

Cryptococcal Immune Reconstitution Inflammatory Syndrome: From Clinical Studies to Animal Experiments

Abstract
:1. Introduction
2. Inflammatory Responses in the CNS during HIV-Associated Cryptococcal Meningoencephalitis
3. Cryptococcal IRIS in HIV/AIDS Patients
4. Cryptococcal PIIRS in HIV-Negative Patients
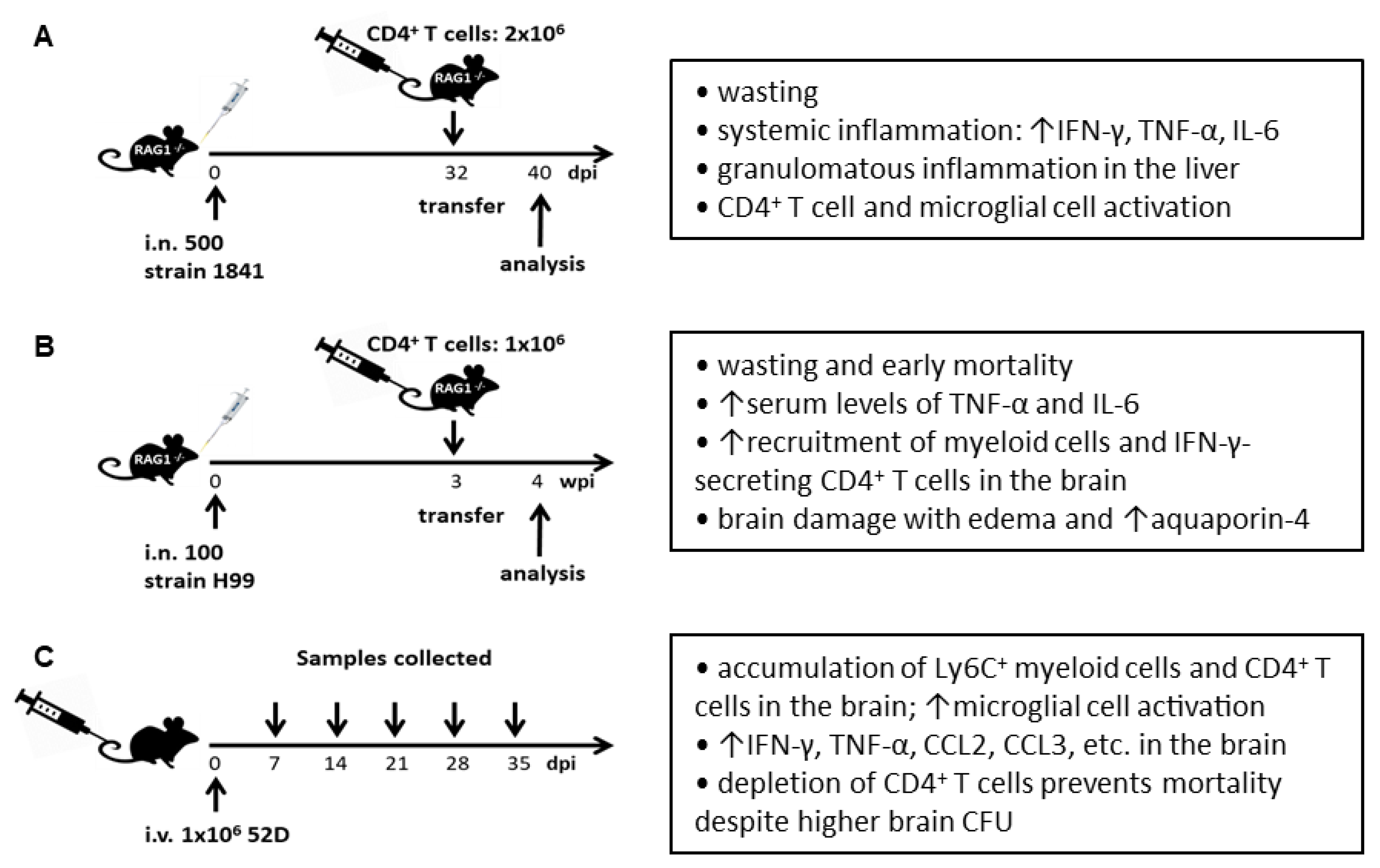
5. Cryptococcal IRIS in Murine Models
6. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Gottfredsson, M.; Perfect, J.R. Fungal meningitis. Semin. Neurol. 2000, 20, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Sorrell, T.C.; Dromer, F.; Fung, E.; Levitz, S.M. Cryptococcosis: Clinical and biological aspects. Med. Mycol. 2000, 38 (Suppl. S1), 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, T.G.; Perfect, J.R. Cryptococcosis in the era of AIDS—100 years after the discovery of Cryptococcus neoformans. Clin. Microbiol. Rev. 1995, 8, 515–548. [Google Scholar] [CrossRef]
- Casadevall, A. Cryptococci at the brain gate: Break and enter or use a Trojan horse? J. Clin. Investig. 2010, 120, 1389–1392. [Google Scholar] [CrossRef] [PubMed]
- May, R.C.; Stone, N.R.; Wiesner, D.L.; Bicanic, T.; Nielsen, K. Cryptococcus: From environmental saprophyte to global pathogen. Nat. Rev. Microbiol. 2016, 14, 106–117. [Google Scholar] [CrossRef]
- Strickland, A.B.; Shi, M. Mechanisms of fungal dissemination. Cell. Mol. Life Sci. 2021, 78, 3219–3238. [Google Scholar] [CrossRef]
- Panackal, A.A.; Williamson, K.C.; van de Beek, D.; Boulware, D.R.; Williamson, P.R. Fighting the Monster: Applying the Host Damage Framework to Human Central Nervous System Infections. mBio 2016, 7, e01906–e01915. [Google Scholar] [CrossRef] [Green Version]
- Panackal, A.A.; Wuest, S.C.; Lin, Y.C.; Wu, T.; Zhang, N.; Kosa, P.; Komori, M.; Blake, A.; Browne, S.K.; Rosen, L.B.; et al. Paradoxical Immune Responses in Non-HIV Cryptococcal Meningitis. PLoS Pathog. 2015, 11, e1004884. [Google Scholar] [CrossRef]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Shi, Z.W.; Strickland, A.B.; Shi, M. Cryptococcus neoformans Infection in the Central Nervous System: The Battle between Host and Pathogen. J. Fungi 2022, 8, 1069. [Google Scholar] [CrossRef]
- Leopold Wager, C.M.; Hole, C.R.; Wozniak, K.L.; Wormley, F.L., Jr. Cryptococcus and Phagocytes: Complex Interactions that Influence Disease Outcome. Front. Microbiol. 2016, 7, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olszewski, M.A.; Zhang, Y.; Huffnagle, G.B. Mechanisms of cryptococcal virulence and persistence. Future Microbiol. 2010, 5, 1269–1288. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.; Lodge, J.; Xue, C. Harnessing the Immune Response to Fungal Pathogens for Vaccine Development. Annu. Rev. Microbiol. 2022, 76, 703–726. [Google Scholar] [CrossRef] [PubMed]
- Pinner, R.W.; Hajjeh, R.A.; Powderly, W.G. Prospects for preventing cryptococcosis in persons infected with human immunodeficiency virus. Clin. Infect. Dis. 1995, 21 (Suppl. S1), S103–S107. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, J.N.; Meintjes, G.; Bicanic, T.; Buffa, V.; Hogan, L.; Mo, S.; Tomlinson, G.; Kropf, P.; Noursadeghi, M.; Harrison, T.S. Cerebrospinal fluid cytokine profiles predict risk of early mortality and immune reconstitution inflammatory syndrome in HIV-associated cryptococcal meningitis. PLoS Pathog. 2015, 11, e1004754. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, A.A.; Brouwer, A.E.; Wuthiekanun, V.; Jaffar, S.; Shattock, R.; Irving, D.; Sheldon, J.; Chierakul, W.; Peacock, S.; Day, N.; et al. IFN-gamma at the site of infection determines rate of clearance of infection in cryptococcal meningitis. J. Immunol. 2005, 174, 1746–1750. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, J.N.; Meintjes, G.; Rebe, K.; Williams, G.N.; Bicanic, T.; Williams, A.; Schutz, C.; Bekker, L.G.; Wood, R.; Harrison, T.S. Adjunctive interferon-gamma immunotherapy for the treatment of HIV-associated cryptococcal meningitis: A randomized controlled trial. AIDS 2012, 26, 1105–1113. [Google Scholar] [CrossRef] [Green Version]
- Pappas, P.G.; Bustamante, B.; Ticona, E.; Hamill, R.J.; Johnson, P.C.; Reboli, A.; Aberg, J.; Hasbun, R.; Hsu, H.H. Recombinant interferon-gamma 1b as adjunctive therapy for AIDS-related acute cryptococcal meningitis. J. Infect. Dis. 2004, 189, 2185–2191. [Google Scholar] [CrossRef] [Green Version]
- Casadevall, A.; Coelho, C.; Cordero, R.J.B.; Dragotakes, Q.; Jung, E.; Vij, R.; Wear, M.P. The capsule of Cryptococcus neoformans. Virulence 2019, 10, 822–831. [Google Scholar] [CrossRef] [Green Version]
- Jang, E.H.; Kim, J.S.; Yu, S.R.; Bahn, Y.S. Unraveling Capsule Biosynthesis and Signaling Networks in Cryptococcus neoformans. Microbiol. Spectr. 2022, e0286622. [Google Scholar] [CrossRef]
- Decote-Ricardo, D.; LaRocque-de-Freitas, I.F.; Rocha, J.D.B.; Nascimento, D.O.; Nunes, M.P.; Morrot, A.; Freire-de-Lima, L.; Previato, J.O.; Mendonca-Previato, L.; Freire-de-Lima, C.G. Immunomodulatory Role of Capsular Polysaccharides Constituents of Cryptococcus neoformans. Front. Med. 2019, 6, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, J.D.; Nascimento, M.T.; Decote-Ricardo, D.; Corte-Real, S.; Morrot, A.; Heise, N.; Nunes, M.P.; Previato, J.O.; Mendonca-Previato, L.; DosReis, G.A.; et al. Capsular polysaccharides from Cryptococcus neoformans modulate production of neutrophil extracellular traps (NETs) by human neutrophils. Sci. Rep. 2015, 5, 8008. [Google Scholar] [CrossRef] [Green Version]
- Bicanic, T.; Meintjes, G.; Rebe, K.; Williams, A.; Loyse, A.; Wood, R.; Hayes, M.; Jaffar, S.; Harrison, T. Immune reconstitution inflammatory syndrome in HIV-associated cryptococcal meningitis: A prospective study. J. Acquir. Immune Defic. Syndr. 2009, 51, 130–134. [Google Scholar] [CrossRef]
- Boulware, D.R.; Meya, D.B.; Bergemann, T.L.; Wiesner, D.L.; Rhein, J.; Musubire, A.; Lee, S.J.; Kambugu, A.; Janoff, E.N.; Bohjanen, P.R. Clinical features and serum biomarkers in HIV immune reconstitution inflammatory syndrome after cryptococcal meningitis: A prospective cohort study. PLoS Med. 2010, 7, e1000384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kambugu, A.; Meya, D.B.; Rhein, J.; O’Brien, M.; Janoff, E.N.; Ronald, A.R.; Kamya, M.R.; Mayanja-Kizza, H.; Sande, M.A.; Bohjanen, P.R.; et al. Outcomes of cryptococcal meningitis in Uganda before and after the availability of highly active antiretroviral therapy. Clin. Infect. Dis. 2008, 46, 1694–1701. [Google Scholar] [CrossRef]
- Lortholary, O.; Fontanet, A.; Memain, N.; Martin, A.; Sitbon, K.; Dromer, F. Incidence and risk factors of immune reconstitution inflammatory syndrome complicating HIV-associated cryptococcosis in France. AIDS 2005, 19, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Shelburne, S.A., 3rd; Darcourt, J.; White, A.C., Jr.; Greenberg, S.B.; Hamill, R.J.; Atmar, R.L.; Visnegarwala, F. The role of immune reconstitution inflammatory syndrome in AIDS-related Cryptococcus neoformans disease in the era of highly active antiretroviral therapy. Clin. Infect. Dis. 2005, 40, 1049–1052. [Google Scholar] [CrossRef] [Green Version]
- Sungkanuparph, S.; Jongwutiwes, U.; Kiertiburanakul, S. Timing of cryptococcal immune reconstitution inflammatory syndrome after antiretroviral therapy in patients with AIDS and cryptococcal meningitis. J. Acquir. Immune Defic. Syndr. 2007, 45, 595–596. [Google Scholar] [CrossRef]
- Lawn, S.D.; Bekker, L.G.; Myer, L.; Orrell, C.; Wood, R. Cryptococcocal immune reconstitution disease: A major cause of early mortality in a South African antiretroviral programme. AIDS 2005, 19, 2050–2052. [Google Scholar] [CrossRef]
- Wiesner, D.L.; Boulware, D.R. Cryptococcus-Related Immune Reconstitution Inflammatory Syndrome (IRIS): Pathogenesis and Its Clinical Implications. Curr. Fungal Infect. Rep. 2011, 5, 252–261. [Google Scholar] [CrossRef]
- Brienze, V.M.S.; Andre, J.C.; Liso, E.; Louis, I.V. Cryptococcal Immune Reconstitution Inflammatory Syndrome: From Blood and Cerebrospinal Fluid Biomarkers to Treatment Approaches. Life 2021, 11, 95. [Google Scholar] [CrossRef] [PubMed]
- Meya, D.B.; Manabe, Y.C.; Boulware, D.R.; Janoff, E.N. The immunopathogenesis of cryptococcal immune reconstitution inflammatory syndrome: Understanding a conundrum. Curr. Opin. Infect. Dis. 2016, 29, 10–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panackal, A.A.; Komori, M.; Kosa, P.; Khan, O.; Hammoud, D.A.; Rosen, L.B.; Browne, S.K.; Lin, Y.C.; Romm, E.; Ramaprasad, C.; et al. Spinal Arachnoiditis as a Complication of Cryptococcal Meningoencephalitis in Non-HIV Previously Healthy Adults. Clin. Infect. Dis. 2017, 64, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eschke, M.; Piehler, D.; Schulze, B.; Richter, T.; Grahnert, A.; Protschka, M.; Muller, U.; Kohler, G.; Hofling, C.; Rossner, S.; et al. A novel experimental model of Cryptococcus neoformans-related immune reconstitution inflammatory syndrome (IRIS) provides insights into pathogenesis. Eur. J. Immunol. 2015, 45, 3339–3350. [Google Scholar] [CrossRef]
- Khaw, Y.M.; Aggarwal, N.; Barclay, W.E.; Kang, E.; Inoue, M.; Shinohara, M.L. Th1-Dependent Cryptococcus-Associated Immune Reconstitution Inflammatory Syndrome Model with Brain Damage. Front. Immunol. 2020, 11, 529219. [Google Scholar] [CrossRef]
- Neal, L.M.; Xing, E.; Xu, J.; Kolbe, J.L.; Osterholzer, J.J.; Segal, B.M.; Williamson, P.R.; Olszewski, M.A. CD4+ T Cells Orchestrate Lethal Immune Pathology despite Fungal Clearance during Cryptococcus neoformans Meningoencephalitis. mBio 2017, 8, e01415-17. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Zhu, W.; Yin, J.; Huang, X.; Sun, L.; Hua, W.; Wang, W.; Zhang, T.; Dai, L.; Wu, H. A preliminary study on the characteristics of Th1/Th2 immune response in cerebrospinal fluid of AIDS patients with cryptococcal meningitis. BMC Infect. Dis. 2021, 21, 500. [Google Scholar] [CrossRef]
- Lortholary, O.; Dromer, F.; Mathoulin-Pelissier, S.; Fitting, C.; Improvisi, L.; Cavaillon, J.M.; Dupont, B. Immune mediators in cerebrospinal fluid during cryptococcosis are influenced by meningeal involvement and human immunodeficiency virus serostatus. J. Infect. Dis. 2001, 183, 294–302. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Dickson, D.W.; Casadevall, A. Pathology of cryptococcal meningoencephalitis: Analysis of 27 patients with pathogenetic implications. Hum. Pathol. 1996, 27, 839–847. [Google Scholar] [CrossRef]
- Mora, D.J.; Fortunato, L.R.; Andrade-Silva, L.E.; Ferreira-Paim, K.; Rocha, I.H.; Vasconcelos, R.R.; Silva-Teixeira, D.N.; Nascentes, G.A.; Silva-Vergara, M.L. Cytokine profiles at admission can be related to outcome in AIDS patients with cryptococcal meningitis. PLoS ONE 2015, 10, e0120297. [Google Scholar] [CrossRef]
- Jarvis, J.N.; Casazza, J.P.; Stone, H.H.; Meintjes, G.; Lawn, S.D.; Levitz, S.M.; Harrison, T.S.; Koup, R.A. The phenotype of the Cryptococcus-specific CD4+ memory T-cell response is associated with disease severity and outcome in HIV-associated cryptococcal meningitis. J. Infect. Dis. 2013, 207, 1817–1828. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.H.; McDonald, R.A.; Wells, J.C.; Huffnagle, G.B.; Lukacs, N.W.; Toews, G.B. The gamma interferon receptor is required for the protective pulmonary inflammatory response to Cryptococcus neoformans. Infect. Immun. 2005, 73, 1788–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardison, S.E.; Herrera, G.; Young, M.L.; Hole, C.R.; Wozniak, K.L.; Wormley, F.L., Jr. Protective immunity against pulmonary cryptococcosis is associated with STAT1-mediated classical macrophage activation. J. Immunol. 2012, 189, 4060–4068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardison, S.E.; Ravi, S.; Wozniak, K.L.; Young, M.L.; Olszewski, M.A.; Wormley, F.L., Jr. Pulmonary infection with an interferon-gamma-producing Cryptococcus neoformans strain results in classical macrophage activation and protection. Am. J. Pathol. 2010, 176, 774–785. [Google Scholar] [CrossRef]
- Herring, A.C.; Lee, J.; McDonald, R.A.; Toews, G.B.; Huffnagle, G.B. Induction of interleukin-12 and gamma interferon requires tumor necrosis factor alpha for protective T1-cell-mediated immunity to pulmonary Cryptococcus neoformans infection. Infect. Immun. 2002, 70, 2959–2964. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, K.; Qureshi, M.H.; Zhang, T.; Koguchi, Y.; Shibuya, K.; Naoe, S.; Saito, A. Interferon-gamma (IFN-gamma)-dependent protection and synthesis of chemoattractants for mononuclear leucocytes caused by IL-12 in the lungs of mice infected with Cryptococcus neoformans. Clin. Exp. Immunol. 1999, 117, 113–122. [Google Scholar] [CrossRef]
- Murdock, B.J.; Huffnagle, G.B.; Olszewski, M.A.; Osterholzer, J.J. Interleukin-17A enhances host defense against cryptococcal lung infection through effects mediated by leukocyte recruitment, activation, and gamma interferon production. Infect. Immun. 2014, 82, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Sionov, E.; Mayer-Barber, K.D.; Chang, Y.C.; Kauffman, K.D.; Eckhaus, M.A.; Salazar, A.M.; Barber, D.L.; Kwon-Chung, K.J. Type I IFN Induction via Poly-ICLC Protects Mice against Cryptococcosis. PLoS Pathog. 2015, 11, e1005040. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, F.; Tompkins, K.C.; McNamara, A.; Jain, A.V.; Moore, B.B.; Toews, G.B.; Huffnagle, G.B.; Olszewski, M.A. Robust Th1 and Th17 immunity supports pulmonary clearance but cannot prevent systemic dissemination of highly virulent Cryptococcus neoformans H99. Am. J. Pathol. 2009, 175, 2489–2500. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Gault, R.A.; Kozel, T.R.; Murphy, W.J. Immunomodulation with CD40 stimulation and interleukin-2 protects mice from disseminated cryptococcosis. Infect. Immun. 2006, 74, 2161–2168. [Google Scholar] [CrossRef]
- Zhou, Q.; Gault, R.A.; Kozel, T.R.; Murphy, W.J. Protection from direct cerebral Cryptococcus infection by interferon-gamma-dependent activation of microglial cells. J. Immunol. 2007, 178, 5753–5761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, Y.; Arora, S.; Erb-Downward, J.R.; McDonald, R.A.; Toews, G.B.; Huffnagle, G.B. Distinct roles for IL-4 and IL-10 in regulating T2 immunity during allergic bronchopulmonary mycosis. J. Immunol. 2005, 174, 1027–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huffnagle, G.B.; Boyd, M.B.; Street, N.E.; Lipscomb, M.F. IL-5 is required for eosinophil recruitment, crystal deposition, and mononuclear cell recruitment during a pulmonary Cryptococcus neoformans infection in genetically susceptible mice (C57BL/6). J. Immunol. 1998, 160, 2393–2400. [Google Scholar] [PubMed]
- Muller, U.; Stenzel, W.; Kohler, G.; Werner, C.; Polte, T.; Hansen, G.; Schutze, N.; Straubinger, R.K.; Blessing, M.; McKenzie, A.N.; et al. IL-13 induces disease-promoting type 2 cytokines, alternatively activated macrophages and allergic inflammation during pulmonary infection of mice with Cryptococcus neoformans. J. Immunol. 2007, 179, 5367–5377. [Google Scholar] [CrossRef] [Green Version]
- Stenzel, W.; Muller, U.; Kohler, G.; Heppner, F.L.; Blessing, M.; McKenzie, A.N.; Brombacher, F.; Alber, G. IL-4/IL-13-dependent alternative activation of macrophages but not microglial cells is associated with uncontrolled cerebral cryptococcosis. Am. J. Pathol. 2009, 174, 486–496. [Google Scholar] [CrossRef] [Green Version]
- Arora, S.; Olszewski, M.A.; Tsang, T.M.; McDonald, R.A.; Toews, G.B.; Huffnagle, G.B. Effect of cytokine interplay on macrophage polarization during chronic pulmonary infection with Cryptococcus neoformans. Infect. Immun. 2011, 79, 1915–1926. [Google Scholar] [CrossRef] [Green Version]
- Scriven, J.E.; Graham, L.M.; Schutz, C.; Scriba, T.J.; Wilkinson, K.A.; Wilkinson, R.J.; Boulware, D.R.; Urban, B.C.; Meintjes, G.; Lalloo, D.G. The CSF Immune Response in HIV-1-Associated Cryptococcal Meningitis: Macrophage Activation, Correlates of Disease Severity, and Effect of Antiretroviral Therapy. J. Acquir. Immune Defic. Syndr. 2017, 75, 299–307. [Google Scholar] [CrossRef]
- Beardsley, J.; Wolbers, M.; Kibengo, F.M.; Ggayi, A.B.; Kamali, A.; Cuc, N.T.; Binh, T.Q.; Chau, N.V.; Farrar, J.; Merson, L.; et al. Adjunctive Dexamethasone in HIV-Associated Cryptococcal Meningitis. N. Engl. J. Med. 2016, 374, 542–554. [Google Scholar] [CrossRef]
- Pirofski, L.A.; Casadevall, A. Immune-Mediated Damage Completes the Parabola: Cryptococcus neoformans Pathogenesis Can Reflect the Outcome of a Weak or Strong Immune Response. mBio 2017, 8, e02063-17. [Google Scholar] [CrossRef] [Green Version]
- Haddow, L.J.; Colebunders, R.; Meintjes, G.; Lawn, S.D.; Elliott, J.H.; Manabe, Y.C.; Bohjanen, P.R.; Sungkanuparph, S.; Easterbrook, P.J.; French, M.A.; et al. Cryptococcal immune reconstitution inflammatory syndrome in HIV-1-infected individuals: Proposed clinical case definitions. Lancet Infect. Dis. 2010, 10, 791–802. [Google Scholar] [CrossRef]
- Sun, H.Y.; Singh, N. Opportunistic infection-associated immune reconstitution syndrome in transplant recipients. Clin. Infect. Dis. 2011, 53, 168–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Lortholary, O.; Alexander, B.D.; Gupta, K.L.; John, G.T.; Pursell, K.; Munoz, P.; Klintmalm, G.B.; Stosor, V.; del Busto, R.; et al. An immune reconstitution syndrome-like illness associated with Cryptococcus neoformans infection in organ transplant recipients. Clin. Infect. Dis. 2005, 40, 1756–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulware, D.R.; Bonham, S.C.; Meya, D.B.; Wiesner, D.L.; Park, G.S.; Kambugu, A.; Janoff, E.N.; Bohjanen, P.R. Paucity of initial cerebrospinal fluid inflammation in cryptococcal meningitis is associated with subsequent immune reconstitution inflammatory syndrome. J. Infect. Dis. 2010, 202, 962–970. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Dorasamy, A.A.; Gosnell, B.I.; Elliott, J.H.; Spelman, T.; Omarjee, S.; Naranbhai, V.; Coovadia, Y.; Ndung’u, T.; Moosa, M.Y.; et al. Clinical and mycological predictors of cryptococcosis-associated immune reconstitution inflammatory syndrome. AIDS 2013, 27, 2089–2099. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Lim, A.; Omarjee, S.; Levitz, S.M.; Gosnell, B.I.; Spelman, T.; Elliott, J.H.; Carr, W.H.; Moosa, M.Y.; Ndung’u, T.; et al. Cryptococcosis-IRIS is associated with lower Cryptococcus-specific IFN-gamma responses before antiretroviral therapy but not higher T-cell responses during therapy. J. Infect. Dis. 2013, 208, 898–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meya, D.B.; Okurut, S.; Zziwa, G.; Cose, S.; Boulware, D.R.; Janoff, E.N. HIV-Associated Cryptococcal Immune Reconstitution Inflammatory Syndrome Is Associated with Aberrant T Cell Function and Increased Cytokine Responses. J. Fungi 2019, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Omarjee, S.; Lim, A.; Spelman, T.; Gosnell, B.I.; Carr, W.H.; Elliott, J.H.; Moosa, M.Y.; Ndung’u, T.; French, M.A.; et al. Chemokine levels and chemokine receptor expression in the blood and the cerebrospinal fluid of HIV-infected patients with cryptococcal meningitis and cryptococcosis-associated immune reconstitution inflammatory syndrome. J. Infect. Dis. 2013, 208, 1604–1612. [Google Scholar] [CrossRef] [Green Version]
- Meya, D.B.; Okurut, S.; Zziwa, G.; Cose, S.; Bohjanen, P.R.; Mayanja-Kizza, H.; Joloba, M.; Boulware, D.R.; Yukari Manabe, C.; Wahl, S.; et al. Monocyte Phenotype and IFN-gamma-Inducible Cytokine Responses Are Associated with Cryptococcal Immune Reconstitution Inflammatory Syndrome. J. Fungi 2017, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Liechty, C.A.; Solberg, P.; Were, W.; Ekwaru, J.P.; Ransom, R.L.; Weidle, P.J.; Downing, R.; Coutinho, A.; Mermin, J. Asymptomatic serum cryptococcal antigenemia and early mortality during antiretroviral therapy in rural Uganda. Trop. Med. Int. Health 2007, 12, 929–935. [Google Scholar] [CrossRef]
- Sungkanuparph, S.; Filler, S.G.; Chetchotisakd, P.; Pappas, P.G.; Nolen, T.L.; Manosuthi, W.; Anekthananon, T.; Morris, M.I.; Supparatpinyo, K.; Kopetskie, H.; et al. Cryptococcal immune reconstitution inflammatory syndrome after antiretroviral therapy in AIDS patients with cryptococcal meningitis: A prospective multicenter study. Clin. Infect. Dis. 2009, 49, 931–934. [Google Scholar] [CrossRef]
- Vlasova-St. Louis, I.; Chang, C.C.; Shahid, S.; French, M.A.; Bohjanen, P.R. Transcriptomic Predictors of Paradoxical Cryptococcosis-Associated Immune Reconstitution Inflammatory Syndrome. Open Forum Infect. Dis. 2018, 5, ofy157. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.A.; Nakouzi, A.; Chang, C.C.; Kuniholm, M.H.; Carreno, L.J.; Wang, T.; Ndung’u, T.; Lewin, S.R.; French, M.A.; Pirofski, L.A. Association between Plasma Antibody Responses and Risk for Cryptococcus-Associated Immune Reconstitution Inflammatory Syndrome. J. Infect. Dis. 2019, 219, 420–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meya, D.B.; Okurut, S.; Zziwa, G.; Rolfes, M.A.; Kelsey, M.; Cose, S.; Joloba, M.; Naluyima, P.; Palmer, B.E.; Kambugu, A.; et al. Cellular immune activation in cerebrospinal fluid from ugandans with cryptococcal meningitis and immune reconstitution inflammatory syndrome. J. Infect. Dis. 2015, 211, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Scriven, J.E.; Rhein, J.; Hullsiek, K.H.; von Hohenberg, M.; Linder, G.; Rolfes, M.A.; Williams, D.A.; Taseera, K.; Meya, D.B.; Meintjes, G.; et al. Early ART After Cryptococcal Meningitis Is Associated with Cerebrospinal Fluid Pleocytosis and Macrophage Activation in a Multisite Randomized Trial. J. Infect. Dis. 2015, 212, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Akilimali, N.A.; Muema, D.M.; Specht, C.; Chang, C.C.; Moosa, M.S.; Levitz, S.M.; Lewin, S.R.; French, M.A.; Ndung’u, T. Cryptococcosis-Associated Immune Reconstitution Inflammatory Syndrome Is Associated with Dysregulation of IL-7/IL-7 Receptor Signaling Pathway in T Cells and Monocyte Activation. J. Acquir. Immune Defic. Syndr. 2019, 80, 596–604. [Google Scholar] [CrossRef]
- Gray, F.; Bazille, C.; Adle-Biassette, H.; Mikol, J.; Moulignier, A.; Scaravilli, F. Central nervous system immune reconstitution disease in acquired immunodeficiency syndrome patients receiving highly active antiretroviral treatment. J. Neurovirol. 2005, 11 (Suppl. S3), 16–22. [Google Scholar] [CrossRef]
- Stone, S.F.; Price, P.; Brochier, J.; French, M.A. Plasma bioavailable interleukin-6 is elevated in human immunodeficiency virus-infected patients who experience herpesvirus-associated immune restoration disease after start of highly active antiretroviral therapy. J. Infect. Dis. 2001, 184, 1073–1077. [Google Scholar] [CrossRef]
- Stone, S.F.; Price, P.; Keane, N.M.; Murray, R.J.; French, M.A. Levels of IL-6 and soluble IL-6 receptor are increased in HIV patients with a history of immune restoration disease after HAART. HIV Med. 2002, 3, 21–27. [Google Scholar] [CrossRef]
- Worsley, C.M.; Suchard, M.S.; Stevens, W.S.; Van Rie, A.; Murdoch, D.M. Multi-analyte profiling of ten cytokines in South African HIV-infected patients with Immune Reconstitution Inflammatory Syndrome (IRIS). AIDS Res. Ther. 2010, 7, 36. [Google Scholar] [CrossRef] [Green Version]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef]
- Lim, A.; Tan, D.; Price, P.; Kamarulzaman, A.; Tan, H.Y.; James, I.; French, M.A. Proportions of circulating T cells with a regulatory cell phenotype increase with HIV-associated immune activation and remain high on antiretroviral therapy. AIDS 2007, 21, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.B.; Yong, Y.K.; Tan, H.Y.; Kamarulzaman, A.; Tan, L.H.; Lim, A.; James, I.; French, M.; Price, P. Immunological profiles of immune restoration disease presenting as mycobacterial lymphadenitis and cryptococcal meningitis. HIV Med. 2008, 9, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Tadokera, R.; Wilkinson, K.A.; Meintjes, G.A.; Skolimowska, K.H.; Matthews, K.; Seldon, R.; Rangaka, M.X.; Maartens, G.; Wilkinson, R.J. Role of the interleukin 10 family of cytokines in patients with immune reconstitution inflammatory syndrome associated with HIV infection and tuberculosis. J. Infect. Dis. 2013, 207, 1148–1156. [Google Scholar] [CrossRef] [Green Version]
- Park, B.J.; Wannemuehler, K.A.; Marston, B.J.; Govender, N.; Pappas, P.G.; Chiller, T.M. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. AIDS 2009, 23, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Pyrgos, V.; Seitz, A.E.; Steiner, C.A.; Prevots, D.R.; Williamson, P.R. Epidemiology of cryptococcal meningitis in the US: 1997–2009. PLoS ONE 2013, 8, e56269. [Google Scholar] [CrossRef] [Green Version]
- Bratton, E.W.; El Husseini, N.; Chastain, C.A.; Lee, M.S.; Poole, C.; Sturmer, T.; Juliano, J.J.; Weber, D.J.; Perfect, J.R. Comparison and temporal trends of three groups with cryptococcosis: HIV-infected, solid organ transplant, and HIV-negative/non-transplant. PLoS ONE 2012, 7, e43582. [Google Scholar] [CrossRef]
- Brizendine, K.D.; Baddley, J.W.; Pappas, P.G. Predictors of mortality and differences in clinical features among patients with Cryptococcosis according to immune status. PLoS ONE 2013, 8, e60431. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, H.; Takayama, A.; Fujiki, Y.; Ito, T.; Kitaoka, H. Refractory Cryptococcus neoformans Meningoencephalitis in an Immunocompetent Patient: Paradoxical Antifungal Therapy-Induced Clinical Deterioration Related to an Immune Response to Cryptococcal Organisms. Case Rep. Neurol. 2015, 7, 204–208. [Google Scholar] [CrossRef]
- Williamson, P.R. Post-infectious inflammatory response syndrome (PIIRS): Dissociation of T-cell-macrophage signaling in previously healthy individuals with cryptococcal fungal meningoencephalitis. Macrophage 2015, 2, e1078. [Google Scholar] [CrossRef] [Green Version]
- Gaiottino, J.; Norgren, N.; Dobson, R.; Topping, J.; Nissim, A.; Malaspina, A.; Bestwick, J.P.; Monsch, A.U.; Regeniter, A.; Lindberg, R.L.; et al. Increased neurofilament light chain blood levels in neurodegenerative neurological diseases. PLoS ONE 2013, 8, e75091. [Google Scholar] [CrossRef]
- Kuhle, J.; Plattner, K.; Bestwick, J.P.; Lindberg, R.L.; Ramagopalan, S.V.; Norgren, N.; Nissim, A.; Malaspina, A.; Leppert, D.; Giovannoni, G.; et al. A comparative study of CSF neurofilament light and heavy chain protein in MS. Mult. Scler. 2013, 19, 1597–1603. [Google Scholar] [CrossRef]
- Yang, B.; Zador, Z.; Verkman, A.S. Glial cell aquaporin-4 overexpression in transgenic mice accelerates cytotoxic brain swelling. J. Biol. Chem. 2008, 283, 15280–15286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yukutake, Y.; Yasui, M. Regulation of water permeability through aquaporin-4. Neuroscience 2010, 168, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Neal, L.M.; Ganguly, A.; Kolbe, J.L.; Hargarten, J.C.; Elsegeiny, W.; Hollingsworth, C.; He, X.; Ivey, M.; Lopez, R.; et al. Chemokine receptor CXCR3 is required for lethal brain pathology but not pathogen clearance during cryptococcal meningoencephalitis. Sci. Adv. 2020, 6, eaba2502. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Ganguly, A.; Zhao, J.; Ivey, M.; Lopez, R.; Osterholzer, J.J.; Cho, C.S.; Olszewski, M.A. CCR2 Signaling Promotes Brain Infiltration of Inflammatory Monocytes and Contributes to Neuropathology during Cryptococcal Meningoencephalitis. mBio 2021, 12, e0107621. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, C.; Sun, D.; Strickland, A.B.; Liu, G.; Shi, M. Quantitative analysis reveals internalisation of Cryptococcus neoformans by brain endothelial cells in vivo. Cell. Microbiol. 2021, 23, e13330. [Google Scholar] [CrossRef]
- Shi, M.; Colarusso, P.; Mody, C.H. Real-time in vivo imaging of fungal migration to the central nervous system. Cell. Microbiol. 2012, 14, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Mody, C.H. Fungal Infection in the Brain: What We Learned from Intravital Imaging. Front. Immunol. 2016, 7, 292. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Pawar, S.; Dutta, O.; Wang, K.; Rivera, A.; Xue, C. Macrophage Mediated Immunomodulation During Cryptococcus Pulmonary Infection. Front. Cell. Infect. Microbiol. 2022, 12, 859049. [Google Scholar] [CrossRef]


{kind=link}
| Sample Sources | Biological Activity | Biomarkers | References |
|---|---|---|---|
| CSF | Uncontrolled fungal growth | ↑fungal burdens | [26,64] |
| Poor inflammatory responses | ↓protein levels | [63,64] | |
| ↓leukocyte counts | [15,63] | ||
| ↓IFN-γ, TNF-α, IL-6, IL-8 | [63] | ||
| High CD8+ T cell response | ↑CSF-blood ratio of CXCR3+CCR5+CD8+ T cells | [67] | |
| High monocyte/macrophage response | ↑CCL2, CCL3, and GM-CSF | [15] | |
| ↑CCL2/CXCL10 and CCL3/CXCL10 ratios | [67] | ||
| ↑CSF-blood ratio of CCL2, CCL3, and CXCL10 | [67] | ||
| Blood | High fungemia | ↑cryptococcal antigen titer | [24,69,70] |
| Low Th1 responses | ↓CD4+ T cell counts | [15,24,26] | |
| ↓secretion of IFN-γ by peripheral CD4+ T cells; ↓activation of peripheral CD4+ T cells | [65,66] | ||
| ↓levels of TNF-α, G-CSF, GM-CSF, and VEGF; ↑CRP | [24] | ||
| High Th2/Th17 responses | ↑levels of IL-4 and IL-17 | [24] | |
| High monocyte response | ↑frequency of peripheral monocytes secreting TNF-α and IL-6 | [68] | |
| Low anti-virus response | ↓type I interferon gene expression | [71] | |
| Poor B cell function | ↓IgM secretion in response to cryptococcal antigens | [72] |
| Sample Sources | Biological Activity | Biomarkers | References |
|---|---|---|---|
| CSF | High T cell responses | ↑CD4+ T cells | [23,73] |
| High inflammatory cytokines | ↑IFN-γ, TNF-α, G-CSF, VEGF, eotaxin | [63] | |
| High monocyte/macrophage activation | ↑monocytes | [73] | |
| ↑sCD14 and sCD163 | [74] | ||
| High NK cell activation | ↑PD-L1 expression on NK cells | [73] | |
| Blood | High monocyte activation | ↑CD14+CD86+ or CD14+HLA-DR+ cells | [75] |
| High CD4+ T cell activation | ↑IL-2+/IL-17+ CD4+ T cells | [66] | |
| High inflammatory cytokines | ↑IL-6 and CRP | [24] | |
| ↑IFN-γ and IL-6 | [79] | ||
| Compensatory anti-inflammation | ↑IL-10 | [79] | |
| ↑Foxp3+CD4+ T cells | [81,82] |
| Sample Sources | Biological Activity | Biomarkers | References |
|---|---|---|---|
| CSF | Axonal damage | ↑NFL | [8,33] |
| High T cell and B responses | ↑CD4+ and CD8+ T cells, B cells | [8,33] | |
| High inflammatory cytokines and chmokines | ↑IFN-γ, IL-6, IL-18, CXCL10, MCP-1, MIP-1α, MCP-3, and MCP-3β | [8] | |
| Low Th2 cytokines | ↓IL-4 | [8] | |
| High innate immune cell responses | ↑NK cells, monocytes, and MyDCs | [8,33] | |
| Compensatory anti-inflammation | ↑IL-10 and IL-13 | [8] | |
| Blood | High T cell activation | ↑CD4+ and CD8+ T cells secreting IFN-γ | [8] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Z.W.; Chen, Y.; Ogoke, K.M.; Strickland, A.B.; Shi, M. Cryptococcal Immune Reconstitution Inflammatory Syndrome: From Clinical Studies to Animal Experiments. Microorganisms 2022, 10, 2419. https://doi.org/10.3390/microorganisms10122419
Shi ZW, Chen Y, Ogoke KM, Strickland AB, Shi M. Cryptococcal Immune Reconstitution Inflammatory Syndrome: From Clinical Studies to Animal Experiments. Microorganisms. 2022; 10(12):2419. https://doi.org/10.3390/microorganisms10122419
Chicago/Turabian StyleShi, Zoe W., Yanli Chen, Krystal M. Ogoke, Ashley B. Strickland, and Meiqing Shi. 2022. "Cryptococcal Immune Reconstitution Inflammatory Syndrome: From Clinical Studies to Animal Experiments" Microorganisms 10, no. 12: 2419. https://doi.org/10.3390/microorganisms10122419