Candida glabrata: A Lot More Than Meets the Eye


1
Laboratory of Fungal Pathogenesis, Centre for DNA Fingerprinting and Diagnostics, Hyderabad 500039, India
2
Graduate Studies, Manipal Academy of Higher Education, Manipal 576104, India
3
Graduate Studies, Regional Centre for Biotechnology, Faridabad 121001, India
*
Author to whom correspondence should be addressed.
Microorganisms 2019, 7(2), 39; https://doi.org/10.3390/microorganisms7020039
Submission received: 19 December 2018
/
Revised: 21 January 2019
/
Accepted: 29 January 2019
/
Published: 30 January 2019
(This article belongs to the Special Issue Molecular Mechanisms of Fungal Virulence and Commensalism)
Abstract
:Candida glabrata is an opportunistic human fungal pathogen that causes superficial mucosal and life-threatening bloodstream infections in individuals with a compromised immune system. Evolutionarily, it is closer to the non-pathogenic yeast Saccharomyces cerevisiae than to the most prevalent Candida bloodstream pathogen, C. albicans. C. glabrata is a haploid budding yeast that predominantly reproduces clonally. In this review, we summarize interactions of C. glabrata with the host immune, epithelial and endothelial cells, and the ingenious strategies it deploys to acquire iron and phosphate from the external environment. We outline various attributes including cell surface-associated adhesins and aspartyl proteases, biofilm formation and stress response mechanisms, that contribute to the virulence of C. glabrata. We further discuss how, C. glabrata, despite lacking morphological switching and secreted proteolytic activity, is able to disarm macrophage, dampen the host inflammatory immune response and replicate intracellularly.
1. Introduction
Candida glabrata is an opportunistic human fungal pathogen that accounts for up to 29% of total Candida bloodstream infections [1,2]. Its prevalence varies with the geographic area [2,3,4]. While C. glabrata is the second most common bloodstream Candida species after C. albicans in Northern Europe and the USA [1,5,6], it ranks as the third or fourth most prevalent invasive Candida pathogen in Asia [4,6,7]. Recent epidemiological surveys have shown a decrease in the frequency of C. albicans and an increased prevalence of non-albicans Candida [3,5,6]. C. glabrata bloodstream infections are commonly found in elderly individuals, diabetic patients and solid organ transplant recipients [6,8,9]. C. glabrata is also a causative agent of urinary tract and vaginal infections [10,11].
Historically, C. glabrata was named Cryptococcus glabrata, and this name was first changed to Torulopsis glabrata and later to Candida glabrata. Since pseudohyphae formation was not found to be a dependable criterion to classify yeasts at the genus level, the Torulopsis and Candida genera were merged under the Candida genus name [10,12]. However, based on molecular evolutionary studies, C. glabrata was later assigned to the genus Nakaseomyces [13]. The current taxonomy of C. glabrata is the Kingdom Fungi, Subkingdom Dikarya, Phlyum Ascomycota, Subphylum Saccharomycotina, Class Saccharomycetes, Order Saccharomycetales, Family Saccharomycetaceae, Genus Nakaseomyces, Clade Nakaseomyces/Candida and Species glabrata (NCBI:txid284593).
Clinically, C. glabrata is primarily diagnosed via culture-based assays viz., colony color (white/pink/purple) on CHROMagar Candida medium and microscopic examination [presence of small-sized (1–4 μm) yeast cells and lack of hyphal structures], and biochemical methods, viz., assimilation of glucose and trehalose sugars [14]. Additionally, the FDA (Food and Drug Administration, USA)-approved T2 Candida panel, which can identify five Candida species, C. albicans, C. glabrata, C. tropicalis, C. parapsilosis and C. krusei, is increasingly being used for rapid Candida species detection in hospital settings [15,16]. In the current review, we summarize the genomic configuration, virulence traits and nutrient acquisition and stress signaling pathways in C. glabrata. Additionally, we provide an overview of C. glabrata-host cell interaction mechanisms.
2. Genomic Architecture
C. glabrata is a haploid budding yeast, and belongs to the non-CTG, Saccharomycetaceae clade in which the CTG codon encodes leucine [17,18]. Contrarily, C. albicans belongs to the CTG clade wherein the CTG codon is translated as serine instead of leucine [17,18]. The genome of the C. glabrata CBS138 strain was sequenced by the Genolevures Consortium using whole genome shotgun sequencing and represented by 1000 contigs [17]. The annotated genome contains 13 chromosomes, named as Chromosome A to M, and is of 12.3 Mb in size [17]. Of a total of 5293 open reading frames (ORFs) in the C. glabrata genome, only 238 (4.5%) ORFs are verified with some experimental evidence for the existence of their gene products [www.candidagenome.org]. The chromosome length varies from 4,91,328 to 14,55,689 bp with the chromosome A and L being the smallest and largest, respectively [www.candidagenome.org]. The C. glabrata genome shows plasticity, and alterations in karyotype and chromosome size have been observed in clinical isolates and the reference strain CBS138 [19,20,21].
The C. glabrata genome displays robust synteny with the genome of the non-pathogenic yeast Saccharomyces cerevisiae, and synteny blocks were found to be present in 88% of the genome of these two yeasts [17,22]. Consistently, C. glabrata contains orthologs of 4870 S. cerevisiae genes [https://yeastmine.yeastgenome.org/], however, it also possesses a set of 337 genes that are absent in S. cerevisiae [23]. Both C. glabrata and S. cerevisiae belong to the Whole Genome Duplication (WGD) group, and are assumed to have arisen from the same tetraploid hybrid ancestor, which existed about 100-200 million years ago [18]. This ancestor is thought to have undergone extensive whole genome duplication brought about by an interspecies hybridization event, which helped the hybrid to regain fertility [24]. Furthermore, compared to the S. cerevisiae lineage, a greater degree of gene loss has occurred in the C. glabrata lineage [17,18]. The C. glabrata genome consistently exhibits lower global redundancy [17].
Although the reductive genome evolution was thought to be linked with the pathogenic life style of C. glabrata with more reliance on the human host for nutrients [17], recent studies do not support this notion [18,25]. As mentioned earlier, within the Saccharomycetaceae family, C. glabrata belongs to the genus Nakaseomyces, which contains three pathogenic species (C. glabrata, C. nivariensis and C. bracarenses) and three environmental species (C. castellii, Kluyveromyces delphensis and K. bacillisporus) [18]. Of these, C. castellii and K. bacillisporus belong to one group, while the remaining four are represented by a group, referred to as the ‘glabrata group’ [18]. Intriguingly, molecular phylogenetic and genome analysis have revealed three human pathogenic species, C. glabrata, C. nivariensis and C. bracarenses of the glabrata group, to be polyphyletic in origin [18,25].
C. glabrata contains a small 20 kb circular mitochondrial genome which contains eleven ORFs, including genes coding for three subunits of the cytochrome C oxidase (CgCox1, 2 and 3), the apocytochrome b (CgCob) and three subunits of the ATP synthase (CgAtp6, 8 and 9) [26], (www.candidagenome.org). It also contains 23 tRNAs, 2 rRNAs and 1 non-coding RNA (www.candidagenome.org).
The reproduction mode in C. glabrata is predominantly clonal, and the opportunistic pathogenic lifestyle of C. glabrata is also thought to have arisen independently from that of C. albicans [25]. As discussed above, C. glabrata belongs to the post-WGD non-CTG clade and appears to be evolutionarily uniquely placed. Phylogenetically, it is closer to S. cerevisiae and only distantly related to the most common pathogenic fungus, C. albicans [17,18]. Furthermore, Nakaseomyces is the only group that possesses the ability to infect humans among the post-WGD clade, and, recently identified C. bracarensis and C. nivariensis are the pathogenic close relatives of C. glabrata [18]. As the capability to infect humans appears to have emerged independently within the Nakaseomyces [18], C. glabrata may have acquired a unique set of pathogenesis attributes. In this review, we discuss the unique features of C. glabrata, as well as the characteristics that it shares with the non-pathogenic yeast S. cerevisiae and the pathogenic yeast C. albicans. Of note, virulence mechanisms of C. bracarensis and C. nivariensis are yet to be identified. Table 1 summarizes the key characteristic features of C. glabrata and C. albicans.
3. Salient Pathobiological Features
The traits that may contribute to the virulence of C. glabrata, are discussed below.
3.1. Adherence
Adherence to the host tissue is an important trait that contributes to colonization and establishment of successful infections. C. glabrata possesses a total of 67 putative adhesins, which may mediate adherence to host cells [27]. These putative glycosylphosphatidylinositol (GPI)-anchored cell wall proteins are composed of a N-terminal ligand-binding domain and a low complexity serine/threonine-rich region with internal tandem repeats followed by a C-terminus GPI-anchor attachment site [27]. Based on their putative ligand-binding regions, the adhesin gene family has been classified into seven sub-families [27]. The Epa (Epithelial Adhesin) sub-family I, containing the PA14 (Anthrax Protective Antigen) ligand-binding domain, is most well studied, and consists of 17 to 23 proteins depending upon the isolate [27,28]. The sequenced fecal isolate CBS 138 contains 17 Epa-encoding genes, while the widely used vaginal isolate BG2 contains 23 Epa-encoding genes [27,28]. The majority of Epa adhesins are encoded by sub-telomeric localized genes and regulated by epigenetic SIR (Silent Information Regulator)-dependent transcriptional silencing [27,29,30,31].
The founding member of the Epa adhesin family, Epa1, is a calcium-dependent lectin, and aids adhesion to epithelial cells [32] and macrophages [33]. The multidrug resistance transcription factor CgPdr1 has been implicated in regulation of the EPA1 gene expression [34]. The Epa6 and Epa7 adhesins have been shown to mediate adherence to epithelial and endothelial cells [30,35]. Furthermore, EPA6 was found to be expressed in the murine urinary tract infection model, due to unavailability of the nicotinic acid, which is a precursor for the CgSir2 histone deacetylase cofactor, nicotinamide adenine dinucleotide (NAD+) [30]. Similarly, differential expression of EPA2, EPA3, EPA7 and EPA22 genes has been reported in response to different environmental cues [36,37,38,39,40,41,42]. Recent analysis has linked expansion of the EPA gene family with virulence of the fungal species of the Nakaseomyces clade, with C. glabrata, C. bracarensis, C. nivariensis and K. delphensis containing 17–23, 12, 9 and 1 Epa adhesins, respectively [18].
With regard to ligands, Epa1, Epa6 and Epa7 were found to bind to oligosaccharides containing a terminal galactose residue [35]. Despite the preference for the terminal galactose, Epa1, 6 and 7 showed glycan ligand specificity, with Epa6 having the widest substrate specificity [35,43]. In accordance, a recent study has predicted many human receptors including mucins, CD43 (leukosialin)/CD45 (receptor-type tyrosine-protein phosphatase) glycoproteins, ceruloplasmin, (sero)transferrin, and fibronectin, for Epa1, 6 and 7 [44]. Furthermore, adherence analysis of S. cerevisiae strains expressing ligand-binding domains of 15 Epa adhesins individually revealed significant, moderate and very weak adherence to human epithelial colorectal adenocarcinoma cells (Caco2) for Epa1, 6 and 7, Epa8, 9, 12, 15 and 23, and Epa2, 3, 11, 13, 19, 20 and 21, respectively [45].
The second adhesin sub-family contains a N-terminal PA14 domain, referred to as the Pwp family (PA14-containing Wall Protein), and contains seven proteins. Of these Pwp1-7, the Pwp7 protein is required for adherence to human endothelial cells in vitro [23]. The number of adhesins in sub-families III, IV, V, VI and VII vary from 3 to 13, and are yet to be characterized for their ligands and functions [27]. Two adhesins, CAGL0L09911p and CAGL0J05159p/CAGL0J05170p, did not belong to any of the subfamily, and were grouped separately [27]. Many adhesins including Epa proteins contained 46-amino acids repeats (Awp2 repeats) in their C-terminus low-complexity regions [27]. One such protein, Aed1 (Adherence to Endothelial cells), belonging to the sub-family III, has been implicated in adherence to human endothelial cells [23]. Of note, a recent study has identified 49 novel protein-coding genes, of which eight are located in close proximity to EPA or PWP genes in the subtelomeric regions [46].
Importantly, many adhesin-encoding genes contain several kilobases of 126–429 bp-long sequences, that are tandemly repeated up to 32-times, called megasatellites [47]. The number of repeats in these mega-satellites may govern the length and functions of adhesions [47]. Moreover, C. glabrata clinical isolates are known to have varied number of adhesin-encoding genes, and a distinct profile of cell wall proteins expressed at the cell surface [41,48,49]. Therefore, the presence of such a large number of adhesin genes and their complex environment-dependent differential regulation are likely to help C. glabrata colonize different host niches as well as to form biofilms on a wide range of surfaces [27,47,48].
3.2. Biofilm Formation
Biofilms are complex extracellular matrix-embedded, multi-layered microbial structures on biotic or abiotic surfaces which are formed by microbe-microbe and microbe-surface interactions [50]. C. glabrata biofilms display antifungal resistance, and are characterized by a compact dense structure of yeast cells nested in an extracellular matrix which is composed of high levels of proteins and carbohydrates including β-1,3 glucan [50,51,52]. Large scale and candidate gene-based studies have identified several genes that are pivotal to biofilm formation in C. glabrata [36,37,50,53,54]. Among these, the adhesin encoded by the EPA6 gene, which is regulated by multiple factors including the CgYak1p kinase, subtelomeric silencing, chromatin remodeling Swi/Snf complex components and the transcriptional factor CgCst6, plays a central role [36,37,53]. Additionally, other adhesins, cell wall proteins and RNA polymerase II mediator complex subunits including Epa3, Epa7, Epa12, Awp4–6, Pwp1, Pwp3, Med12, Med13 and Med15 have also been implicated in biofilm formation [36,37,40,53,54]. Gene expression profiling analysis has revealed differential expression of adhesin-encoding genes in biofilms formed under in vitro and in vivo conditions [55]. Furthermore, increased and decreased levels of stress proteins and glycolytic enzymes, respectively, have been reported during the biofilm mode of growth [56]. The ability of C. glabrata to form biofilms has also been found to be enhanced in the high-iron environment [57]. Moreover, recent studies have shown a close association between the capacity to form biofilms and colonize murine organs in the systemic candidiasis model [57,58], however, the precise mechanisms underlying this relationship are unknown.
3.3. Aspartyl Proteases
Secreted aspartyl proteases are key fungal virulence factors [28]. However, despite the lack of any secreted proteolytic activity [59], the C. glabrata genome contains eleven genes (CgYPS1-11) that code for a family of eleven putative GPI-anchored aspartyl proteases, known as Yapsins [60]. CgYPS genes have been shown to be regulated by various environmental conditions including low pH, thermal stress, macrophage and neutrophil internalization [60,61,62,63]. Consistently, CgYapsins are pivotal to the regulation of several cellular processes such as maintenance of the cell wall architecture, pH and vacuole homeostasis, biofilm formation and interaction with the host [58,60,61,64]. The latter includes suppression of the production of pro-inflammatory cytokine IL-1β in human THP-1 macrophages, facilitating survival in macrophages and virulence in the models of murine systemic candidiasis and Drosophila melanogaster [58,60,65,66].
Of eleven CgYapsins, CgYps1 is uniquely required to survive acid stress, as CgYPS1 deletion led to low intracellular pH, high ROS (reactive oxygen species) levels and cell death under low pH and acidic environmental conditions [61]. This specific role of CgYPS1 in intracellular pH homeostasis has been attributed, in part, to regulation of the ATPase activity of the plasma membrane proton pump, CgPma1 [61]. Additionally, the mutant lacking eleven yapsins was found to display large acidic vacuole, elevated metal ion susceptibility, mis-sorting of the vacuole hydrolase carboxypeptidase Y (CPY), diminished vacuolar ATPase activity and perturbed polyphosphate and energy homeostasis (Figure 1) [64]. Furthermore, infection with the Cgyps11 mutant, generated by the CRISPR-Cas system, resulted in slower death of fruit flies lacking adapter of the Toll signaling pathway MyD88 [67]. Lastly, the Cgyps1-11Δ mutant displayed sunken cell walls containing higher chitin and reduced β-glucan and mannan levels (Figure 1) [58,64]. CgYapsins have also been implicated in shedding Epa1 adhesin off the cell wall, as Epa1 release into the medium was drastically reduced in the Cgyps1-11Δ mutant compared to wild-type cells (Figure 1) [60]. Although CgYapsins rank amongst the major virulence factors of C. glabrata, a direct link between the yapsin enzyme activity and different cellular processes is yet to be established.
3.4. Colony and Mating-Type Switching
Phenotypic plasticity is the ability of a given genotype to produce different phenotypes in varied environmental conditions [10]. C. glabrata is known to exhibit four different-colored colony types, white, light brown, dark brown and very dark brown, in the presence of copper sulfate or phloxine B [68]. Switching between these phenotypes, that was found to be common, spontaneous and reversible [68], had also been shown to occur at sites of colonization in vaginitis patients [69]. Moreover, C. glabrata is also known to undergo spontaneous reversible switching from a regular to an irregular wrinkled colony type [68]. Importantly, morphological switching from the yeast to the hyphal form has not been reported [28], although C. glabrata displayed pseudohyphal structures in response to nitrogen starvation and carbon dioxide exposure [70,71]. Furthermore, disruption of the transcription factor CgAce2 is known to lead to cell aggregation, due to defects in cell separation, and hypervirulence [72,73].
C. glabrata has three mating type (MAT)-like loci, CgMTL1 (MAT), CgMTL2 (HMR) and CgMTL3 (HML), whose genomic configuration is similar to that of the S. cerevisae MTL loci [74]. Contrarily, C. albicans contains a single MTL locus that regulates mating and cell type (Figure 2) [75]. The MAT locus in S. cerevisiae, consisting of the active MAT locus, and silent HMLα and HMRa loci, resides on the chromosome III (Figure 2), and the mating type is determined by the presence of MATa or MATα allele [75]. However, unlike S. cerevisiae, the MAT loci are present on different chromosomes in C. glabrata with CgMTL1 and CgMTL3 on chromosome B, and CgMTL2 on chromosome E (Figure 2) [76]. CgMTL1 locus encodes either ‘a’ (Cga1gene) or ‘alpha’ (Cgα1 or Cgα2 genes) information while CgMTL2 and CgMTL3 code for ‘a’ and ‘α’ information, respectively [74,76]. While CgMTL1 and CgMTL2 are transcriptionally active, CgMTL3 is subjected to subtelomeric silencing [77]. Despite having two mating types, a and α, C. glabrata lacks spontaneous mating-type switching. However, ectopic expression of the S. cerevisiae endonuclease-encoding HO gene is known to result in efficient mating-type switching and lethality in C. glabrata [78]. Mating-type switching has also been reported at sites of colonization in vaginitis patients [69]. Of note, genomic recombination between different clades and evidence for an active sexual cycle have recently been reported in C. glabrata [25]. Altogether, the above-mentioned switching mechanisms may generate phenotypic diversity that is likely to assist C. glabrata in adapting to different environmental conditions.
3.5. Stress Response Mechanisms
C. glabrata is highly tolerant to oxidative, cell wall, osmotic and endoplasmic reticulum (ER) stress [79]. The environmental stress response in C. glabrata is governed by two transcriptional regulators CgMsn2 and CgMsn4 [38], while the oxidative stress response is regulated by the sole catalase CgCta1 [80], two superoxide dismutases CgSod1 and CgSod2 [81], and glutathione biosynthetic enzymes CgGsh1 and CgGsh2 [82,83]. CgCTA1 expression is regulated by several stress response transcriptional regulators including CgYap1, CgMsn2, CgMsn4 and CgSkn7 [80,84,85,86]. Additionally, lack of the histone deacetylase CgHst1 led to high expression of CgCTA1 and elevated resistance to oxidative stress, which was dependent upon CgMsn4 [87]. Despite an essential role of CgCta1 in resistance to the hydrogen peroxide-generated oxidative stress in vitro, the Cgcta1Δ mutant was not attenuated for virulence in the murine disseminated candidiasis model which suggests the existence of a CgCta1-independent resistance mechanism in vivo [80]. Recently, Kounatidis et al. have identified the transcriptional adapter of the Spt-Ada-Gcn5 acetyltransferase (SAGA) complex, CgAda2, by screening a library of 196 transcription factor mutants, to be essential for ROS survival in the Drosophila larvae model [88]. Furthermore, tryptophan-based pigment production is also known to aid in survival of the ROS stress [89].
The protein kinase C (PKC)-mediated signaling pathway safeguards the cell wall in C. glabrata, and its terminal mitogen-activated protein kinase (MAPK) CgSlt2, along with other cascade components, is required for survival of the cell wall stress [54,90,91]. Intriguingly, the PKC pathway is also regulated by the ER stress [92]. In addition, components of the calcineurin signaling have also been found to be essential for maintenance of the cell wall integrity [93,94]. Calcineurin signaling is also known to play a pivotal role in the virulence of C. glabrata [93,94]. The response of C. glabrata to osmotic stress is yet to be characterized in detail. However, the high osmolarity glycerol pathway [HOG] is known to be activated by sorbic acid, and its terminal MAPK CgHog1 is required for survival of weak acid and osmotic stress [95,96]. For survival of the ER stress, essentiality of the key ER stress sensor, CgIre1 endoribonuclease, was attributed to its role in activation of the non-canonical unfolded protein response pathway [97]. Furthermore, lack of CgIre1 led to diminished virulence in both immunosuppressed and immunocompetent mice [97]. Contrarily, deletion of the transcriptional co-activator CgAda2 conferred resistance to the ER stressor, tunicamycin, and hypervirulence in the immunocompromised mice [98]. Besides Ire1 signaling, two other stress-responsive pathways, PKC-mediated cell wall integrity and calcineurin signaling, have also been implicated in regulating the transcriptional response to the ER stress [94,97]. Both these signaling cascades are also required to survive the azole and echinocandin antifungal stress [54,91,99,100].
Of three common antifungal drugs, polyenes, azoles and echinocandins, for treatment of systemic Candida infections, C. glabrata is intrinsically less susceptible to azoles which impede ergosterol biosynthesis by inhibiting the cytochrome P450-dependent lanosterol 14α-demethylase enzyme [101]. Recently, a substantial number of azole-resistant C. glabrata isolates have also been found to be resistant to cell-wall targeting drugs echinocandins, which inhibit the β-glucan synthase enzyme [101,102,103]. Moreover, due to renal toxicity, the use of polyene antifungals, which bind to ergosterol in the plasma membrane and disrupt cell membrane functions, is largely limited [101]. Hence, several studies have been conducted to advance our understanding of the signaling pathways that C. glabrata utilizes to cope/counteract antifungal stress. The PKC-mediated cell wall integrity pathway has been shown to be required for the transcriptional activation of multidrug efflux pumps, which is the most frequent azole resistance mechanism in clinical settings world-wide [91,101,104,105]. Other azole resistance mechanisms include mitochondrial dysfunction and overexpression of the sterol biosynthetic target enzyme [28,101]. Resistance towards echinocandins is primarily due to mutations in the CgFKS1 and CgFKS2 genes that code for β-1,3-glucan synthase [103]. Both calcineurin and PKC signaling have been implicated in echinocandin resistance in C. glabrata [93,103,106]. Polyene resistance is less frequent in C. glabrata, and mutations in genes encoding an ergosterol biosynthetic enzyme Erg6 and components of the mismatch repair pathway have been associated with polyene resistance [103,107,108,109]. Because of space limitations, the factors, which confer tolerance and/or resistance to antifungal drugs in C. glabrata are not discussed here; readers may refer to recent reviews on this topic [101,103].
Overall, the high resistance to diverse stressful conditions may significantly contribute to the survival of C. glabrata in varied host niches.
3.6. Nutrient Acquisition Pathways
A prerequisite to thrive in the host is the ability to acquire nutrition from the nutrient-limited host environment. C. glabrata has rewired its metabolic machinery, and developed novel nutrient uptake mechanisms. It has lost 5, 4, 3, 3 and 6 genes involved in galactose metabolism, phosphate metabolism, cell rescue, defense and virulence, nitrogen and sulfur metabolism, and allantoin catabolism, respectively [17]. C. glabrata also lacks genes involved in thiamine, pyridoxine and nicotinic acid biosynthetic pathways [28]. The loss of nicotinic acid synthesis genes was thought to be due to close association of C. glabrata with the mammalian host, however, recent studies report that the nicotinic acid auxotrophy is a trait of the Nakaseomyces clade, which contains both environmental and pathogenic species [18].
3.6.1. Carbon Acquisition
C. glabrata is a facultatively anaerobic, crabtree-positive yeast which prefers fermentation over respiration in the presence of oxygen [110,111]. C. glabrata can assimilate both glucose and trehalose, but lacks genes for assimilation of galactose and sucrose [17,28]. Importantly, C. glabrata can utilize l-amino acids such as glutamate, aspartate and proline as the sole carbon and nitrogen source [112]. C. glabrata is also able to grow in very low concentration of glucose, and its glucose sensing pathway resembles to that of S. cerevisiae [113]. The C. glabrata genome encodes eleven hexose transporters, two transcriptional regulators CgRgt1 and CgMig1, and two glucose sensors, CgRgt2 and CgSnf3. CgSnf3 acts as the high-affinity glucose sensor which is essential for growth under glucose-limited conditions and in macrophages [113]. C. glabrata also has a set of six conserved duplicated gene paralogs encoding glycolytic enzymes, Eno1/Eno2, Pyc1/Pyc2, Glk1/Emi2, Hxk1/Hxk2, Tdh2/Tdh3 and Cdc19/Pyk2 [18], that may aid in the increased carbon flux through glycolysis, and contribute to its respiro-fermentative lifestyle [114].
3.6.2. Phosphate Acquisition
The macronutrient phosphorus, in the form of inorganic phosphate, is pivotal to the biosynthesis of cellular moieties and regulation of various metabolic processes. The PHO (phosphate signal transduction) pathway executes the phosphate starvation response in C. glabrata via the transcription factor CgPho4, which itself is regulated by the cyclin (CgPho80)/cyclin-dependent protein kinase (CDK; CgPho85)/CDK inhibitor (CgPho81) complex [115]. However, the C. glabrata genome lacks an ortholog of the S. cerevisiae PHO5 gene which codes for a phosphate starvation-inducible acid phosphatase [115]. Instead, C. glabrata genome uniquely codes for a family of three phosphatases, CgPmu1-3 (C. glabrata phosphomutase-like protein 1–3), which possess different substrate specificity [116]. CgPho4 was found to be required for the phosphate starvation-induced phosphatase activity, and deletion of the putative CgPho4 nuclear exporter, CgMsn5, led to increased phosphatase activity under phosphate-surplus conditions [115]. Of three CgPmu proteins, secreted phosphatase activity of only CgPmu2 was induced in response to phosphate starvation [116]. Consistent with this, CgPMU2 transcript levels were lower in high-phosphate conditions and substantially higher during phosphate starvation [116]. Furthermore, CgPMU2 and CgPHO gene expression was found to be regulated by CgPho4 in a largely CgPho2 coactivator-independent manner [115,117]. This reduced dependence on CgPho2 appears to have substantially broadened the target gene set of CgPho4, which included genes involved in phosphate homeostasis as well as adherence, cell wall biosynthesis, non-phosphate-related stress response and carbohydrate metabolism [118]. Recently, CgPmu3 has been shown to be a thiamine phosphatase that is transcriptionally regulated by the transcription factor CgThi3 in response to thiamine starvation [119]. Overall, C. glabrata appears to have neofunctionalized the CgPmu family to compensate for the loss of the ancestral Pho5 phosphatase [116,119]. Although CgPmu2 was required to grow in the presence of organic phosphate compounds as the sole phosphate source in vitro [116], it remains to be determined whether neofunctionalization of the CgPMU gene family confers a growth advantage in the mammalian host.
3.6.3. Iron Acquisition
Of known fungal iron uptake mechanisms, siderophore-mediated uptake of Fe3+, reductive iron acquisition, haemoglobin/haem uptake [120], all systems are operational in C. glabrata except for the receptor-mediated haem uptake [121,122]. The sole xenosiderophore transporter CgSit1 typifies the siderophore-mediated iron uptake system in C. glabrata [123]. CgSit1 has been shown to be pivotal to survival in the iron-limited host environment [123]. Although C. glabrata exhibits haemolytic activity in vitro and possesses hemolysins, it lacks the haem receptor [121,122,124]. C. glabrata is also unable to utilize the host iron proteins, haemoglobin, and transferrin, as iron sources [125]. Recently, putative cell surface-associated, cysteine-rich Common in Fungal Extracellular Membrane (CFEM) domain-containing protein (CgCcw14), haem oxygenase (CgHmx1) and intracellular iron trafficking machinery components including vacuolar (CgCcc1, CgSmf1, CgSmf3, and CgFth1) and mitochondrial (CgMmt1, CgMmt2, CgAtm1, CgMrs3, and CgMrs4) iron transporters have been identified in C. glabrata [122,125], however, their role in iron metabolism is yet to be fully characterized.
The reductive iron acquisition system in C. glabrata consists of three ferric reductases, multicopper ferroxidase CgFet3 (oxidizes Fe2+ to Fe3+), an iron transporter/permease CgFtr1 (facilitates the passage of Fe3+ across the membrane) and a copper ion transporter CgCcc2 (loads copper on to CgFet3) [122]. Disruption of the high-affinity iron uptake components resulted in perturbed iron homoeostasis, debilitated survival under in vitro iron-limiting conditions and attenuated virulence [122]. Despite the presence of three ferric reductase genes, C. glabrata exhibited no surface ferric reductase activity, and it has been postulated that extracellular ferric reduction may be achieved through a secreted molecule [125]. C. glabrata could also utilize ferritin and ferric chloride as iron sources in a pH-dependent manner via reductive high-affinity iron uptake system [125].
Furthermore, C. glabrata has been reported to respond to iron-deplete condition via expression of the high-affinity iron permease CgFtr1 on the plasma membrane, and to iron-replete condition by trafficking CgFtr1 to the vacuole [57]. This retrograde trafficking of CgFtr1 from the plasma membrane to the vacuole is dependent on the sole class III phosphoinositide 3-kinase kinase, CgVps34 [57]. With regard to signaling pathways, Hog1-mediated MAPK has been shown to be essential for survival under high iron stress, as a lack of CgHog1 resulted in elevated intracellular iron and mitochondrial iron content and cell death in the high-iron environment [96].
The iron regulon in C. glabrata is comprised of a set of 51 genes that undergo reciprocal regulation in response to low and high environmental iron conditions [96]. C. glabrata possesses a unique hybrid iron regulatory network that consists of orthologs of the S. cerevisiae positive master iron regulator Aft1 and mRNA-encoding iron-requiring enzyme-degrading protein Cth2, as well as the C. albicans positive iron regulator Sef1 [126]. Of note, three bZip transcription factors, CgYap1, CgYap5 and CgYap7 have also been implicated in regulation of the heme biosynthesis, iron-excess stress response and iron-sulfur cluster biogenesis, respectively [127]. Additionally, CgHap5, a subunit of the CCAAT-binding complex, was found to interact with CgYap5, as well as being pivotal to the CgYap5-mediated iron stress response [128]. Figure 3 depicts major iron acquisition pathways in C. glabrata.
4. C. glabrata-Host Interaction
During its commensal and pathogenic life style, C. glabrata is thought to interact with host epithelial, endothelial and immune cells [10]. The in vivo and in vitro models, that have largely been used to study the pathogenesis of C. glabrata, include Mus musculus (mice), D. melanogaster (fruit fly) and Galleria mellonella (wax moth), and epithelial and endothelial cell lines, macrophage cell lines, human neutrophils and reconstituted oral epithelia, respectively [23,60,89,129,130,131,132]. A brief account of C. glabrata-host cell interaction is described below.
4.1. C. glabrata-Epithelial Cell Interaction
Adhesion of C. glabrata to epithelial cells and extracellular matrix is a prerequisite for mucosal colonization. Although the in vitro adherence of C. glabrata to epithelial cells is primarily mediated by the Epa1 adhesin [32], Epa6 and Epa7 have been shown to be pivotal to adherence under specific environmental conditions [30,36]. Epa6 is also known to mediate binding of C. glabrata to the human extracellular matrix protein, fibronectin [133]. Furthermore, increased adherence to Chinese Hamster Ovary derived-Lec2 cells has been reported for C. glabrata strains carrying the hyperactive CgPDR1 allele, which was attributed to the elevated expression of Epa1 [34]. Unlike C. albicans, C. glabrata elicited production of the granulocyte monocyte colony-stimulating factor (GM-CSF) in oral epithelial cells but caused no significant cytotoxicity [129]. This GM-CSF production in oral epithelial cells was later shown to be dependent upon the lactosylceramide receptor CDw17-mediated activation of NF (nuclear factor)-kB [134]. Similarly, C. glabrata could neither degrade the E-Cadherin protein present in the adherens junctions of the oral mucosal epithelium [135] nor invade the reconstituted human oral epithelium in vitro [130]. However, C. glabrata cells producing tryptophan-based pigment have been reported to cause increased damage to human oral epithelial TR146 cells [89]. A recent study has shown C. glabrata to induce phosphorylation of the ephrin type-A receptor 2 (EphA2), that binds to β-glucan, in immortalized normal human oral keratinocytes (OKF6/TERT-2) [136]. Furthermore, although the Toll-like receptor 2 (TLR-2) has also been implicated in recognition of C. glabrata and induction of the NF-κB-dependent release of TNF-α and IL-6 cytokines in rat tracheal epithelial cells [132], more studies are required to better understand the immune response of epithelial cells to association with C. glabrata.
4.2. C. glabrata-Endothelial Cell Interaction
The two main events that lead to systemic infection are dissemination (entry of the pathogen into the bloodstream) and tissue invasion (entry into surrounding tissues from the bloodstream). In an in vitro model of the human umbilical vein endothelial cells (HUVEC), C. glabrata was found to be able to cross the endothelial barrier [137]. Furthermore, two GPI-anchored cell wall proteins, CgPwp7 and CgAed1, have been shown to be required for adherence to HUVEC, as mutants lacking these adhesins were 2-3-fold less adherent [23]. However, unlike EPA genes, CgPWP7 and CgAED1 were not transcriptionally regulated by silencing [23]. In addition, two enzymes of the N-linked glycosylation system, α-1,6-mannosyltransferase (CgAnp1) and α-1,2-mannosyltransferase (CgMnn2), have also been implicated in adherence, as mutants lacking these enzymes were hyperadherent to the human microvascular endothelial cells HMEC-1 [138]. The C. glabrata Cgsir3Δ mutant, which expresses higher levels of Epa adhesins [36], has been reported to display mannose- and galactose-dependent strong binding to the coronary endothelium, through the coronary endothelial luminal membrane lectinic G protein-coupled receptors (GPCRs) including endothelin-2 and α-adrenergic 1A receptor, that led to altered cardiac functions [139]. Although this study underscored the binding of GPCRs to C. glabrata, C. glabrata-endothelial cell interaction are yet to be analyzed in depth.
4.3. C. glabrata-Neutrophil Interaction
Neutrophils are major players of the host defense system against fungal infections [140]. However, the role of neutrophils in the control of C. glabrata infections is not well-studied. Human neutrophils have been reported to release neutrophil extracellular traps after engulfment of C. glabrata cells [141]. Additionally, human neutrophils, after phagocytosis, killed and dumped C. glabrata, which may aid in activation of the immune response [142]. Intriguingly, C. glabrata was preferentially taken up by monocytes in the whole blood infection model, and infiltration of predominantly monocytes was observed in mouse kidneys [143]. Consistent with this, C. glabrata-activated neutrophils secreted monocyte chemoattractants, MIP-1α and MIP-1β, leading to increased migration of monocytes to the site of neutrophil-C. glabrata confrontation [143]. However, neutrophils of the dectin-2-/- knock out mice, which displayed elevated susceptibility to C. glabrata infections, have been reported to mount a deficient oxidative burst, pointing towards a role of the dectin-2 receptor and neutrophils in the control of C. glabrata infections [144]. The response of C. glabrata to the neutrophil environment has also been studied with genes involved in oxidative stress, gluconeogenesis, glyoxylate cycle, and methionine metabolism displaying upregulation [63]. Although a pigment derived from tryptophan is known to protect C. glabrata against the neutrophil attack [89], a detailed characterization of C. glabrata-neutrophil interaction is yet to be done.
4.4. C. glabrata-Natural Killer Cell Interaction
A recent study has highlighted the role of Natural Killer cells, which are effector lymphatic cells of the innate and adaptive immune system, in combating C. glabrata infections [145]. The Epa1, Epa6 and Epa7 adhesins were shown to be specifically recognized by the NKp46/NCR1 receptor, resulting in the clearance of disseminated infection [145]. Additionally, the murine dendritic cells have also been reported to produce IFN-β through the toll-like receptor TLR7, thereby, underscoring the role of IFN-I signaling in modulation of the host response to C. glabrata infection [146].
4.5. C. glabrata-Macrophage Interaction
Macrophages are the primary effectors of the innate immune system [140]. Besides engulfing and killing the fungal pathogen, they also facilitate recruitment of other immune cells, through cytokine and chemokine production, at the site of infection [140]. C. glabrata survives and replicates in human and murine macrophages without adversely affecting macrophages [60,65,147]. Among the pathogen recognition receptors, the C-type lectin receptors, dectin 1 and dectin-2, which recognize cell wall β-glucan, and α-mannan and β-glucan, respectively, have been implicated in the recognition of C. glabrata [144,148]. C. glabrata infection did not substantially activate any MAPK pathway including Erk1/2 (Extracellular signal-regulated kinases), SAPK/JNK (Stress-activated protein kinases/Jun amino-terminal kinases) and NF-κB signaling [149]. In accordance, macrophages did not produce TNF-α, IL-6, IL-8, IL-12, and IFN-γ pro-inflammatory cytokines, however, GM-CSF production has been reported upon C. glabrata infection [65]. Additionally, the spleen tyrosine kinase, Syk, is known to be phosphorylated in response to C. glabrata infection which resulted in the NLRP3 inflammasome-dependent production of IL-1β in human THP-1 macrophages [58]. The family of eleven cell surface-associated proteases (CgYapsins) has been shown to be required to keep the Syk pathway activation in check, as lack of these proteases led to increased IL-1β release and killing of C. glabrata cells [58].
C. glabrata is also known to impede maturation of the phagosome in macrophages [65,147], and the phosphoinositide 3-kinase subunits (CgVps15 and CgVsps34), mannosyltransferases (CgMnn10 and CgMnn11) and vesicular trafficking proteins (CgLdb17 and CgSla2) contributed to the inhibition of phagolysosomal acidification [147,149,150]. Of note, CgMnn10 and CgMnn11 have also been implicated in the ammonia extrusion-dependent environmental alkalinization which may partly account for their role in impeding phagolysosomal acidification [149]. C. glabrata activates pexophagy and autophagy processes, undergoes chromatin remodeling and evades immune responses to survive and replicate in macrophages [84,147,150]. Lastly, despite encountering iron restriction, ROS and carbon starvation, C. glabrata has equipped itself well with strategies to proliferate in macrophages, including transcriptional reconfiguration of cellular pathways [60,65,147]. Genes belonging to the glyoxylate cycle, β-oxidation of fatty acids, gluconeogenesis, methyl citrate cycle and proteolysis have been shown to be upregulated, while genes encoding glycolytic enzymes and ribosomal translational machinery components were found to be downregulated in macrophage-internalized C. glabrata cells [60]. Furthermore, the continuous contact with macrophages during microevolution studies led to a change in the morphology of C. glabrata cells from yeast to pseudohyphae which was attributed to a mutation in the chitin synthase-encoding gene CgCHS2 [151]. Altogether, because of its ability to subvert the immune response and replicate in macrophages in vitro, macrophages are assumed to be the Trojan horses for C. glabrata [58,143,151]. Figure 4 summarizes key aspects of C. glabrata-macrophage interaction.
5. Conclusions
C. glabrata occupies a unique position in the phylogenetic tree and appears to possess requisite attributes to establish successful infections in the human host. However, it is significantly less pathogenic than C. albicans. The lack of invasive hyphal forms, secreted proteolytic activity and invasins, and limited nutrient plasticity including non-utilization of haemoglobin as an iron source are likely to contribute to the low pathogenicity of C. glabrata. Research over the last two decades has brought many unexpected biological features of C. glabrata to the fore. Owing to its reduced susceptibility to azole antifungals and emerging resistance to echinocandins, effective treatment of C. glabrata infections remains a clinical challenge. Hence, future research, focusing on adhesins, proteases, stress response regulators and nutrient acquisition machinery, that may modulate interaction with the host, is likely to elucidate precise mechanisms underneath the commensal and the opportunistic life style of this important pathogen.
Author Contributions
R.K. conceived and designed the content of the review. K.K., F.A. and M.S.S. prepared the figures. K.K., F.A., M.S.S. and R.K. wrote the manuscript.
Funding
Research in Kaur laboratory is funded by the Wellcome Trust/DBT India Alliance Senior Fellowship to RK [IA/S/15/1/501831], and grants from the Department of Biotechnology [BT/HRD/NBA/37/01/2014], and the Science and Engineering Research Board, Department of Science and Technology [EMR/2016/005375], Government of India, to R.K. K.K. and M.S.S. are recipients of the Shyama Prasad Mukherjee Fellowship and Junior Research Fellowship, respectively, sponsored by the Council of Scientific & Industrial Research, New Delhi, India. F.A. is a recipient of the Junior Research Fellowship sponsored by the University Grants Commission, New Delhi, India.
Acknowledgments
We apologize to all colleagues whose work could not be cited in this review owing to space constraints.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pfaller, M.A.; Messer, S.A.; Moet, G.J.; Jones, R.N.; Castanheira, M. Candida bloodstream infections: Comparison of species distribution and resistance to echinocandin and azole antifungal agents in Intensive Care Unit (ICU) and non-ICU settings in the SENTRY antimicrobial surveillance program (2008–2009). Int. J. Antimicrob. Agents 2011, 38, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Montagna, M.T.; Lovero, G.; Borghi, E.; Amato, G.; Andreoni, S.; Campion, L.; Lo Cascio, G.; Lombardi, G.; Luzzaro, F.; Manso, E.; et al. Candidemia in intensive care unit: A nationwide prospective observational survey (GISIA-3 study) and review of the European literature from 2000 through 2013. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 661–674. [Google Scholar] [PubMed]
- Yapar, N. Epidemiology and risk factors for invasive candidiasis. Ther. Clin. Risk Manag. 2014, 10, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Sood, P.; Rudramurthy, S.M.; Chen, S.; Kaur, H.; Capoor, M.; Chhina, D.; Rao, R.; Eshwara, V.K.; Xess, I.; et al. Incidence, characteristics and outcome of ICU-acquired candidemia in India. Intensive Care Med. 2014, 41, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Diekema, D.; Arbefeville, S.; Boyken, L.; Kroeger, J.; Pfaller, M. The changing epidemiology of healthcare-associated candidemia over three decades. Diagn. Microbiol. Infect. Dis. 2012, 73, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Guinea, J. Global trends in the distribution of Candida species causing candidemia. Clin. Microbiol. Infect. 2014, 20, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.H.; Chakrabarti, A.; Li, R.Y.; Patel, A.K.; Watcharananan, S.P.; Liu, Z.; Chindamporn, A.; Tan, A.L.; Sun, P.L.; Wu, U.I.; et al. Incidence and species distribution of candidaemia in Asia: A laboratory-based surveillance study. Clin. Microbiol. Infect. 2015, 21, 946–953. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Andes, D.R.; Diekema, D.J.; Horn, D.L.; Reboli, A.C.; Rotstein, C.; Franks, B.; Azie, N.E. Epidemiology and outcomes of invasive candidiasis due to non-albicans species of Candida in 2,496 patients: Data from the Prospective Antifungal Therapy (PATH) registry 2004–2008. PLoS ONE 2014, 9, e101510. [Google Scholar] [CrossRef]
- Khatib, R.; Johnson, L.B.; Fakih, M.G.; Riederer, K.; Briski, L. Current trends in candidemia and species distribution among adults: Candida glabrata surpasses C. albicans in diabetic patients and abdominal sources. Mycoses 2016, 59, 781–786. [Google Scholar] [CrossRef]
- Fidel, P.L.; Vazquez, J.A.; Sobel, J.D. Candida glabrata: Review of epidemiology, pathogenesis, and clinical disease with comparison to C. albicans. Clin. Microbiol. Rev. 1999, 12, 80–96. [Google Scholar] [CrossRef]
- Achkar, J.M.; Fries, B.C. Candida infections of the genitourinary tract. Clin. Microbiol. Rev. 2010, 23, 253–273. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J.A. A history of research on yeasts 8: Taxonomy. Yeast 2004, 21, 1141–1193. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P. Phylogenetic circumscription of Saccharomyces, Kluyveromyces and other members of the Saccharomycetaceae, and the proposal of the new genera Lachancea, Nakaseomyces, Naumovia, Vanderwaltozyma and Zygotorulaspora. FEMS Yeast Res. 2003, 4, 233–245. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. FEMS Microbiol. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef]
- Zacharioudakis, I.; Zervou, F.; Mylonakis, E. T2 Magnetic Resonance Assay: Overview of available data and clinical implications. J. Fungi 2018, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Clancy, C.J.; Nguyen, M.H. Diagnosing invasive candidiasis. J. Clin. Microbiol. 2018, 56, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dujon, B.; Sherman, D.; Fischer, G.; Durrens, P.; Casaregola, S.; Lafontaine, I.; De Montigny, J.; Marck, C.; Neuvéglise, C.; Talla, E.; et al. Genome evolution in yeasts. Nature 2004, 430, 35–44. [Google Scholar] [CrossRef]
- Gabaldón, T.; Martin, T.; Marcet-Houben, M.; Durrens, P.; Bolotin-Fukuhara, M.; Lespinet, O.; Arnaise, S.; Boisnard, S.; Aguileta, G.; Atanasova, R.; et al. Comparative genomics of emerging pathogens in the Candida glabrata clade. BMC Genomics 2013, 14, 623. [Google Scholar] [CrossRef]
- Jong, H.S.; Myung, J.C.; Jeong, W.S.; Jung, S.I.; Cho, D.; Seung, J.K.; Soo, H.K.; Myung, G.S.; Soon, P.S.; Dong, W.R. Changes in karyotype and azole susceptibility of sequential bloodstream isolates from patients with Candida glabrata candidemia. J. Clin. Microbiol. 2007, 45, 2385–2391. [Google Scholar]
- Bader, O.; Schwarz, A.; Kraneveld, E.A.; Tangwattanchuleeporn, M.; Schmidt, P.; Jacobsen, M.D.; Gross, U.; de Groot, P.W.J.; Weig, M. Gross karyotypic and phenotypic alterations among different progenies of the Candida glabrata CBS138/ATCC2001 reference strain. PLoS ONE 2012, 7, 1–8. [Google Scholar] [CrossRef]
- Ahmad, K.M.; Ishchuk, O.P.; Hellborg, L.; Jørgensen, G.; Skvarc, M.; Stenderup, J.; Jørck-Ramberg, D.; Polakova, S.; Piškur, J. Small chromosomes among Danish Candida glabrata isolates originated through different mechanisms. Antonie van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2013, 104, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Fischer, G.; Rocha, E.P.C.; Brunet, F.; Vergassola, M.; Dujon, B. Highly variable rates of genome rearrangements between hemiascomycetous yeast lineages. PLoS Genet. 2006, 2, 0253–0261. [Google Scholar] [CrossRef] [PubMed]
- Desai, C.; Mavrianos, J.; Chauhan, N. Candida glabrata Pwp7p and Aed1p are required for adherence to human endothelial cells. FEMS Yeast Res. 2011, 11, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Marcet-Houben, M.; Gabaldón, T. Beyond the whole-genome duplication: Phylogenetic evidence for an ancient interspecies hybridization in the baker’s yeast lineage. PLoS Biol. 2015, 13, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Carreté, L.; Ksiezopolska, E.; Pegueroles, C.; Gómez-Molero, E.; Saus, E.; Iraola-Guzmán, S.; Loska, D.; Bader, O.; Fairhead, C.; Gabaldón, T. Patterns of genomic variation in the opportunistic pathogen Candida glabrata suggest the existence of mating and a secondary association with humans. Curr. Biol. 2018, 28, 15–27.e7. [Google Scholar] [CrossRef] [PubMed]
- Koszul, R.; Malpertuy, A.; Frangeul, L.; Bouchier, C.; Wincker, P.; Thierry, A.; Duthoy, S.; Ferris, S.; Hennequin, C.; Dujon, B. The complete mitochondrial genome sequence of the pathogenic yeast Candida (Torulopsis) glabrata. FEBS Lett. 2003, 534, 39–48. [Google Scholar] [CrossRef]
- De Groot, P.W.J.; Kraneveld, E.A.; Qing, Y.Y.; Dekker, H.L.; Groß, U.; Crielaard, W.; De Koster, C.G.; Bader, O.; Klis, F.M.; Weig, M. The cell wall of the human pathogen Candida glabrata: Differential incorporation of novel adhesin-like wall proteins. Eukaryot. Cell 2008, 7, 1951–1964. [Google Scholar] [CrossRef]
- Kaur, R.; Domergue, R.; Zupancic, M.L.; Cormack, B.P. A yeast by any other name: Candida glabrata and its interaction with the host. Curr. Opin. Microbiol. 2005, 8, 378–384. [Google Scholar] [CrossRef]
- De Las Peñas, A.; Pan, S.J.; Castaño, I.; Alder, J.; Cregg, R.; Cormack, B.P. Virulence-related surface glycoproteins in the yeast pathogen Candida glabrata are encoded in subtelomeric clusters and subject to RAP1- and SIR-dependent transcriptional silencing. Genes Dev. 2003, 17, 2245–2258. [Google Scholar] [CrossRef]
- Domergue, R. Nicotinic acid limitation regulates silencing of Candida adhesins during UTI. Science 2005, 308, 866–870. [Google Scholar] [CrossRef]
- López-Fuentes, E.; Gutiérrez-Escobedo, G.; Timmermans, B.; Van Dijck, P.; De Las Peñas, A.; Castaño, I. Candida glabrata’s genome plasticity confers a unique pattern of expressed cell wall proteins. J. Fungi 2018, 4, 67. [Google Scholar] [CrossRef]
- Cormack, B.P. An adhesin of the yeast pathogen Candida glabrata mediating adherence to human epithelial cells. Science 1999, 285, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, D.M.; Vyas, V.K. The Candida glabrata adhesin Epa1p causes adhesion, phagocytosis, and cytokine secretion by innate immune cells. FEMS Yeast Res. 2012, 12, 398–414. [Google Scholar] [CrossRef] [PubMed]
- Vale-Silva, L.A.; Moeckli, B.; Torelli, R.; Posteraro, B.; Sanglard, D. Upregulation of the adhesin gene EPA1 mediated by PDR1 in Candida glabrata leads to enhanced host colonization. mSphere 2016, 1, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zupancic, M.L.; Frieman, M.; Smith, D.; Alvarez, R.A.; Cummings, R.D.; Cormack, B.P. Glycan microarray analysis of Candida glabrata adhesin ligand specificity. Mol. Microbiol. 2008, 68, 547–559. [Google Scholar] [CrossRef]
- Castaño, I.; Pan, S.J.; Zupancic, M.; Hennequin, C.; Dujon, B.; Cormack, B.P. Telomere length control and transcriptional regulation of subtelomeric adhesins in Candida glabrata. Mol. Microbiol. 2005, 55, 1246–1258. [Google Scholar] [CrossRef]
- Iraqui, I.; Garcia-Sanchez, S.; Aubert, S.; Dromer, F.; Ghigo, J.M.; D’Enfert, C.; Janbon, G. The Yak1p kinase controls expression of adhesins and biofilm formation in Candida glabrata in a Sir4p-dependent pathway. Mol. Microbiol. 2005, 55, 1259–1271. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Gregori, C.; Jennings, A.M.; Quintin, J.; Ferrandon, D.; Butler, G.; Kuchler, K.; Ammerer, G.; Schüller, C. Candida glabrata environmental stress response involves Saccharomyces cerevisiae Msn2/4 orthologous transcription factors. Mol. Microbiol. 2008, 69, 603–620. [Google Scholar] [CrossRef]
- Mundy, R.D.; Cormack, B. Expression of Candida glabrata adhesins after exposure to chemical preservatives. J. Infect. Dis. 2009, 199, 1891–1898. [Google Scholar] [CrossRef]
- Kraneveld, E.A.; de Soet, J.J.; Deng, D.M.; Dekker, H.L.; de Koster, C.G.; Klis, F.M.; Crielaard, W.; de Groot, P.W.J. Identification and differential gene expression of adhesin-like wall proteins in Candida glabrata biofilms. Mycopathologia. 2011, 172, 415–427. [Google Scholar] [CrossRef]
- Gómez-Molero, E.; de Boer, A.D.; Dekker, H.L.; Moreno-Martínez, A.; Kraneveld, E.A.; Ichsan; Chauhan, N.; Weig, M.; de Soet, J.J.; de Koster, C.G.; et al. Proteomic analysis of hyperadhesive Candida glabrata clinical isolates reveals a core wall proteome and differential incorporation of adhesins. FEMS Yeast Res. 2015, 15, fov098. [Google Scholar]
- Juárez-Cepeda, J.; Orta-Zavalza, E.; Cañas-Villamar, I.; Arreola-Gómez, J.; Pérez-Cornejo, G.P.; Hernández-Carballo, C.Y.; Gutiérrez-Escobedo, G.; Castaño, I.; De Las Peñas, A. The EPA2 adhesin encoding gene is responsive to oxidative stress in the opportunistic fungal pathogen Candida glabrata. Curr. Genet. 2015, 61, 529–544. [Google Scholar] [CrossRef]
- Maestre-reyna, M.; Diderrich, R.; Stefan, M.; Eulenburg, G.; Kalugin, V.; Brückner, S. Structural basis for promiscuity and specificity during Candida glabrata invasion of host epithelia. Proc. Natl. Acad. Sci. USA 2012, 109, 16864–16869. [Google Scholar] [CrossRef]
- Ielasi, F.S.; Alioscha-perez, M.; Donohue, D.; Claes, S.; Sahli, H.; Schols, D. Lectin-glycan interaction network-based identification of host receptors of microbial pathogenic adhesins. mBio 2016, 7, 1–17. [Google Scholar] [CrossRef]
- Diderrich, R.; Kock, M.; Maestre-Reyna, M.; Keller, P.; Steuber, H.; Rupp, S.; Essen, L.O.; Mösch, H.U. Structural hot spots determine functional diversity of the Candida glabrata epithelial adhesin family. J. Biol. Chem. 2015, 290, 19597–19613. [Google Scholar] [CrossRef]
- Linde, J.; Duggan, S.; Weber, M.; Horn, F.; Sieber, P.; Hellwig, D.; Riege, K.; Marz, M.; Martin, R.; Guthke, R.; et al. Defining the transcriptomic landscape of Candida glabrata by RNA-Seq. Nucleic Acids Res. 2015, 43, 1392–1406. [Google Scholar] [CrossRef]
- Thierry, A.; Bouchier, C.; Dujon, B.; Richard, G.F. Megasatellites: A peculiar class of giant minisatellites in genes involved in cell adhesion and pathogenicity in Candida glabrata. Nucleic Acids Res. 2008, 36, 5970–5982. [Google Scholar] [CrossRef]
- Vale-Silva, L.; Beaudoing, E.; Tran, V.D.T.; Sanglard, D. Comparative genomics of two sequential Candida glabrata clinical isolates. G3 2017, 7, 2413–2426. [Google Scholar] [CrossRef]
- Salazar, S.B.; Wang, C.; Musterkotter, M.; Okamoto, M.; Takahashi-Nakaguchi, A.; Chibana, H.; Lopes, M.M.; Güldener, U.; Butler, G.; Mira, N.P. Comparative genomic and transcriptomic analyses unveil novel features of azole resistance and adaptation to the human host in Candida glabrata. FEMS Yeast Res. 2017, 18, 1–11. [Google Scholar] [CrossRef]
- D’Enfert, C.; Janbon, G. Biofilm formation in Candida glabrata: What have we learnt from functional genomics approaches? FEMS Yeast Res. 2015, 16, 1–13. [Google Scholar]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: Quantification, structure and matrix composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef]
- Mitchell, K.F.; Taff, H.T.; Cuevas, M.A.; Reinicke, E.L.; Sanchez, H.; Andes, D.R. Role of matrix β-1,3 glucan in antifungal resistance of non-albicans Candida biofilms. Antimicrob. Agents Chemother. 2013, 57, 1918–1920. [Google Scholar] [CrossRef]
- Riera, M.; Mogensen, E.; d’Enfert, C.; Janbon, G. New regulators of biofilm development in Candida glabrata. Res. Microbiol. 2012, 163, 297–307. [Google Scholar] [CrossRef]
- Schwarzmuller, T.; Ma, B.; Hiller, E.; Istel, F.; Tscherner, M.; Brunke, S.; Ames, L.; Firon, A.; Green, B.; Cabral, V.; et al. Systematic phenotyping of a large-scale Candida glabrata deletion collection reveals novel antifungal tolerance genes. PLoS Pathog. 2014, 10, e1004211. [Google Scholar] [CrossRef]
- Kucharíková, S.; Neirinck, B.; Sharma, N.; Vleugels, J.; Lagrou, K.; Van Dijck, P. In vivo Candida glabrata biofilm development on foreign bodies in a rat subcutaneous model. J. Antimicrob. Chemother. 2015, 70, 846–856. [Google Scholar] [CrossRef]
- Jayampath Seneviratne, C.; Wang, Y.; Jin, L.; Abiko, Y.; Samaranayake, L.P. Proteomics of drug resistance in Candida glabrata biofilms. Proteomics 2010, 10, 1444–1454. [Google Scholar] [CrossRef]
- Sharma, V.; Purushotham, R.; Kaur, R. The phosphoinositide 3-kinase regulates retrograde trafficking of the iron permease CgFtr1 and iron homeostasis in Candida glabrata. J. Biol. Chem. 2016, 291, 24715–24734. [Google Scholar] [CrossRef]
- Rasheed, M.; Battu, A.; Kaur, R. Aspartyl proteases in Candida glabrata are required for suppression of the host innate immune response. J. Biol. Chem. 2018, 293, 6410–6433. [Google Scholar] [CrossRef]
- Kantarcioǧlu, A.S.; Yücel, A. Phospholipase and protease activities in clinical Candida isolates with reference to the sources of strains. Mycoses 2002, 45, 160–165. [Google Scholar] [CrossRef]
- Kaur, R.; Ma, B.; Cormack, B.P. A family of glycosylphosphatidylinositol-linked aspartyl proteases is required for virulence of Candida glabrata. Proc. Natl. Acad. Sci. USA 2007, 104, 7628–7633. [Google Scholar] [CrossRef]
- Bairwa, G.; Kaur, R. A novel role for a glycosylphosphatidylinositol-anchored aspartyl protease, CgYps1, in the regulation of pH homeostasis in Candida glabrata. Mol. Microbiol. 2011, 79, 900–913. [Google Scholar] [CrossRef]
- Miyazaki, T.; Izumikawa, K.; Yamauchi, S.; Inamine, T.; Nagayoshi, Y.; Saijo, T.; Seki, M.; Kakeya, H.; Yamamoto, Y.; Yanagihara, K.; et al. The glycosylphosphatidylinositol-linked aspartyl protease Yps1 is transcriptionally regulated by the calcineurin-Crz1 and Slt2 MAPK pathways in Candida glabrata. FEMS Yeast Res. 2011, 11, 449–456. [Google Scholar] [CrossRef]
- Fukuda, Y.; Tsai, H.F.; Myers, T.G.; Bennett, J.E. Transcriptional profiling of Candida glabrata during phagocytosis by neutrophils and in the infected mouse spleen. Infect. Immun. 2013, 81, 1325–1333. [Google Scholar] [CrossRef]
- Bairwa, G.; Rasheed, M.; Taigwal, R.; Sahoo, R.; Kaur, R. GPI (glycosylphosphatidylinositol)-linked aspartyl proteases regulate vacuole homoeostasis in Candida glabrata. Biochem. J. 2014, 458, 323–334. [Google Scholar] [CrossRef]
- Seider, K.; Brunke, S.; Schild, L.; Jablonowski, N.; Wilson, D.; Majer, O.; Barz, D.; Haas, A.; Kuchler, K.; Schaller, M.; et al. The facultative intracellular pathogen Candida glabrata subverts macrophage cytokine production and phagolysosome maturation. J. Immunol. 2011, 187, 3072–3086. [Google Scholar] [CrossRef]
- Quintin, J.; Asmar, J.; Matskevich, A.A.; Lafarge, M.-C.; Ferrandon, D. The Drosophila toll pathway controls but does not clear Candida glabrata infections. J. Immunol. 2013, 190, 2818–2827. [Google Scholar] [CrossRef]
- Enkler, L.; Richer, D.; Marchand, A.L.; Ferrandon, D.; Jossinet, F. Genome engineering in the yeast pathogen Candida glabrata using the CRISPR-Cas9 system. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Lachke, S.A.; Joly, S.; Daniels, K.; Soll, D.R. Phenotypic switching and filamentation in Candida glabrata. Microbiology 2002, 148, 2661–2674. [Google Scholar] [CrossRef]
- Brockert, P.J.; Lachke, S.A.; Srikantha, T.; Pujol, C.; Galask, R.; Soll, D.R. Phenotypic switching and mating type switching of Candida glabrata at sites of colonization. Infect. Immun. 2003, 71, 7109–7118. [Google Scholar] [CrossRef]
- Csank, C.; Haynes, K. Candida glabrata displays pseudohyphal growth. FEMS Microbiol. Lett. 2000, 189, 115–120. [Google Scholar] [CrossRef]
- Sasani, E.; Khodavaisy, S.; Agha Kuchak Afshari, S.; Darabian, S.; Aala, F.; Rezaie, S. Pseudohyphae formation in Candida glabrata due to CO2 exposure. Curr. Med. Mycol. 2016, 2, 49–52. [Google Scholar] [CrossRef]
- Kamran, M.; Calcagno, A.; Findon, H.; Bignell, E.; Jones, M.D.; Warn, P.; Denning, D.W.; Butler, G.; Mühlschlegel, F.A.; Haynes, K.; et al. Inactivation of transcription factor gene ACE2 in the fungal pathogen Candida glabrata results in hypervirulence. Eukaryot. Cell 2004, 3, 546–552. [Google Scholar] [CrossRef]
- MacCallum, D.M.; Findon, H.; Kenny, C.C.; Butler, G.; Haynes, K.; Odds, F.C. Different consequences of ACE2 and SWI5 gene disruptions for virulence of pathogenic and nonpathogenic yeasts. Infect. Immun. 2006, 74, 5244–5248. [Google Scholar] [CrossRef]
- Srikantha, T.; Lachke, S.A.; Soll, D.R. Three mating type-like loci in Candida glabrata. Eukaryot. Cell 2003, 2, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.A.; Heitman, J. Fungal mating-type loci. Curr. Biol. 2003, 13, R792–R795. [Google Scholar] [CrossRef]
- Robledo-Márquez, K.; Gutiérrez-Escobedo, G.; Yáñez-Carrillo, P.; Vidal-Aguiar, Y.; Briones-Martín-del-Campo, M.; Orta-Zavalza, E.; De Las Peñas, A.; Castaño, I. Candida glabrata encodes a longer variant of the mating type (MAT) alpha2 gene in the mating type-like MTL3 locus, which can form homodimers. FEMS Yeast Res. 2016, 16, 1–12. [Google Scholar] [CrossRef]
- Ramírez-Zavaleta, C.Y.; Salas-Delgado, G.E.; de Las Peñas, A.; Castaño, I. Subtelomeric silencing of the MTL3 locus of Candida glabrata requires yKu70, yKu80, and Rif1 proteins. Eukaryot. Cell 2010, 9, 1602–1611. [Google Scholar] [CrossRef]
- Boisnard, S.; Li, Y.Z.; Arnaise, S.; Sequeira, G.; Raffoux, X.; Enache-Angoulvant, A.; Bolotin-Fukuhara, M.; Fairhead, C. Efficient mating-type switching in Candida glabrata induces cell death. PLoS ONE 2015, 10, 1–18. [Google Scholar] [CrossRef]
- Nikolaou, E.; Agrafioti, I.; Stumpf, M.; Quinn, J.; Stansfield, I.; Brown, A.J. Phylogenetic diversity of stress signalling pathways in fungi. BMC Evol. Biol. 2009, 9, 1–18. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Briones-Martin-del-Campo, M.; Cañas-Villamar, I.; Montalvo-Arredondo, J.; Riego-Ruiz, L.; Castaño, I.; De Las Peñas, A. High resistance to oxidative stress in the fungal pathogen Candida glabrata is mediated by a single catalase, Cta1p, and is controlled by the transcription factors Yap1p, Skn7p, Msn2p, and Msn4p. Eukaryot. Cell 2008, 7, 814–825. [Google Scholar] [CrossRef]
- Briones-Martin-Del-Campo, M.; Orta-Zavalza, E.; Caïas-Villamar, I.; Gutiïrrez-Escobedo, G.; Juïrez-Cepeda, J.; Robledo-Mïrquez, K.; Arroyo-Helguera, O.; Castaïo, I.; De Las Peïas, A. The superoxide dismutases of Candida glabrata protect against oxidative damage and are required for lysine biosynthesis, DNA integrity and chronological life survival. Microbiology 2015, 161, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Desai, P.R.; Rai, M.N.; Kaur, R.; Ganesan, K.; Bachhawat, A.K. Glutathione biosynthesis in the yeast pathogens Candida glabrata and Candida albicans: Essential in C. glabrata, and essential for virulence in C. albicans. Microbiology 2011, 157, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Escobedo, G.; Orta-Zavalza, E.; Castaño, I.; De Las Peñas, A. Role of glutathione in the oxidative stress response in the fungal pathogen Candida glabrata. Curr. Genet. 2013, 59, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Gratz, N.; Kovarik, P.; Schüller, C. Autophagy supports Candida glabrata survival during phagocytosis. Cell. Microbiol. 2010, 12, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Klopf, E.; Gratz, N.; Marcet-Houben, M.; Hiller, E.; Rupp, S.; Gabaldón, T.; Kovarik, P.; Schüller, C. Regulation of Candida glabrata oxidative stress resistance is adapted to host environment. FEBS Lett. 2011, 585, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Saijo, T.; Miyazaki, T.; Izumikawa, K.; Mihara, T.; Takazono, T.; Kosai, K.; Imamura, Y.; Seki, M.; Kakeya, H.; Yamamoto, Y.; et al. Skn7p is involved in oxidative stress response and virulence of Candida glabrata. Mycopathologia 2010, 169, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Orta-Zavalza, E.; Guerrero-Serrano, G.; Gutierrez-Escobedo, G.; Canas-Villamar, I.; Juarez-Cepeda, J.; Castano, I.; De Las Penas, A. Local silencing controls the oxidative stress response and the multidrug resistance in Candida glabrata. Mol. Microbiol. 2013, 88, 1135–1148. [Google Scholar] [CrossRef] [PubMed]
- Kounatidis, I.; Ames, L.; Mistry, R.; Ho, H.-L.; Haynes, K.; Ligoxygakis, P. A host-pathogen interaction screen identifies ada2 as a mediator of Candida glabrata defences against reactive oxygen species. G3 2018, 8, 1637–1647. [Google Scholar] [CrossRef] [PubMed]
- Brunke, S.; Seider, K.; Almeida, R.S.; Heyken, A.; Fleck, C.B.; Brock, M.; Barz, D.; Rupp, S.; Hube, B. Candida glabrata tryptophan-based pigment production via the Ehrlich pathway. Mol. Microbiol. 2010, 76, 25–47. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Inamine, T.; Yamauchi, S.; Nagayoshi, Y.; Saijo, T.; Izumikawa, K.; Seki, M.; Kakeya, H.; Yamamoto, Y.; Yanagihara, K.; et al. Role of the Slt2 mitogen-activated protein kinase pathway in cell wall integrity and virulence in Candida glabrata. FEMS Yeast Res. 2010, 10, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Borah, S.; Shivarathri, R.; Kaur, R. The Rho1 GTPase-activating protein CgBem2 is required for survival of azole stress in Candida glabrata. J. Biol. Chem. 2011, 286, 34311–34324. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sasaki, M.; Ito, F.; Aoyama, T.; Sato-Okamoto, M.; Takahashi-Nakaguchi, A.; Chibana, H.; Shibata, N. Cooperation between ER stress and calcineurin signaling contributes to the maintenance of cell wall integrity in Candida glabrata. Fungal Biol. 2018, 122, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Yamauchi, S.; Inamine, T.; Nagayoshi, Y.; Saijo, T.; Izumikawa, K.; Seki, M.; Kakeya, H.; Yamamoto, Y.; Yanagihara, K.; et al. Roles of calcineurin and Crz1 in antifungal susceptibility and virulence of Candida glabrata. Antimicrob. Agents Chemother. 2010, 54, 1639–1643. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-L.; Konieczka, J.H.; Springer, D.J.; Bowen, S.E.; Zhang, J.; Silao, F.G.S.; Bungay, A.A.C.; Bigol, U.G.; Nicolas, M.G.; Abraham, S.N.; et al. Convergent evolution of calcineurin pathway roles in thermotolerance and virulence in Candida glabrata. G3 2012, 2, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Jandric, Z.; Gregori, C.; Klopf, E.; Radolf, M.; Schüller, C. Sorbic acid stress activates the Candida glabrata high osmolarity glycerol MAP kinase pathway. Front. Microbiol. 2013, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.K.; Suneetha, K.J.; Kaur, R. The mitogen-activated protein kinase CgHog1 is required for iron homeostasis, adherence and virulence in Candida glabrata. FEBS J. 2015, 282, 2142–2166. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Nakayama, H.; Nagayoshi, Y.; Kakeya, H.; Kohno, S. Dissection of Ire1 functions reveals stress response mechanisms uniquely evolved in Candida glabrata. PLoS Pathog. 2013, 9, e1003160. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.-J.; Chang, Y.-L.; Chen, Y.-L. Deletion of ADA2 increases antifungal drug susceptibility and virulence in Candida glabrata. Antimicrob. Agents Chemother. 2018, 62, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Castano, I.; Cormack, B.P. Functional genomic analysis of fluconazole susceptibility in the pathogenic yeast Candida glabrata: Roles of calcium signaling and mitochondria. Antimicrob. Agents Chemother. 2004, 48, 1600–1613. [Google Scholar] [CrossRef] [PubMed]
- Singh-Babak, S.D.; Babak, T.; Diezmann, S.; Hill, J.A.; Xie, J.L.; Chen, Y.L.; Poutanen, S.M.; Rennie, R.P.; Heitman, J.; Cowen, L.E. Global analysis of the evolution and mechanism of echinocandin resistance in Candida glabrata. PLoS Pathog. 2012, 8, e1002718. [Google Scholar] [CrossRef] [PubMed]
- Whaley, S.G.; Rogers, P.D. Azole Resistance in Candida glabrata. Curr. Infect. Dis. Rep. 2016, 18, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Messer, S.A.; Rhomberg, P.R.; Pfaller, M.A. Antifungal susceptibility patterns of a global collection of fungal isolates: Results of the SENTRY antifungal surveillance program (2013). Diagn. Microbiol. Infect. Dis. 2016, 85, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Healey, K.; Perlin, D.S. Fungal resistance to echinocandins and the MDR phenomenon in Candida glabrata. J. Fungi 2018, 4, 105. [Google Scholar]
- Sanglard, D.; Ischer, F.; Calabrese, D.; Majcherczyk, P.A.; Bille, J. The ATP binding cassette transporter gene CgCDR1 from Candida glabrata is involved in the resistance of clinical isolates to azole antifungal agents. Antimicrob. Agents Chemother. 1999, 43, 2753–2765. [Google Scholar] [CrossRef]
- Izumikawa, K.; Kakeya, H.; Tsai, H.F.; Grimberg, B.; Bennett, J.E. Function of Candida glabrata ABC transporter gene, PDH1. Yeast 2003, 20, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Cota, J.M.; Grabinski, J.L.; Talbert, R.L.; Burgess, D.S.; Rogers, P.D.; Edlind, T.D.; Wiederhold, N.P. Increases in SLT2 expression and chitin content are associated with incomplete killing of Candida glabrata by caspofungin. Antimicrob. Agents Chemother. 2008, 52, 1144–1146. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, P.; Tronchin, G.; Bergès, T.; Hennequin, C.; Chabasse, D.; Bouchara, J.P. Reduced susceptibility to polyenes associated with a missense mutation in the ERG6 gene in a clinical isolate of Candida glabrata with pseudohyphal growth. Antimicrob. Agents Chemother. 2007, 51, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, P.; Tronchin, G.; Larcher, G.; Ernoult, E.; Bergès, T.; Chabasse, D.; Bouchara, J.P. A nonsense mutation in the ERG6 gene leads to reduced susceptibility to polyenes in a clinical isolate of Candida glabrata. Antimicrob. Agents Chemother. 2008, 52, 3701–3709. [Google Scholar] [CrossRef]
- Healey, K.R.; Zhao, Y.; Perez, W.B.; Lockhart, S.R.; Sobel, J.D.; Farmakiotis, D.; Kontoyiannis, D.P.; Sanglard, D.; Taj-Aldeen, S.J.; Alexander, B.D.; et al. Prevalent mutator genotype identified in fungal pathogen Candida glabrata promotes multi-drug resistance. Nat. Commun. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Van Urk, H.; Voll, W.S.L.; Scheffers, W.A.; Van Dijken, J.P. Transient-state analysis of metabolic fluxes in crabtree-positive and crabtree-negative yeasts. Appl. Environ. Microbiol. 1990, 56, 281–287. [Google Scholar]
- Legrand, J.; Bolotin-Fukuhara, M.; Bourgais, A.; Fairhead, C.; Sicard, D. Life-history strategies and carbon metabolism gene dosage in the Nakaseomyces yeasts. FEMS Yeast Res. 2016, 16, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Freese, S.; Vogts, T.; Speer, F.; Schäfer, B.; Passoth, V.; Klinner, U. C- and N-catabolic utilization of tricarboxylic acid cycle-related amino acids by Scheffersomyces stipitis and other yeasts. Yeast 2011, 28, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.S.; Chew, S.Y.; Rangasamy, P.; Mohd Desa, M.N.; Sandai, D.; Chong, P.P.; Than, L.T.L. SNF3 as high affinity glucose sensor and its function in supporting the viability of Candida glabrata under glucose-limited environment. Front. Microbiol. 2015, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Conant, G.C.; Wolfe, K.H. Increased glycolytic flux as an outcome of whole-genome duplication in yeast. Mol. Syst. Biol. 2007, 3, 129. [Google Scholar] [CrossRef] [PubMed]
- Kerwin, C.L.; Wykoff, D.D. Candida glabrata PHO4 is necessary and sufficient for Pho2-independent transcription of phosphate starvation genes. Genetics 2009, 182, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Orkwis, B.R.; Davies, D.L.; Kerwin, C.L.; Sanglard, D.; Wykoff, D.D. Novel acid phosphatase in Candida glabrata suggests selective pressure and niche specialization in the phosphate signal transduction pathway. Genetics 2010, 186, 885–895. [Google Scholar] [CrossRef] [PubMed]
- Kerwin, C.L.; Wykoff, D.D. De novo generation of a phosphate starvation-regulated promoter in Candida glabrata. FEMS Yeast Res. 2012, 12, 980–989. [Google Scholar] [CrossRef]
- He, B.Z.; Zhou, X.; O’Shea, E.K. Evolution of reduced co-activator dependence led to target expansion of a starvation response pathway. eLife 2017, 6, 1–26. [Google Scholar] [CrossRef]
- Nahas, J.V.; Iosue, C.L.; Shaik, N.F.; Selhorst, K.; He, B.Z.; Wykoff, D.D. Dynamic changes in yeast phosphatase families allow for specialization in phosphate and thiamine starvation. G3 2018, 8, 2333–2343. [Google Scholar] [CrossRef]
- Kornitzer, D. Fungal mechanisms for host iron acquisition. Curr. Opin. Microbiol. 2009, 12, 377–383. [Google Scholar] [CrossRef]
- Weissman, Z.; Kornitzer, D. A family of Candida cell surface haem-binding proteins involved in haemin and haemoglobin-iron utilization. Mol. Microbiol. 2004, 53, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.K.; Suneetha, K.J.; Kaur, R. A systematic analysis reveals an essential role for high-affinity iron uptake system, haemolysin and CFEM domain-containing protein in iron homoeostasis and virulence in Candida glabrata. Biochem. J. 2014, 463, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Nevitt, T.; Thiele, D.J. Host iron withholding demands siderophore utilization for Candida glabrata to survive macrophage killing. PLoS Pathog. 2011, 7, e1001322. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Samaranayake, L.P.; Yau, J.Y.Y. Candida species exhibit differential in vitro hemolytic activities. J. Clin. Microbiol. 2001, 39, 2971–2974. [Google Scholar] [CrossRef] [PubMed]
- Gerwien, F.; Safyan, A.; Wisgott, S.; Brunke, S.; Kasper, L.; Hube, B. The fungal pathogen Candida glabrata does not depend on surface ferric reductases for iron acquisition. Front. Microbiol. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gerwien, F.; Safyan, A.; Wisgott, S.; Hille, F.; Kaemmer, P.; Linde, J.; Brunke, S.; Kasper, L.; Hube, B. A novel hybrid iron regulation network combines features from pathogenic and nonpathogenic yeasts. MBio 2016, 7, e01782-16. [Google Scholar] [CrossRef] [PubMed]
- Merhej, J.; Thiebaut, A.; Blugeon, C.; Pouch, J.; Ali Chaouche, M.E.A.; Camadro, J.M.; Le Crom, S.; Lelandais, G.; Devaux, F. A network of paralogous stress response transcription factors in the human pathogen Candida glabrata. Front. Microbiol. 2016, 7, 645. [Google Scholar] [CrossRef]
- Thiébaut, A.; Delaveau, T.; Benchouaia, M.; Boeri, J.; Garcia, M.; Lelandais, G.; Devaux, F. The CCAAT-binding complex controls respiratory gene expression and iron homeostasis in Candida glabrata. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Li, L.; Dongari-Bagtzoglou, A. Oral epithelium- Candida glabrata interactions in vitro. Oral Microbiol. Immunol. 2007, 22, 182–187. [Google Scholar] [CrossRef]
- Silva, S.; Henriques, M.C.; Hayes, A.; Oliveira, R.; Azeredo, J.; Williams, D.W. Candida glabrata and Candida albicans co-infection of an in vitro oral epithelium. J. Oral Pathol. Med. 2011, 40, 421–427. [Google Scholar] [CrossRef]
- Ho, H.L.; Haynes, K. Candida glabrata: New tools and technologies-expanding the toolkit. FEMS Yeast Res. 2015, 15, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bai, J.; Wu, C.-L.; Wu, Y.; Zhao, Y.; Song, W.-M.; Luo, X.-P. Candida glabrata induced infection of rat tracheal epithelial cells is mediated by TLR-2 induced activation of NF-κB. Microb. Pathog. 2015, 91, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Zajac, D.; Karkowska-Kuleta, J.; Bochenska, O.; Rapala-Kozik, M.; Kozik, A. Interaction of human fibronectin with Candida glabrata epithelial adhesin 6 (Epa6). Acta Biochim. Pol. 2016, 63, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Dongari-Bagtzoglou, A. Epithelial GM-CSF induction by Candida glabrata. J. Dent. Res. 2009, 88, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Pärnänen, P.; Meurman, J.H.; Samaranayake, L.; Virtanen, I. Human oral keratinocyte E-cadherin degradation by Candida albicans and Candida glabrata. J. Oral Pathol. Med. 2010, 39, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Swidergall, M.; Solis, N.V.; Lionakis, M.S.; Filler, S.G. EphA2 is an epithelial cell pattern recognition receptor for fungal β-glucans. Nat. Microbiol. 2018, 3, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torrado, R.; Querol, A. Saccharomyces cerevisiae show low levels of traversal across the human blood brain barrier in vitro. F1000 Res. 2017, 6, 944. [Google Scholar] [CrossRef]
- West, L.; Lowman, D.W.; Mora-Montes, H.M.; Grubb, S.; Murdoch, C.; Thornhill, M.H.; Gow, N.A.R.; Williams, D.; Haynes, K. Differential virulence of Candida glabrata glycosylation mutants. J. Biol. Chem. 2013, 288, 22006–22018. [Google Scholar] [CrossRef]
- Torres-Tirado, D.; Knabb, M.; Castaño, I.; Patrón-Soberano, A.; De Las Peñas, A.; Rubio, R. Candida glabrata binds to glycosylated and lectinic receptors on the coronary endothelial luminal membrane and inhibits flow sense and cardiac responses to agonists. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R24–R32. [Google Scholar] [CrossRef]
- Netea, M.G.; Joosten, L.A.B.; Van Der Meer, J.W.M.; Kullberg, B.J.; Van De Veerdonk, F.L. Immune defense against Candida fungal infections. Nat. Rev. Immunol. 2015, 15, 630–642. [Google Scholar] [CrossRef]
- Johnson, C.J.; Kernien, J.F.; Hoyer, A.R.; Nett, J.E. Mechanisms involved in the triggering of neutrophil extracellular traps (NETs) by Candida glabrata during planktonic and biofilm growth. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Essig, F.; Hünniger, K.; Dietrich, S.; Figge, M.T.; Kurzai, O. Human neutrophils dump Candida glabrata after intracellular killing. Fungal Genet. Biol. 2015, 84, 37–40. [Google Scholar] [CrossRef]
- Duggan, S.; Essig, F.; Hünniger, K.; Mokhtari, Z.; Bauer, L.; Lehnert, T.; Brandes, S.; Häder, A.; Jacobsen, I.D.; Martin, R.; et al. Neutrophil activation by Candida glabrata but not Candida albicans promotes fungal uptake by monocytes. Cell. Microbiol. 2015, 17, 1259–1276. [Google Scholar] [CrossRef] [PubMed]
- Ifrim, D.C.; Bain, J.M.; Reid, D.M.; Oosting, M.; Verschueren, I.; Gow, N.A.R.; van Krieken, J.H.; Brown, G.D.; Kullberg, B.J.; Joosten, L.A.B.; et al. Role of dectin-2 for host defense against systemic infection with Candida glabrata. Infect. Immun. 2014, 82, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Vitenshtein, A.; Charpak-Amikam, Y.; Yamin, R.; Bauman, Y.; Isaacson, B.; Stein, N.; Berhani, O.; Dassa, L.; Gamliel, M.; Gur, C.; et al. NK cell recognition of Candida glabrata through binding of NKp46 and NCR1 to fungal ligands Epa1, Epa6, and Epa7. Cell Host Microbe 2016, 20, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, C.; Majer, O.; Frohner, I.E.; Lesiak-Markowicz, I.; Hildering, K.-S.; Glaser, W.; Stockinger, S.; Decker, T.; Akira, S.; Muller, M.; et al. Conventional dendritic cells mount a Type I IFN response against Candida spp. requiring novel phagosomal TLR7-mediated IFN- Signaling. J. Immunol. 2011, 186, 3104–3112. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.N.; Balusu, S.; Gorityala, N.; Dandu, L.; Kaur, R. Functional genomic analysis of candida glabrata-macrophage interaction: Role of chromatin remodeling in virulence. PLoS Pathog. 2012, 8, e1002863. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.M.; Shen, H.; Zhang, T.; Huang, X.; Liu, X.Q.; Guo, S.Y.; Zhao, J.J.; Wang, C.F.; Yan, L.; Xu, G.T.; et al. Dectin-1 plays an important role in host defense against systemic Candida glabrata infection. Virulence 2017, 5594, 1–14. [Google Scholar] [CrossRef]
- Kasper, L.; Seider, K.; Gerwien, F.; Allert, S.; Brunke, S.; Schwarzmüller, T.; Ames, L.; Zubiria-Barrera, C.; Mansour, M.K.; Becken, U.; et al. Identification of Candida glabrata genes involved in pH modulation and modification of the phagosomal environment in macrophages. PLoS ONE 2014, 9, e96015. [Google Scholar] [CrossRef]
- Seider, K.; Gerwien, F.; Kasper, L.; Allert, S.; Brunke, S.; Jablonowski, N.; Schwarzmüller, T.; Barz, D.; Rupp, S.; Kuchler, K.; et al. Immune evasion, stress resistance, and efficient nutrient acquisition are crucial for intracellular survival of Candida glabrata within macrophages. Eukaryot. Cell 2014, 13, 170–183. [Google Scholar] [CrossRef]
- Brunke, S.; Seider, K.; Fischer, D.; Jacobsen, I.D.; Kasper, L.; Jablonowski, N.; Wartenberg, A.; Bader, O.; Enache-Angoulvant, A.; Schaller, M.; et al. One small step for a yeast—Microevolution within macrophages renders Candida glabrata hypervirulent due to a single point mutation. PLoS Pathog. 2014, 10, e1004478. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of pleiotropic defects associated with lack of CgYapsins. The Cgyps1-11Δ mutant displays increased ROS and diminished ATP levels, enlarged acidic vacuole with higher amount of polyphosphate, missorting of the vacuolar carboxypeptidase Y to the medium and constitutive activation of the protein kinase C-mediated cell wall integrity (CWI) pathway. CgSlt2 is the terminal MAPK of the CWI pathway. Furthermore, the cell wall of the Cgyps1-11Δ mutant contains lower amounts of β-glucan and mannan, and higher amount of chitin. Processing of the adhesin Epa1 from the cell wall is also reduced in the Cgyps1-11Δ mutant. The altered cell wall composition in the mutant may contribute to diminished biofilm formation, increased activation of macrophages and reduced colonization and virulence in the systemic candidiasis model. In line with mutant phenotypes, CgYPS genes are induced in response to diverse stresses including macrophage internalization and thermal, pH and cell wall stress. V, M, N and CW indicate vacuole, mitochondria, nucleus and cell wall, respectively.
Figure 1.
Schematic representation of pleiotropic defects associated with lack of CgYapsins. The Cgyps1-11Δ mutant displays increased ROS and diminished ATP levels, enlarged acidic vacuole with higher amount of polyphosphate, missorting of the vacuolar carboxypeptidase Y to the medium and constitutive activation of the protein kinase C-mediated cell wall integrity (CWI) pathway. CgSlt2 is the terminal MAPK of the CWI pathway. Furthermore, the cell wall of the Cgyps1-11Δ mutant contains lower amounts of β-glucan and mannan, and higher amount of chitin. Processing of the adhesin Epa1 from the cell wall is also reduced in the Cgyps1-11Δ mutant. The altered cell wall composition in the mutant may contribute to diminished biofilm formation, increased activation of macrophages and reduced colonization and virulence in the systemic candidiasis model. In line with mutant phenotypes, CgYPS genes are induced in response to diverse stresses including macrophage internalization and thermal, pH and cell wall stress. V, M, N and CW indicate vacuole, mitochondria, nucleus and cell wall, respectively.

Figure 2.
Schematic representation of the mating-type (MAT)-like loci in C. glabrata (A), S. cerevisiae (B) and C. albicans (C). C. glabrata and S. cerevisiae has three mating-type loci while C. albicans has one mating-type locus. MTL1, MAT and MTL loci determine the strain mating type in C. glabrata, S. cerevisiae and C. albicans, respectively. The other two mating-type loci in C. glabrata (MTL2 and MTL3) and S. cerevisiae (HMR and HML), encoding a and α information, remain silent. S. cerevisiae has all three mating-type loci on the chromosome III, while the C. albicans MTL locus is present on the chromosome 5. In contrast, C. glabrata mating-type-like loci exist on two different chromosomes with CgMTL1 and CgMTL3, and CgMTL2 being present on the chromosome B and E, respectively. S. cerevisiae and C. glabrata have HO endonuclease site within the α1 gene while C. albicans lack HO site.
Figure 2.
Schematic representation of the mating-type (MAT)-like loci in C. glabrata (A), S. cerevisiae (B) and C. albicans (C). C. glabrata and S. cerevisiae has three mating-type loci while C. albicans has one mating-type locus. MTL1, MAT and MTL loci determine the strain mating type in C. glabrata, S. cerevisiae and C. albicans, respectively. The other two mating-type loci in C. glabrata (MTL2 and MTL3) and S. cerevisiae (HMR and HML), encoding a and α information, remain silent. S. cerevisiae has all three mating-type loci on the chromosome III, while the C. albicans MTL locus is present on the chromosome 5. In contrast, C. glabrata mating-type-like loci exist on two different chromosomes with CgMTL1 and CgMTL3, and CgMTL2 being present on the chromosome B and E, respectively. S. cerevisiae and C. glabrata have HO endonuclease site within the α1 gene while C. albicans lack HO site.

Figure 3.
A schematic depicting major iron acquisition and homeostasis pathways in C. glabrata. The iron acquisition mechanisms of C. glabrata include the high-affinity iron uptake system mainly comprised of the CgFtr1 permease and the CgFet3 ferroxidase, and the siderophore uptake system consisting of the sole xenosiderophore transporter CgSit1. In response to low-iron, genes of the reductive iron acquisition system are upregulated in the transcriptional factor CgAft1/2-dependent manner. Additionally, the CgCth2 protein, in an iron-limited environment, degrades mRNAs that code for iron-consuming enzymes. Contrarily, C. glabrata responds to high iron by retrograde trafficking of the components of the high-affinity iron uptake system from the plasma membrane. CgFtr1 is transported to the vacuole in a phosphoinositide-3-kinase (CgVps34)-dependent manner, while CgFet3 is trafficked to an intracellular organelle. The factor/s responsible for the retrograde trafficking of CgFet3 are unknown. In addition, the transcriptional factor CgYap5 regulates genes involved in the iron-surplus response, and iron-sulfur cluster biogenesis is upregulated in the high-iron environment. The functions of ferric reductases (CgFre) are yet to be deciphered, as C. glabrata does not exhibit cell surface-associated ferric reductase activity.
Figure 3.
A schematic depicting major iron acquisition and homeostasis pathways in C. glabrata. The iron acquisition mechanisms of C. glabrata include the high-affinity iron uptake system mainly comprised of the CgFtr1 permease and the CgFet3 ferroxidase, and the siderophore uptake system consisting of the sole xenosiderophore transporter CgSit1. In response to low-iron, genes of the reductive iron acquisition system are upregulated in the transcriptional factor CgAft1/2-dependent manner. Additionally, the CgCth2 protein, in an iron-limited environment, degrades mRNAs that code for iron-consuming enzymes. Contrarily, C. glabrata responds to high iron by retrograde trafficking of the components of the high-affinity iron uptake system from the plasma membrane. CgFtr1 is transported to the vacuole in a phosphoinositide-3-kinase (CgVps34)-dependent manner, while CgFet3 is trafficked to an intracellular organelle. The factor/s responsible for the retrograde trafficking of CgFet3 are unknown. In addition, the transcriptional factor CgYap5 regulates genes involved in the iron-surplus response, and iron-sulfur cluster biogenesis is upregulated in the high-iron environment. The functions of ferric reductases (CgFre) are yet to be deciphered, as C. glabrata does not exhibit cell surface-associated ferric reductase activity.

Figure 4.
A schematic illustrating major facets of C. glabrata-macrophage interaction. C. glabrata cells are internalized by macrophages probably via the Dectin receptor-mediated endocytosis. The C. glabrata Epa1 adhesin also facilitates binding to macrophages. After phagocytosis, C. glabrata first resides in the early endosome [decorated with EEA1 (Early Endosome Antigen 1)] and later in the late endosome [marked with LAMP1 (Lysosomal-associated membrane protein 1)], with cell replication occurring in the unacidified phagolysosome. During the course of intracellular proliferation, C. glabrata remodels its chromatin, activates autophagy and upregulates genes involved in the β-oxidation of fatty acids, gluconeogenesis, glyoxylate cycle and oxidative stress. Metabolic and stress pathways reconfiguration helps C. glabrata survive the macrophage internal milieu which is limited for iron and glucose, and contains high levels of ROS. On the other hand, the infected macrophage weakly activates the Dectin-mediated Syk signaling through Syk phosphorylation, and secretes small amount of the pro-inflammatory cytokine IL-1β in a Syk-NLRP3 inflammasome-dependent manner. The transcriptional response of the macrophage to C. glabrata internalization primarily consists of induction of genes involved in signal transduction and negative regulation of the inflammatory response and cytokine secretion processes.
Figure 4.
A schematic illustrating major facets of C. glabrata-macrophage interaction. C. glabrata cells are internalized by macrophages probably via the Dectin receptor-mediated endocytosis. The C. glabrata Epa1 adhesin also facilitates binding to macrophages. After phagocytosis, C. glabrata first resides in the early endosome [decorated with EEA1 (Early Endosome Antigen 1)] and later in the late endosome [marked with LAMP1 (Lysosomal-associated membrane protein 1)], with cell replication occurring in the unacidified phagolysosome. During the course of intracellular proliferation, C. glabrata remodels its chromatin, activates autophagy and upregulates genes involved in the β-oxidation of fatty acids, gluconeogenesis, glyoxylate cycle and oxidative stress. Metabolic and stress pathways reconfiguration helps C. glabrata survive the macrophage internal milieu which is limited for iron and glucose, and contains high levels of ROS. On the other hand, the infected macrophage weakly activates the Dectin-mediated Syk signaling through Syk phosphorylation, and secretes small amount of the pro-inflammatory cytokine IL-1β in a Syk-NLRP3 inflammasome-dependent manner. The transcriptional response of the macrophage to C. glabrata internalization primarily consists of induction of genes involved in signal transduction and negative regulation of the inflammatory response and cytokine secretion processes.


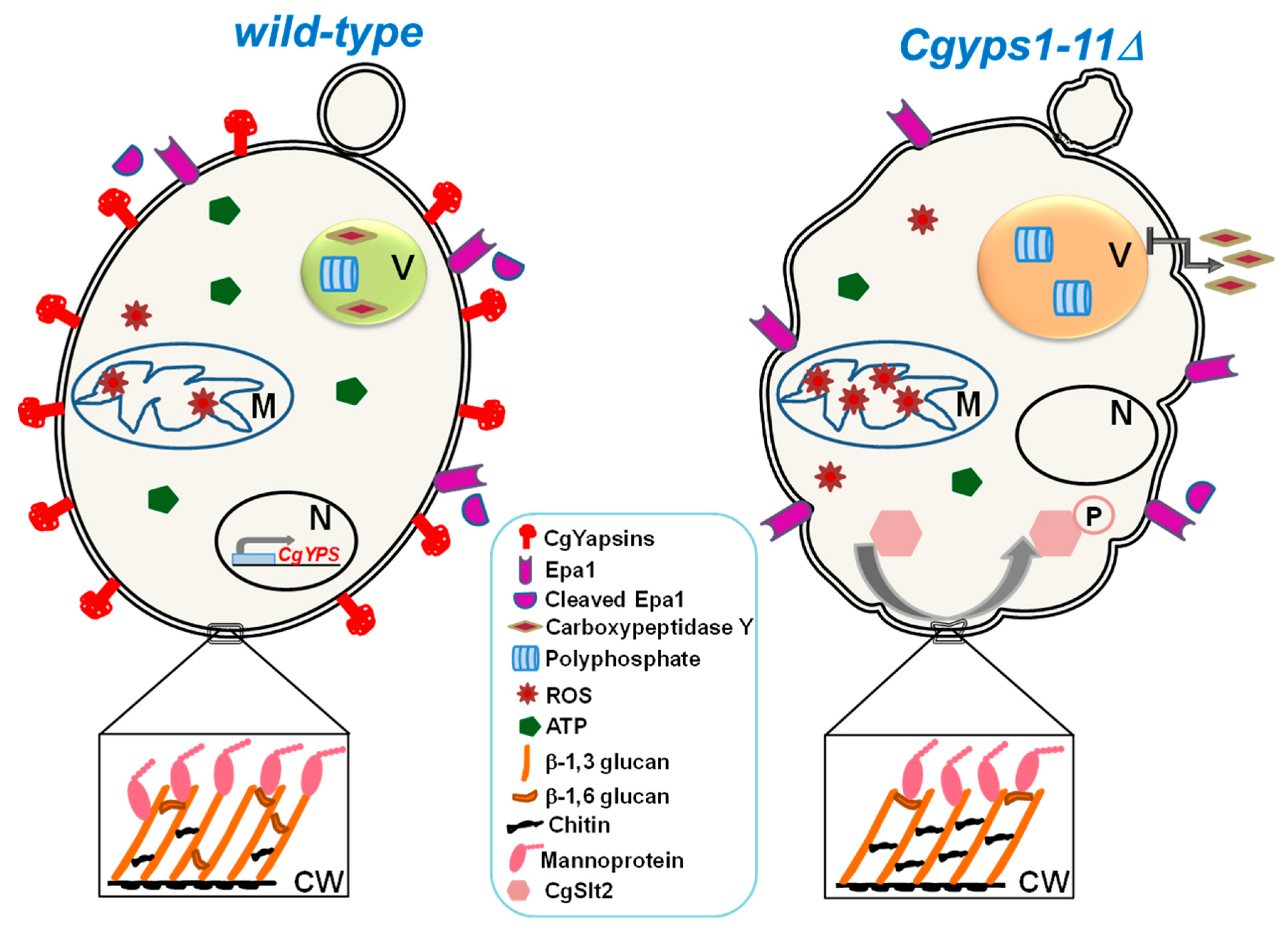{kind=link}
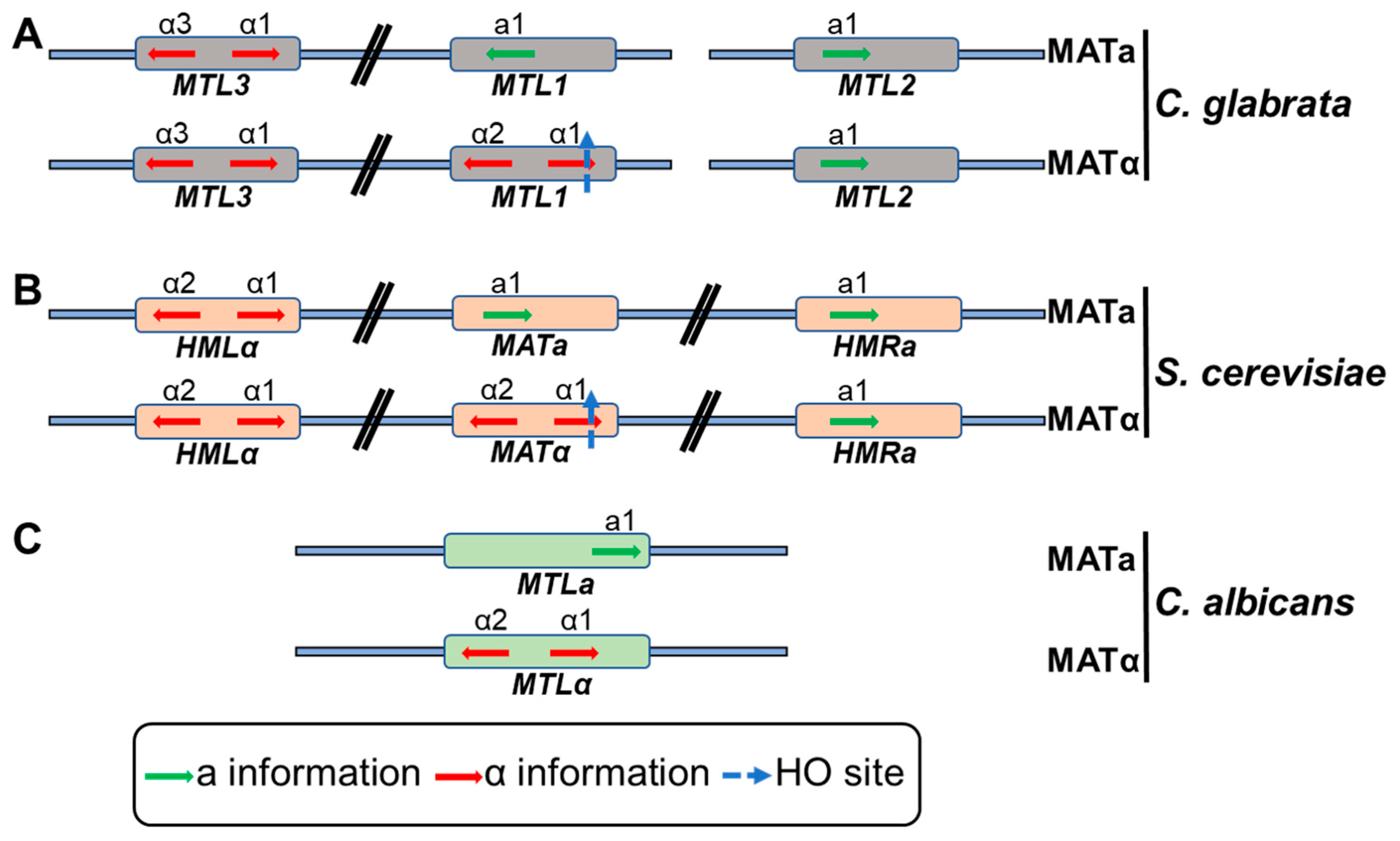{kind=link}
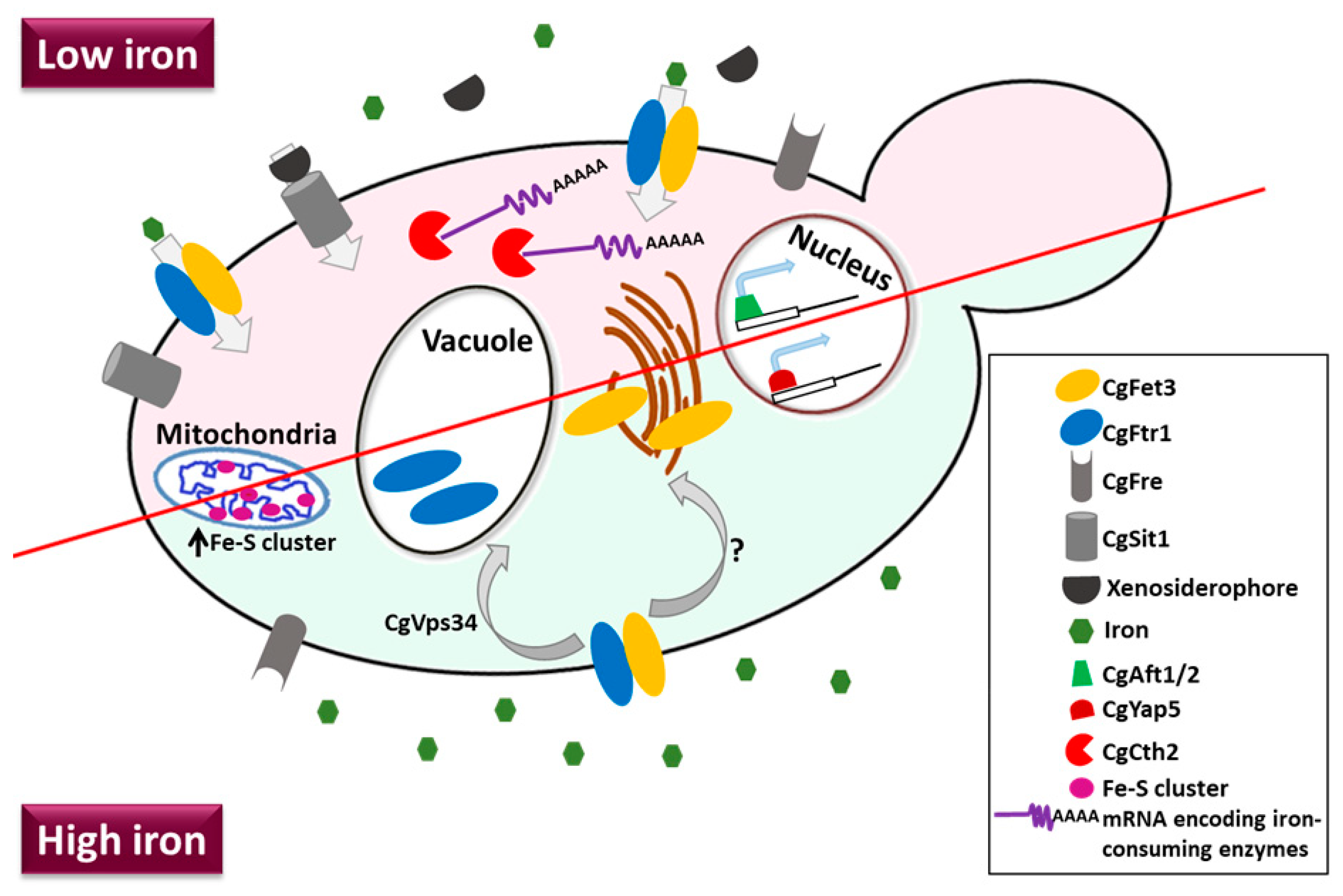{kind=link}
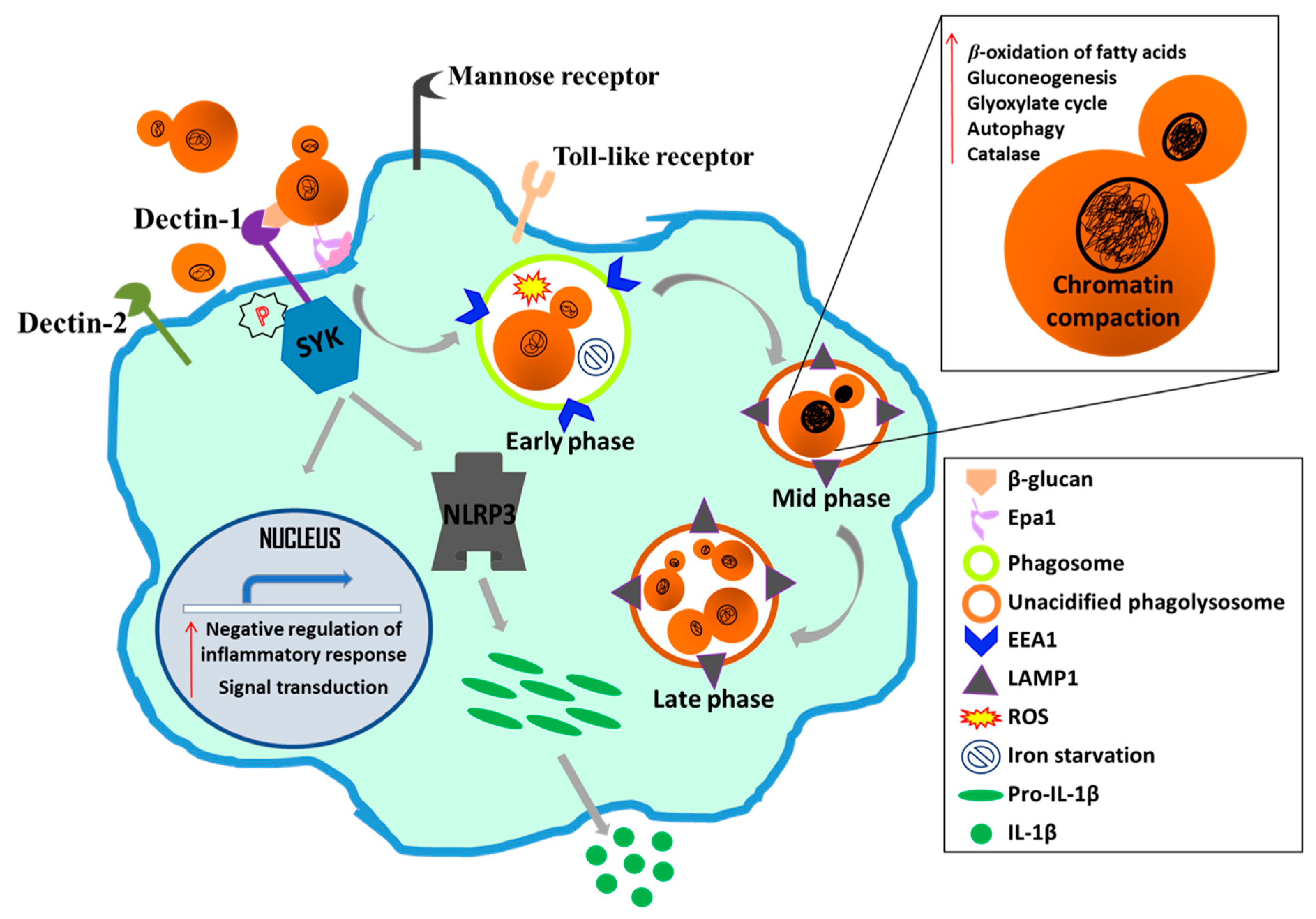{kind=link}
Table 1.
Comparison of morphological and pathogenesis traits of C. glabrata and C. albicans.
| Feature | Candida glabrata | Candida albicans |
|---|---|---|
| Ploidy | Haploid | Diploid |
| Cellular morphology | Yeast | Yeast, pseudohyphae and hyphae |
| Cell size | 1–4 µm | 4–6 µm |
| Phylogeny | Non-CTG clade | CTG clade |
| Phenotypic switching | Present | Present |
| Carbon assimilation | Glucose and trehalose | Glucose, trehalose, maltose and galactose |
| Auxotrophy | Niacin, thiamine, pyridoxine | None |
| Crab tree | Positive | Negative |
| Mitochondrial function | Petite positive | Petite negative |
| Mating genes | Present | Present |
| Haem receptor | Absent | Present |
| Haemoglobin and transferrin utilization | Absent | Present |
| Innate azole resistance | Present | Absent |
| Secretory aspartyl proteases | Absent | Present |
| Life style | Probably commensal, and pathogenic | Commensal and pathogenic |
| Major sites of infection | Vaginal, oral, disseminated | Vaginal, oral, disseminated |
| Major adhesins | Lectins (Epa) | Lectins (Als and Hwp) |
| Biofilm | Present | Present |
| Invasion | Not known | Induced endocytosis and active penetration |
| Damage to host cells | No significant damage | Substantial damage |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kumar, K.; Askari, F.; Sahu, M.S.; Kaur, R. Candida glabrata: A Lot More Than Meets the Eye. Microorganisms 2019, 7, 39. https://doi.org/10.3390/microorganisms7020039
AMA Style
Kumar K, Askari F, Sahu MS, Kaur R. Candida glabrata: A Lot More Than Meets the Eye. Microorganisms. 2019; 7(2):39. https://doi.org/10.3390/microorganisms7020039
Chicago/Turabian StyleKumar, Kundan, Fizza Askari, Mahima Sagar Sahu, and Rupinder Kaur. 2019. "Candida glabrata: A Lot More Than Meets the Eye" Microorganisms 7, no. 2: 39. https://doi.org/10.3390/microorganisms7020039
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.