
Piezophilic Phenotype Is Growth Condition Dependent and Correlated with the Regulation of Two Sets of ATPase in Deep-Sea Piezophilic Bacterium Photobacterium profundum SS9
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Construction of Gene Deletion Mutant
2.3. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR)
2.4. Quantification of Intracellular ATP
3. Results
3.1. Organization of Two Sets of ATPase Operons in the Genome of Strain SS9R
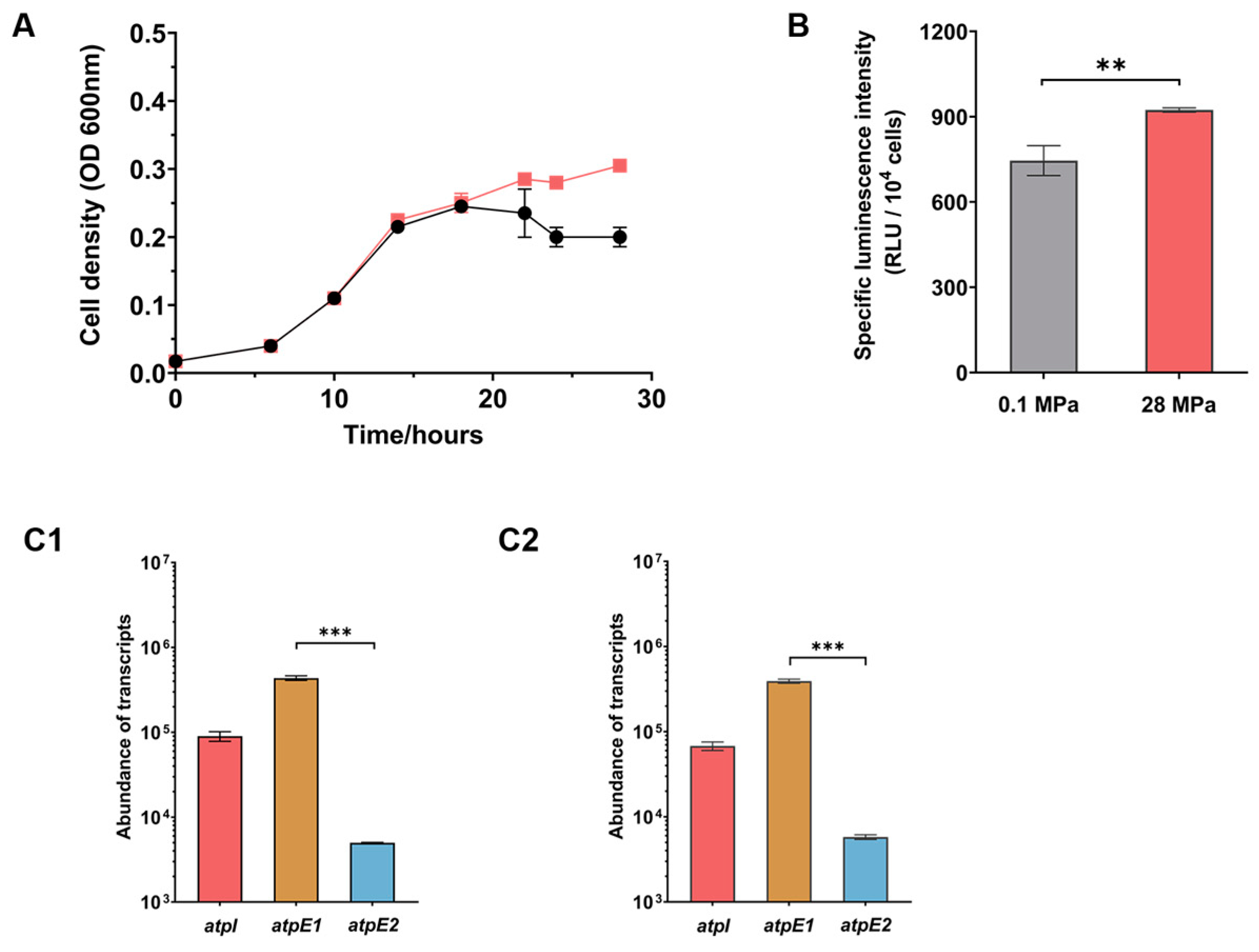
3.2. ATPase-I Acts As the Dominant ATPase under Conventional Cultural Condition
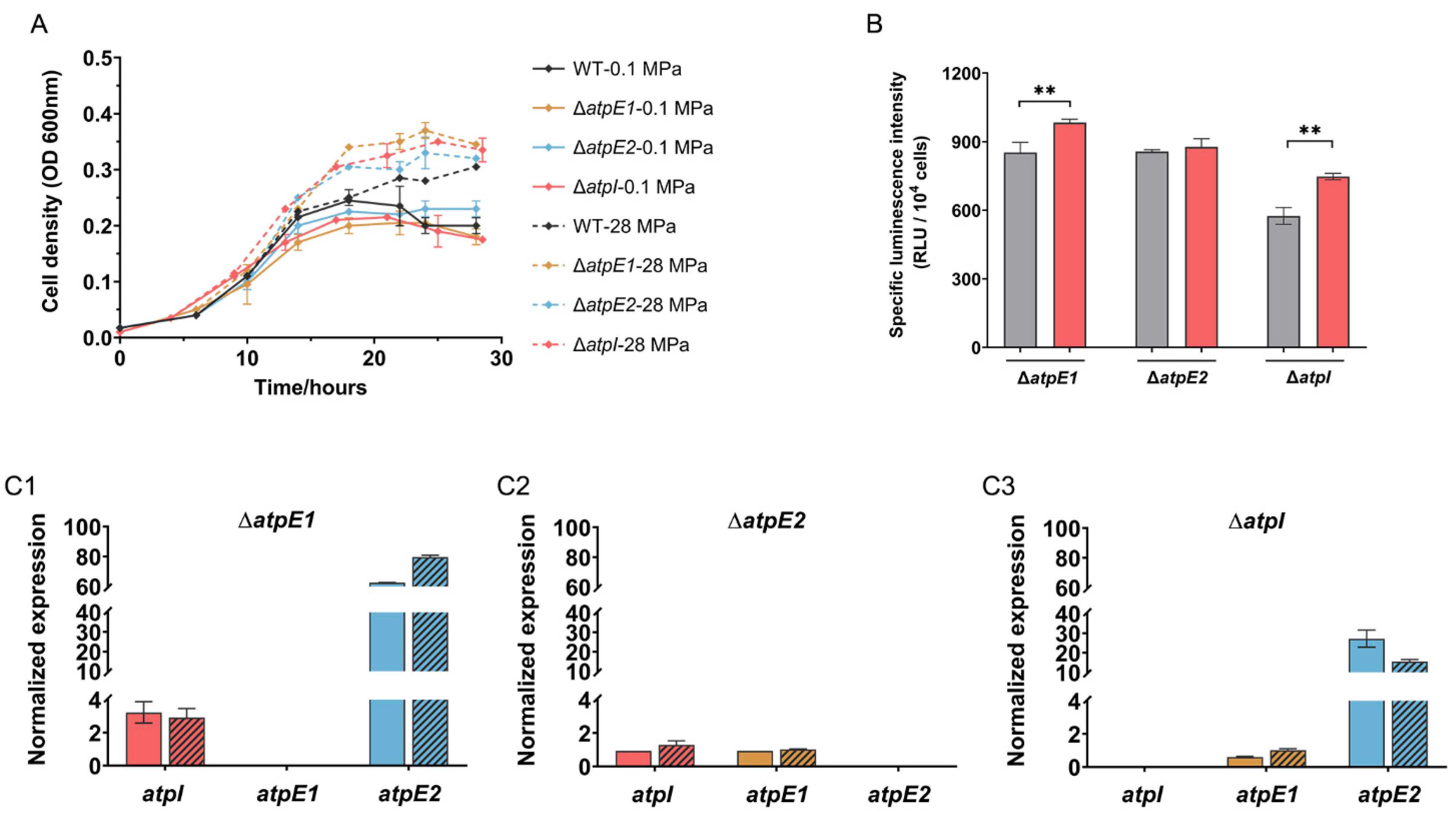
3.3. Impairment in ATPase-I Results in Up-regulated Expression of ATPase-II
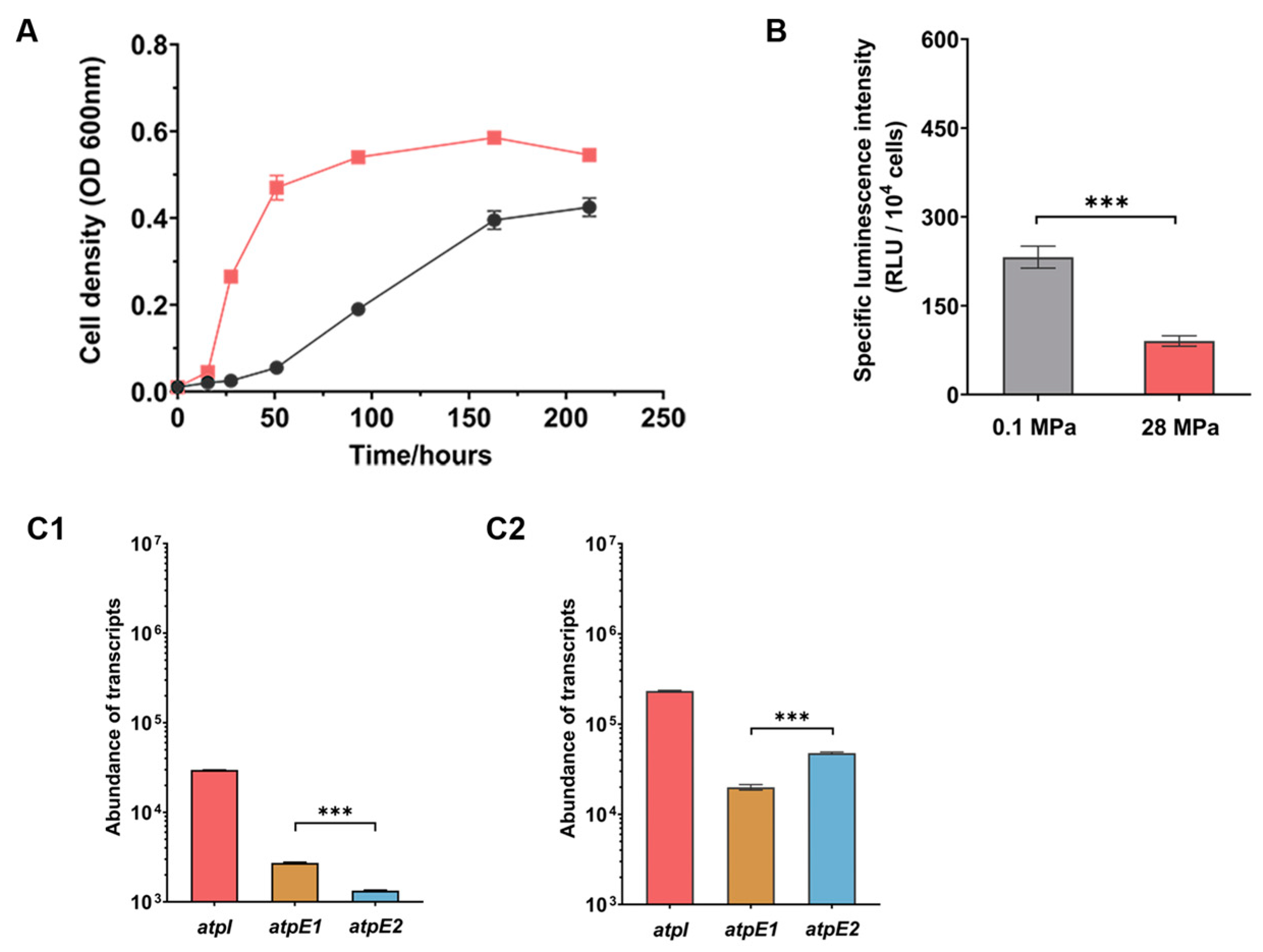
3.4. SS9R Cells Had a More Pronounced Piezophilic Phenotype When Cultivated in MG Medium
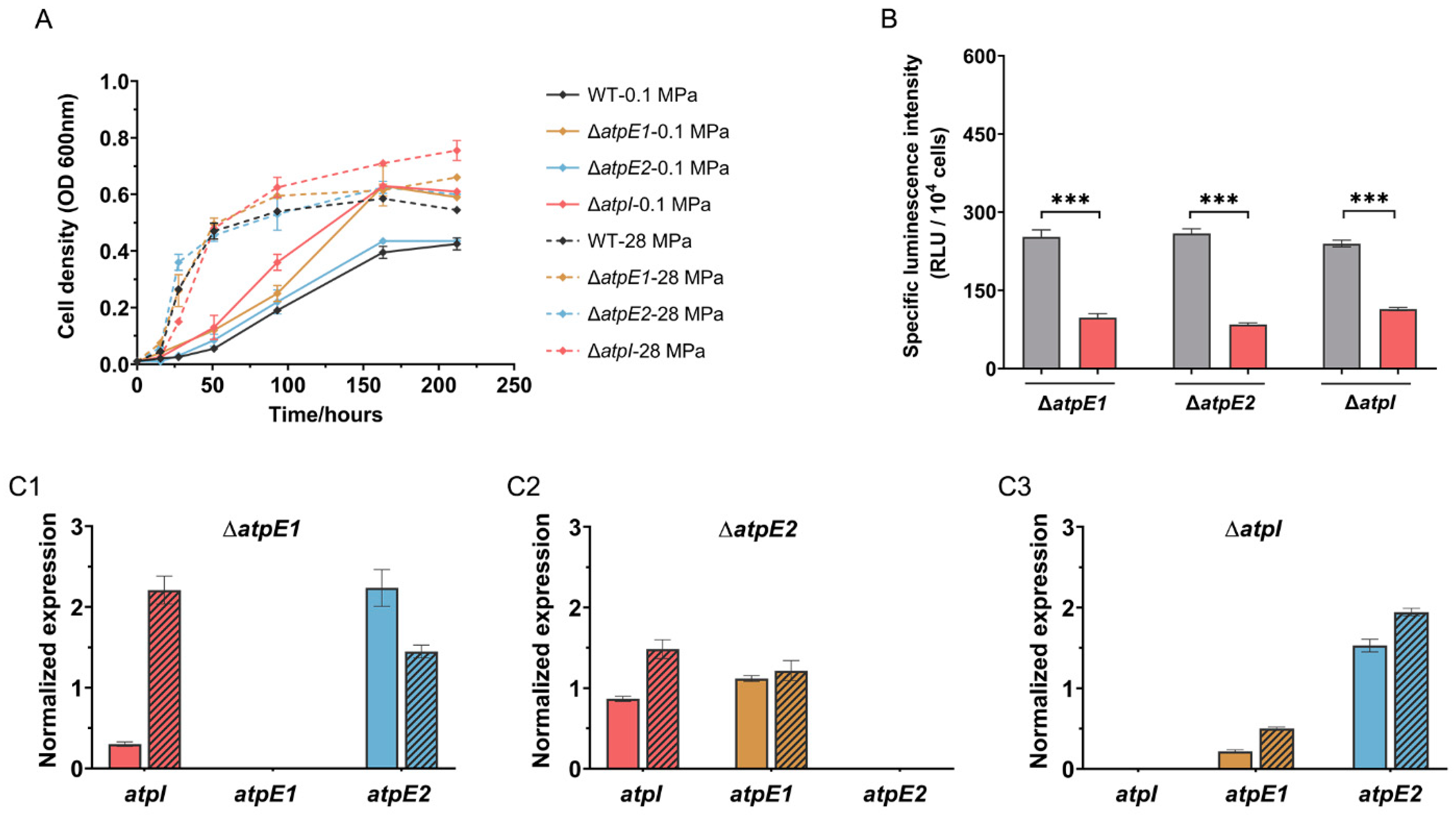
3.5. ATPase-II Is Preferred When Cultivated in MG Medium
3.6. Mutated ATPase-I Improved Growth at 0.1 MPa in MG Medium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bartlett, D.H. Pressure effects on in vivo microbial processes. Biochim. Biophys. Acta 2002, 1595, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Gross, M.; Jaenicke, R. Pressure-induced dissociation of tight couple ribosomes. FEBS Lett. 1990, 267, 239–241. [Google Scholar] [CrossRef] [Green Version]
- Balny, C.; Masson, P.; Heremans, K. High pressure effects on biological macromolecules: From structural changes to alteration of cellular processes. Biochim. Biophys. Acta 2002, 1595, 3–10. [Google Scholar] [CrossRef]
- Kato, C.; Nogi, Y.; Arakawa, S. Isolation, Cultivation, and Diversity of Deep-Sea Piezophiles. In High-Pressure Microbiology; American Society of Microbiology: Washington, DC, USA, 2008. [Google Scholar] [CrossRef]
- Amrani, A.; Bergon, A.; Holota, H.; Tamburini, C.; Garel, M.; Ollivier, B.; Imbert, J.; Dolla, A.; Pradel, N. Transcriptomics Reveal Several Gene Expression Patterns in the Piezophile Desulfovibrio hydrothermalis in Response to Hydrostatic Pressure. PLoS ONE 2014, 9, e106831. [Google Scholar] [CrossRef] [PubMed]
- Amrani, A.; van Helden, J.; Bergon, A.; Aouane, A.; Ben Hania, W.; Tamburini, C.; Loriod, B.; Imbert, J.; Ollivier, B.; Pradel, N.; et al. Deciphering the adaptation strategies of Desulfovibrio piezophilus to hydrostatic pressure through metabolic and transcriptional analyses. Environ. Microbiol. Rep. 2016, 8, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Ohke, Y.; Sakoda, A.; Kato, C.; Sambongi, Y.; Kawamoto, J.; Kurihara, T.; Tamegai, H. Regulation of cytochrome c- and quinol oxidases, and piezotolerance of their activities in the deep-sea piezophile Shewanella violacea DSS12 in response to growth conditions. Biosci. Biotechnol. Biochem. 2013, 77, 1522–1528. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Nakasone, K.; Tamegai, H.; Kato, C.; Usami, R.; Horikoshi, K. Pressure regulation of soluble cytochromes c in a deep-sea piezophilic bacterium, Shewanella violacea. J. Bacteriol. 2000, 182, 2945–2952. [Google Scholar] [CrossRef] [Green Version]
- Tamegai, H.; Kawano, H.; Ishii, A.; Chikuma, S.; Nakasone, K.; Kato, C. Pressure-regulated biosynthesis of cytochrome bd in piezo- and psychrophilic deep-sea bacterium Shewanella violacea DSS12. Extremophiles 2005, 9, 247–253. [Google Scholar] [CrossRef]
- Kilic, V.; Kilic, G.A.; Kutlu, H.M.; Martinez-Espinosa, R.M. Nitrate reduction in Haloferax alexandrinus: The case of assimilatory nitrate reductase. Extremophiles 2017, 21, 551–561. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, F.P.; Xu, J.; Mehmood, M.A.; Xiao, X. Physiological and evolutionary studies of NAP systems in Shewanella piezotolerans WP3. ISME J. 2011, 5, 843–855. [Google Scholar] [CrossRef] [Green Version]
- Li, X.G.; Zhang, W.J.; Xiao, X.; Jian, H.H.; Jiang, T.; Tang, H.Z.; Qi, X.Q.; Wu, L.F. Pressure-Regulated Gene Expression and Enzymatic Activity o the Two Periplasmic Nitrate Reductases in the Deep-Sea Bacterium Shewanella piezotolerans WP3. Front. Microbiol. 2018, 9, 3173. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.; Jian, H.H.; Zhang, Y.X.; Xiao, X. The Two Sets of DMSO Respiratory Systems of Shewanella piezotolerans WP3 Are Involved in Deep Sea Environmental Adaptation. Front. Microbiol. 2016, 7, 1418. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.; Jian, H.H.; Xiao, X. Deep-Sea Bacterium Shewanella piezotolerans WP3 Has Two Dimethyl Sulfoxide Reductases in Distinct Subcellular Locations. Appl. Environ. Microbiol. 2017, 83, e01262-17. [Google Scholar] [CrossRef] [Green Version]
- Brandt, K.; Muller, D.B.; Hoffmann, J.; Hubert, C.; Brutschy, B.; Deckers-Hebestreit, G.; Muller, V. Functional production of the Na+ F1F(O) ATP synthase from Acetobacterium woodii in Escherichia coli requires the native AtpI. J. Bioenerg. Biomembr. 2013, 45, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Dreyfus, G.; Guimaraes-Motta, H.; Silva, J.L. Effect of hydrostatic pressure on the mitochondrial ATP synthase. Biochem. -Us 1988, 27, 6704–6710. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.O.; Creczynski-Pasa, T.B.; Scofano, H.M.; Graber, P.; Mignaco, J.A. High hydrostatic pressure perturbs the interactions between CF0F1 subunits and induces a dual effect on activity. Int. J. Biochem. Cell Biol. 2004, 36, 920–930. [Google Scholar] [CrossRef]
- Okuno, D.; Nishiyama, M.; Noji, H. Single-Molecule Analysis of the Rotation of F1-ATPase under High Hydrostatic Pressure. Biophys. J. 2014, 106, 393A. [Google Scholar] [CrossRef] [Green Version]
- Nogi, Y.; Masui, N.; Kato, C. Photobacterium profundum sp. nov., a new, moderately barophilic bacterial species isolated from a deep-sea sediment. Extremophiles 1998, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- DeLong, E.F.; Franks, D.G.; Yayanos, A.A. Evolutionary relationships of cultivated psychrophilic and barophilic deep-sea bacteria. Appl. Environ. Microb. 1997, 63, 2105–2108. [Google Scholar] [CrossRef] [Green Version]
- Vezzi, A.; Campanaro, S.; D’Angelo, M.; Simonato, F.; Vitulo, N.; Lauro, F.M.; Cestaro, A.; Malacrida, G.; Simionati, B.; Cannata, N.; et al. Life at depth: Photobacterium profundum genome sequence and expression analysis. Science 2005, 307, 1459–1461. [Google Scholar] [CrossRef] [Green Version]
- Lauro, F.M.; Eloe, E.A.; Liverani, N.; Bertoloni, G.; Bartlett, D.H. Conjugal vectors for cloning, expression, and insertional mutagenesis in gram-negative bacteria. Biotechniques 2005, 38, 708–712. [Google Scholar] [CrossRef]
- Welch, T.J.; Bartlett, D.H. Identification of a regulatory protein required for pressure-responsive gene expression in the deep-sea bacterium Photobacterium species strain SS9. Mol. Microbiol. 1998, 27, 977–985. [Google Scholar] [CrossRef]
- Eloe, E.A.; Lauro, F.M.; Vogel, R.F.; Bartlett, D.H. The deep-sea bacterium Photobacterium profundum SS9 utilizes separate flagellar systems for swimming and swarming under high-pressure conditions. Appl. Environ. Microbiol. 2008, 74, 6298–6305. [Google Scholar] [CrossRef] [Green Version]
- El-Hajj, Z.W.; Allcock, D.; Tryfona, T.; Lauro, F.M.; Sawyer, L.; Bartlett, D.H.; Ferguson, G.P. Insights into piezophily from genetic studies on the deep-sea bacterium, Photobacterium profundum SS9. Ann. N. Y. Acad. Sci. 2010, 1189, 143–148. [Google Scholar] [CrossRef]
- Campanaro, S.; Pascale, F.D.; Telatin, A.; Schiavon, R.; Bartlett, D.H.; Valle, G. The transcriptional landscape of the deep-sea bacterium Photobacterium profundum in both a toxR mutant and its parental strain. BMC Genomics 2012, 13, 567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamegai, H.; Nishikawa, S.; Haga, M.; Bartlett, D.H. The respiratory system of the piezophile Photobacterium profundum SS9 grown under various pressures. Biosci. Biotechnol. Biochem. 2012, 76, 1506–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.D.; Santini, C.L.; Zhang, W.J.; Barbe, V.; Mangenot, S.; Guyomar, C.; Garel, M.; Chen, H.T.; Li, X.G.; Yin, Q.J.; et al. Genomic and physiological analysis reveals versatile metabolic capacity of deep-sea Photobacterium phosphoreum ANT-2200. Extremophiles 2016, 20, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Chi, E.; Bartlett, D.H. Use of a reporter gene to follow high-pressure signal transduction in the deep-sea bacterium Photobacterium sp. strain SS9. J. Bacteriol. 1993, 175, 7533–7540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufault-Thompson, K.; Jian, H.; Cheng, R.; Li, J.; Wang, F.; Zhang, Y. A Genome-Scale Model of Shewanella piezotolerans Simulates Mechanisms of Metabolic Diversity and Energy Conservation. Msystems 2017, 2, e00165-16. [Google Scholar] [CrossRef] [Green Version]
- Yin, Q.J.; Zhang, W.J.; Qi, X.Q.; Zhang, S.D.; Jiang, T.; Li, X.G.; Chen, Y.; Santini, C.L.; Zhou, H.; Chou, I.M.; et al. High Hydrostatic Pressure Inducible Trimethylamine N-Oxide Reductase Improves the Pressure Tolerance of Piezosensitive Bacteria Vibrio fluvialis. Front. Microbiol. 2018, 8, 2646. [Google Scholar] [CrossRef] [Green Version]
- Le Bihan, T.; Rayner, J.; Roy, M.M.; Spagnolo, L. Photobacterium profundum under pressure: A MS-based label-free quantitative proteomics study. PLoS ONE 2013, 8, e60897. [Google Scholar] [CrossRef] [Green Version]
- Urbanczyk, H.; Ast, J.C.; Dunlap, P.V. Phylogeny, genomics, and symbiosis of Photobacterium. FEMS Microbiol. Rev. 2011, 35, 324–342. [Google Scholar] [CrossRef] [PubMed]
- Cooper, V.S.; Vohr, S.H.; Wrocklage, S.C.; Hatcher, P.J. Why genes evolve faster on secondary chromosomes in bacteria. PLoS Comput. Biol. 2010, 6, e1000732. [Google Scholar] [CrossRef]
- Suzuki, T.; Ozaki, Y.; Sone, N.; Feniouk, B.A.; Yoshida, M. The product of uncI gene in F1Fo-ATP synthase operon plays a chaperone-like role to assist c-ring assembly. Proc. Natl. Acad. Sci. USA 2007, 104, 20776–20781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gay, N.J. Construction and characterization of an Escherichia coli strain with a uncI mutation. J. Bacteriol. 1984, 158, 820–825. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, Y.; Suzuki, T.; Kuruma, Y.; Ueda, T.; Yoshida, M. UncI protein can mediate ring-assembly of c-subunits of FoF1-ATP synthase in vitro. Biochem. Biophys. Res. Commun. 2008, 367, 663–666. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SS9R | ΔatpE1 | ΔatpE2 | Δ atpI | |||||
|---|---|---|---|---|---|---|---|---|
| 0.1 MPa | 28 MPa | 0.1 MPa | 28 MPa | 0.1 MPa | 28 MPa | 0.1 MPa | 28 MPa | |
| atpI | 20.13 ± 1.06 #,*** | 3.58 ± 0.19 *** | 1.89 ± 0.18 * | 2.75 ± 0.21 *** | 6.77 ± 0.23 *** | 4.18 ± 0.32 *** | - | - |
| atpE1 | 0.52 ± 0.05 ** | 0.23 ± 0.01 *** | - | - | 0.62 ± 0.02 *** | 0.27 ± 0.03 *** | 0.19 ± 0.02 ** | 0.11 ± 0.00 *** |
| atpE2 | 8.48 ± 1.32 *** | 9.08 ± 0.97 *** | 0.46 ± 0.15 | 0.16 ± 0.01 *** | - | - | 0.48 ± 0.02 ** | 1.14 ± 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, A.-Q.; Zhang, W.-J.; Li, X.-G.; Bao, X.-C.; Qi, X.-Q.; Wu, L.-F.; Bartlett, D.H. Piezophilic Phenotype Is Growth Condition Dependent and Correlated with the Regulation of Two Sets of ATPase in Deep-Sea Piezophilic Bacterium Photobacterium profundum SS9. Microorganisms 2023, 11, 637. https://doi.org/10.3390/microorganisms11030637
Li A-Q, Zhang W-J, Li X-G, Bao X-C, Qi X-Q, Wu L-F, Bartlett DH. Piezophilic Phenotype Is Growth Condition Dependent and Correlated with the Regulation of Two Sets of ATPase in Deep-Sea Piezophilic Bacterium Photobacterium profundum SS9. Microorganisms. 2023; 11(3):637. https://doi.org/10.3390/microorganisms11030637
Chicago/Turabian StyleLi, An-Qi, Wei-Jia Zhang, Xue-Gong Li, Xu-Chong Bao, Xiao-Qing Qi, Long-Fei Wu, and Douglas H. Bartlett. 2023. "Piezophilic Phenotype Is Growth Condition Dependent and Correlated with the Regulation of Two Sets of ATPase in Deep-Sea Piezophilic Bacterium Photobacterium profundum SS9" Microorganisms 11, no. 3: 637. https://doi.org/10.3390/microorganisms11030637