Isolation and Characterization of Bacillus cereus Phage vB_BceP-DLc1 Reveals the Largest Member of the Φ29-Like Phages


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Isolation and Purification of DLc1
2.3. Preparation of DLc1 Phage Stock with High Titer
2.4. Transmission Electron Microscopy
2.5. Genome Sequencing and Analysis
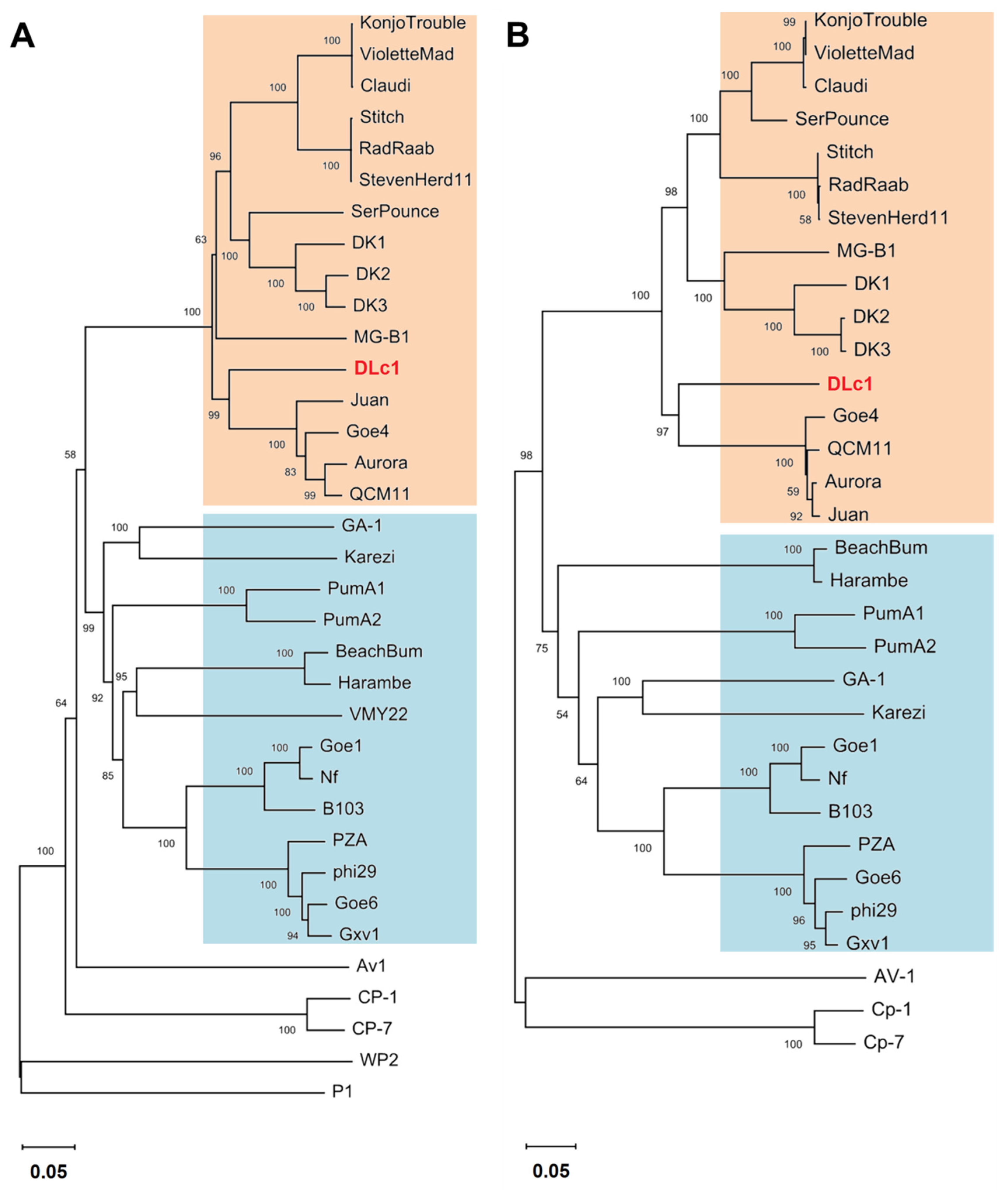
2.6. Phylogenetic Analysis
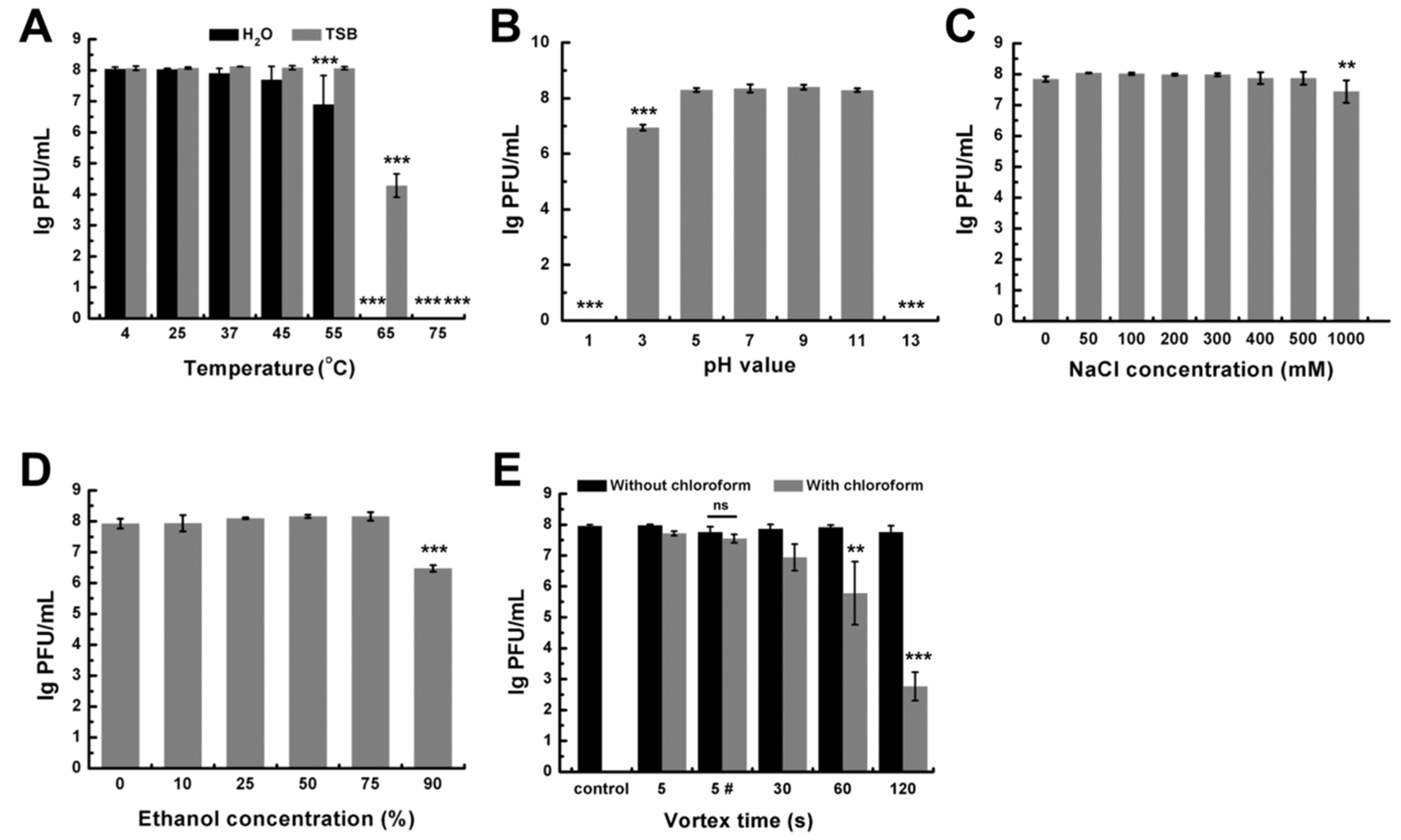
2.7. Stability of DLc1
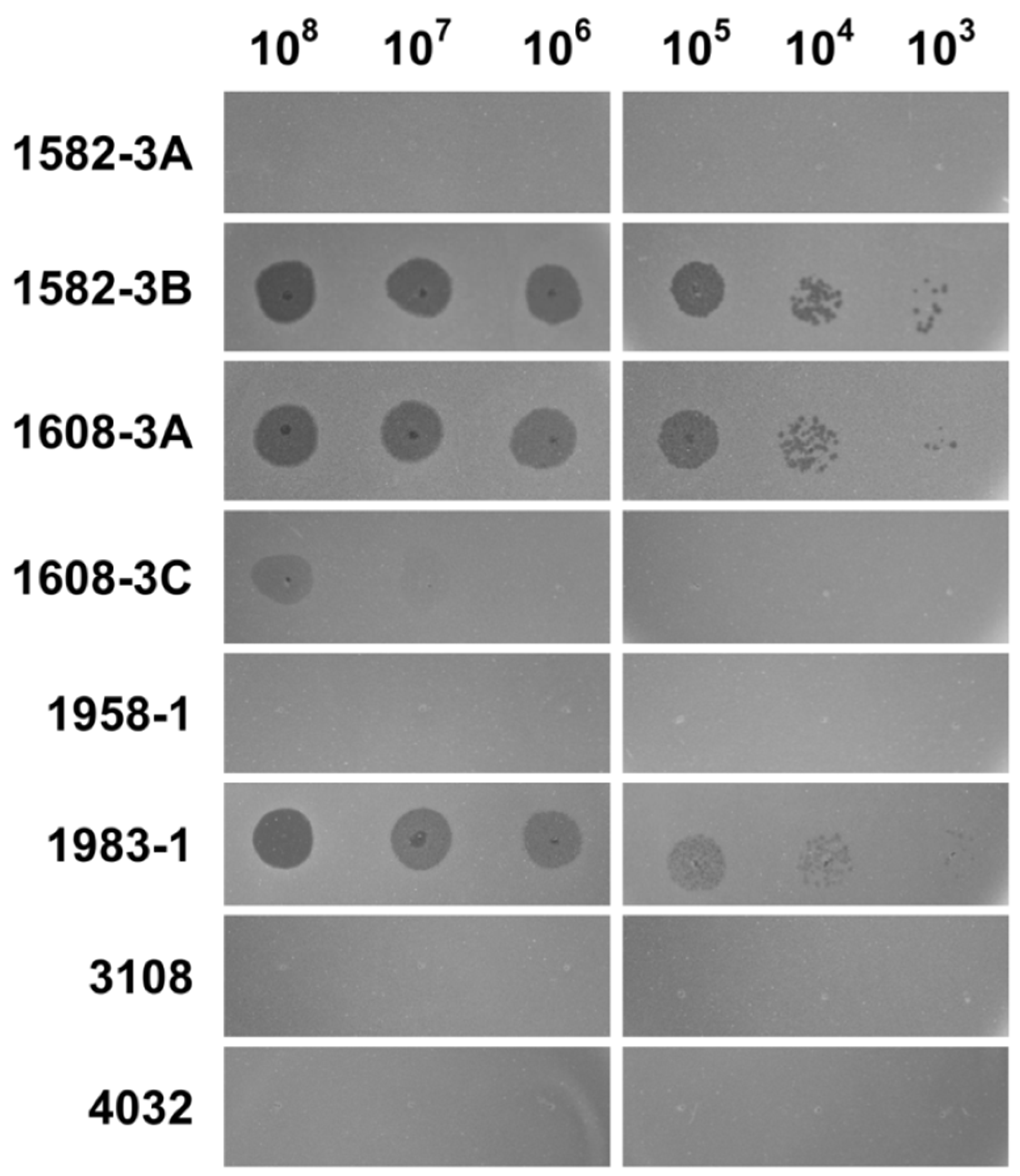
2.8. Host Range Determination
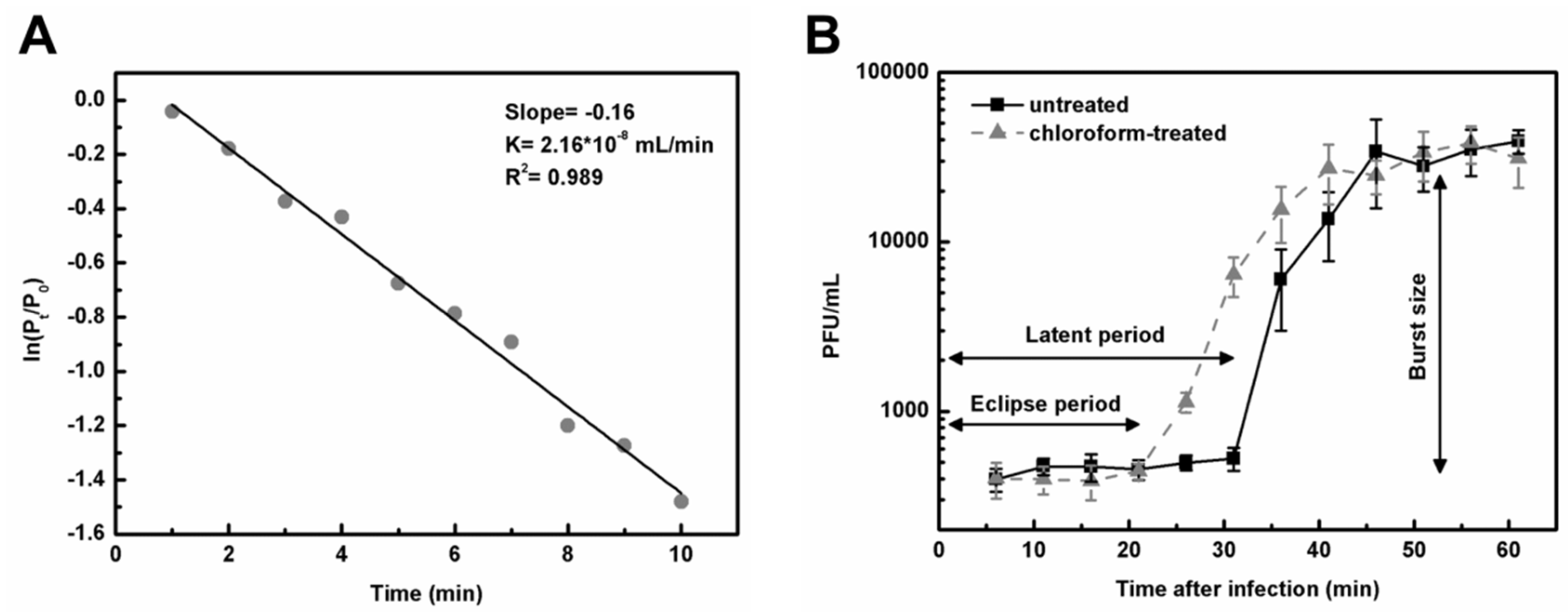
2.9. Adsorption Rate Constant k
2.10. One-Step Growth Curves
2.11. Phage Adsorption Assays
2.12. Statistics
2.13. Genomic Data Availability
3. Results
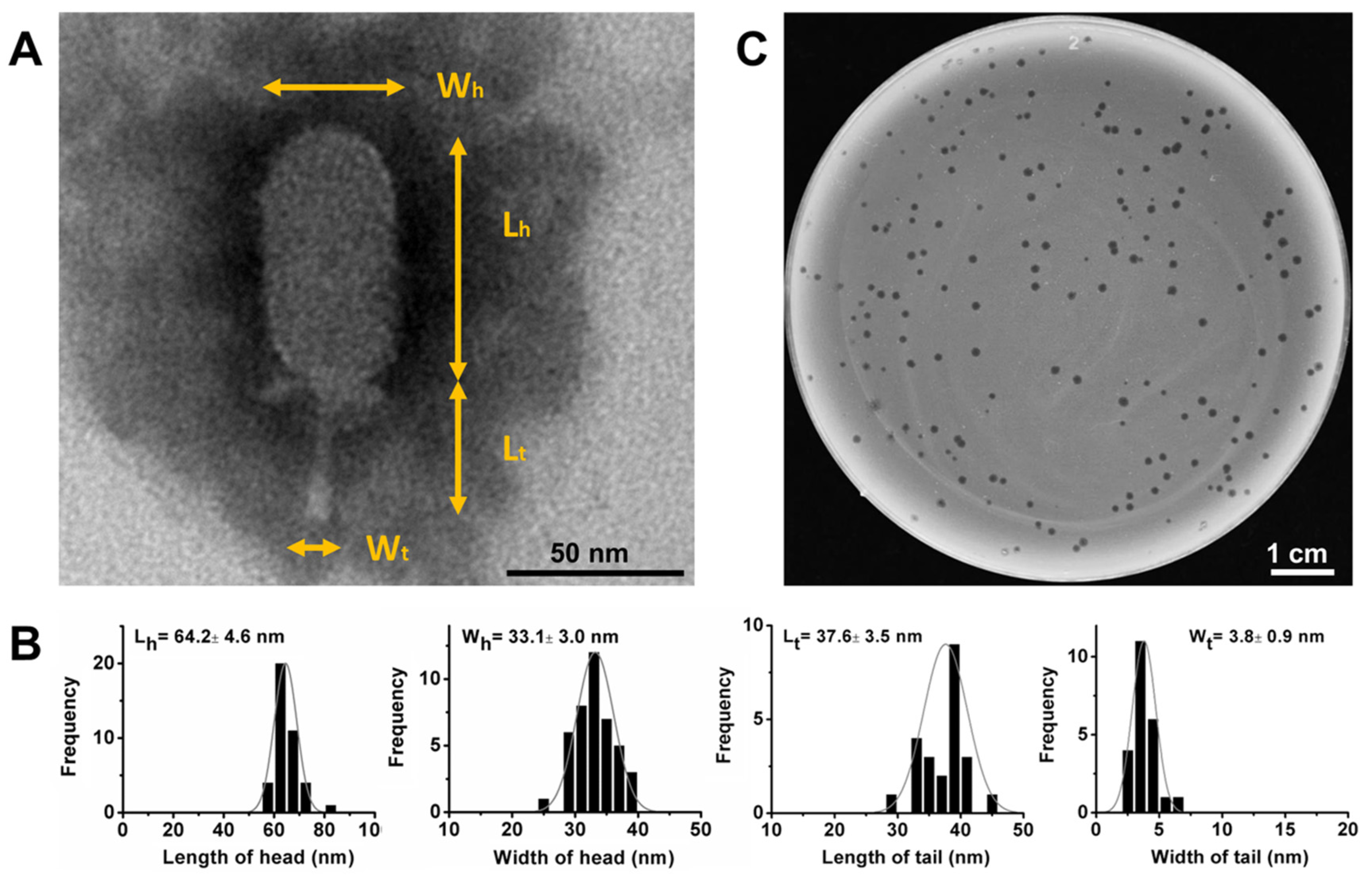
3.1. Isolation and Morphological Characteristics of DLc1
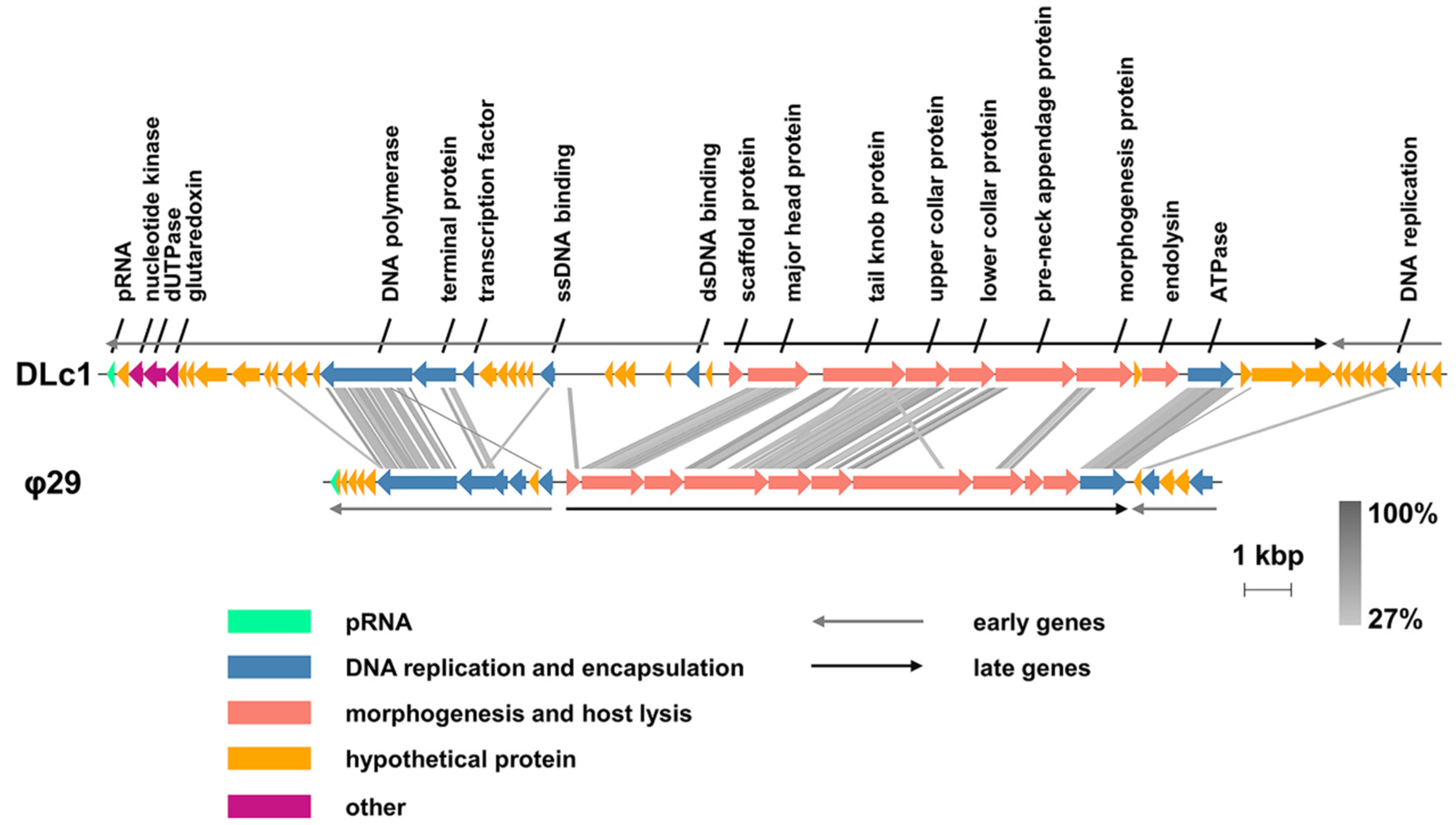
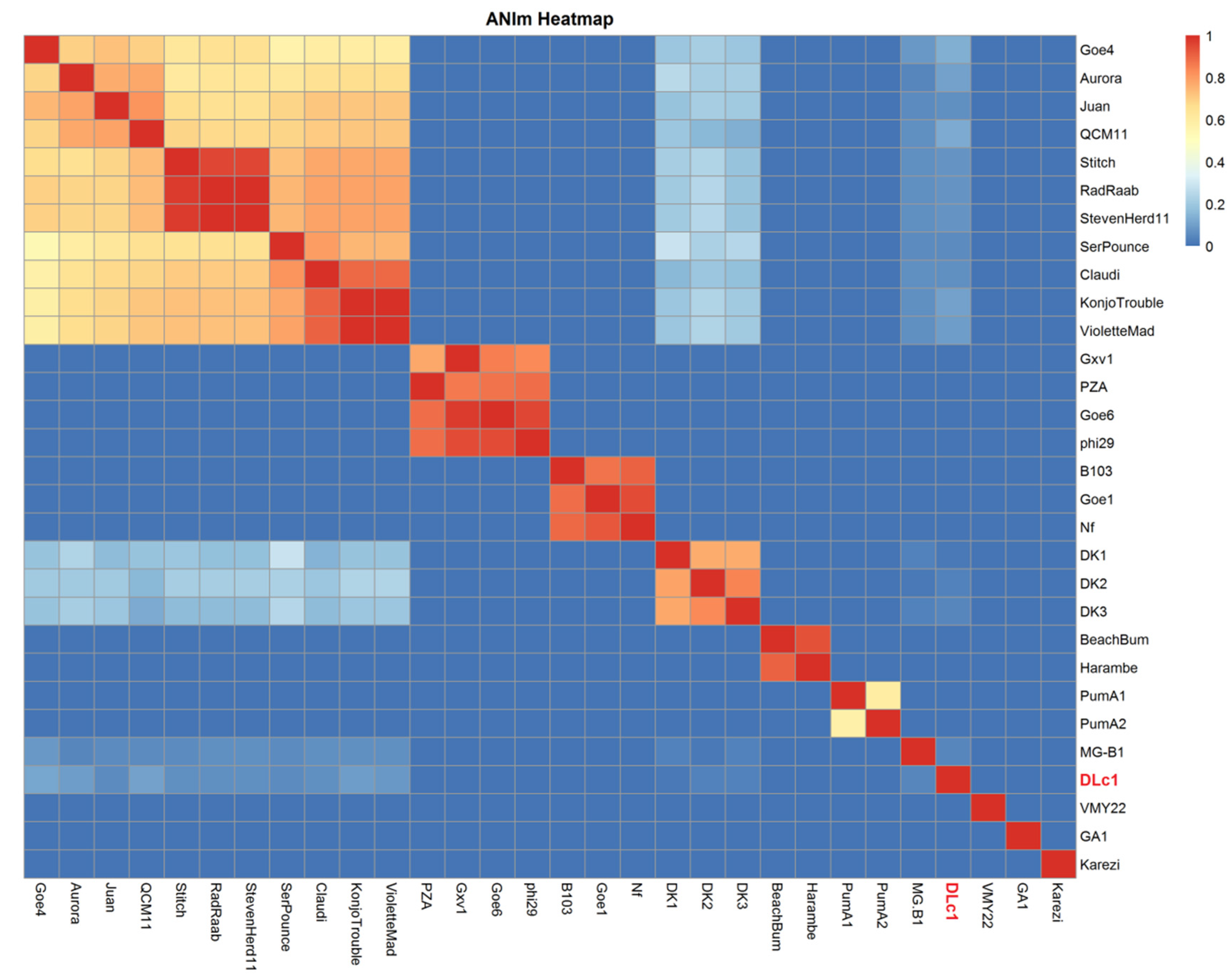
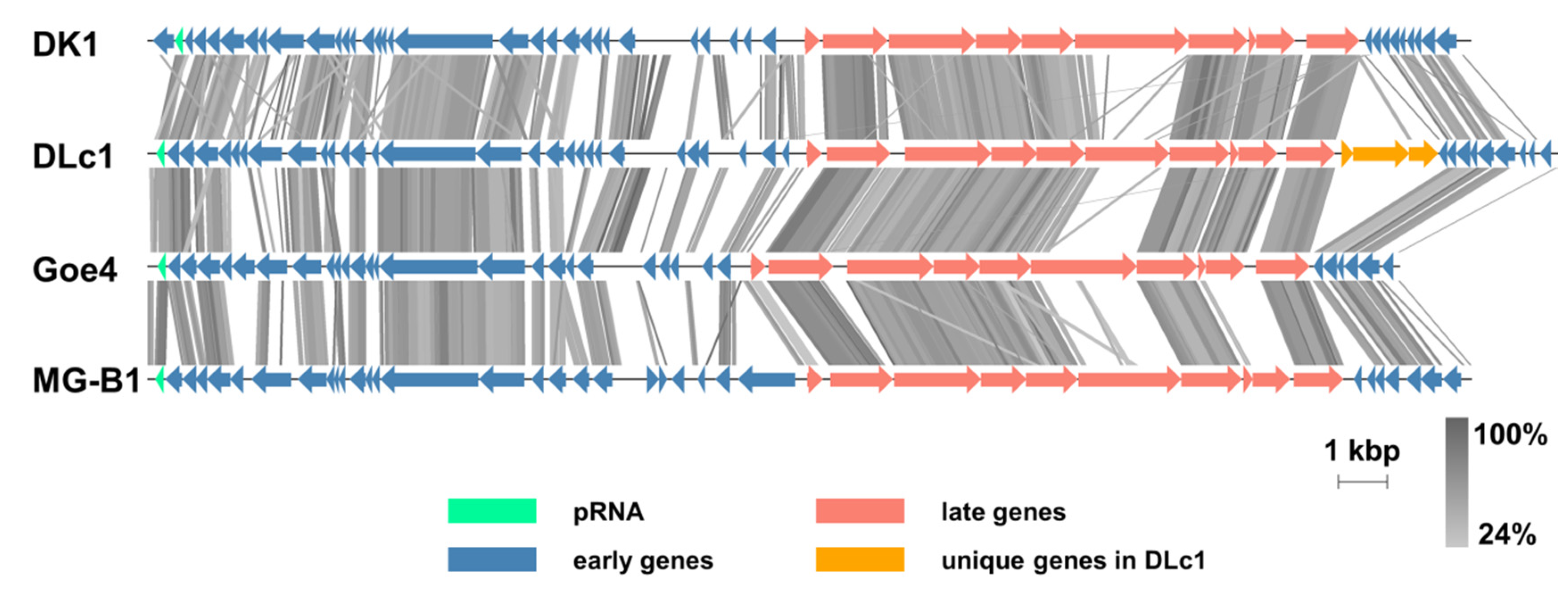
3.2. Genomic and Phylogenetic Analysis
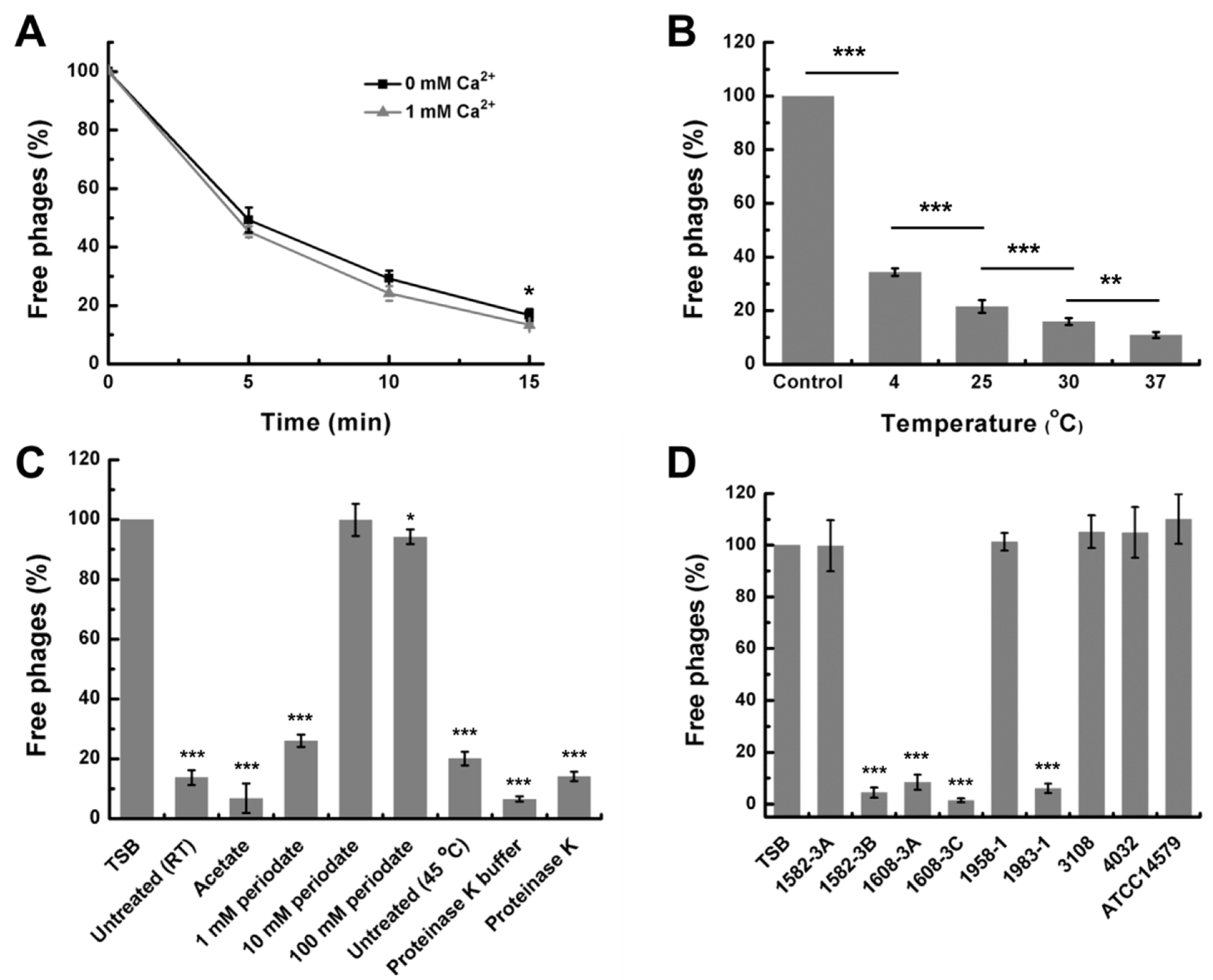
3.3. Stability of DLc1 to Environmental Stresses
3.4. Host Range of DLc1
3.5. Adsorption Rate Constant and One-Step Growth Curves
3.6. Phage Adsorption Assay
4. Discussion
4.1. Classification Proposal
4.2. Genomic Features
4.3. Interaction between DLc1 and Host
4.4. The Application Potential
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jessberger, N.; Kranzler, M.; Da Riol, C.; Schwenk, V.; Buchacher, T.; Dietrich, R.; Ehling-Schulz, M.; Märtlbauer, E. Assessing the toxic potential of enteropathogenic Bacillus cereus. Food Microbiol. 2019, 84, 103276. [Google Scholar] [CrossRef]
- Bratcher, D.F. Bacillus species (Anthrax). In Principles and Practice of Pediatric Infectious Diseases, 5th ed.; Long, S., Prober, C., Fischer, M., Eds.; Elsevier: Philadelphia, PA, USA, 2017; pp. 770–773.e2. [Google Scholar]
- Prod’hom, G.; Bille, J. Aerobic Gram-positive Bacilli. In Infectious Diseases, 4th ed.; Cohen, J., Powderly, W.G., Opal, S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1537–1552.e2. [Google Scholar]
- Yu, S.; Yu, P.; Wang, J.; Li, C.; Guo, H.; Liu, C.; Kong, L.; Yu, L.; Wu, S.; Lei, T.; et al. A study on prevalence and characterization of Bacillus cereus in ready-to-eat foods in China. Front. Microbiol. 2020, 10, 3043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.; Yu, S.; Wang, J.; Guo, H.; Zhang, Y.; Liao, X.; Zhang, J.; Wu, S.; Gu, Q.; Xue, L.; et al. Bacillus cereus isolated from vegetables in China: Incidence, genetic diversity, virulence genes, and antimicrobial resistance. Front. Microbiol. 2019, 10, 948. [Google Scholar] [CrossRef] [PubMed]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage diversity, genomics and phylogeny. Nat. Rev. Microbiol. 2020, 18, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage therapy: A renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehman, S.; Ali, Z.; Khan, M.; Bostan, N.; Naseem, S. The dawn of phage therapy. Rev. Med. Virol. 2019, 29, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Pires, D.P.; Costa, A.R.; Azeredo, J. Phage therapy: Going temperate? Trends Microbiol. 2019, 27, 368–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, P.; Buckling, A. Bacteria-phage antagonistic coevolution in soil. Science 2011, 332, 106–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, S.B.; Green, S.I.; Liu, C.G.; Salazar, K.C.; Clark, J.R.; Terwilliger, A.L.; Kaplan, H.B.; Maresso, A.W.; Trautner, B.W.; Ramig, R.F. Constructing and characterizing bacteriophage libraries for phage therapy of human infections. Front. Microbiol. 2019, 10, 2537. [Google Scholar] [CrossRef] [Green Version]
- Gurney, J.; Brown, S.P.; Kaltz, O.; Hochberg, M.E. Steering phages to combat bacterial pathogens. Trends Microbiol. 2020, 28, 85–94. [Google Scholar] [CrossRef]
- Gillis, A.; Mahillon, J. Phages preying on Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis: Past, present and future. Viruses 2014, 6, 2623–2672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, X.; Zhang, C.; Fang, Y.; Zhang, Q.; Lin, L.; Tang, B.; Wei, Y. Isolation and characterization of glacier VMY22, a novel lytic cold-active bacteriophage of Bacillus cereus. Virol. Sin. 2015, 30, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Ding, Y.; Wu, Q.; Wang, J.; Zhang, J.; Li, H.; Yu, S.; Yu, P.; Gao, T.; Zeng, H.; et al. Genome sequencing and characterization of three Bacillus cereus-specific phages, DK1, DK2, and DK3. Arch. Virol. 2019, 164, 1927–1929. [Google Scholar] [CrossRef] [PubMed]
- Okinaka, R.T.; Keim, P. The Phylogeny of Bacillus cereus sensu lato. Microbiol. Spectr. 2016, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bazinet, A.L. Pan-genome and phylogeny of Bacillus cereus sensu lato. BMC Evol. Biol. 2017, 17, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.H.; Walter, M.H.; Yang, L.; Chen, S.C.; Winston, V.; Thomas, M.A. Predicting genome terminus sequences of Bacillus cereus-group bacteriophage using next generation sequencing data. BMC Genom. 2017, 18, 350. [Google Scholar] [CrossRef]
- Redondo, R.A.F.; Kupczok, A.; Stift, G.; Bollback, J.P. Complete genome sequence of the novel phage MG-B1 infecting Bacillus weihenstephanensis. Genome Announc. 2013, 1, e00216-13. [Google Scholar] [CrossRef] [Green Version]
- Schilling, T.; Hoppert, M.; Hertel, R. Genomic analysis of the recent viral isolate vB_BthP-Goe4 reveals increased diversity of φ29-like phages. Viruses 2018, 10, 624. [Google Scholar] [CrossRef] [Green Version]
- Salas, M. Protein-priming of DNA replication. Annu. Rev. Biochem. 1991, 60, 39–71. [Google Scholar] [CrossRef]
- Salas, M. Phage φ29 and its relatives. In The Bacteriophages, 2nd ed.; Calendar, R., Ed.; Oxford University Press: NewYork, NY, USA, 2006; pp. 315–330. [Google Scholar]
- Xu, Z.; Sun, Y.; Weber, J.K.; Cao, Y.; Wang, W.; Jasinski, D.; Guo, P.; Zhou, R.; Li, J. Directional mechanical stability of bacteriophage φ29 motor’s 3WJ-pRNA: Extraordinary robustness along portal axis. Sci. Adv. 2017, 3, e1601684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A.M.Q.; Lefkowitz, E.J.; Mushegian, A.R.; Adams, M.J.; Dutilh, B.E.; Gorbalenya, A.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; Knowles, N.J.; et al. Changes to taxonomy and the international code of virus classification and nomenclature ratified by the international committee on taxonomy of viruses. Arch. Virol. 2018, 163, 2601–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Liang, Y.; Huang, S.; Zhang, J.; Wang, J.; Chen, H.; Ye, Y.; Gao, X.; Wu, Q.; Tan, Z. Isolation and characterization of the novel phages vB_VpS_BA3 and vB_VpS_CA8 for lysing Vibrio parahaemolyticus. Front. Microbiol. 2020, 11, 259. [Google Scholar] [CrossRef] [PubMed]
- Phage Protocols. Available online: https://cpt.tamu.edu/phage-links/phage-protocols/ (accessed on 3 July 2019).
- Imam, M.; Alrashid, B.; Patel, F.; Dowah, A.S.A.; Brown, N.; Millard, A.; Clokie, M.R.J.; Galyov, E.E. vB_PaeM_MIJ3, a novel jumbo phage infecting Pseudomonas aeruginosa, possesses unusual genomic features. Front. Microbiol. 2019, 10, 2772. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2020. Available online: https://www.R-project.org/ (accessed on 21 August 2020).
- Pang, R.; Wu, S.; Zhang, F.; Huang, J.; Wu, H.; Zhang, J.; Li, Y.; Ding, Y.; Zhang, J.; Chen, M.; et al. The genomic context for the evolution and transmission of community-associated Staphylococcus aureus ST59 through the food chain. Front. Microbiol. 2020, 11, 422. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 2019, 47, D351–D360. [Google Scholar] [CrossRef] [Green Version]
- Nawrocki, E.P.; Eddy, S.R. Infernal 1.1: 100-fold faster RNA homology searches. Bioinformatics 2013, 29, 2933–2935. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Schluttenhofer, C.; Pattanaik, S.; Patra, B.; Yuan, L. Analyses of catharanthus roseus and arabidopsis thaliana WRKY transcription factors reveal involvement in jasmonate signaling. BMC Genom. 2014, 15, 502. [Google Scholar] [CrossRef] [Green Version]
- Mazzocco, A.; Waddell, T.E.; Lingohr, E.; Johnson, R.P. Enumeration of bacteriophages using the small drop plaque assay system. In Bacteriophages: Methods and Protocols; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: New York, NY, USA, 2009; Volume 1, pp. 81–85. [Google Scholar]
- Hyman, P.; Abedon, S.T. Practical methods for determining phage growth parameters. In Bacteriophages: Methods and Protocols; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: New York, NY, USA, 2009; Volume 1, pp. 175–202. [Google Scholar]
- Schuch, R.; Pelzek, A.J.; Nelson, D.C.; Fischetti, V.A. The PlyB endolysin of bacteriophage vB_BanS_Bcp1 exhibits broad-spectrum bactericidal activity against Bacillus cereus sensu lato isolates. Appl. Environ. Microbiol. 2019, 85, e00003-19. [Google Scholar] [CrossRef] [Green Version]
- Kropinski, A.M.; Mazzocco, A.; Waddell, T.E.; Lingohr, E.; Johnson, R.P. Enumeration of bacteriophages by double agar overlay plaque assay. In Bacteriophages: Methods and Protocols; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: New York, NY, USA, 2009; Volume 1, pp. 69–76. [Google Scholar]
- Methods for Counting Bacteria. Available online: http://mathbench.org.au/microbiology/methods-for-counting-bacteria/ (accessed on 12 May 2020).
- Kropinski, A.M. Measurement of the rate of attachment of bacteriophage to cells. In Bacteriophages: Methods and Protocols; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: New York, NY, USA, 2009; Volume 1, pp. 151–155. [Google Scholar]
- Kong, M.; Ryu, S. Bacteriophage PBC1 and its endolysin as an antimicrobial agent against Bacillus cereus. Appl. Environ. Microbiol. 2015, 81, 2274–2283. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, M.C.H.; van Alphen, L.B.; Harboe, A.; Li, J.; Christensen, B.B.; Szymanski, C.M.; Brøndsted, L. Bacteriophage F336 recognizes the capsular phosphoramidate modification of Campylobacter jejuni NCTC11168. J. Bacteriol. 2011, 193, 6742–6749. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.; Ding, Y.; Wu, Q.; Wang, J.; Zhang, J.; Yu, S.; Yu, P.; Liu, C.; Kong, L.; Feng, Z.; et al. Prevalence, virulence genes, antimicrobial susceptibility, and genetic diversity of Bacillus cereus isolated from pasteurized milk in China. Front. Microbiol. 2018, 9, 533. [Google Scholar] [CrossRef] [Green Version]
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. Virus Taxonomy, Classification and Nomenclature of Viruses; Elsevier: Amsterdam, The Netherlands, 2012; pp. 79–85. [Google Scholar]
- Adriaenssens, E.M.; Brister, J.R. How to name and classify your phage: An informal guide. Viruses 2017, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Chistol, G.; Hetherington, C.L.; Tafoya, S.; Aathavan, K.; Schnitzbauer, J.; Grimes, S.; Jardine, P.J.; Bustamante, C. A viral packaging motor varies its DNA rotation and step size to preserve subunit coordination as the capsid fills. Cell 2014, 157, 702–713. [Google Scholar] [CrossRef] [Green Version]
- Farley, M.M.; Tu, J.; Kearns, D.B.; Molineux, I.J.; Liu, J. Ultrastructural analysis of bacteriophage φ29 during infection of Bacillus subtilis. J. Struct. Biol. 2017, 197, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Kampf, G. Efficacy of ethanol against viruses in hand disinfection. J. Hosp. Infect. 2018, 98, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Tolstoy, I.; Kropinski, A.M.; Brister, J.R. Bacteriophage taxonomy: An evolving discipline. In Bacteriophage Therapy—From Lab to Clinical Practice; Azeredo, J., Sillankorva, S., Eds.; Humana Press: New York, NY, USA, 2018; pp. 57–71. [Google Scholar]
- Ciufo, S.; Kannan, S.; Sharma, S.; Badretdin, A.; Clark, K.; Turner, S.; Brover, S.; Schoch, C.L.; Kimchi, A.; DiCuccio, M. Using average nucleotide identity to improve taxonomic assignments in prokaryotic genomes at the NCBI. Int. J. Syst. Evol. Microbiol. 2018, 68, 2386–2392. [Google Scholar] [CrossRef]
- D’Souza, A.W.; Potter, R.F.; Wallace, M.; Shupe, A.; Patel, S.; Sun, X.; Gul, D.; Kwon, J.H.; Andleeb, S.; Burnham, C.A.D.; et al. Spatiotemporal dynamics of multidrug resistant bacteria on intensive care unit surfaces. Nat. Commun. 2019, 10, 4569. [Google Scholar] [CrossRef] [Green Version]
- Jean, N.L.; Rutherford, T.J.; Löwe, J. FtsK in motion reveals its mechanism for double-stranded DNA translocation. Proc. Natl. Acad. Sci. USA 2020, 117, 14202–14208. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.N.; Penade’s, J.R.; Chen, J. Genetic transduction by phages and chromosomal islands: The new and noncanonical. PLoS Pathog. 2019, 15, e1007878. [Google Scholar] [CrossRef] [Green Version]
- Rakhuba, D.V.; Kolomiets, E.I.; Dey, E.S.; Novik, G.I. Bacteriophage receptors, mechanisms of phage adsorption and penetration into host cell. Pol. J. Microbiol. 2010, 59, 145–155. [Google Scholar] [CrossRef]
- Young, F.E. Requirement of glucosylated teichoic acid for adsorption of phage in Bacillus subtilis 168. Proc. Natl. Acad. Sci. USA 1967, 58, 2377–2384. [Google Scholar] [CrossRef] [Green Version]
- Sao-Jose, C.; Baptista, C.; Santos, M.A. Bacillus subtilis operon encoding a membrane receptor for bacteriophage SPP1. J. Bacteriol. 2004, 186, 8337–8346. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Zhang, Y.; Liu, Z.; Dong, Z.; Xie, C.; Bravo, A.; Soberón, M.; Mahillon, J.; Sun, M.; Peng, D. The CRISPR-Cas systems were selectively inactivated during evolution of Bacillus cereus group for adaptation to diverse environments. ISME J. 2020, 14, 1479–1493. [Google Scholar] [CrossRef]
- Abedon, S.T. Lysis from without. Bacteriophage 2017, 1, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Dominy, B.N.; Perl, D.; Schmid, F.X.; Brooks, C.L. The effects of ionic strength on protein stability: The cold shock protein family. J. Mol. Biol. 2002, 319, 541–554. [Google Scholar] [CrossRef]
- Endersen, L.; O’Mahony, J.; Hill, C.; Ross, R.P.; McAuliffe, O.; Coffey, A. Phage therapy in the food industry. Annu. Rev. Food Sci. Technol. 2014, 5, 327–349. [Google Scholar] [CrossRef]
- El-Arabi, T.F.; Griffiths, M.W.; She, Y.M.; Villegas, A.; Lingohr, E.J.; Kropinski, A.M. Genome sequence and analysis of a broad-host range lytic bacteriophage that infects the Bacillus cereus group. Virol. J. 2013, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.; Bandara, N.; Shin, E.; Ryu, S.; Kim, K. Prevalence of Bacillus cereus bacteriophages in fermented foods and characterization of phage JBP901. Res. Microbiol. 2011, 162, 791–797. [Google Scholar] [CrossRef]
- Schachtele, C.F.; De Sain, C.V.; Hawley, L.A.; Anderson, D.L. Transcription during the development of bacteriophage φ29: Production of host- and φ29-specific ribonucleic acid. J. Virol. 1972, 10, 1170–1178. [Google Scholar] [CrossRef] [Green Version]
- Ahamed, S.K.; Roy, B.; Basu, U.; Dutta, S.; Ghosh, A.N.; Bandyopadhyay, B.; Giri, N. Genomic and proteomic characterizations of sfin-1, a novel lytic phage infecting multidrug-resistant Shigella spp. and Escherichia coli C. Front. Microbiol. 2019, 10, 1876. [Google Scholar] [CrossRef] [Green Version]
- Bull, H.B.; Breese, K. Interaction of alcohols with proteins. Biopolymers 1978, 17, 2121–2131. [Google Scholar] [CrossRef]
- Carlson, K. Appendix: Working with bacteriophages: Common techniques and methodological approaches. In Bacteriophages-Biology and Applications; Kutter, E., Sulakvelidze, A., Eds.; CRC Press: New York, NY, USA, 2005; pp. 437–494. [Google Scholar]
- Lauster, D.; Klenk, S.; Ludwig, K.; Nojoumi, S.; Behren, S.; Adam, L.; Stadtmüller, M.; Saenger, S.; Zimmler, S.; Hönzke, K.; et al. Phage capsid nanoparticles with defined ligand arrangement block influenza virus entry. Nat. Nanotechnol. 2020, 15, 373–379. [Google Scholar] [CrossRef]
- Hernando-Pérez, M.; Miranda, R.; Aznar, M.; Carrascosa, J.L.; Schaap, I.A.T.; Reguera, D.; de Pablo, P.J. Direct measurement of phage phi29 stiffness provides evidence of internal pressure. Small 2012, 8, 2366–2370. [Google Scholar] [CrossRef]
- Asakura, T.; Adachi, K.; Schwartz, E. Stabilizing effect of various organic solvents on protein. J. Biol. Chem. 1978, 253, 6423–6425. [Google Scholar] [PubMed]
- Herskovits, T.T.; Gadegbeku, B.; Jaillet, H. On the structural stability and solvent denaturation of proteins. J. Biol. Chem. 1970, 245, 2588–2598. [Google Scholar] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Strain Name | ST | Sensitivity a | No. | Strain Name | ST | Sensitivity |
|---|---|---|---|---|---|---|---|
| 1 | ATCC14579 | 921 | − | 40 | 3858-1B | 205 | − |
| 2 | 1582-3A | 4 | − | 41 | 3958 | 205 | − |
| 3 | 1582-3B | 4 | + | 42 | 3982-3A | 205 | − |
| 4 | 1608-3A | 4 | + | 43 | 4182-3C | 217 | − |
| 5 | 1608-3C | 4 | + | 44 | 2233-1 | 371 | − |
| 6 | 1958-1 | 4 | − | 45 | 2233-2 | 371 | − |
| 7 | 1983-1 | 4 | + | 46 | 2432-2 | 462 | − |
| 8 | 3108 | 4 | − | 47 | 3482-2C | 512 | − |
| 9 | 4032 | 4 | − | 48 | 3483 | 512 | − |
| 10 | 233-1 | 18 | − | 49 | 82 | 770 | − |
| 11 | 233-2 | 18 | − | 50 | 108-1 | 770 | − |
| 12 | 3332 | 24 | − | 51 | 208 | 770 | − |
| 13 | 3332-1A | 24 | − | 52 | 1058-2 | 770 | − |
| 14 | 3332-2A | 24 | − | 53 | 1682-2B | 770 | − |
| 15 | 791 | 26 | − | 54 | 2008-2 | 770 | − |
| 16 | 1058-1 | 26 | − | 55 | 3308-1A | 770 | − |
| 17 | 2132-4 | 26 | − | 56 | 3208 | 869 | − |
| 18 | 2583 | 26 | − | 57 | 3233 | 869 | − |
| 19 | 2932 | 26 | − | 58 | 2083-2 | 938 | − |
| 20 | 4182-1C | 26 | − | 59 | 2008-3 | 962 | − |
| 21 | 1782-2B | 32 | − | 60 | 2208 | 1001 | − |
| 22 | 2083-1 | 59 | − | 61 | 3808 | 1032 | − |
| 23 | 58 | 90 | − | 62 | 276-1C | 1084 | − |
| 24 | 232 | 90 | − | 63 | Y608 | 1225 | − |
| 25 | 1833-1C | 90 | − | 64 | 3008-1B | 1237 | − |
| 26 | 2833 | 90 | − | 65 | 3732 | 1327 | − |
| 27 | 3532 | 90 | − | 66 | 2833-2A | 1328 | − |
| 28 | 1682-3B | 92 | − | 67 | 4233 | 1329 | − |
| 29 | 1782-3A | 92 | − | 68 | 4233-1A | 1329 | − |
| 30 | Y1808 | 92 | − | 69 | 892-1 | 1330 | − |
| 31 | 2042-2 | 104 | − | 70 | 3283 | 1332 | − |
| 32 | 3232 | 144 | − | 71 | 1958-2 | 1418 | − |
| 33 | 3258-2A | 144 | − | 72 | 2132-1 | 1419 | − |
| 34 | 1682-2C | 158 | − | 73 | 1782-1A | 1435 | − |
| 35 | Y1683 | 164 | − | 74 | 3132-3A | 1439 | − |
| 36 | 1782-3B | 164 | − | 75 | 2833-1A | 1440 | − |
| 37 | Y641 | 172 | − | 76 | 2833-3A | 1440 | − |
| 38 | 1127 | 184 | − | 77 | 1558-2A | 1565 | − |
| 39 | 183 | 205 | − | 78 | 1558-3B | 1565 | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Yuan, X.; Li, N.; Wang, J.; Yu, S.; Zeng, H.; Zhang, J.; Wu, Q.; Ding, Y. Isolation and Characterization of Bacillus cereus Phage vB_BceP-DLc1 Reveals the Largest Member of the Φ29-Like Phages. Microorganisms 2020, 8, 1750. https://doi.org/10.3390/microorganisms8111750
Li C, Yuan X, Li N, Wang J, Yu S, Zeng H, Zhang J, Wu Q, Ding Y. Isolation and Characterization of Bacillus cereus Phage vB_BceP-DLc1 Reveals the Largest Member of the Φ29-Like Phages. Microorganisms. 2020; 8(11):1750. https://doi.org/10.3390/microorganisms8111750
Chicago/Turabian StyleLi, Chun, Xiaoming Yuan, Na Li, Juan Wang, Shubo Yu, Haiyan Zeng, Jumei Zhang, Qingping Wu, and Yu Ding. 2020. "Isolation and Characterization of Bacillus cereus Phage vB_BceP-DLc1 Reveals the Largest Member of the Φ29-Like Phages" Microorganisms 8, no. 11: 1750. https://doi.org/10.3390/microorganisms8111750