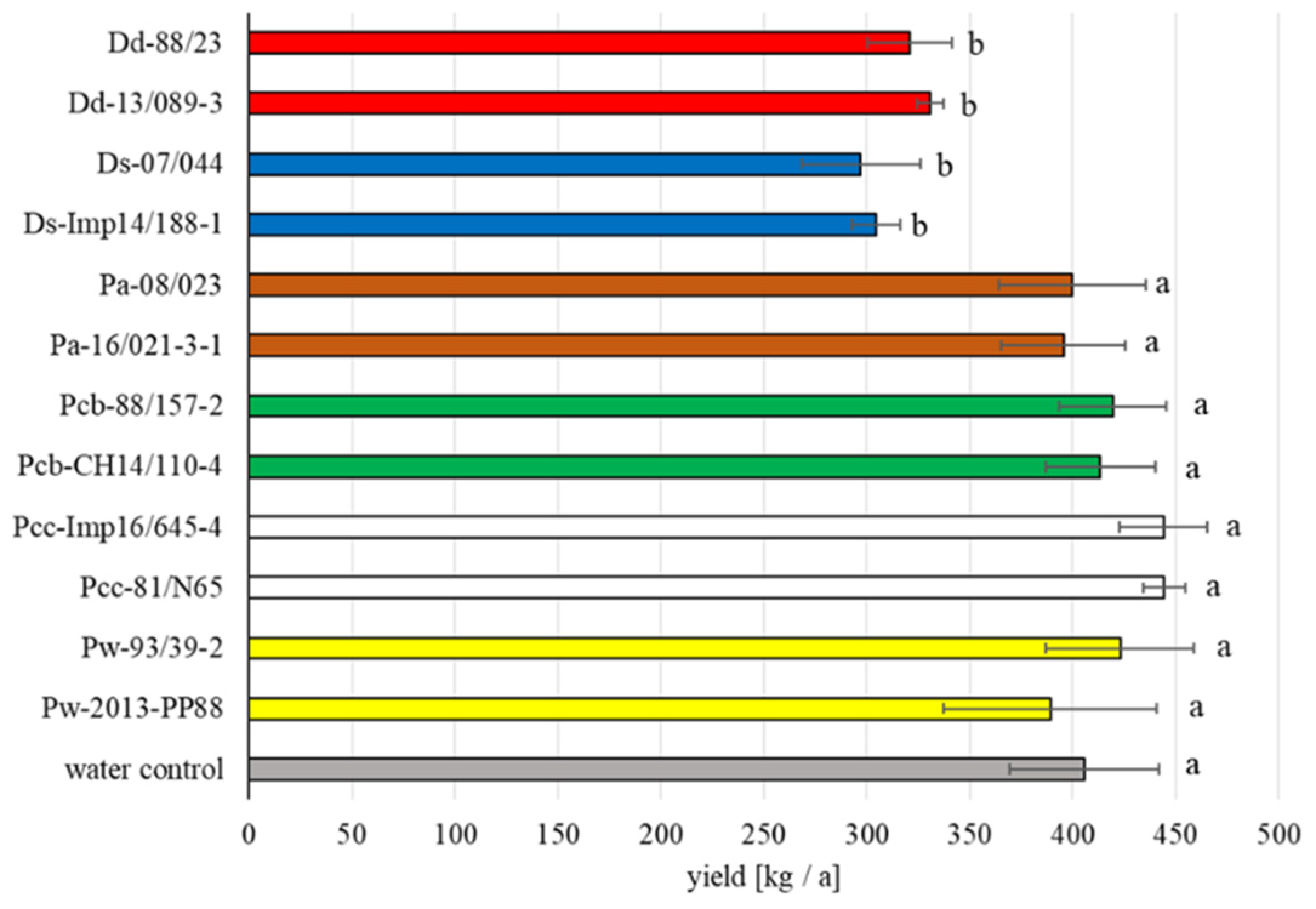
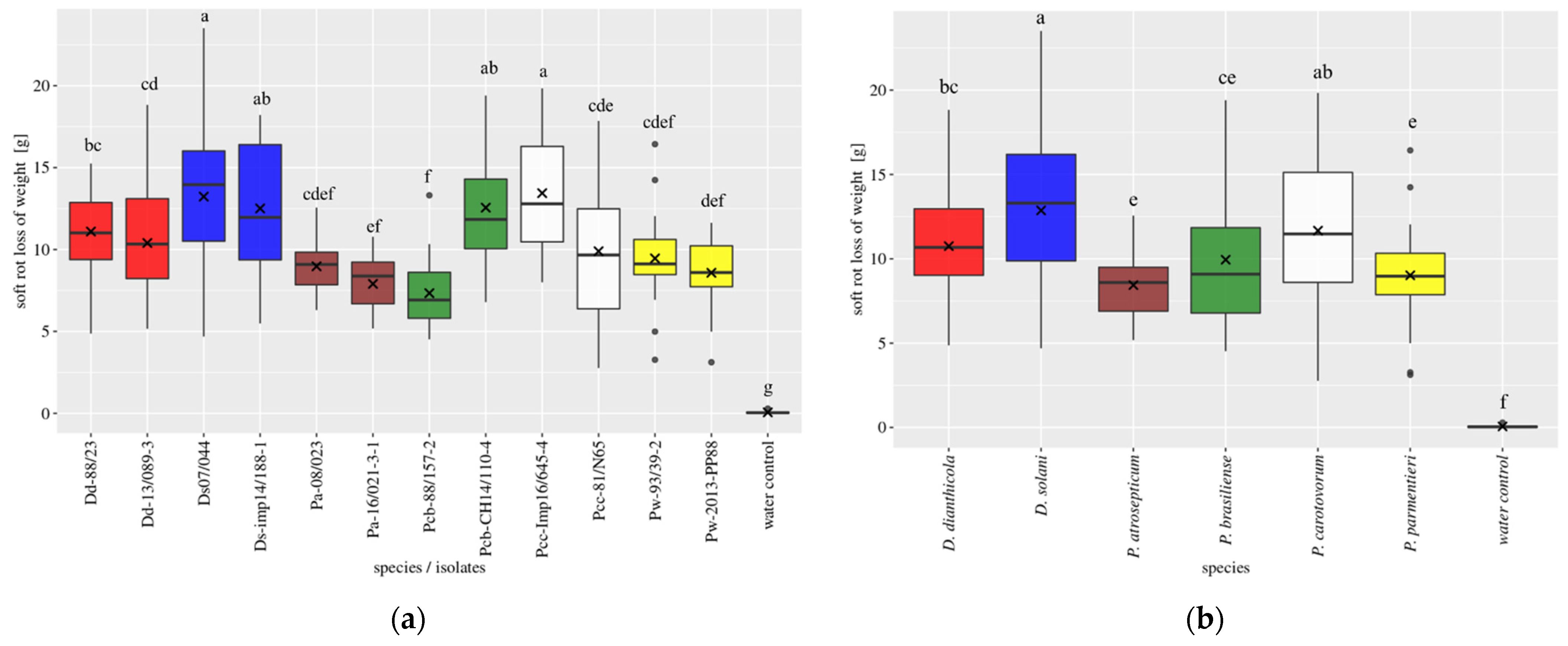
3.1. Field Trials
3.2. Tuber Slice Trial
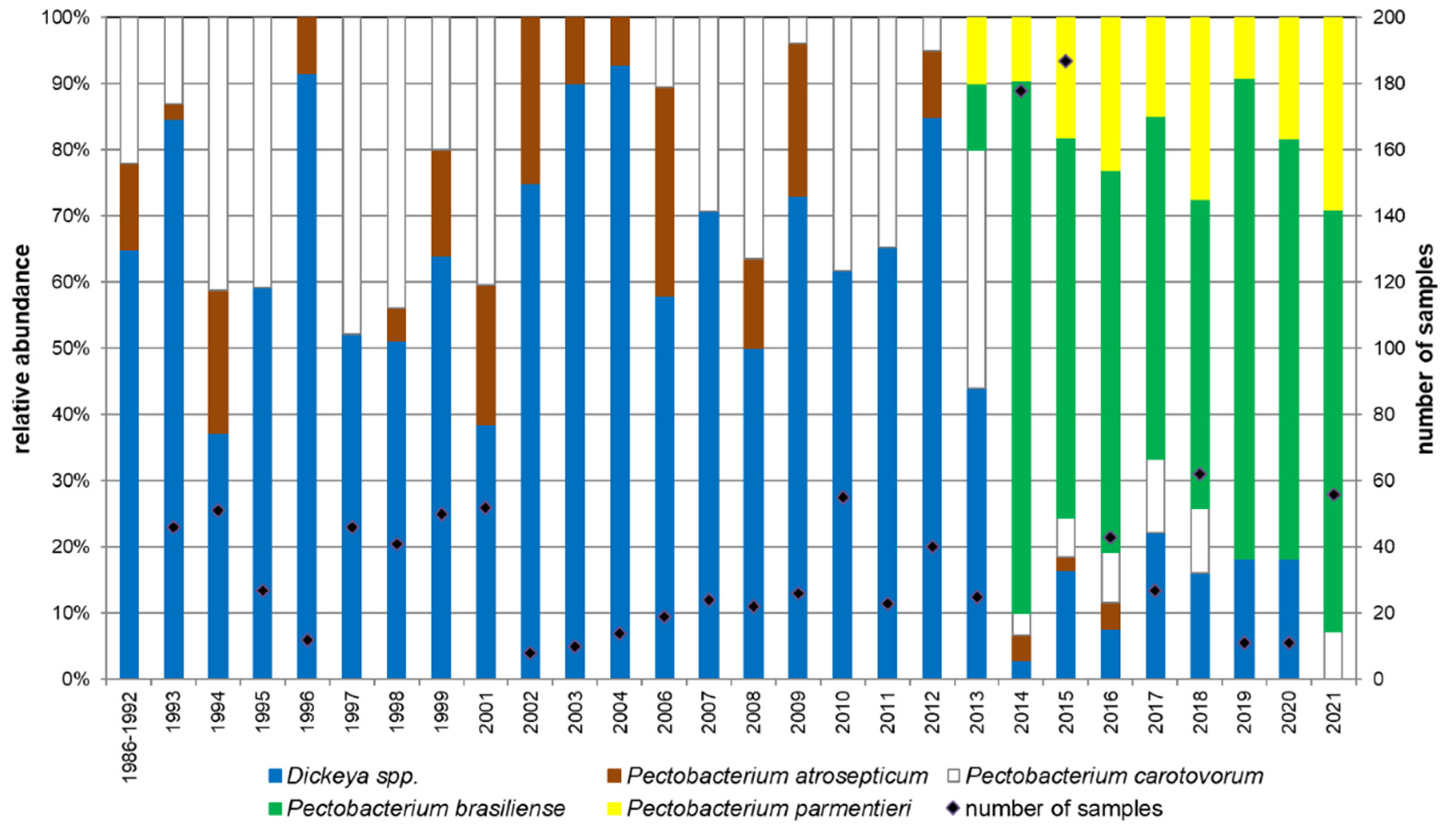
3.3. Strain Survey
4. Discussion
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dupuis, B.; Nkuriyingoma, P.; Van Gijsegem, F. Chapter 8 Economic impact of Pectobacterium and Dickeya species on potato crops: A review and case study. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Van Gijsegem, F., Toth, I., Van der Wolf, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Khayi, S.; Cigna, J.; Chong, T.M.; Quêtu-Laurent, A.; Chan, K.-G.; Hélias, V.; Faure, D. Transfer of the potato plant isolates of Pectobacterium wasabiae to Pectobacterium parmentieri sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5379–5383. [Google Scholar] [CrossRef]
- Van Der Wolf, J.M.; De Haan, E.G.; Kastelein, P.; Krijger, M.; De Haas, B.H.; Velvis, H.; Mendes, O.; Kooman-Gersmann, M.; Van Der Zouwen, P.S. Virulence of Pectobacterium carotovorum subsp. brasiliense on potato compared with that of other Pectobacterium and Dickeya species under climatic conditions prevailing in the Netherlands. Plant Pathol. 2016, 66, 571–583. [Google Scholar] [CrossRef]
- Toth, I.K.; Van Der Wolf, J.M.; Saddler, G.S.; Lojkowska, E.; Helias, V.; Pirhonen, M.; Tsror, L.; Elphinstone, J.G. Dickeya species: An emerging problem for potato production in Europe. Plant Pathol. 2011, 60, 385–399. [Google Scholar] [CrossRef]
- De Werra, P.; Kopp, C.; Häberli, M.; Stöcker, I.; Keil, A.; Debonneville, C.; Oberhänsli, T.; Altenbach, D.; Keiser, A. Monitoring potato seed lots to control blackleg in fields in Switzerland and southern Germany. Plant Pathol. 2020, 69, 1331–1346. [Google Scholar] [CrossRef]
- Zaczek-Moczydłowska, M.A.; Fleming, C.C.; Young, G.K.; Campbell, K.; O’Hanlon, R. Pectobacterium and Dickeya species detected in vegetables in Northern Ireland. Eur. J. Plant Pathol. 2019, 154, 635–647. [Google Scholar] [CrossRef] [Green Version]
- Czajkowski, R.L.; Grabe, G.J.; van der Wolf, J.M. Distribution of Dickeya spp. and Pectobacterium carotovorum subsp carotovorum in naturally infected seed potatoes. Eur. J. Plant Pathol. 2009, 125, 263–275. [Google Scholar] [CrossRef]
- Dupuis, B.; Schaerer, S.; Gilliand, H.; Cazelles, O. The Dickeya and Pectobacterium situation in Switzerland. In Proceedings of the Dickeya Workshop, Emmeloord, The Netherlands, 10–12 November 2010. [Google Scholar]
- Cazelles, O.; Schwaerzel, R. Enquête sur les bactérioses causées par Erwinia dans les cultures de plants de pommes de terre en Suisse romande. Rev. Suisse Agric. 1992, 24, 215–218. [Google Scholar]
- De Werra, P.; Bussereau, F.; Kellenberger, I.; Dupuis, B.; Schaerer, S.; Keiser, A. Pomme de terre: L’Empire Pectobacterium contre-attaque. Rech. Agron. Suisse 2015, 6, 256–263. [Google Scholar]
- Van der Wolf, J.M.; Acuña, I.; De Boer, S.H.; Brurberg, M.B.; Cahill, G.; Charkowski, A.O.; Coutinho, T.; Davey, T.; Dees, M.W.; Degefu, Y.; et al. Diseases Caused by Pectobacterium and Dickeya Species Around the World: Switzerland. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Van Gijsegem, F., Toth, I., Van der Wolf, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 215–261. [Google Scholar] [CrossRef]
- Skelsey, P.; Humphris, S.N.; Campbell, E.J.; Toth, I.K. Threat of establishment of non-indigenous potato blackleg and tuber soft rot pathogens in Great Britain under climate change. PLoS ONE 2018, 13, e0205711. [Google Scholar] [CrossRef]
- Tsror, L.; Erlich, O.; Lebiush, S.; Zig, U.; van de Haar, J.J. Recent outbreak of Erwinia chrysanthemi in Israel—Epidemiology and monitoring in seed tubers. In Proceedings of the 11th International Conference on Plant Pathogenic Bacteria, Edinburgh, Scotland, 10–14 July 2006; p. 70. [Google Scholar]
- Tsror, L.; Erlich, O.; Lebiush, S.; Hazanovsky, M.; Zig, U.; Slawiak, M.; Grabe, G.; Van Der Wolf, J.M.; Van De Haar, J.J. Assessment of recent outbreaks of Dickeya sp. (syn. Erwinia chrysanthemi) slow wilt in potato crops in Israel. Eur. J. Plant Pathol. 2009, 123, 311–320. [Google Scholar] [CrossRef]
- Pérombelon, M.C.M. Potato diseases caused by soft rot erwinias: An overview of pathogenesis. Plant Pathol. 2002, 51, 1–12. [Google Scholar] [CrossRef]
- Van der Wolf, J.; Czajkowski, R.L.; Velvis, H. Effectieve kolonisatie van aardappelplanten door Dickeya-soorten (Erwinia chrysanthemi): Themanummer fytobacteriologie. Gewasbescherming Jaargang 2009, 4, 169–171. [Google Scholar]
- Sacristán, S.; García-Arenal, F. The evolution of virulence and pathogenicity in plant pathogen populations. Mol. Plant Pathol. 2008, 9, 369–384. [Google Scholar] [CrossRef]
- Rouffiange, J.; Gerardin, D.; Riot, G.; Thévoz, E.; Kellenberger, I.; Schaerer, S.; Dupuis, B. Sensibilité au champ de la pomme de terre à la maladie de la jambe noire provoquée par Dickeya spp. Rech. Agron. Suisse 2014, 5, 96–103. [Google Scholar]
- Czajkowski, R.; de Boer, W.J.; van der Zouwen, P.S.; Kastelein, P.; Jafra, S.; de Haan, E.G.; van den Bovenkamp, G.W.; van der Wolf, J.M. Virulence of ‘Dickeya solani’ and Dickeya dianthicola biovar-1 and -7 strains on potato (Solanum tuberosum). Plant Pathol. 2012, 62, 597–610. [Google Scholar] [CrossRef]
- Gill, E.D.; Schaerer, S.; Dupuis, B. Factors impacting blackleg development caused by Dickeya spp. in the field. Eur. J. Plant Pathol. 2014, 140, 317–327. [Google Scholar] [CrossRef]
- Laurila, J.; Hannukkala, A.; Nykyri, J.; Pasanen, M.; Hélias, V.; Garlant, L.; Pirhonen, M. Symptoms and yield reduction caused by Dickeya spp. strains isolated from potato and river water in Finland. Eur. J. Plant Pathol. 2010, 126, 249–262. [Google Scholar] [CrossRef]
- Raoul des Essarts, Y.; Pédron, J.; Blin, P.; Van Dijk, E.; Faure, D.; Van Gijsegem, F. Common and distinctive adaptive traits expressed in Dickeya dianthicola and Dickeya solani pathogens when exploiting potato plant host. Environ. Microbiol. 2019, 21, 1004–1018. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, L.; Humphris, S.; Baeyen, S.; Maes, M.; Van Vaerenbergh, J.; Elphinstone, J.; Saddler, G.; Toth, I. Draft Genome Sequences of Four Dickeya dianthicola and Four Dickeya solani Strains. Genome Announc. 2013, 1, e00087-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khayi, S.; Blin, P.; Pédron, J.; Chong, T.-M.; Chan, K.G.; Moumni, M.; Hélias, V.; Van Gijsegem, F.; Faure, D. Population genomics reveals additive and replacing horizontal gene transfers in the emerging pathogen Dickeya solani. BMC Genom. 2015, 16, 788. [Google Scholar] [CrossRef] [Green Version]
- Pédron, J.; Schaerer, S.; Kellenberger, I.; Van Gijsegem, F. Early Emergence of Dickeya solani Revealed by Analysis of Dickeya Diversity of Potato Blackleg and Soft Rot Causing Pathogens in Switzerland. Microorganisms 2021, 9, 1187. [Google Scholar] [CrossRef] [PubMed]
- Rouffiange, J.; Gerardin, D.; Kellenberger, I.; Schaerer, S.; Dupuis, B. Potato susceptibility to aerial stem rot caused by Dickeya spp. Agrar. Schweiz 2013, 4, 424–431. [Google Scholar]
- Gerardin, D.; Rouffiange, J.; Kellenberger, I.; Schaerer, S.; Dupuis, B. Potato susceptibility to soft rot caused by Dickeya spp. Rech. Agron. Suisse 2013, 4, 288–295. [Google Scholar]
- Laurila, J.; Joutsjoki, T.; Lehtinen, A.; Ahola, V.; Hannukkala, A.; Pirhonen, M. Characterisation of erwinias causing blackleg and soft rot in Finland. Nord. Assoc. Agric. Sci. 2006, 2, 25. [Google Scholar]
- Ge, T.; Ekbataniamiri, F.; Johnson, S.; Larkin, R.; Hao, J. Interaction between Dickeya dianthicola and Pectobacterium parmentieri in Potato Infection under Field Conditions. Microorganisms 2021, 9, 316. [Google Scholar] [CrossRef] [PubMed]
- Nassar, A.; Darrasse, A.; Lemattre, M.; Kotoujansky, A.; Dervin, C.; Vedel, R.; Bertheau, Y. Characterization of Erwinia chrysanthemi by pectinolytic isozyme polymorphism and restriction fragment length polymorphism analysis of PCR-amplified fragments of pel genes. Appl. Environ. Microbiol. 1996, 62, 2228–2235. [Google Scholar] [CrossRef] [Green Version]
- De Boer, S.H.; Ward, L.J. PCR detection of Erwinia carotovora subsp atroseptica associated with potato tissue. Phytopathology 1995, 85, 854–858. [Google Scholar] [CrossRef]
- Duarte, V.; De Boer, S.; Ward, L.; De Oliveira, A.M.R. Characterization of atypical Erwinia carotovora strains causing blackleg of potato in Brazil. J. Appl. Microbiol. 2004, 96, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.W.; Kwon, S.W.; Go, S.J. PCR-based specific and sensitive detection of Pectobacterium carotovorum ssp. carotovorum by primers generated from a URP-PCR fingerprinting-derived polymorphic band. Plant Pathol. 2003, 52, 127–133. [Google Scholar] [CrossRef]
- Kim, M.H.; Cho, M.S.; Kim, B.K.; Choi, H.J.; Hahn, J.H.; Kim, C.; Kang, M.J.; Kim, S.H.; Park, D.S. Quantitative Real-Time Polymerase Chain Reaction Assay for Detection of Pectobacterium wasabiae Using YD Repeat Protein Gene-Based Primers. Plant Dis. 2012, 96, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, L.; Humphris, S.; Saddler, G.S.; Parkinson, N.M.; Bertrand, V.; Elphinstone, J.G.; Toth, I.K. Detection of phytopathogens of the genusDickeyausing a PCR primer prediction pipeline for draft bacterial genome sequences. Plant Pathol. 2012, 62, 587–596. [Google Scholar] [CrossRef]
- Dubois Gill, E.; Helias, V.; Schaerer, S.; Dupuis, B. Efficient inoculation method of potato tubers with Dickeya spp. In Proceedings of the 19th Triennial Conference of the European Association for Potato Research, Brussels, Belgium, 6–11 July 2014. [Google Scholar]
- Haynes, K.G.; Potts, W.J.E.; Goth, R.W. Evaluation of the reliability of determining soft rot resistance in potatoes by the tuber slice method. Am. Potato J. 1997, 74, 265–275. [Google Scholar] [CrossRef]
- Addinsoft. Statistical Software for Excel; XLSTAT Statistical and Data Analysis Solution, Version 2021.2; Addinsoft: Paris, France, 2021. [Google Scholar]
- Pérombelon, M.C.M. Potato blackleg: Epidemiology, host-pathogen interaction and control. Neth. J. Plant Pathol. 1992, 98, 135–146. [Google Scholar] [CrossRef]
- Du Raan, S.; Coutinho, T.A.; van der Waals, J.E. Cardinal temperature differences, determined in vitro, between closely related species and subspecies of pectinolytic bacteria responsible for blackleg and soft rot on potatoes. Eur. J. Plant Pathol. 2016, 144, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Motyka-Pomagruk, A.; Zoledowska, S.; Sledz, W.; Lojkowska, E. The occurrence of bacteria from different species of Pectobacteriaceae on seed potato plantations in Poland. Eur. J. Plant Pathol. 2021, 159, 309–325. [Google Scholar] [CrossRef]
- De Werra, P.; Bussereau, F.; Keiser, A.; Ziegler, D. First Report of Potato Blackleg Caused by Pectobacterium carotovorum subsp. brasiliense in Switzerland. Plant Dis. 2015, 99, 551–552. [Google Scholar] [CrossRef]
- Van der Merwe, J.J.; Coutinho, T.A.; Korsten, L.; van der Waals, J.E. Pectobacterium carotovorum subsp brasiliensis causing blackleg on potatoes in South Africa. Eur. J. Plant Pathol. 2010, 126, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.S.H.; Liew, E.C.Y. Genus-level change in aggressiveness with continuous invasions: A phylogenetically-informed Bayesian quantile regression. Biol. Invasions 2020, 22, 1931–1946. [Google Scholar] [CrossRef]
- Ansermet, M.; Schaerer, S.; Kellenberger, I.; Tallant, M.; Dupuis, B. Influence of seed-borne and soil-carried inocula of Dickeya spp. on potato plant transpiration and symptom expression. Eur. J. Plant Pathol. 2016, 145, 459–467. [Google Scholar] [CrossRef]
- Dupuis, B.; Riot, G.; Kellenberger, I.; Schaerer, S. Effect of lenticels and vascular infection with Dickeya spp. on blackleg symptom expression in potato fields. In Proceedings of the EUPHRESCO Dickeya, Gdansk, Poland, 23 November 2015. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Isolates | Year of Isolation |
|---|---|---|---|
| Dickeya | dianthicola | 13/089-3 | 2013 |
| dianthicola | 88/23 | 1988 | |
| Dickeya | solani | 07/044 | 2007 |
| solani | 14/188-1 | 2014 | |
| Pectobacterium | atrosepticum | 08/023 | 2008 |
| atrosepticum | 16/021-3-1 | 2016 | |
| Pectobacterium | parmentieri | 93/39-2 | 1993 |
| parmentieri | 2013-PP88 | 2013 | |
| Pectobacterium | carotovorum | Imp16/645-4 | 2016 |
| carotovorum | 81/N65 | 1981 | |
| Pectobacterium | brasiliense | 88/157-2 | 1988 |
| brasiliense | CH14/110-4 | 2014 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Werra, P.; Debonneville, C.; Kellenberger, I.; Dupuis, B. Pathogenicity and Relative Abundance of Dickeya and Pectobacterium Species in Switzerland: An Epidemiological Dichotomy. Microorganisms 2021, 9, 2270. https://doi.org/10.3390/microorganisms9112270
de Werra P, Debonneville C, Kellenberger I, Dupuis B. Pathogenicity and Relative Abundance of Dickeya and Pectobacterium Species in Switzerland: An Epidemiological Dichotomy. Microorganisms. 2021; 9(11):2270. https://doi.org/10.3390/microorganisms9112270
Chicago/Turabian Stylede Werra, Patrice, Christophe Debonneville, Isabelle Kellenberger, and Brice Dupuis. 2021. "Pathogenicity and Relative Abundance of Dickeya and Pectobacterium Species in Switzerland: An Epidemiological Dichotomy" Microorganisms 9, no. 11: 2270. https://doi.org/10.3390/microorganisms9112270