Mycobacterium bovis Infection in Red Foxes in Four Animal Tuberculosis Endemic Areas in France

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
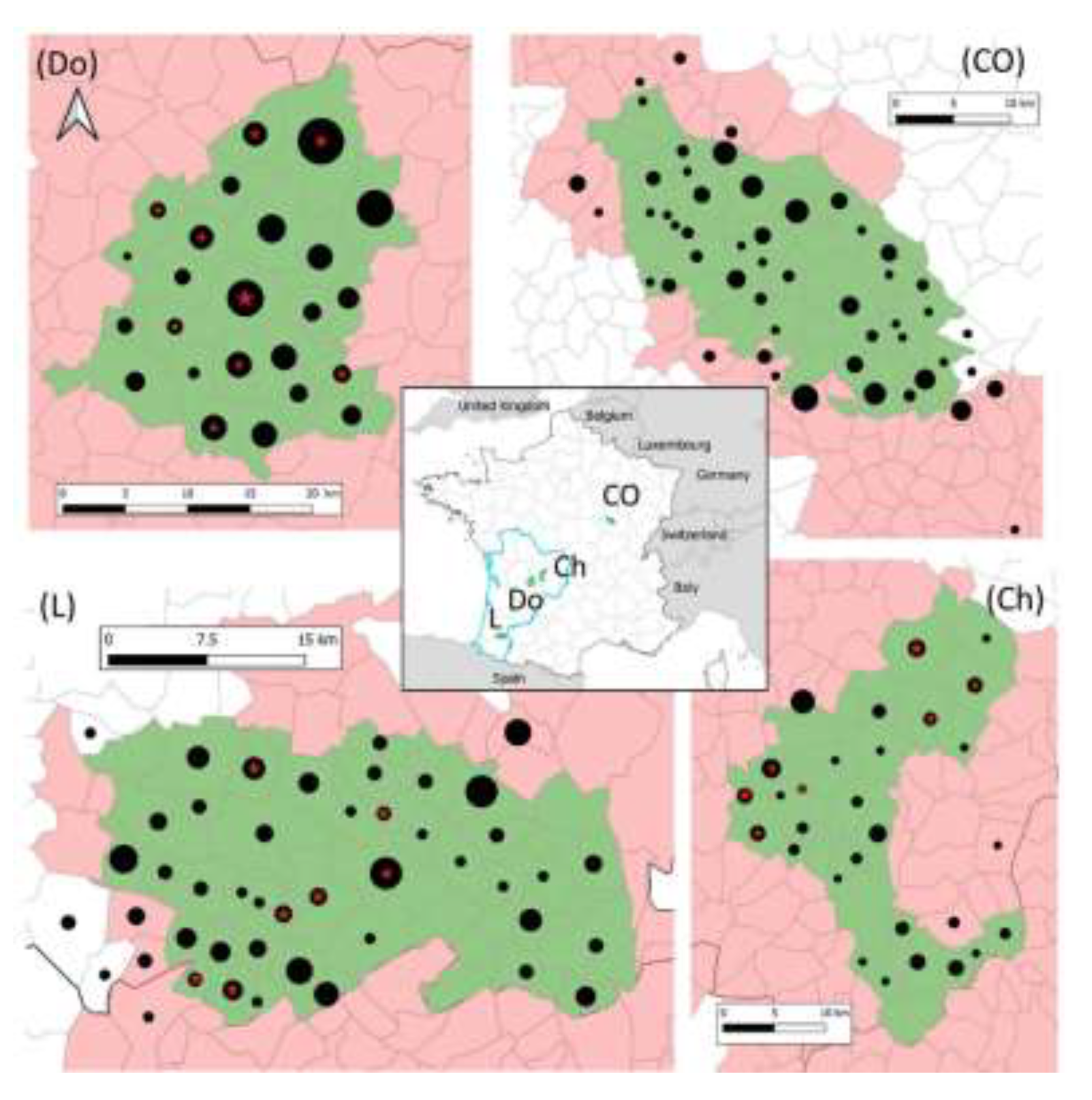
2.2. Study Areas
2.3. Fox Sampling and Sample Collection
2.4. Detection of M. bovis Infection
2.5. Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Santos, N.; Richomme, C.; Nunes, T.; Vicente, J.; Alves, P.C.; de la Fuente, J.; Correia-Neves, M.; Boschiroli, M.-L.; Delahay, R.; Gortázar, C. Quantification of the Animal Tuberculosis Multi-Host Community Offers Insights for Control. Pathogens 2020, 9, 421. [Google Scholar] [CrossRef] [PubMed]
- Haydon, D.T.; Cleaveland, S.; Taylor, L.H.; Laurenson, M.K. Identifying reservoirs of infection: A conceptual and practical challenge. Emerg. Infect. Dis. 2002, 8, 1468–1473. [Google Scholar] [CrossRef] [PubMed]
- The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, e05500. [CrossRef]
- Réveillaud, É.; Desvaux, S.; Boschiroli, M.-L.; Hars, J.; Faure, É.; Fediaevsky, A.; Cavalerie, L.; Chevalier, F.; Jabert, P.; Poliak, S.; et al. Infection of Wildlife by Mycobacterium bovis in France Assessment Through a National Surveillance System, Sylvatub. Front. Vet. Sci. 2018, 5, 262. [Google Scholar] [CrossRef] [PubMed]
- Delahay, R.J.; Smith, G.C.; Barlow, A.M.; Walker, N.; Harris, A.; Clifton-Hadley, R.S.; Cheeseman, C.L. Bovine tuberculosis infection in wild mammals in the South-West region of England: A survey of prevalence and a semi-quantitative assessment of the relative risks to cattle. Vet. J. 2007, 173, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Corner, L.A.L.; Murphy, D.; Gormley, E. Mycobacterium bovis infection in the Eurasian badger (Meles meles): The disease, pathogenesis, epidemiology and control. J. Comp. Pathol. 2011, 144, 1–24. [Google Scholar] [CrossRef]
- Matos, A.C.; Figueira, L.; Martins, M.H.; Pinto, M.L.; Matos, M.; Coelho, A.C. New Insights into Mycobacterium bovis Prevalence in Wild Mammals in Portugal. Transbound. Emerg. Dis. 2016, 63, e313–e322. [Google Scholar] [CrossRef]
- Michelet, L.; De Cruz, K.; Hénault, S.; Tambosco, J.; Richomme, C.; Réveillaud, É.; Gares, H.; Moyen, J.-L.; Boschiroli, M.L. Mycobacterium bovis Infection of Red Fox, France. Emerg. Infect. Dis. 2018, 24, 1150–1153. [Google Scholar] [CrossRef] [Green Version]
- Sergeant, E. Epitools Epidemiological Calculators; Ausvet. 2018. Available online: http://epitools.ausvet.com.au (accessed on 29 January 2020).
- Lesellier, S.; Boschiroli, M.-L.; Barrat, J.; Wanke, C.; Salguero, F.J.; Garcia-Jimenez, W.L.; Nunez, A.; Godinho, A.; Spiropoulos, J.; Palmer, S.; et al. Detection of live M. bovis BCG in tissues and IFN-γ responses in European badgers (Meles meles) vaccinated by oropharyngeal instillation or directly in the ileum. BMC Vet. Res. 2019, 15, 445. [Google Scholar] [CrossRef]
- Zhang, J.; Abadia, E.; Refregier, G.; Tafaj, S.; Boschiroli, M.L.; Guillard, B.; Andremont, A.; Ruimy, R.; Sola, C. Mycobacterium tuberculosis complex CRISPR genotyping: Improving efficiency, throughput and discriminative power of “spoligotyping” with new spacers and a microbead-based hybridization assay. J. Med. Microbiol. 2010, 59, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Hénault, S.; Karoui, C.; Boschiroli, M.L. A PCR-based method for tuberculosis detection in wildlife. Dev. Biol. (Basel) 2006, 126, 123–132; discussion 325–326. [Google Scholar]
- Thierry, D.; Brisson-Noël, A.; Vincent-Lévy-Frébault, V.; Nguyen, S.; Guesdon, J.L.; Gicquel, B. Characterization of a Mycobacterium tuberculosis insertion sequence, IS6110, and its application in diagnosis. J. Clin. Microbiol. 1990, 28, 2668–2673. [Google Scholar] [CrossRef] [Green Version]
- QGIS.org. QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: http://qgis.org (accessed on 29 January 2020).
- RStudio Team. RStudio: Integrated Development for R. RStudio, PBC, Boston, MA. Available online: http://www.rstudio.com/ (accessed on 29 January 2020).
- Martín-Atance, P.; Palomares, F.; González-Candela, M.; Revilla, E.; Cubero, M.J.; Calzada, J.; León-Vizcaíno, L. Bovine tuberculosis in a free ranging red fox (Vulpes vulpes) from Doñana National Park (Spain). J. Wildl. Dis. 2005, 41, 435–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millán, J.; Jiménez, M.A.; Viota, M.; Candela, M.G.; Peña, L.; León-Vizcaíno, L. Disseminated bovine tuberculosis in a wild red fox (Vulpes vulpes) in southern Spain. J. Wildl. Dis. 2008, 44, 701–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delahay, R.J.; De Leeuw, A.N.S.; Barlow, A.M.; Clifton-Hadley, R.S.; Cheeseman, C.L. The Status of Mycobacterium bovis Infection in UK Wild Mammals: A Review. Vet. J. 2002, 164, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.C.; Figueira, L.; Martins, M.H.; Matos, M.; Morais, M.; Dias, A.P.; Pinto, M.L.; Coelho, A.C. Disseminated Mycobacterium bovis Infection in Red Foxes (Vulpes vulpes) with Cerebral Involvement Found in Portugal. Vector Borne Zoonotic Dis. 2014, 14, 531–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsot, M.; Béral, M.; Scoizec, A.; Mathevon, Y.; Durand, B.; Courcoul, A. Herd-level risk factors for bovine tuberculosis in French cattle herds. Prev. Vet. Med. 2016, 131, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Richomme, C.; Boadella, M.; Courcoul, A.; Durand, B.; Drapeau, A.; Corde, Y.; Hars, J.; Payne, A.; Fediaevsky, A.; Boschiroli, M.L. Exposure of Wild Boar to Mycobacterium tuberculosis Complex in France since 2000 Is Consistent with the Distribution of Bovine Tuberculosis Outbreaks in Cattle. PLoS ONE 2013, 8, e77842. [Google Scholar] [CrossRef] [Green Version]
- Dell’Arte, G.L.; Laaksonen, T.; Norrdahl, K.; Korpimäki, E. Variation in the diet composition of a generalist predator, the red fox, in relation to season and density of main prey. Acta Oecologica 2007, 31, 276–281. [Google Scholar] [CrossRef]
- Macdonald, D.W. The Red Fox, Vulpes vulpes, as a Predator upon Earthworms, Lumbricus terrestris. Zeitschrift für Tierpsychologie 1980, 52, 171–200. [Google Scholar] [CrossRef]
- Payne, A.; Chappa, S.; Hars, J.; Dufour, B.; Gilot-Fromont, E. Wildlife visits to farm facilities assessed by camera traps in a bovine tuberculosis-infected area in France. Eur J. Wildl Res 2015, 62, 33–42. [Google Scholar] [CrossRef]
- Payne, A. Rôle de la faune sauvage dans le système multi-hôtes de Mycobacterium bovis et risque de transmission entre faune sauvage et bovins: Étude expérimentale en Côte d’Or. Ph.D. Thesis, Université Claude Bernard - Lyon I, Villeurbanne, France, 14 March 2014. [Google Scholar]
- Tolhurst, B.; Ward, A.I.; Delahay, R.J. A study of fox (Vulpes vulpes) visits to farm buildings in Southwest England and the implications for disease management. Eur. J. Wildl. Res. 2011, 57, 1227–1230. [Google Scholar] [CrossRef]
- Campbell, E.L.; Byrne, A.W.; Menzies, F.D.; McBride, K.R.; McCormick, C.M.; Scantlebury, M.; Reid, N. Interspecific visitation of cattle and badgers to fomites: A transmission risk for bovine tuberculosis? Ecol. Evol. 2019, 9, 8479–8489. [Google Scholar] [CrossRef] [PubMed]
- Tolhurst, B.A.; Delahay, R.J.; Walker, N.J.; Ward, A.I.; Roper, T.J. Behaviour of badgers (Meles meles) in farm buildings: Opportunities for the transmission of Mycobacterium bovis to cattle? Appl. Anim. Behav. Sci. 2009, 117, 103–113. [Google Scholar] [CrossRef]
- Payne, A.; Philipon, S.; Hars, J.; Dufour, B.; Gilot-Fromont, E. Wildlife interactions on baited places and waterholes in a French area infected by bovine tuberculosis. Front. Vet. Sci. 2016, 3, 122. [Google Scholar] [CrossRef] [Green Version]
- Robertson, A.; Delahay, R.J.; McDonald, R.A.; Aylett, P.; Henderson, R.; Gowtage, S.; Chambers, M.A.; Carter, S.P. Behaviour of European badgers and non-target species towards candidate baits for oral delivery of a tuberculosis vaccine. Prev. Vet. Med. 2016, 135, 95–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, A.; Ruette, S.; Schmidt, R.; Duhayer, J.; Colombe, M.; Lesellier, S.; Richomme, C.; Rossi, S. Premiers tests de distribution d’appâts en vue de la vaccination orale du blaireau contre la tuberculose bovine en France. Impact des espèces non cibles/ Impact of non-target species on a candidate baits deployment in the perspective of orally vaccinate badgers against bovine tuberculosis. Épidémiol. et santé anim. 2019, 75, 143–160. [Google Scholar]
- Gallagher, J.; Monies, R.; Gavier-Widen, M.; Rule, B. Role of infected, non-diseased badgers in the pathogenesis of tuberculosis in the badger. Vet. Rec. 1998, 142, 710–714. [Google Scholar] [CrossRef]
- Gallagher, M.J.; Clifton-Hadley, R.S. Tuberculosis in badgers; a review of the disease and its significance for other animals. Res. Vet. Sci. 2000, 69, 203–217. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Region | Nouvelle-Aquitaine | Burgundy – Franche-Comté | ||
|---|---|---|---|---|
| Department | Dordogne (Do) | Charente (Ch) | Landes (L) | Côte-d’Or (CO) |
| Surface of the area (in km2) | 525 | 539 | 504 | 484 |
| Number of cattle outbreaks | 35 | 13 | 31 | 22 |
| Mean prevalence in badger (# infected/# analyzed) | 11.1% (15/135) | 9.0% (29/321) | 13.2% (20/152) | 6.2% (27/432) |
| Mean prevalence in wild boar (# infected/# analyzed) | 4.3% (8/185) | 7.8% (20/255) | 17.9% (7/39) | 2.2% (6/269) |
| Number of cases in wild deer | 2 | 1 | 0 | 2 |
| Region | Department | Sampling Collection | Tissues Used for M. bovis Detection | Data Analysis | ||||
|---|---|---|---|---|---|---|---|---|
| Apparent Prevalence | Exposure Routes | Excretion Routes | Fox Infection/Cattle Outbreak Pastures Spatial Relationship | |||||
| Burgundy – Franche-Comté | Côte-d’Or | June 2018 – November 2019 | Pool (RP LN + resp. LN), mes. LN | x | x | - | - | |
| Nouvelle-Aquitaine | Charente | x | x | - | - | |||
| Landes | x | x | - | |||||
| Dordogne | Season #1 | March 2017 to August 2018 | Pool (RP LN + resp. LN), mes. LN, feces 1, urine 1,2, oropharyngeal swab 1 | x | x | x | x | |
| Season #2 | August 2018 to November 2019 | Separately RP LN, resp. LN, mes. LN | - | x | - | - | ||
| Number of foxes | Prevalence (%) | 95% Confidence Interval | ||
|---|---|---|---|---|
| Analyzed | Infected | |||
| Charente | 98 | 9 | 9.2 | 4.3–16.7 |
| Dordogne (season #1) | 184 | 13 | 7.1 | 3.8–11.8 |
| Landes | 140 | 7 | 5.0 | 2.0–10.0 |
| Nouvelle-Aquitaine * | 422 | 29 | 6.9 | 4.6–9.7 |
| Côte-d’Or | 146 | 0 | - | 0–2.5 |
| M. bovis Detection | |||
|---|---|---|---|
| Tissues Analyzed | Total Analyzed | Culture Positive | PCR Positive |
| LN mesenteric | 41 | 4 | 28 |
| LN RP + resp pooled | 29 | 2 | 26 |
| LN RP separately | 12 | 3 | 1 |
| LN resp separately | 12 | 1 | 8 |
| Feces | 12 | 0 | 5 (1) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richomme, C.; Réveillaud, E.; Moyen, J.-L.; Sabatier, P.; De Cruz, K.; Michelet, L.; Boschiroli, M.L. Mycobacterium bovis Infection in Red Foxes in Four Animal Tuberculosis Endemic Areas in France. Microorganisms 2020, 8, 1070. https://doi.org/10.3390/microorganisms8071070
Richomme C, Réveillaud E, Moyen J-L, Sabatier P, De Cruz K, Michelet L, Boschiroli ML. Mycobacterium bovis Infection in Red Foxes in Four Animal Tuberculosis Endemic Areas in France. Microorganisms. 2020; 8(7):1070. https://doi.org/10.3390/microorganisms8071070
Chicago/Turabian StyleRichomme, Céline, Edouard Réveillaud, Jean-Louis Moyen, Perrine Sabatier, Krystel De Cruz, Lorraine Michelet, and Maria Laura Boschiroli. 2020. "Mycobacterium bovis Infection in Red Foxes in Four Animal Tuberculosis Endemic Areas in France" Microorganisms 8, no. 7: 1070. https://doi.org/10.3390/microorganisms8071070