
Requirement of the LtsA Protein for Formation of the Mycolic Acid-Containing Layer on the Cell Surface of Corynebacterium glutamicum



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media and Growth Conditions
2.2. Analysis of Temperature- and Lysozyme-Sensitivity in C. glutamicum
2.3. Construction of GFP-Fused ltsA Genes and Microscopic Observation of Intracellular Localization of LtsA-GFP Fusion
2.4. Construction of the Recombinant Strains Carrying Mutant ltsA Alleles
2.5. Preparation of Polyclonal Rabbit Antiserum Containing Anti-LtsA Antibodies
2.6. Observation of Intracellular Localization of LtsA by Indirect Immunofluorescence Hybridization
2.7. Selective Staining of Peptidoglycan and Mycolic Acid-Containing Layers in C. glutamicum
2.8. Microscopic Observation of Bulge Formation in C. glutamicum
3. Results
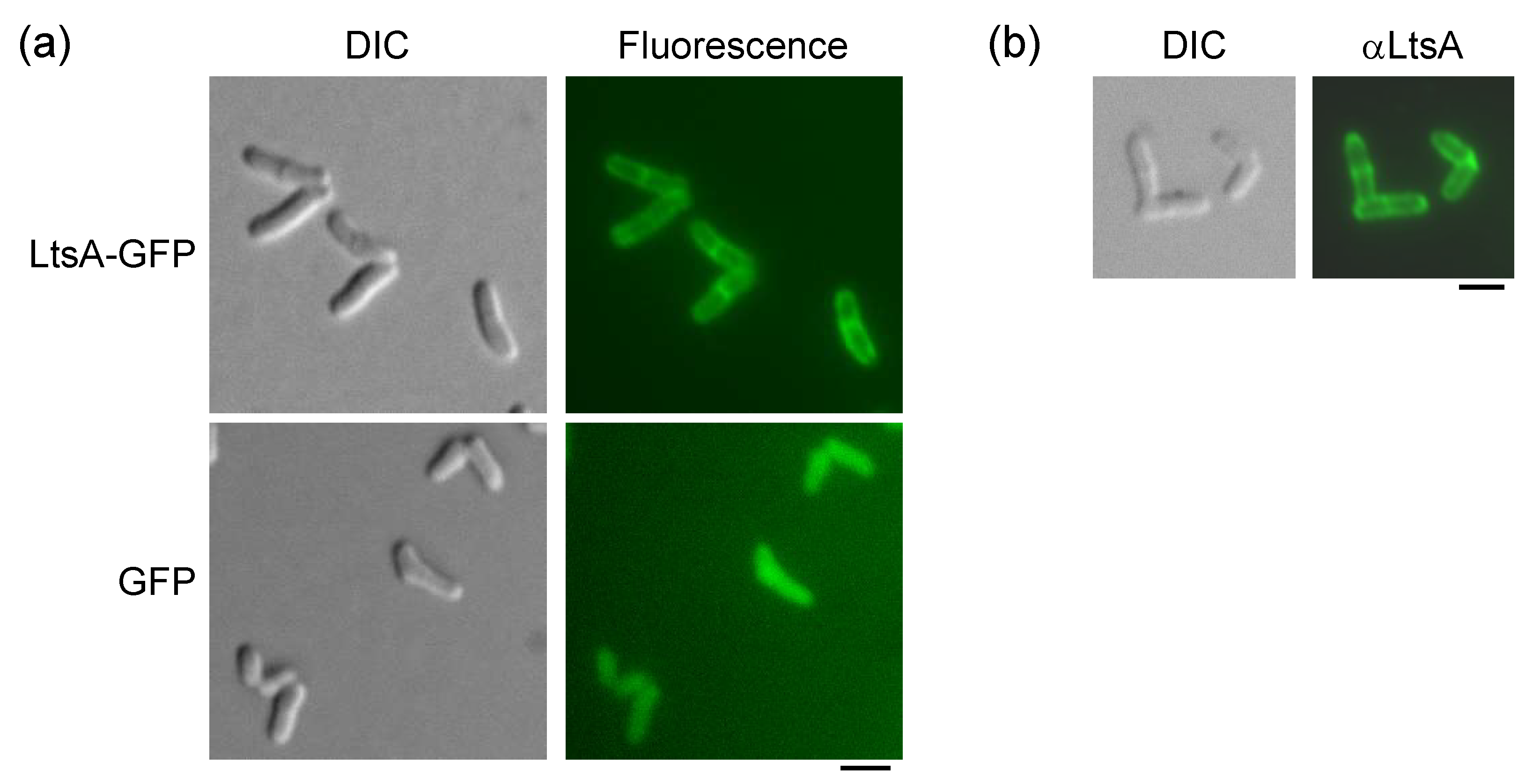
3.1. Intracellular Localization of LtsA Protein in C. glutamicum Cells
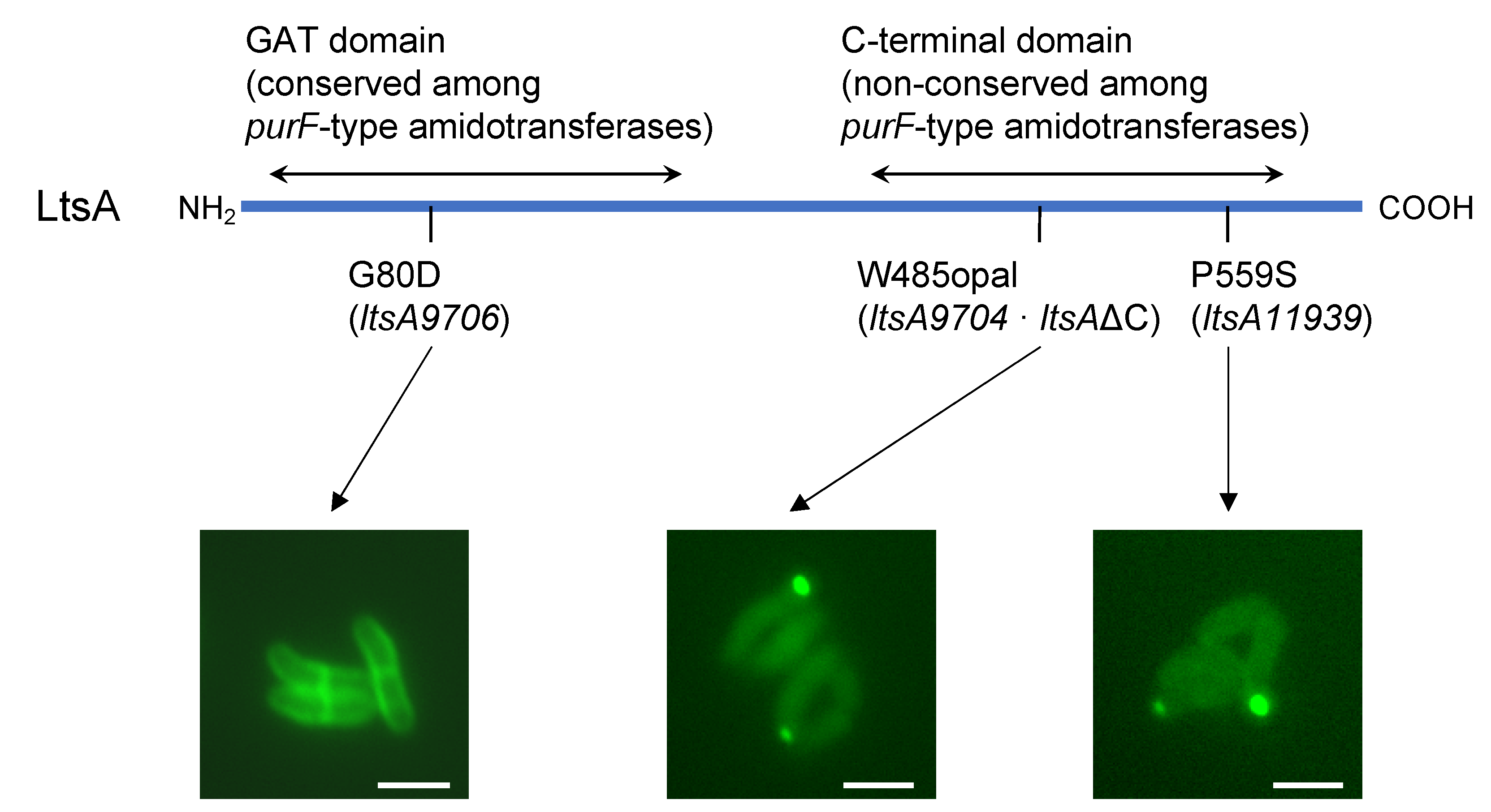
3.2. Requirement of C-Terminal Domain for Peripheral Localization of the LtsA Protein in C. glutamicum
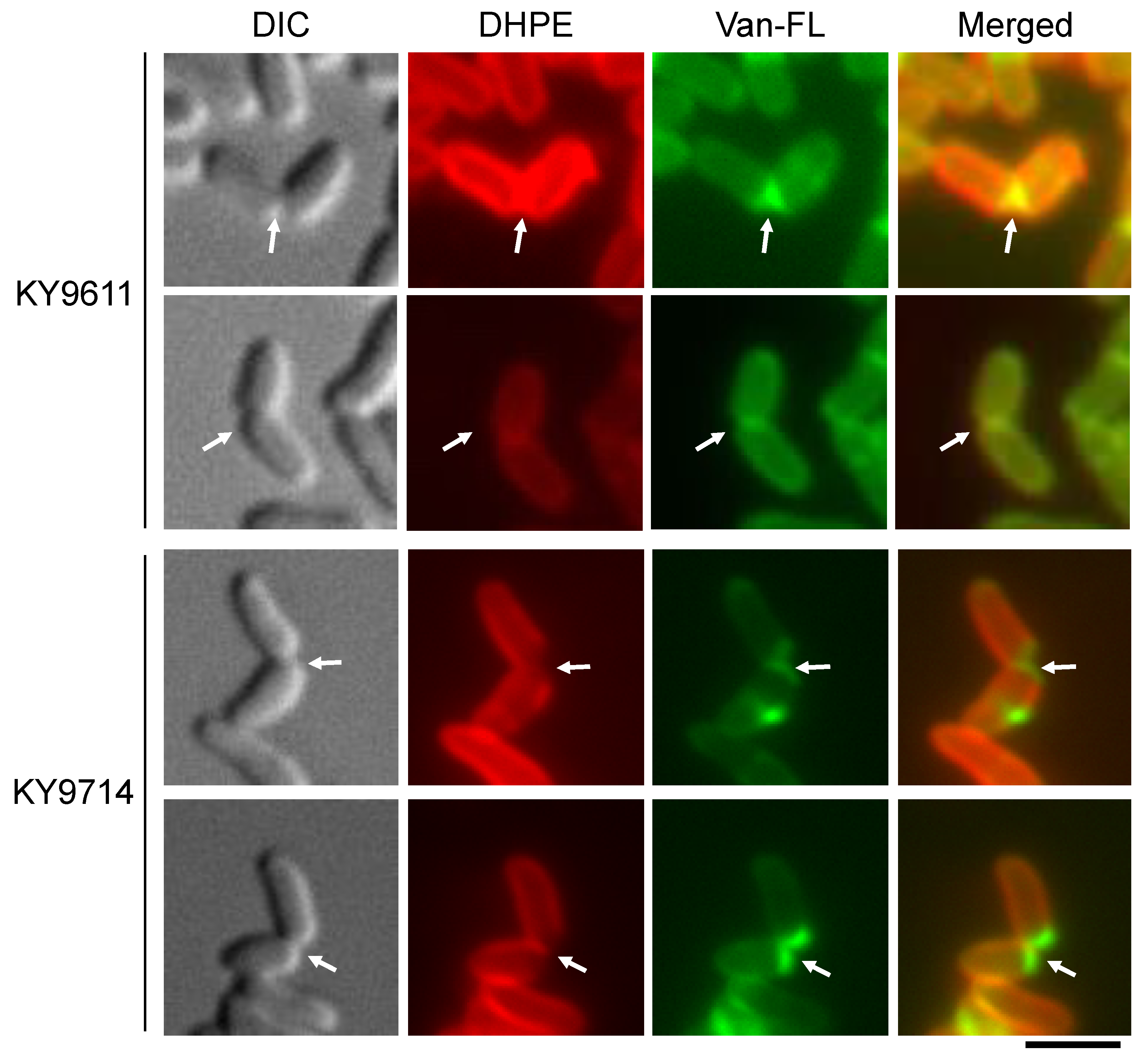
3.3. Defect in Mycolic Acid-Containing Layer Formation in the C. glutamicum ltsA Mutant
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kinoshita, S.; Udaka, S.; Shimono, M. Studies on the amino acid fermentation. Part 1. Production of L-glutamic acid by various microorganisms. J. Gen. Appl. Microbiol. 1957, 50, 193–205. [Google Scholar] [CrossRef]
- Udaka, S. Screening method for microorganisms accumulating metabolites and its use in the isolation of Micrococcus glutamicus. J. Bacteriol. 1960, 79, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, K.; Kitada, S.; Kinoshita, S. Studies on lysine fermentation I. The control mechanism on lysine accumulation by homoserine and threonine. J. Gen. Appl. Microbiol. 1961, 7, 145–154. [Google Scholar] [CrossRef]
- Sano, K.; Shiio, I. Microbial production of L-lysine III. Production by mutants resistant to S-(2-aminoethyl)-L-cysteine. J. Gen. Appl. Microbiol. 1970, 16, 373–391. [Google Scholar]
- Tsuchida, T.; Yoshinaga, F.; Kubota, K.; Momose, H. Production of L-valine by 2-thiazolealanine resistant mutants derived from glutamic acid producing bacteria. Agric. Biol. Chem. 1975, 39, 1319–1322. [Google Scholar] [CrossRef]
- Shiio, I.; Nakamori, S. Microbial production of L-threonine. Part II. Production by α-amino-β-hydroxyvaleric acid resistant mutants of glutamate producing bacteria. Agric. Biol. Chem. 1970, 34, 448–456. [Google Scholar]
- Baritugo, K.A.; Kim, H.T.; David, Y.; Choi, J.I.; Hong, S.H.; Jeong, K.J.; Choi, J.H.; Joo, J.C.; Park, S.J. Metabolic engineering of Corynebacterium glutamicum for fermentative production of chemicals in biorefinery. Appl. Microbiol. Biotechnol. 2018, 102, 3915–3937. [Google Scholar] [CrossRef]
- Becker, J.; Rohles, C.M.; Wittmann, C. Metabolically engineered Corynebacterium glutamicum for bio-based production of chemicals, fuels, materials, and healthcare products. Metab. Eng. 2018, 50, 122–141. [Google Scholar] [CrossRef] [PubMed]
- Nara, T.; Samejima, H.; Kinoshita, S. Effect of penicillin on amino acid fermentation. Agric. Biol. Chem. 1964, 28, 120–124. [Google Scholar] [CrossRef]
- Shiio, I.; Otsuka, S.I.; Takahashi, M. Effect of biotin on the bacterial formation of glutamic acid. I. Glutamate formation and cellular premeability of amino acids. J. Biochem. 1962, 51, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Takinami, K.; Yoshii, H.; Tsuri, H.; Okada, H. Biochemical effects of fatty acid and its derivatives on L-glutamic acid fermentation. Part III. Biotin-Tween 60 relationship in the accumulation of L-glutamic acid and the growth of Brevibacterium lactofermentum. Agric. Biol. Chem. 1965, 29, 351–359. [Google Scholar]
- Nakamura, J.; Hirano, S.; Ito, H.; Wachi, M. Mutations of the Corynebacterium glutamicum NCgl1221 gene, encoding a mechanosensitive channel homolog, induce L-glutamic acid production. Appl. Environ. Microbiol. 2007, 73, 4491–4498. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, K.; Murata, J.; Konishi, T.; Yabe, I.; Nakamatsu, T.; Kawasaki, H. Glutamate is excreted across the cytoplasmic membrane through the NCgl1221 channel of Corynebacterium glutamicum by passive diffusion. Biosci. Biotechnol. Biochem. 2012, 76, 1422–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, M.; Börngen, K.; Nomura, T.; Battle, A.R.; Marin, K.; Martinac, B.; Krämer, R. Glutamate efflux mediated by Corynebacterium glutamicum MscCG, Escherichia coli MscS, and their derivatives. Biochim. Biophys. Acta 2013, 1828, 1230–1240. [Google Scholar] [CrossRef]
- Hirasawa, T.; Wachi, M. Glutamate fermentation-2: Mechanism of L-glutamate overproduction in Corynebacterium glutamicum. In Amino Acid Fermentation; Yokota, A., Ikeda, M., Eds.; Springer: Tokyo, Japan, 2017; pp. 57–72. [Google Scholar] [CrossRef]
- Nakayama, Y. Corynebacterium glutamicum mechanosensing: From osmoregulation to L-glutamate secretion for the avian microbiota-gut-brain axis. Microorganisms 2021, 9, 201. [Google Scholar] [CrossRef]
- Wachi, M. Amino acid exporters in Corynebacterium glutamicum. In Microbiology Monographs: Volume 23 Corynebacterium glutamicum; Inui, M., Toyoda, K., Eds.; Springer: Tokyo, Japan, 2020; pp. 267–284. [Google Scholar]
- Houssin, C.; de Sousa d’Auria, C.; Constantinesco, F.; Dietrich, C.; Labarre, C.; Bayan, N. Architecture and biogenesis of the cell envelope of Corynebacterium glutamicum. In Microbiology Monographs: Volume 23 Corynebacterium glutamicum; Inui, M., Toyoda, K., Eds.; Springer: Tokyo, Japan, 2020; pp. 25–60. [Google Scholar]
- Daffé, M. The cell envelope of corynebacteria. In Handbook of Corynebacterium glutamicum; Eggeling, L., Bott, M., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 121–148. [Google Scholar]
- Hirasawa, T.; Wachi, M.; Nagai, K. A mutation in the Corynebacterium glutamicum ltsA gene causes susceptibility to lysozyme, temperature-sensitive growth, and L-glutamate production. J. Bacteriol. 2000, 182, 2696–2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirasawa, T.; Wachi, M.; Nagai, K. L-Glutamate production by lysozyme-sensitive Corynebacterium glutamicum ltsA mutant strains. BMC Biotechnol. 2001, 1, 9. [Google Scholar] [CrossRef]
- Mei, B.G.; Zalkin, H. Amino-terminal deletions define a glutamine amide transfer domain in glutamine phosphoribosylpyrophosphate amidotransferase and other PurF-type amidotransferases. J. Bacteriol. 1990, 172, 3512–3514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, B.; Zalkin, H. A cysteine-histidine-aspartate catalytic triad is involved in glutamine amide transfer function in purF-type glutamine amidotransferases. J. Biol. Chem. 1989, 264, 16613–16619. [Google Scholar] [CrossRef]
- Massière, F.; Badet-Denisot, M.A. The mechanism of glutamine-dependent amidotransferases. Cell. Mol. Life Sci. 1998, 54, 205–222. [Google Scholar] [CrossRef]
- Levefaudes, M.; Patin, D.; de Sousa-d’Auria, C.; Chami, M.; Blanot, D.; Herve, M.; Arthur, M.; Houssin, C.; Mengin-Lecreulx, D. Diaminopimelic acid amidation in Corynebacteriales: New insights into the role of LtsA in peptidoglycan modification. J. Biol. Chem. 2015, 290, 13079–13094. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, Y.; Hirasawa, T.; Hayakawa, K.; Nagai, K.; Wachi, M. Fluorescent phospholipid analogs as microscopic probes for detection of the mycolic acid-containing layer in Corynebacterium glutamicum: Detecting alterations in the mycolic acid-containing layer following ethambutol treatment. Biosci. Biotechnol. Biochem. 2005, 69, 2051–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirasawa, T.; Kumagai, Y.; Nagai, K.; Wachi, M. A Corynebacterium glutamicum rnhA recG double mutant showing lysozyme-sensitivity, temperature-sensitive growth, and UV-sensitivity. Biosci. Biotechnol. Biochem. 2003, 67, 2416–2424. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, A.; Tauch, A.; Jager, W.; Kalinowski, J.; Thierbach, G.; Puhler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Hiraga, S.; Ichinose, C.; Niki, H.; Yamazoe, M. Cell cycle-dependent duplication and bidirectional migration of SeqA-associated DNA-protein complexes in E. coli. Mol. Cell. 1998, 1, 381–387. [Google Scholar] [CrossRef]
- Boehlein, S.K.; Richards, N.G.; Walworth, E.S.; Schuster, S.M. Arginine 30 and asparagine 74 have functional roles in the glutamine dependent activities of Escherichia coli asparagine synthetase B. J. Biol. Chem. 1994, 269, 26789–26795. [Google Scholar] [CrossRef]
- Mitani, Y.; Meng, X.; Kamagata, Y.; Tamura, T. Characterization of LtsA from Rhodococcus erythropolis, an enzyme with glutamine amidotransferase activity. J. Bacteriol. 2005, 187, 2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dajkovic, A.; Tesson, B.; Chauhan, S.; Courtin, P.; Keary, R.; Flores, P.; Marliere, C.; Filipe, S.R.; Chapot-Chartier, M.P.; Carballido-Lopez, R. Hydrolysis of peptidoglycan is modulated by amidation of meso-diaminopimelic acid and Mg2+ in Bacillus subtilis. Mol. Microbiol. 2017, 104, 972–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, E.; Rolain, T.; Courtin, P.; Hols, P.; Chapot-Chartier, M.P. Identification of the amidotransferase AsnB1 as being responsible for meso-diaminopimelic acid amidation in Lactobacillus plantarum peptidoglycan. J. Bacteriol. 2011, 193, 6323–6330. [Google Scholar] [CrossRef] [Green Version]
- Daniel, R.A.; Errington, J. Control of cell morphogenesis in bacteria: Two distinct ways to make a rod-shaped cell. Cell 2003, 113, 767–776. [Google Scholar] [CrossRef] [Green Version]
- Marienfeld, S.; Uhlemann, E.M.; Schmid, R.; Kramer, R.; Burkovski, A. Ultrastructure of the Corynebacterium glutamicum cell wall. Antonie Van. Leeuwenhoek 1997, 72, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Puech, V.; Chami, M.; Lemassu, A.; Laneelle, M.A.; Schiffler, B.; Gounon, P.; Bayan, N.; Benz, R.; Daffe, M. Structure of the cell envelope of corynebacteria: Importance of the non-covalently bound lipids in the formation of the cell wall permeability barrier and fracture plane. Microbiology 2001, 147, 1365–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, K.; Armstrong, E.L.; Kunugi, K.A.; Kilburn, J.O. Inhibition by ethambutol of mycolic acid transfer into the cell wall of Mycobacterium smegmatis. Antimicrob. Agents Chemother. 1979, 16, 240–242. [Google Scholar] [CrossRef] [Green Version]
- Takayama, K.; Kilburn, J.O. Inhibition of synthesis of arabinogalactan by ethambutol in Mycobacterium smegmatis. Antimicrob. Agents Chemother. 1989, 33, 1493–1499. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Rodriguez-Rivera, F.P.; Lim, H.C.; Bell, J.C.; Bernhardt, T.G.; Bertozzi, C.R.; Theriot, J.A. Sequential assembly of the septal cell envelope prior to V snapping in Corynebacterium glutamicum. Nat. Chem. Biol. 2019, 15, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Kacem, R.; De Sousa-D’Auria, C.; Tropis, M.; Chami, M.; Gounon, P.; Leblon, G.; Houssin, C.; Daffe, M. Importance of mycoloyltransferases on the physiology of Corynebacterium glutamicum. Microbiology 2004, 150, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Portevin, D.; de Sousa, D.; Auria, C.; Houssin, C.; Grimaldi, C.; Chami, M.; Daffé, M.; Guilhot, C. A polyketide synthase catalyzes the last condensation step of mycolic acid biosynthesis in mycobacteria and related organisms. Proc. Natl. Acad. Sci. USA 2004, 101, 314. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Liu, J. AsnB is involved in natural resistance of Mycobacterium smegmatis to multiple drugs. Antimicrob. Agents Chemother. 2006, 50, 250–255. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumagai, Y.; Hirasawa, T.; Wachi, M. Requirement of the LtsA Protein for Formation of the Mycolic Acid-Containing Layer on the Cell Surface of Corynebacterium glutamicum. Microorganisms 2021, 9, 409. https://doi.org/10.3390/microorganisms9020409
Kumagai Y, Hirasawa T, Wachi M. Requirement of the LtsA Protein for Formation of the Mycolic Acid-Containing Layer on the Cell Surface of Corynebacterium glutamicum. Microorganisms. 2021; 9(2):409. https://doi.org/10.3390/microorganisms9020409
Chicago/Turabian StyleKumagai, Yutaro, Takashi Hirasawa, and Masaaki Wachi. 2021. "Requirement of the LtsA Protein for Formation of the Mycolic Acid-Containing Layer on the Cell Surface of Corynebacterium glutamicum" Microorganisms 9, no. 2: 409. https://doi.org/10.3390/microorganisms9020409