
Characterization and Antimicrobial Resistance of Environmental and Clinical Aeromonas Species Isolated from Fresh Water Ornamental Fish and Associated Farming Environment in Sri Lanka

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.1.1. Fish Farming Environment
2.1.2. Apparently Healthy Fish
2.1.3. Diseased Fish
2.2. Isolation of Aeromonads and Phenotypic Characterization
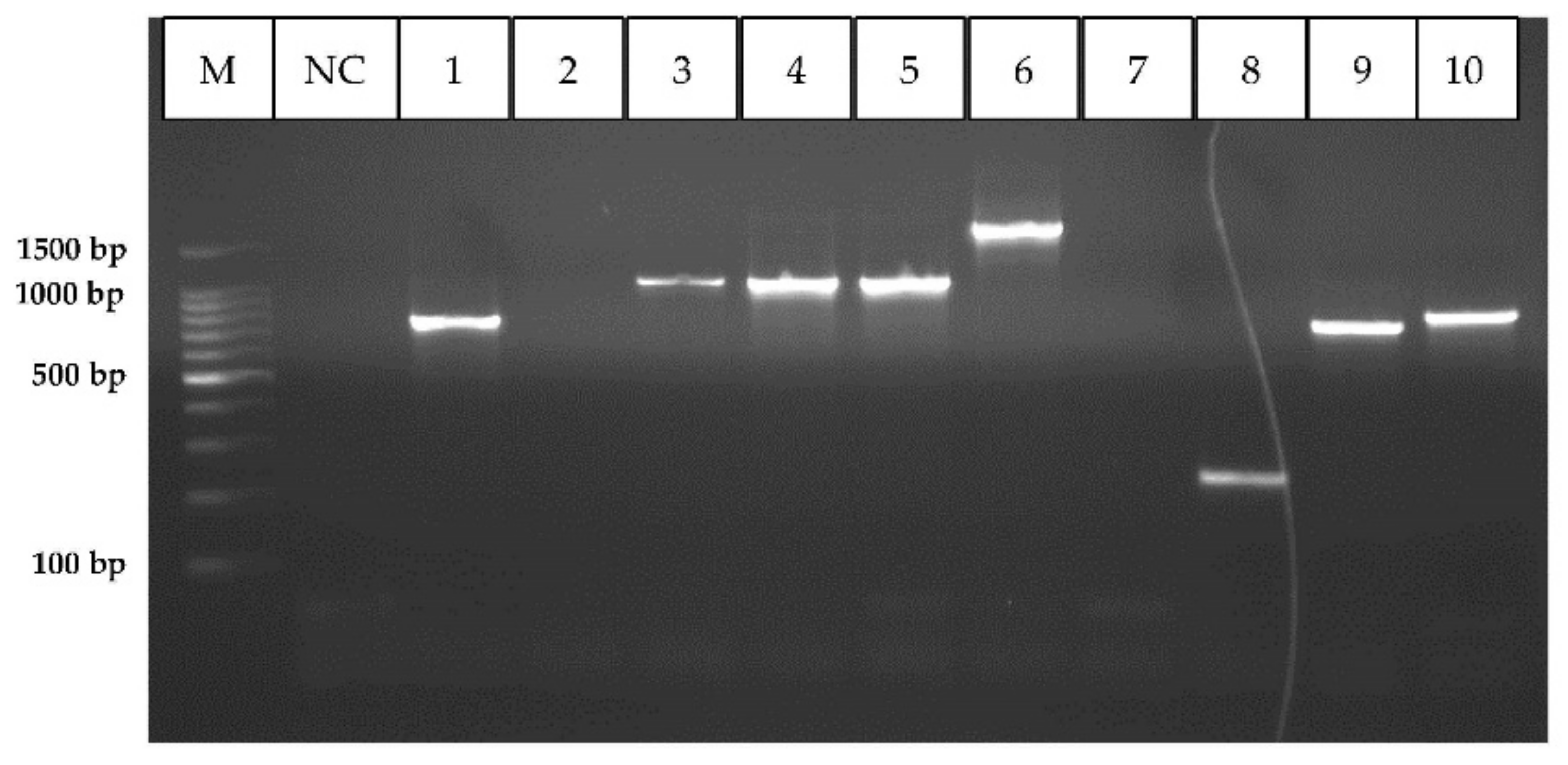
2.3. DNA Extraction, Genetic Characterization and Phylogenetic Analysis
2.3.1. Genus Identification

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Reference |
|---|---|---|
| 16S rRNA forward | 5′ AGAGTTTGATCATGGCTCAG 3′ | [31] |
| 16S rRNA reverse | 5′ GGTTACCTTGTTACGACTT 3′ | |
| gyrB 3F | 5′ TCCGGCGGTCTGCACGGCGT 3′ | [34] |
| gyrB 14R | 5′ TTGTCCGGGTTGTACTCGTC 3′ | |
| hep35 | 5′ TGCGGGTYAARGATBTKGATTT 3′ | [35] |
| hep36 | 5′ CARCACATGCGTRTARAT 3′ | |
| IntI1.F | 5′ GGG TCA AGG ATC TGG ATT TCG 3′ | [36] |
| IntI1.R | 5′ ACA TGC GTG TAA ATC ATC GTC G 3′ | |
| hep58 | 5′ TCATGGCTTGTTATGACTGT 3′ | [35] |
| hep59 | 5′ GTAGGGCTTATTATGCACGC 3′ | |
| hep74 | 5′ CGGGATCCCGGACGGCATGCACGATTTGTA 3′ | [37] |
| hep51 | 5′ GATGCCATCGCAAGTACGAG 3′ |
2.3.2. Species Identification
2.3.3. Nucleotide Sequence Accession Numbers
2.3.4. Phylogenetic Analysis
2.4. Antimicrobial Susceptibility Testing
2.5. PCR Amplification of Integrons and Determination of the Class of Integrons
2.6. Amplification of the Gene Cassettes Regions of Class 1 and Class 2 Integrons
2.7. Statistical Analysis
3. Results
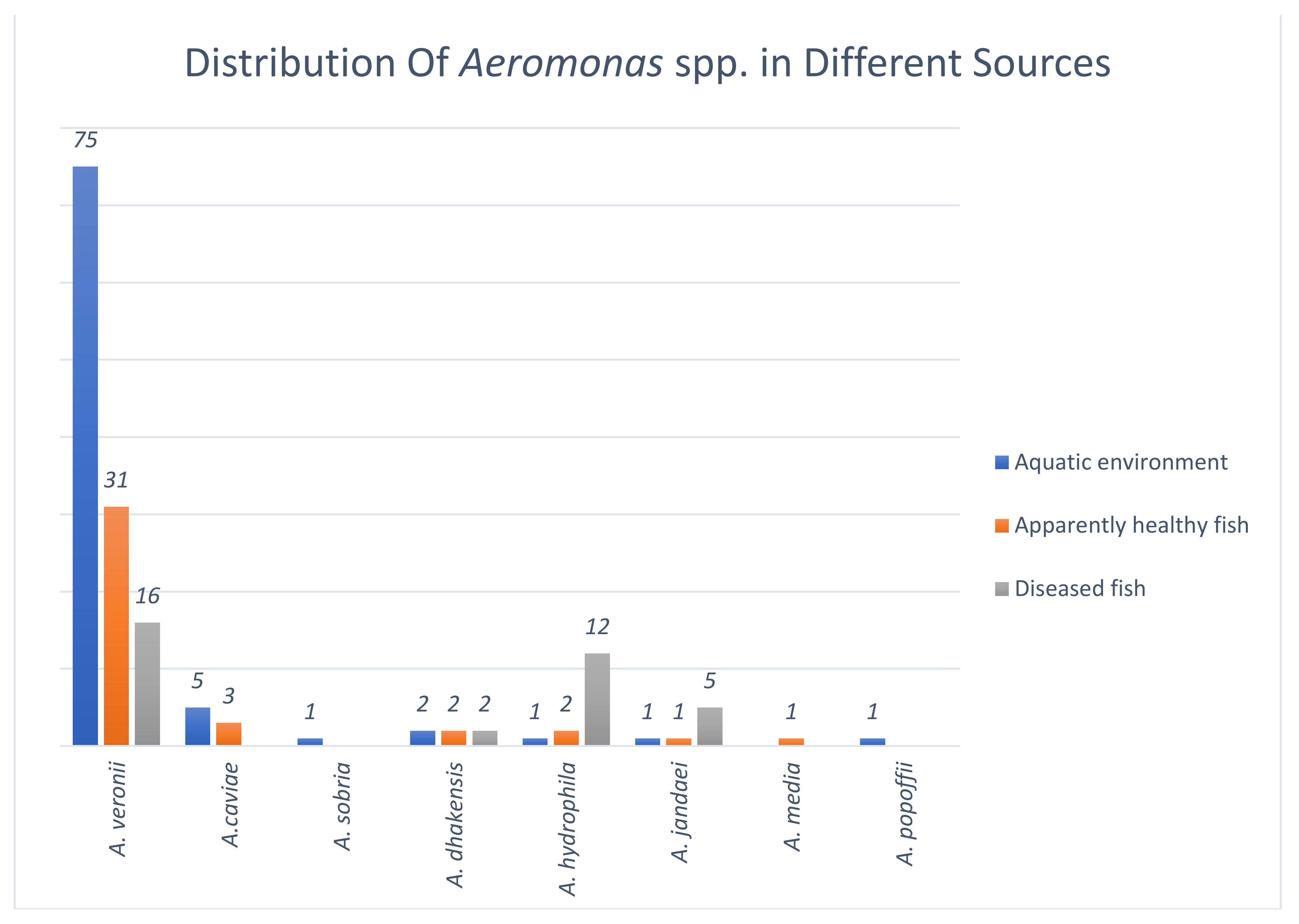
3.1. Identification of Aeromonas spp.
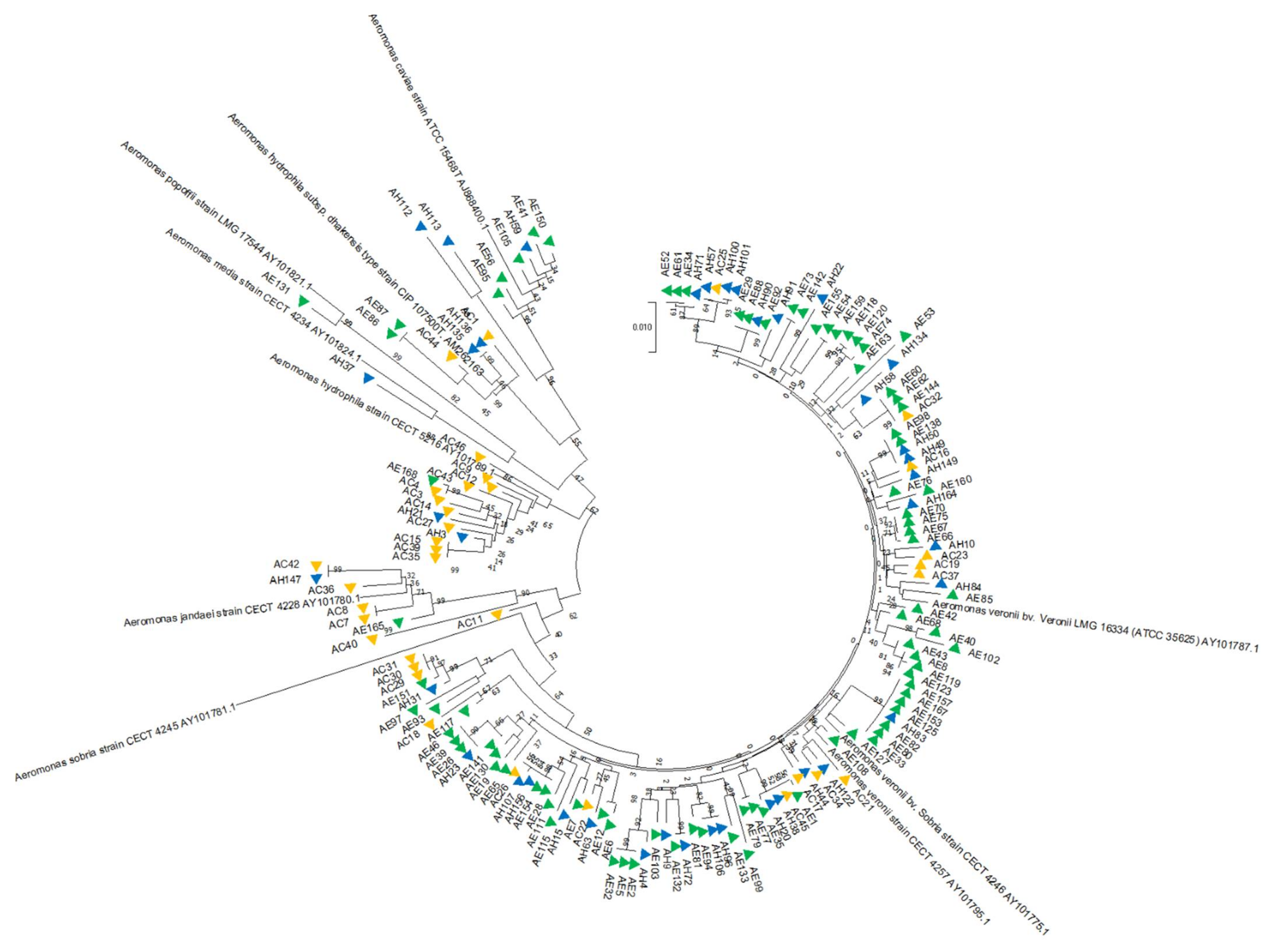
3.1.1. Phylogenetic Analysis
3.1.2. Species Distribution of Aeromonas among Isolates from the Aquatic Environment
3.1.3. Species Distribution of Aeromonas among Isolates from Apparently Healthy Ornamental Fish
3.1.4. Species Distribution of Clinical Aeromonas Isolates from Ornamental Fish
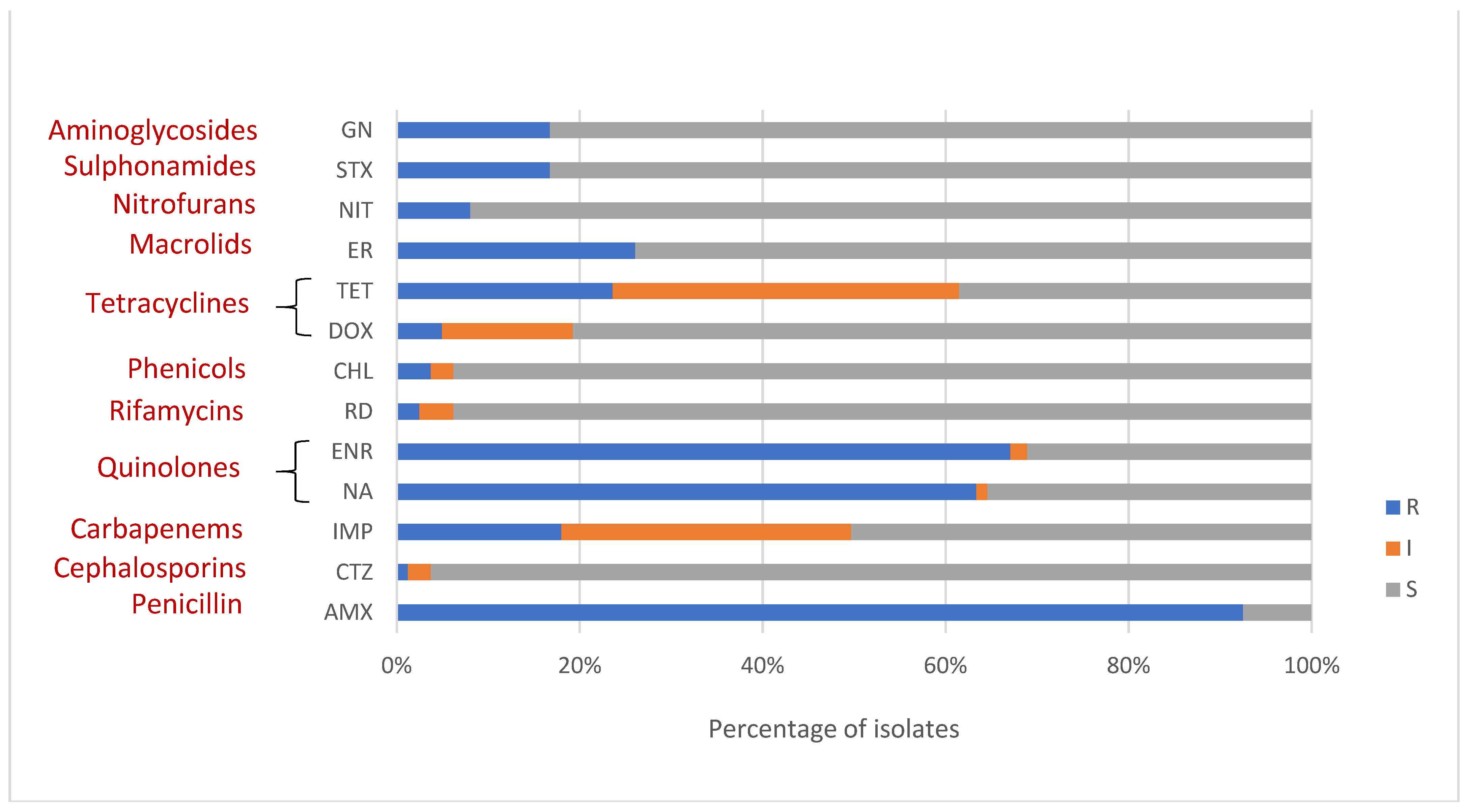
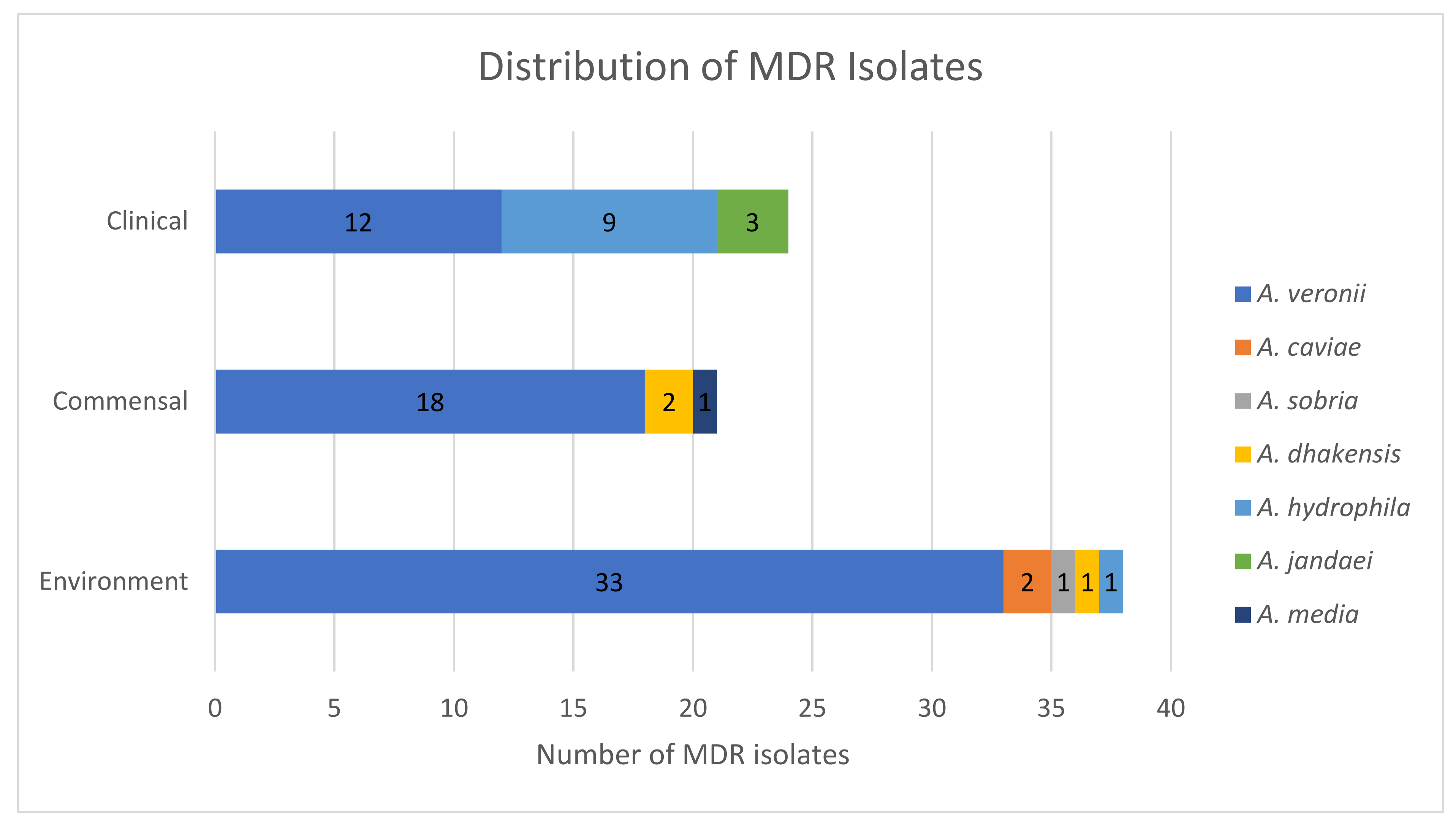
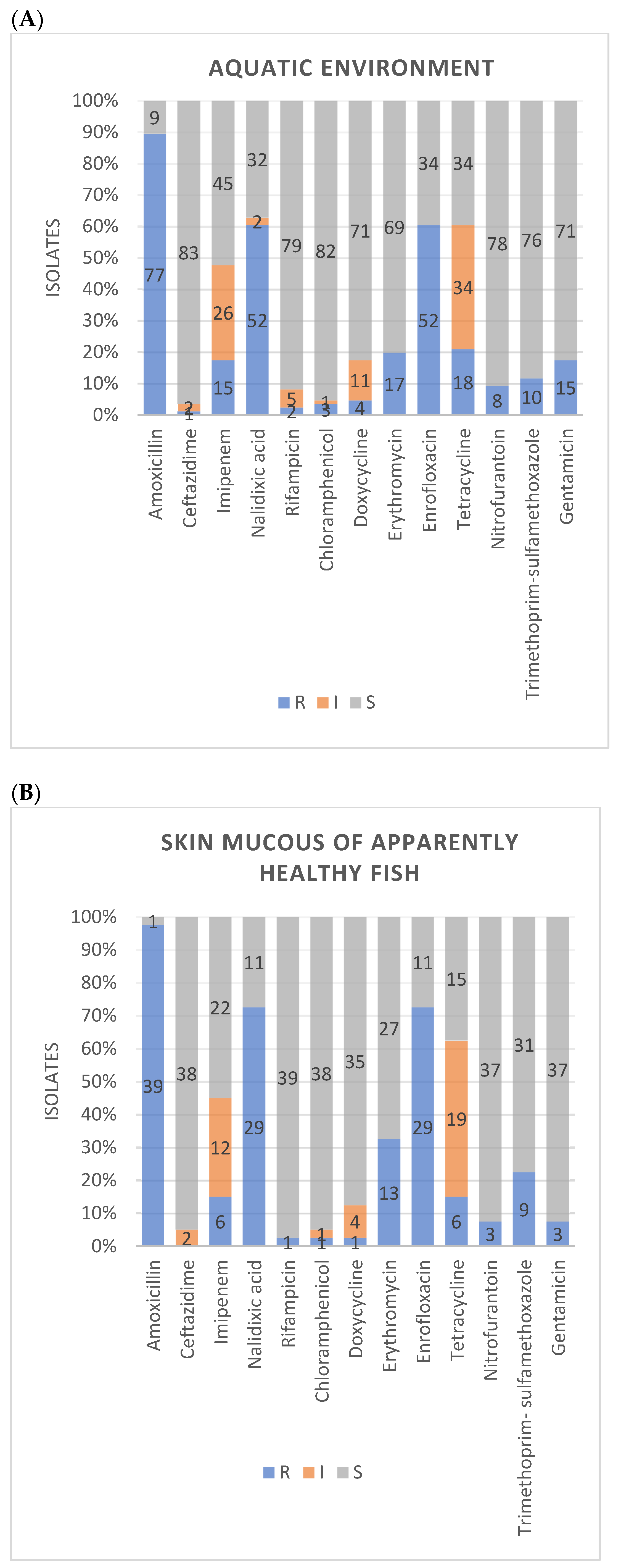
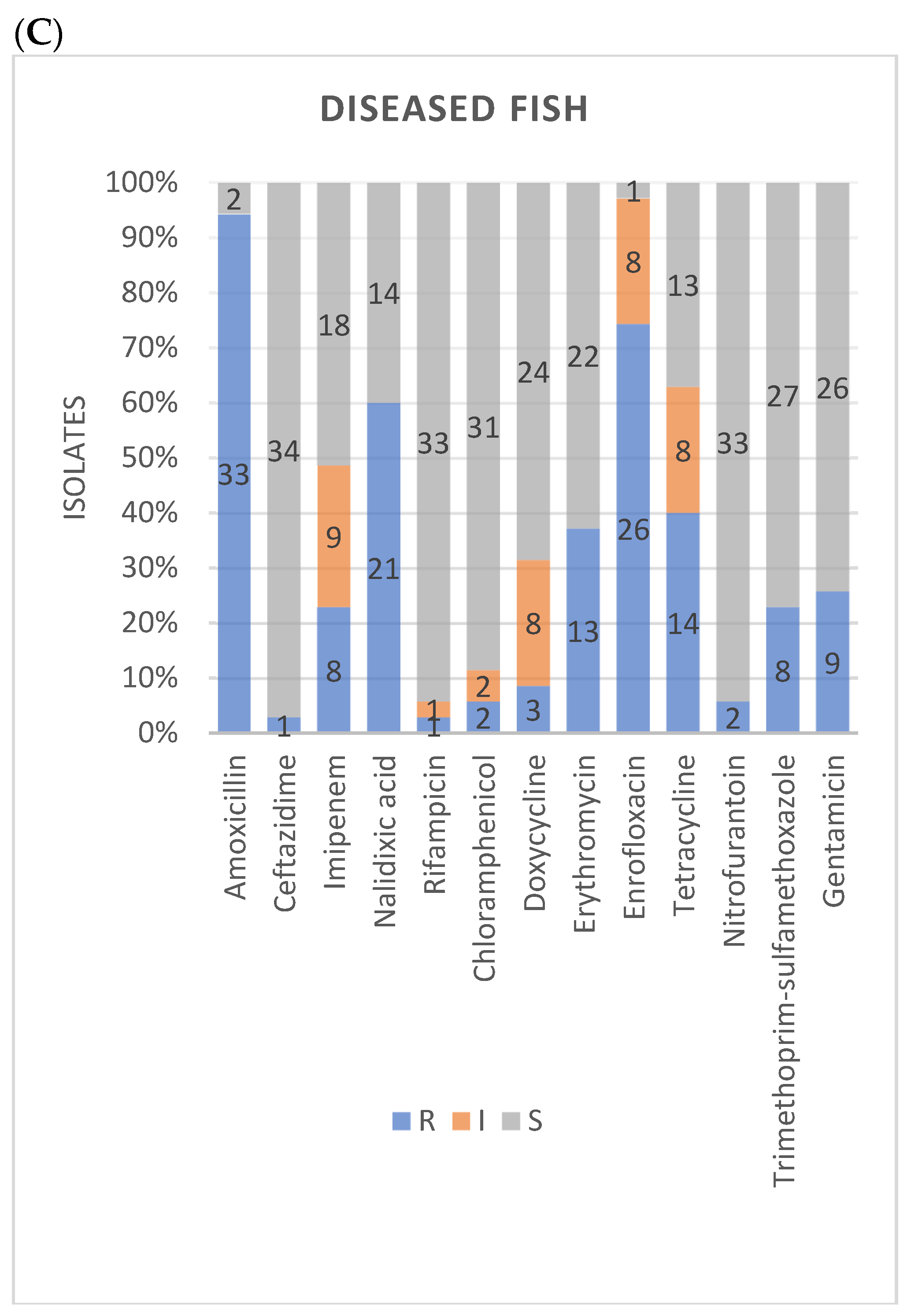
3.2. Antimicrobial Susceptibility
3.3. Detection of Class 1 and Class 2 Integrons
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ndi, O.L.; Barton, M.D. Incidence of Class 1 Integron and Other Antibiotic Resistance Determinants in Aeromonas Spp. from Rainbow Trout Farms in Australia. J. Fish Dis. 2011, 34, 589–599. [Google Scholar] [CrossRef]
- Ghenghesh, K.S.; El-Mohammady, H.; Levin, S.Y.; Zorgani, A.; Tawil, K. Antimicrobial Resistance Profile of Aeromonas Species Isolated from Libya. Libyan J. Med. 2013, 8, 1–3. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. The Genus Aeromonas: Taxonomy, Pathogenicity, and Infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaz-hidalgo, R.; Figueras, M.J. Review Article Aeromonas Spp. Whole Genomes and Virulence Factors Implicated in Fish Disease. J. Fish Dis. 2013, 36, 371–388. [Google Scholar] [CrossRef]
- Syrova, E.; Kohoutova, L.; Dolejska, M.; Papezikova, I.; Kutilova, I.; Cizek, A.; Navratil, S.; Minarova, H.; Palikova, M. Antibiotic Resistance and Virulence Factors in Mesophilic Aeromonas Spp. from Czech Carp Fisheries Eva Syrova. J. Appl. Microbiol. 2018, 125, 1702–1713. [Google Scholar] [CrossRef]
- Hossain, S.; Silva, B.C.J.D.; Wickramanayake, M.V.K.S.; Dahanayake, P.S.; Wimalasena, S.H.M.P.; Heo, G. Incidence of Antimicrobial Resistance Genes and Class 1 Integron Gene Cassettes in Multidrug-Resistant Motile Aeromonas Sp. Isolated from Ornamental Guppy (Poecilia Reticulata). Lett. Appl. Microbiol. 2019, 69, 1–10. [Google Scholar] [CrossRef]
- Saengsitthisak, B.; Chaisri, W.; Punyapornwithaya, V.; Mekteirat, R.; Klayraung, S.; Bernard, J.K.; Pikulkaew, S. Occurrence and Antimicrobial Susceptibility Profiles of Multidrug-Resistant Aeromonads Isolated from Freshwater Ornamental Fish in Chiang Mai Province. Pathogens 2020, 9, 973. [Google Scholar] [CrossRef] [PubMed]
- Hatha, M.; Vivekanandhan, A.A.; Julie Joice, G.; Christol. Antibiotic Resistance Pattern of Motile Aeromonads from Farm Raised Fresh Water Fish. Int. J. Food Microbiol. 2005, 98, 131–134. [Google Scholar] [CrossRef]
- Parker, J.L.; Shaw, J.G. Aeromonas Spp. Clinical Microbiology and Disease. J. Infect. 2011, 62, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Willmon, E.; Burgos, F.A.; Ray, C.L.; Hanson, T.; Arias, C. Biofilm and Sediment Are Major Reservoirs of Virulent Aeromonas Hydrophila (VAh) in Catfish Production Ponds Wenlong Cai. J. Aquat. Anim. Health 2019, 31, 112–120. [Google Scholar] [CrossRef]
- Ormanci, S.; Yucel, N. Bio Film Formation on Polystyrene and Glass Surface by Aeromonas Species Isolated from Different Sources. J. Food Process. Preserv. 2017, 41, e13223. [Google Scholar] [CrossRef]
- Dias, C.; Borges, A.; Saavedra, M.J.; Simoes, M. Biofilm Formation and Multidrug Resistant Aeromonas Spp. from Wild Animals. J. Glob. Antimicrob. Resist. 2018, 12, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Heuer, O.E.; Kruse, H.; Grave, K.; Collignon, P.; Karunasagar, I.; Angulo, F.J. Human Health Consequences of Use of Antimicrobial Agents in Aquaculture. Clin. Infect. Dis. 2009, 49, 1248–1253. [Google Scholar] [CrossRef] [PubMed]
- Manage, P.M. Heavy Use of Antibiotics in Aquaculture: Emerging Human and Animal Health Problems – A Review. Sri Lanka J. Aquat. Sci. 2018, 23, 13. [Google Scholar] [CrossRef]
- Burridge, L.; Weis, J.; Cabello, F.; Pizarro, J. Chemical Use in Salmon Aquaculture: A Review of Current Practices and Possible Environmental Effects. Aquaculture 2010, 306, 7–23. [Google Scholar] [CrossRef]
- Hossain, S.; Heo, G. Ornamental Fish: A Potential Source of Pathogenic and Multidrug-Resistant Motile Aeromonas Spp. Lett. Appl. Microbiol. 2021, 72, 2–12. [Google Scholar] [CrossRef]
- Akinbowale, O.L.; Peng, H.; Barton, M.D. Antimicrobial Resistance in Bacteria Isolated from Aquaculture Sources in Australia. J. Appl. Microbiol. 2006, 100, 1103–1113. [Google Scholar] [CrossRef]
- Miller, R.A.; Harbottle, H. Antimicrobial Drug Resistance in Fish Pathogens. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Chung, T.H.; Yi, S.W.; Shin, G.W. Antibiotic Resistance and Repetitive-Element PCR Fingerprinting in Aeromonas Veronii Isolates. J. Fish Dis. 2017, 40, 821–829. [Google Scholar] [CrossRef]
- Aravena-román, M.; Inglis, T.J.J.; Henderson, B.; Riley, T.V.; Chang, B.J. Antimicrobial Susceptibilities of Aeromonas Strains Isolated from Clinical and Environmental Sources to 26 Antimicrobial Agents. Antimicrob. Agents Chemother. 2012, 56, 1110–1112. [Google Scholar] [CrossRef] [Green Version]
- Dias, C.; Mota, V.; Martinez-Murcia, A.; Saavedra, M.J. Antimicrobial Resistance Patterns of Aeromonas Spp. Isolated from Ornamental Fish. J. Aquac. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Preena, P.G.; Dharmaratnam, A.; Raj, N.S.; Kumar, T.V.A.; Raja, S.A.; Swaminathan, T.R. Antibiotic Susceptibility Pattern of Bacteria Isolated from Freshwater Ornamental Fish, Guppy Showing Bacterial Disease. Biologia (Bratisl.) 2019, 74, 1055–1062. [Google Scholar] [CrossRef]
- Goñi-urriza, M.; Pineau, L.; Capdepuy, M.; Roques, C.; Caumette, P.; Quentin, C. Antimicrobial Resistance of Mesophilic Aeromonas Spp. Isolated from Two European Rivers. J. Antimicrob. Chemother. 2000, 46, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Piotrowska, M.; Popowska, M. Insight into the Mobilome of Aeromonas Strains. Front. Microbiol. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Bao, X.; Ji, L.; Chen, L.; Liu, J.; Miao, J.; Chen, D.; Bian, H.; Li, Y. Resistance Integrons: Class 1, 2 and 3 Integrons. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cambray, G.; Guerout, A.; Mazel, D. Integrons. Annu. Rev. Genet. 2010, 44, 141–166. [Google Scholar] [CrossRef]
- Hall, R.M. Integrons and Gene Cassettes: Hotspots of Diversity in Bacterial Genomes. Ann. N.Y. Acad. Sci. 2012, 1267, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Ling, K.H.; Lim, L.Y. The Status of Ornamental Fish Industry in Singapore. Singapore J. Pri. Ind 2005, 32, 59–69. [Google Scholar]
- Shotts, E.B., Jr.; Vanderwork, V.L.; Campbell, L.M. Occurrence of R Factors Associated with Aeromonas Hydrophila Isolates from Aquarium Fish and Waters. J. Fish. Board Canada 1976, 33, 736–740. [Google Scholar] [CrossRef]
- Cunningham, F.L.; Hanson-Dorr, K.C.; Ford, L.; Middleton, D.R.; Crain, A.; Durst, L.; Ware, C.; Griffin, M.J.; Mischke, C.C.; Wan, X.; et al. Environmental Factor (s) and Animal Vector (s) Associated with Atypical Aeromonas Hydrophila Abundance and Dissemination among Channel Catfish Ponds. J. World Aquac. Soc. 2020, 51, 750–762. [Google Scholar] [CrossRef]
- Borrell, N.; Acinas, S.G.; Figueras, M.; Marti, A.J. Identification of Aeromonas Clinical Isolates by Restriction Fragment Length Polymorphism of PCR-Amplified 16S RRNA Genes. J. Clin. Microbiol. 1997, 35, 1671–1674. [Google Scholar] [CrossRef] [Green Version]
- Jagoda, S.S.S.D.S.; Tan, E.; Arulkanthan, A.; Kinoshita, S.; Watabe, S.; Asakawa, S. Draft Genome Sequence of Aeromonas Hydrophila Strain Ae34, Isolated from a Septicemic and Moribund Koi Carp (Cyprinus Carpio Koi), a Freshwater Aquarium Fish. Genome Announc. 2014, 2, e00572-14. [Google Scholar] [CrossRef] [Green Version]
- Jagoda, S.S.S.D.S.; Honein, K.; Arulkanthan, A.; Ushio, H.; Asakawa, S. Genome Sequencing and Annotation of Aeromonas Veronii Strain Ae52, a Multidrug-Resistant Isolate from Septicaemic Gold Fish (Carassius Auratus) in Sri Lanka. Genomics Data 2017, 11, 46–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanez, M.A.; Catalan, V.; Apraiz, D.; Figueras, M.J.; Martinez-murcia, A.J. Phylogenetic Analysis of Members of the Genus Aeromonas Based on GyrB Gene Sequences. Int. J. Syst. Evol. Microbiol. 2003, 53, 875–883. [Google Scholar] [CrossRef] [Green Version]
- White, P.A.; Mciver, C.J.; Deng, Y.; Rawlinson, W.D. Characterisation of Two New Gene Cassettes, AadA5 and DfrA17. FEMS Microbiol. Lett. 2000, 182, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Mazel, D.; Dychinco, B.; Webb, V.A.; Davies, J. Antibiotic Resistance in the ECOR Collection: Integrons and Identification of a Novel Aad Gene. Antibiot. Agents Chemother. 2000, 44, 1568–1574. [Google Scholar] [CrossRef] [Green Version]
- White, P.A.; Iver, C.J.M.C.; Rawlinson, W.D. Integrons and Gene Cassettes in the Enterobacteriaceae. Antimicrob. Agents Chemother. 2001, 45, 2658–2661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Tamura, K. Estimation of the Number of Nucleotide Substitutions When There Are Strong Transition-Transversion and G+C-Content Biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing for Bacteria Isolated from Aquatic Animals, 3rd ed.; CLSI supplement VET04; Clinical and Laboratoty standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Clinical and Laboratory Standards Institute. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, 3rd ed.; CLSI guideline M45; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Clinical and Laboratory Standards Institute. M100. Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; Clinical and Laboratoty standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Jacobs, L.; Chenia, H.Y. Characterization of Integrons and Tetracycline Resistance Determinants in Aeromonas Spp. Isolated from South African Aquaculture Systems. Int. J. Food Microbiol. 2007, 114, 295–306. [Google Scholar] [CrossRef]
- Fernandez-Bravo, A.; Figueras, M.J. An Update on the Genus Aeromonas: Taxonomy, Epidemiology, and Pathogenicity. Microorganisms 2020, 8, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harnisz, M.; Korzeniewska, E. Science of the Total Environment The Prevalence of Multidrug-Resistant Aeromonas Spp. in the Municipal Wastewater System and Their Dissemination in the Environment. Sci. Total Environ. 2018, 626, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Baron, S.; Granier, S.A.; Larvor, E.; Jouy, E.; Cineux, M.; Wilhelm, A.; Gassilloud, B.; Bouquin, S.L.; Kempf, I. Aeromonas Diversity and Antimicrobial Susceptibility in Freshwater — An Attempt to Set Generic Epidemiological Cut-Off Values. Front. Microbiol. 2017, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usui, M.; Tagaki, C.; Fukuda, A.; Okubo, T.; Boonla, C.; Suzuki, S.; Seki, K.; Takada, H.; Tamura, Y. Use of Aeromonas Spp. as General Indicators of Antimicrobial Susceptibility among Bacteria in Aquatic Environments in Thailand. Front. Microbiol. 2016, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.S.; Bruun, M.S.; Dalsgaard, I.; Pedersen, K.; Larsen, J.L. Occurrence of Antimicrobial Resistance in Fish-Pathogenic and Environmental Bacteria Associated with Four Danish Rainbow Trout Farms. Appl. Environ. Microbiol. 2000, 66, 4908–4915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaz-Hidalgo, R.; Alperi, A.; Bujan, N.; Romalde, J.L.; Figueras, M.J. Comparison of Phenotypical and Genetic Identification of Aeromonas Strains Isolated from Diseased Fish. Syst. Appl. Microbiol. 2010, 33, 149–153. [Google Scholar] [CrossRef]
- Martinez-Murcia, A.J.; Monera, A.; Saavedra, M.J.; Oncina, R.; Lopez-Alvarez, M.; Lara, E.; Figueras, M.J. Multilocus Phylogenetic Analysis of the Genus Aeromonas. Syst. Appl. Microbiol. 2011, 34, 189–199. [Google Scholar] [CrossRef]
- Otero-Olarra, J.E.; Curiel-Quesada, E.; Baltazar-Cruz, J.; Aguilera-Arreola, M.G.; Perez-Valdespino, A. Low Cassette Variability in Class 2 and Class 1 Integrons of Aeromonas Spp. Isolated from Environmental Samples. Microb. Drug Rsistance 2020, 26, 794–801. [Google Scholar] [CrossRef]
- Jagoda, S.S.S.D.S.; Wijewardana, T.G.; Arulkanthan, A.; Igarashi, Y.; Tan, E.; Kinoshita, S.; Watabe, S.; Asakawa, S. Characterization and Antimicrobial Susceptibility of Motile Aeromonads Isolated from Freshwater Ornamental Fish Showing Signs of Septicaemia. Dis. Aquat. Organ. 2014, 109, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Wang, N.; Pan, Z.H.; Lu, C.P.; Liu, Y.J. Identity and Virulence Properties of Aeromonas Isolates from Diseased Fish, Healthy Controls and Water Environment in China. Lett. Appl. Microbiol. 2012, 55, 224–233. [Google Scholar] [CrossRef]
- Sicuro, B.; Pastorino, P.; Barisone, S.; Dellerba, D.; Menconi, V.; Righetti, M.; De Vita, V.; Prearo, M. Prevalence and Antibiotic Sensitivity of Bacteria Isolated from Imported Ornamental Fish in Italy: A Translocation of Resistant Strains ? Prev. Vet. Med. 2020, 175, 104880. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.S.; Bruun, M.S.; Dalsgaard, I.; Larsen, J.L. Incidence, Distribution, and Spread of Tetracycline Resistance Determinants and Integron-Associated Antibiotic Resistance Genes among Motile Aeromonads from a Fish Farming Environment. Appl. Environ. Microbiol. 2001, 67, 5675–5682. [Google Scholar] [CrossRef] [Green Version]
- Pessoa, R.B.G.; de Oliveira, W.F.; Marques, D.S.C.; dos Santos Correia, M.T.; de Carvalho, E.V.M.M.; Coelho, L.C.B.B. The Genus Aeromonas: A General Approach. Microb. Pathog. 2019, 130, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Roberts, H.E. Fundamentlas of Ornamental Fish Health; Wiley-Blackwell: Ames, IA, USA, 2010. [Google Scholar]
- Hernández-Montañez, Z.; López-Ramírez, M.P.; Delgado-Balbuena, L.; Dendooven, L.; Bello-López, J.M. Mesophilic Strains of Aeromonas Spp. Can Acquire the Multidrug Resistance Plasmid PRAS1 in Horizontal Transfer Experiments at Low Temperatures. Ann. Microbiol. 2015, 65, 827–831. [Google Scholar] [CrossRef]
- Santos, L.; Ramos, F. Antimicrobial Resistance in Aquaculture: Current Knowledge and Alternatives to Tackle the Problem. Int. J. Antimicrob. Agents 2018, 52, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.T.; Lubbers, B.V.; Schwarz, S.; Watts, J.L. Applying Definitions for Multidrug Resistance, Extensive Drug Resistance and Pandrug Resistance to Clinically Significant Livestock and Companion Animal Bacterial Pathogens. J. Antimicrob. Chemother. 2018, 73, 1460–1463. [Google Scholar] [CrossRef]
- Hossain, S.; De Silva, B.C.J.; Wimalasena, S.H.M.P.; Pathirana, H.N.K.S.; Dahanayake, P.S.; Heo, G.-J. Distribution of Antimicrobial Resistance Genes and Class 1 Integron Gene Cassette Arrays in Motile Aeromonas Spp. Isolated from Goldfish (Carassius Auratus). Microb. drug Rsistance 2018, 24, 1217–1225. [Google Scholar] [CrossRef]
- John, N.; Hatha, A.A.M. Prevalence, Distribution and Drug Resistance of Motile Aeromonads in Freshwater Ornamental Fishes. Indian J. Fish 2012, 59, 161–164. [Google Scholar]
- Nguyen, H.N.K.; Van, T.T.H.; Nguyen, H.T.; Smooker, P.M.; Shimeta, J.; Coloe, P.J. Molecular Characterization of Antibiotic Resistance in Pseudomonas and Aeromonas Isolates from Catfish of the Mekong Delta, Vietnam. Vet. Microbiol. 2014, 171, 397–405. [Google Scholar] [CrossRef]
- Sreedharan, K.; Philip, R.; Singhe, I.S.B. Virulence Potential and Antibiotic Susceptibility Pattern of Motile Aeromonads Associated with Freshwater Ornamental Fish Culture Systems: A Possible Threat to Public Health. Brazilian J. Microbiol. 2012, 43, 754–765. [Google Scholar] [CrossRef] [Green Version]
- Picão, R.C.; Cardoso, J.P.; Campana, E.H.; Nicoletti, A.G.; Petrolini, F.V.B.; Assis, D.M.; Juliano, L.; Gales, A.C. The Route of Antimicrobial Resistance from the Hospital Effluent to the Environment: Focus on the Occurrence of KPC-Producing Aeromonas Spp. and Enterobacteriaceae in Sewage. Diagn. Microbiol. Infect. Dis. 2013, 76, 80–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajdács, M.; Abrok, M.; Lazar, A.; Janvari, L.; Toth, A.; Terhes, G.; Burian, K. Detection of VIM, NDM and OXA-48 Producing Carbapenem Resistant Enterobacterales among Clinical Isolates in Southern Hungary. Acta Microbiol. Immunol. Hung. 2020, 67, 209–215. [Google Scholar] [CrossRef]
- Rosso, F.; Cedano, J.A.; Parra-lara, L.G.; Sanz, A.M.; Velez, J.F.; Hormaza, M.P.; Moncada, P.A.; Correa, A. Emerging Carbapenem-Resistant Aeromonas Spp. Infections in Cali, Colombia. Brazilian J. Infect. Dis. 2019, 23, 336–342. [Google Scholar] [CrossRef]
- Hilt, E.E.; Fitzwater, S.P.; Ward, K.; Maurice, A.D.S.; Chandrasekaran, S.; Garner, O.B.; Yang, S. Carbapenem Resistant Aeromonas Hydrophila Carrying Bla CphA 7 Isolated From Two Solid Organ Transplant Patients. Front. Cell. Infect. Microbiol. 2020, 10, 624. [Google Scholar] [CrossRef]
- Rose, S.; Hill, R.; Bermudez, L.E.; Miller-Morgan, T. Imported Ornamental Fish Are Colonized with Antibiotic-Resistant Bacteria. J. Fish Dis. 2013, 36, 533–542. [Google Scholar] [CrossRef]
- Krumperman, P.H. Multiple Antibiotic Resistance Indexing of Escherichia Coli to Identify High-Risk Sources of Fecal Contamination of Foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Sadat, A.; El-Sherbiny, H.; Zakaria, A.; Ramadan, H.; Awad, A. Prevalence, Antibiogram and Virulence Characterization of Vibrio Isolates from Fish and Shellfish in Egypt: A Possible Zoonotic Hazard to Humans. J. Appl. Microbiol. 2021, 131, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Igbinosa, I.H.; Beshiru, A.; Odjadjare, E.E.; Ateba, C.N.; Igbinosa, E.O. Microbial Pathogenesis Pathogenic Potentials of Aeromonas Species Isolated from Aquaculture and Abattoir Environments. Microb. Pathog. 2017, 107, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Gillings, M.; Boucher, Y.; Labbate, M.; Holmes, A.; Krishnan, S.; Holley, M.; Stokes, H.W. The Evolution of Class 1 Integrons and the Rise of Antibiotic Resistance. J. Bacteriol. 2008, 190, 5095–5100. [Google Scholar] [CrossRef] [Green Version]
- Ranjbar, R.; Salighehzadeh, R.; Sharifiyazdi, H. Antimicrobial Resistance and Incidence of Integrons in Aeromonas Species Isolated from Diseased Freshwater Animals and Water Samples in Iran. Antibiotics 2019, 8, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]







| A. veronii | A. caviae | A. sobria | A. dhakensis | A. hydrophila | A. jandaei | A. media | A. popoffii | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Source | ||||||||||
| Aquatic environment | ||||||||||
| Effluent water | 23 | 1 | 1 | 1 | ||||||
| Pond sediment | 36 | 3 | 1 | |||||||
| Biofilm | 16 | 2 | 1 | 1 | ||||||
| Apparently healthy ornamental fish | ||||||||||
| Poecilia reticulata (Guppy) | 6 | 2 | 2 | |||||||
| Carassius auratus (Goldfish) | 11 | 1 | 1 | |||||||
| Xiphophorus helleri (Swordtail) | 4 | 1 | ||||||||
| Xiphophorus maculatus (Platy) | 8 | 1 | ||||||||
| Poecilia sphenops (Molly) | 2 | 1 | ||||||||
| Diseased fish | 16 | 2 | 12 | 5 | ||||||
| Amoxicillin | Nalidixic acid | Rifampicin | Chloramphenicol | Doxycycline | Erythromycin | Enrofloxacin | Tetracycline | Nitrofurantoin | Trimethoprim-sulfamethoxazole | Gentamicin | Ceftazidime | Imipenem | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | ||
| A. veronii | R | 111 (91.0) | 80 (65.6) | 1 (0.8) | 4 (3.3) | 5 (4.1) | 26 (21.3) | 78 (63.9) | 28 (23.0) | 10 (8.2) | 18 (14.8) | 21 (17.2) | 2 (1.6) | 26 (21.3) |
| I | 2 (1.6) | 5 (4.1) | 3 (2.5) | 17 (13.9) | 52 (42.6) | 3 (2.5) | 40 (32.8) | |||||||
| S | 11 (9.0) | 40 (32.8) | 116 (95.1) | 115 (94.3) | 100 (82.0) | 96 (78.7) | 44 (36.1) | 42 (34.4) | 112 (91.8) | 104 (85.2) | 101 (82.8) | 117 (95.9) | 56 (45.9) | |
| A. caviae | R | 8 (100) | 4 (50.0) | 3 (37.5) | 6 (75) | 1 (12.5) | ||||||||
| I | 2 (25) | 1 (12.5) | ||||||||||||
| S | 4 (50.0) | 8 (100) | 8 (100) | 6 (75) | 5 (62.5) | 2 (25) | 6 (75) | 8 (100) | 8 (100) | 8 (100) | 8 (100) | 8 (100) | ||
| A. sobria | R | 1 (100) | 1 (100) | 1 (100) | 1 (100) | |||||||||
| I | 1 (100) | 1 (100) | ||||||||||||
| S | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | |||||||
| A. dhakensis | R | 6 (100) | 4 (66.7) | 2 (33.3) | 1 (16.7) | 1 (16.7) | 2 (33.3) | 6 (100) | 2 (33.3) | 2 (33.3) | 2 (33.3) | |||
| I | 1 (16.7) | 1 (16.7) | 1 (16.7) | 1 (16.7) | ||||||||||
| S | 2 (33.3) | 4 (66.7) | 5 (83.3) | 5 (83.3) | 4 (66.7) | 3 (50.0) | 4 (66.7) | 4 (66.7) | 5 (83.3) | 5 (83.3) | 5 (83.3) | |||
| A. hydrophila | R | 15 (100) | 7 (46.7) | 1 (6.7) | 2 (13.3) | 8 (53.3) | 11 (73.3) | 5 (33.3) | 1 (6.7) | 3 (20.0) | 2 (13.3) | 2 (13.3) | ||
| I | 1 (6.7) | 3 (20.0) | 2 (13.3) | 5 (33.3) | 2 (13.3) | |||||||||
| S | 8 (53.3) | 13 (86.6) | 15 (100) | 10 (66.7) | 7 (46.7) | 2 (13.3) | 5 (33.3) | 14 (93.3) | 12 (60.0) | 13 (86.6) | 15 (100) | 11 (73.3) | ||
| A. jandaei | R | 6 (85.7) | 5 (71.4) | 1 (14.3) | 2 (28.6) | 5 (71.4) | 1 (14.3) | 3 (42.9) | 1 (14.3) | 1 (14.3) | ||||
| I | 1 (14.3) | 1 (14.3) | 1 (14.3) | 6 (85.7) | ||||||||||
| S | 1 (14.3) | 2 (28.6) | 7 (100) | 5 (71.4) | 7 (100) | 5 (71.4) | 1 (14.3) | 5 (71.4) | 7 (100) | 4 (57.1) | 6 (85.7) | 7 (100) | ||
| A. media | R | 1 (100) | 1 (100) | 1 (100) | 1 (100) | |||||||||
| I | 1 (100) | 1 (100) | ||||||||||||
| S | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | |||||||
| A. popoffii | R | 1 (100) | ||||||||||||
| I | ||||||||||||||
| S | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhanapala, P.M.; Kalupahana, R.S.; Kalupahana, A.W.; Wijesekera, D.P.H.; Kottawatta, S.A.; Jayasekera, N.K.; Silva-Fletcher, A.; Jagoda, S.S.S.d.S. Characterization and Antimicrobial Resistance of Environmental and Clinical Aeromonas Species Isolated from Fresh Water Ornamental Fish and Associated Farming Environment in Sri Lanka. Microorganisms 2021, 9, 2106. https://doi.org/10.3390/microorganisms9102106
Dhanapala PM, Kalupahana RS, Kalupahana AW, Wijesekera DPH, Kottawatta SA, Jayasekera NK, Silva-Fletcher A, Jagoda SSSdS. Characterization and Antimicrobial Resistance of Environmental and Clinical Aeromonas Species Isolated from Fresh Water Ornamental Fish and Associated Farming Environment in Sri Lanka. Microorganisms. 2021; 9(10):2106. https://doi.org/10.3390/microorganisms9102106
Chicago/Turabian StyleDhanapala, Pavithra M., Ruwani S. Kalupahana, Anil W. Kalupahana, D.P.H. Wijesekera, Sanda A. Kottawatta, Niromi K. Jayasekera, Ayona Silva-Fletcher, and S.S.S. de S. Jagoda. 2021. "Characterization and Antimicrobial Resistance of Environmental and Clinical Aeromonas Species Isolated from Fresh Water Ornamental Fish and Associated Farming Environment in Sri Lanka" Microorganisms 9, no. 10: 2106. https://doi.org/10.3390/microorganisms9102106