
Diversity and Identification of Plant-Parasitic Nematodes in Wheat-Growing Ecosystems

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Occurrence of Plant-Parasitic Nematodes
3.2. Morphological Measurements
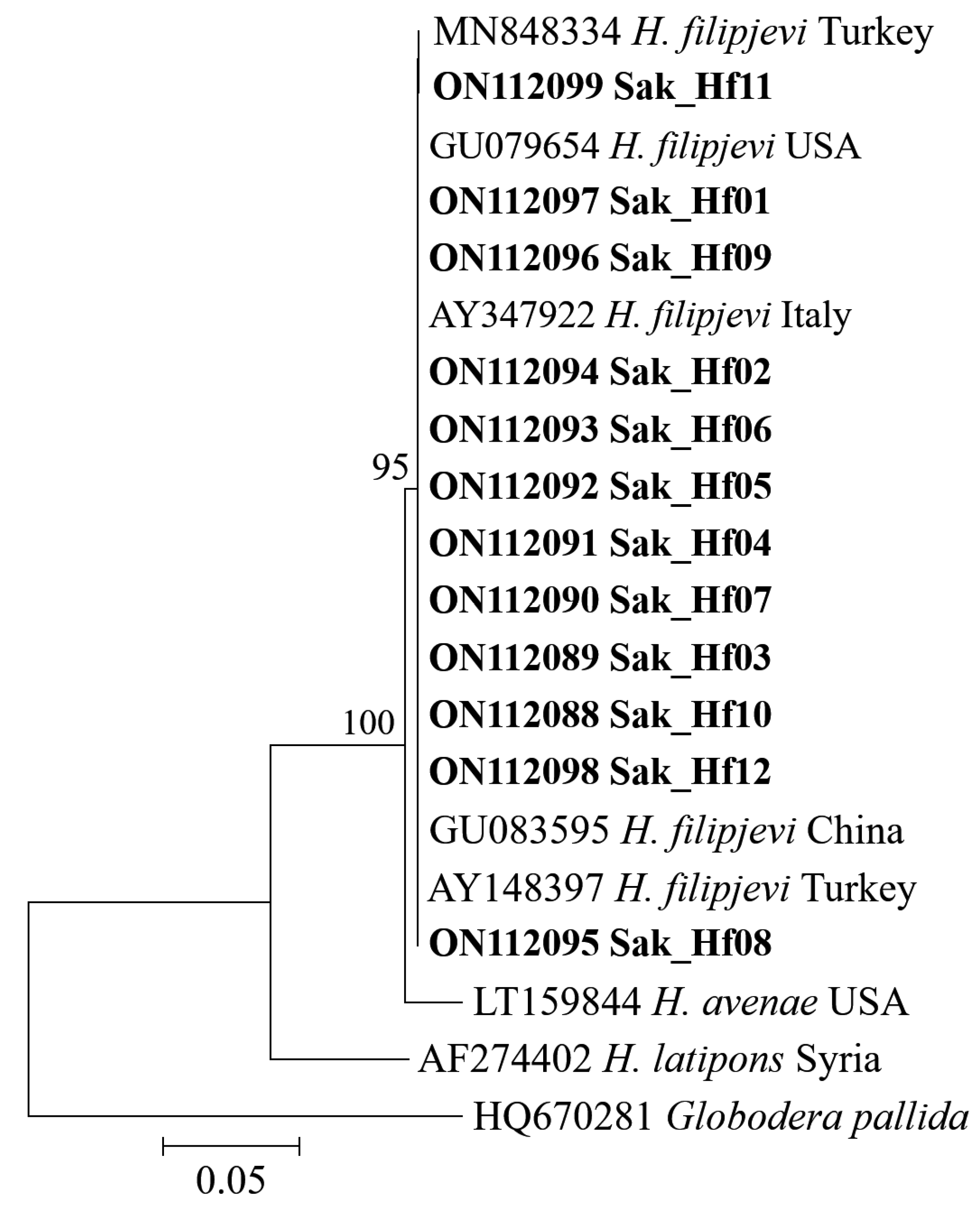
3.3. Molecular Features
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food Balance. 2010–2019. Available online: https://www.fao.org/faostat/en/#data/FBS (accessed on 12 January 2022).
- Turkish Statistical Institute (TUIK). Available online: https://biruni.tuik.gov.tr/medas/?kn=92&locale=tr (accessed on 12 January 2022).
- Zencirci, N.; Baloch, F.S.; Habyarimana, E.; Chung, G. Wheat Landraces; Springer: New York, NY, USA, 2021; pp. 124–215. [Google Scholar]
- Chitwood, D.J. Research on plant-parasitic nematode biology conducted by the United States Department of Agriculture-Agricultural Research Service. Pest Manag. Sci. 2003, 59, 748–753. [Google Scholar] [CrossRef]
- Koenning, S.R.; Overstreet, C.; Noling, J.W.; Donald, P.A.; Becker, J.O.; Fortnum, B.A. Survey of crop losses in response to phytoparasitic nematodes in the United States for 1994. J. Nematol. 1999, 31, 587–618. [Google Scholar]
- Smiley, R.W.; Nicol, J.M. Nematodes which challenge global wheat production. Wheat Sci. Trade 2009, 1, 171–187. [Google Scholar] [CrossRef]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef]
- Smiley, R.W.; Whittaker, R.G.; Gourlie, J.A.; Easley, S.A.; Ingham, R.E. Plant-parasitic nematodes associated with reduced wheat yield in Oregon: Heterodera avenae. J. Nematol. 2005, 37, 297–307. [Google Scholar]
- Walia, K.; Kanwar, R.S.; Bajaj, H.K. Biodiversity of nemic fauna associated with wheat in two districts of Haryana. Indian J. Nematol. 2005, 35, 224–225. [Google Scholar]
- Turner, S.J.; Subbotin, S.A. Cyst nematodes. In Plant Nematology; Perry, R.N., Moens, M., Eds.; CAB Internatiıonal: Wallingford, UK, 2013; pp. 109–143. [Google Scholar]
- Seid, A.; Imren, M.; Ali, M.A.; Toumi, F.; Paulitz, T.; Dababat, A.A. Genetic Resistance of Wheat towards Plant-Parasitic Nematodes: Current Status and Future Prospects. Biotech Stud. 2021, 30, 43–62. [Google Scholar] [CrossRef]
- Sobczak, M.; Golinowski, W. Cyst nematodes and syncytia. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Jones, J., Gheysen, G., Fenoll, C., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 61–82. [Google Scholar]
- Dababat, A.A.; Fourie, H. Nematode parasites of cereals. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; Sikora, R.A., Coyne, D., Hallmann, J., Timper, P., Eds.; CAB International: Wallingford, UK, 2018; pp. 163–221. [Google Scholar]
- Subbotin, S.; Sturhan, D.; Rumpenhorst, H.J.; Moens, M. Molecular and morphological characterisation of the Heterodera avenae species complex (Tylenchida: Heteroderidae). Nematology 2003, 5, 515–538. [Google Scholar] [CrossRef] [Green Version]
- Subbotin, S.A.; Toumi, F.; Elekçioğlu, I.H.; Waeyenberge, L.; Maafi, Z.T. DNA barcoding, phylogeny and phylogeography of the cyst nematode species of the Avenae group from the genus Heterodera (Tylenchida: Heteroderidae). Nematology 2018, 20, 671–702. [Google Scholar] [CrossRef] [Green Version]
- Ahin, E.; Nicol, J.; Elekcioglu, I.; Yorgancilar, O.; Yildirim, A.; Tulek, A.; Hekimhan, H.; Yorgancilar, A.; Kilinc, A.T.; Bolat, N.; et al. Frequency and diversity of cereal nematodes on the Central Anatolian Plateau of Turkey. In Cereal Cyst Nematodes: Status, Research and Outlook; Riley, I.T., Nicol, J.M., Dababat, A.A., Eds.; CIMMYT: Ankara, Turkey, 2009; pp. 100–105. [Google Scholar]
- Toktay, H.; Imren, M.; Öcal, A.; Waeyenberge, L.; Viaene, N.; Dababat, A.A. Incidence of cereal cyst nematodes in the East Anatolia Region in Turkey. Russ. J. Nematol. 2015, 23, 29–40. [Google Scholar]
- Imren, M.; Yildiz, S.; Toktay, H.; Duman, N.; Dababat, A.A. Morphometric and genetic variability among Mediterranean cereal cyst nematode (Heterodera latipons) populations in Turkey. Turk. J. Zool. 2018, 42, 625–636. [Google Scholar] [CrossRef]
- Imren, M.; Özer, G.; Paulitz, T.C.; Morgounov, A.; Dababat, A.A. Plant-parasitic nematodes associated with wheat in central, eastern, and southeastern Kazakhstan. Plant Dis. 2021, 105, 2299–2305. [Google Scholar] [CrossRef] [PubMed]
- Imren, M.; Ozer, G.; Duman, N.; Dababat, A. Phenotypic and genotypic characterization of wheat and barley varieties for resistance to cereal cyst nematode (Heterodera latipons). Genet. Resour. Crop Evol. 2021, 68, 2131–2141. [Google Scholar] [CrossRef]
- Hooper, D.J. Extraction of Free-Living Stages from Soil. In Laboratory Methods for Work with Plant and Soil Nematodes; Southey, J.F., Ed.; Her Majesty’s Stationery Office: London, UK, 1986; pp. 5–30. [Google Scholar]
- Fenwick, D.W. Methods for the recovery and counting of cysts of Heterodera schachtii from soil. J. Helminthol. 1940, 18, 155–172. [Google Scholar] [CrossRef]
- Carneiro, R.M.D.G.; Lima, F.S.D.O.; Correia, V.R. Methods and tools currently used for the identification of plant parasitic nematodes. Nematol Concepts Diagn. Control. 2017, 1, 19–52. [Google Scholar] [CrossRef] [Green Version]
- Golden, A.M. Morphology and identification of cyst nematodes. In Cyst nematodes; Springer: Boston, MA, USA, 1986; pp. 23–45. [Google Scholar] [CrossRef]
- Dababat, A.A.; Erginbaş-Orakci, G.; Toktay, H.; Imren, M.; Akin, B.; Braun, H.-J.; Dreisigacker, S.; Elekcioğlu, I.H.; Morgounov, A. Resistance of winter wheat to Heterodera filipjevi in Turkey. Turk. J. Agric. For. 2014, 38, 180–186. [Google Scholar] [CrossRef]
- S’Jacob, J.J.; Van Bezooijen, J. Manual for Practical Work in Nematology; Landbouwhogeschool: Wageningen, The Netherlands, 1984; pp. 40–70. [Google Scholar]
- Southey, J.F. Principles of Sampling for Nematodes. In Laboratory Methods for Work with Plant and Soil Nematodes; Southey, J.F., Ed.; Her Majesty’s Stationery Office: London, UK, 1986; 402p. [Google Scholar]
- Joyce, S.A.; Reid, A.; Driver, F.; Curran, J. Application of polymerase chain reaction (PCR) methods to the identification of entomopathogenic nematodes. In Genetics of Entomopathogenic Nematode-Bacterium Complexes, Proceedings of Symposium and Workshop; European Commission, DG XII: Luxembourg, 1994; pp. 178–187. [Google Scholar]
- Subbotin, S.A. Molecular identification of nematodes using polymerase chain reaction (PCR). In Techniques for Work with Plant and Soil Nematodes; CAB international: Wallingford, UK, 2020; p. 218. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Briefings Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Schwartz, T.; Pickett, B.E.; He, S.; Klem, E.B.; Scheuermann, R.H.; Passarotti, M.; Kaufman, S.; O’Leary, M.A. A RESTful API for Access to Phylogenetic Tools via the CIPRES Science Gateway. Evol. Bioinform. 2015, 11, EBO-S21501. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E. A mathematical theory of communication. Bell. Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Lumeran, B.T. Assemblage of Gastropods in the Rocky Intertidal Zone of Asry Beach, Kingdom of Bahrain. In Inverte-Brates-Ecophysiology and Management; Ray, S., Diarte-Plata, G., Escamilla-Montes, R., Eds.; Intech Open: Rijeka, Croatia, 2019; pp. 140–165. [Google Scholar]
- Siddiqi, M.R. Tylenchida Parasites of Plants and Insects; CABI Publishing: Wallingford, UK, 2000; pp. 450–848. [Google Scholar]
- A Handoo, Z. A Key and Compendium to Species of the Heterodera avenae Group (Nematoda: Heteroderidae). J. Nematol. 2002, 34, 250–262. [Google Scholar] [PubMed]
- Imren, M.; Elekcioğlu, I.H. Diyarbakır ili Buğday, Sebze ve bağ Alanlarinda Önemli Bitki Paraziti Nematod Türlerinin Belirlenmesi. Master’s Thesis, Fen Bilimleri Enstitüsü, Mersin, Turkey, 2008. [Google Scholar]
- Öcal, A.; Elekcioğlu, İ.H. Plant Parasitic Nematode Species Associated With Barley (Hordeum Vulgare) and Wheat (Triticum Spp. L.) In Adiyaman Province. In Nematodes of Small Grain Cereals; FAO: Ankara, Turkey, 2015; p. 149. [Google Scholar]
- İmren, M.; Toktay, H.; Kutuk, H.; Dababat, A.A. Occurrence and identification of cereal cyst nematode, Heterodera filipjevi (Nemata: Heteroderidae), in Bolu province of Turkey. Nematropica 2016, 46, 154–161. [Google Scholar]
- Toktay, H.; Imren, M.; Akyol, B.G.; Evlice, E.; Riley, I.T.; Dababat, A. Phytophagous nematodes in cereal fields in Niğde Province, Turkey. Turk. J. Entomol. 2020, 44, 559–569. [Google Scholar] [CrossRef]
- Subbotin, S.A.; Mundo-Ocampo, M.; Baldwin, J.G. Systematics of Cyst Nematodes (Nematoda: Heteroderinae), Part B; Brill: Leiden, The Netherlands, 2010. [Google Scholar] [CrossRef]
- Wouts, W.M.; Sturhan, D. Heterodera aucklandica sp. n.(Nematoda: Heteroderidae) from a New Zealand native grass, with notes on the species of the H. avenae group. N. Z. J. Zool. 1995, 22, 199–207. [Google Scholar] [CrossRef]
- Rivoal, R.; Valette, S.; Bekal, S.; Gauthier, J.P.; Yahyaoui, A. Genetic and phenotypic diversity in the graminaceous cyst nem-atode complex, inferred from PCR-RFLP of ribosomal DNA and morphometric analysis. Eur. J. Plant Pathol. 2003, 109, 227–241. [Google Scholar] [CrossRef]
- Valdeolivas, A.; Romero, M.D. Morphometric relationships of some members of the Heterodera avenae complex (Nematoda: Heteroderidae). Nematologica 1990, 36, 292–303. [Google Scholar] [CrossRef]
- Baldwin, J.G. Evolution of cyst and noncyst-forming Heteroderinae. Annu. Rev. Phytopathol. 1992, 30, 271–290. [Google Scholar] [CrossRef]
- Bossis, M.; Mugniéry, D. Specific status of six Globodera parasites of solanaceous plants studied by means of two-dimensional gel electrophoresis with a comparison of gel patterns by a computed system. Fundam. Appl. Nematol. 1993, 16, 47–56. [Google Scholar]
- Ferris, V.R.; Ferris, J.M.; Faghihi, J. Variation in spacer ribosomal DNA in some cyst-forming species of plant parasitic nem-atodes. Fundam. Appl. Nematol. 1993, 16, 177–184. [Google Scholar]
- Hyman, B.C.; Powers, T.O. Integration of molecular data with systematics of plant parasitic nematodes. Annu. Rev. Phytopathol. 1991, 29, 89–107. [Google Scholar] [CrossRef] [PubMed]
- Subbotin, S.A.; Vera, I.C.D.P.; Mundo-Ocampo, M.; Baldwin, J.G. Identification, phylogeny and phylogeography of cir-cumfenestrate cyst nematodes (Nematoda: Heteroderidae) as inferred from analysis of ITS-rDNA. Nematology 2011, 13, 805–824. [Google Scholar] [CrossRef] [Green Version]
- Powers, T. Nematode Molecular Diagnostics: From Bands to Barcodes. Annu. Rev. Phytopathol. 2004, 42, 367–383. [Google Scholar] [CrossRef] [PubMed]
- Bekal, S.; Gauthier, J.P.; Rivoal, R. Genetic diversity among a complex of cereal cyst nematodes inferred from RFLP analysis of the ribosomal internal transcribed spacer region. Genome 1997, 40, 479–486. [Google Scholar] [CrossRef]




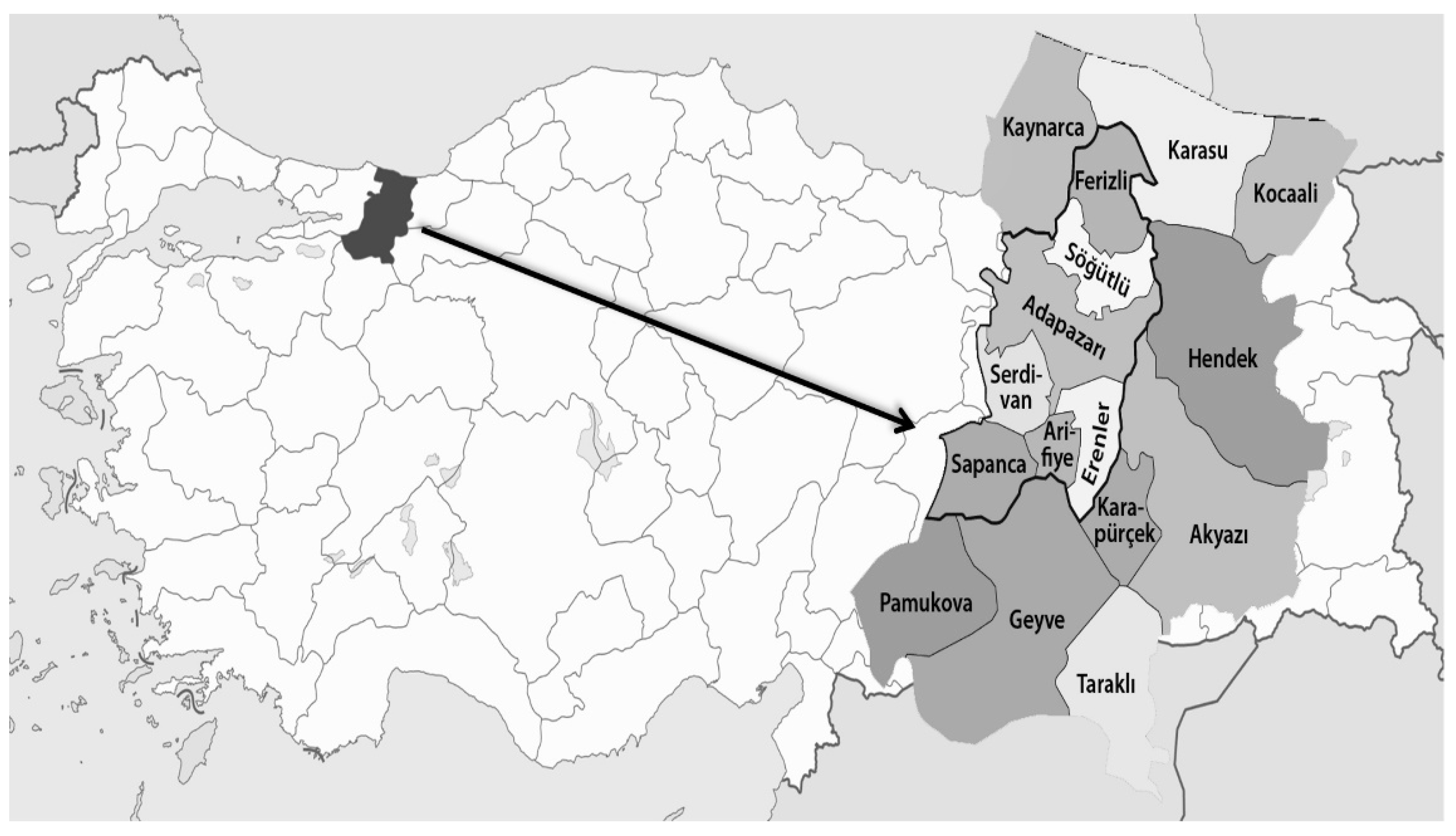{kind=link}
{kind=link}
{kind=link}
| No. | District | Location | Latitude | Longitude |
|---|---|---|---|---|
| 1 | Hendek | Beylice | 40°83′13″ | 30°84′80″ |
| 2 | Beylice | 40°83′04″ | 30°84′24″ | |
| 3 | Bıçkıatik | 40°83′85″ | 30°85′69″ | |
| 4 | Beylice | 40°83′85″ | 30°88′34″ | |
| 5 | Beylice | 40°83′28″ | 30°86′14″ | |
| 6 | Ferizli | Karadiken | 40°96′13″ | 30°47′70″ |
| 7 | Sebiller | 40°96′13″ | 30°43′25″ | |
| 8 | Doğancı | 40°98′44″ | 30°43′06″ | |
| 9 | Karadiken | 40°95′59″ | 30°51′48″ | |
| 10 | Karadiken | 40°96′15″ | 30°43′27″ | |
| 11 | Kaynarca | Kızılcaali | 40°98′66″ | 30°40′71″ |
| 12 | Kızılcaali | 40°98′27″ | 30°40′58″ | |
| 13 | Merkez | 41°03′33″ | 30°30′77″ | |
| 14 | Küçükkaynarca | 41°03′38″ | 30°32′48″ | |
| 15 | Geyve | Merkez | 40°51′81″ | 30°30′91″ |
| 16 | Merkez | 40°51′78″ | 30°30′94″ | |
| 17 | Merkez | 40°51′74″ | 30°31′06″ | |
| 18 | Merkez | 40°50′41″ | 30°30′55″ | |
| 19 | Merkez | 40°51′18″ | 30°29′81″ | |
| 20 | Ilıca | 40°45′25″ | 30°38′10″ | |
| 21 | Çamlık | 40°45′24″ | 30°37′93″ | |
| 22 | Taraklı | Hacıyakup | 40°44′69″ | 30°37′94″ |
| 23 | Hacıyakup | 40°43′95″ | 30°43′85″ | |
| 24 | Hacıyakup | 40°43′98″ | 30°43′90″ | |
| 25 | Aksu | 40°44′64″ | 30°45′37″ | |
| 26 | Aksu | 40°44′62″ | 30°46′51″ | |
| 27 | Aksu | 40°44′52″ | 30°46′59″ | |
| 28 | Aksu | 40°44′37″ | 30°47′00″ | |
| 29 | Söğütlü | Merkez | 40°44′28″ | 30°43′80″ |
| 30 | Merkez | 40°44′13″ | 30°43′91″ | |
| 31 | Merkez | 40°86′42″ | 30°49′01″ | |
| 32 | Akyazı | Merkez | 40°89′69″ | 30°46′43″ |
| 33 | Merkez | 40°67′62″ | 30°62′37″ | |
| 34 | Şerefiye | 40°61′73″ | 30°67′32″ | |
| 35 | Şerefiye | 40°61′68″ | 30°67′43″ | |
| 36 | Merkez | 40°05′33″ | 30°85′00″ | |
| 37 | Pamukova | Merkez | 40°05′32″ | 30°85′06″ |
| 38 | Mekece | 40°45′44″ | 30°04′85″ | |
| 39 | Mekece | 40°45′51″ | 30°04′72″ | |
| 40 | Mekece | 40°45′48″ | 30°04′28″ |
| No. | Districts | Locations | Nematode Genera |
|---|---|---|---|
| 1 | Hendek | Beylice | Pratylenchus; Dorylaimida; Basiria |
| 2 | Beylice | Ditylenchus | |
| 3 | Bıçkıatik | Dorylaimida | |
| 4 | Beylice | Dorylaimida | |
| 5 | Beylice | Dorylaimida | |
| 6 | Ferizli | Karadiken | Scutylenchus; Filenchus; Dorylaimida |
| 7 | Sebiller | Helicotylenchus; Dorylaimida | |
| 8 | Doğancı | Helicotylenchus; Scutylenchus; Filenchus | |
| 9 | Karadiken | Dorylaimida | |
| 10 | Karadiken | Filenchus | |
| 11 | Kaynarca | Kızılcaali | Dorylaimida |
| 12 | Kızılcaali | Filenchus; Aphelenchus | |
| 13 | Merkez | Helicotylenchus; Scutylenchus; Dorylaimida | |
| 14 | Küçük Kaynarca | Tylenchus; Dorylaimida | |
| 15 | Geyve | Town center | Merlinius; Filenchus; Ditylenchus |
| 16 | Town center | Pratylenchus; Scutylenchus | |
| 17 | Town center | Pratylenchus | |
| 18 | Town center | Scutylenchus; Filenchus; Dorylaimida | |
| 19 | Town center | Ditylenchus; Dorylaimida | |
| 20 | Ilıca | Pratylenchus; Paratrophurus; Filenchus; Ditylenchus; Dorylaimida | |
| 21 | Çamlık | Pratylenchus; Merlinius; Dorylaimida | |
| 22 | Taraklı | Hacıyakup | Merlinius; Filenchus; Dorylaimida |
| 23 | Hacıyakup | Merlinius; Pratylenchus; Dorylaimida | |
| 24 | Hacıyakup | Ditylenchus | |
| 25 | Aksu | Scutylenchus; Filenchus | |
| 26 | Aksu | Filenchus | |
| 27 | Aksu | Ditylenchus; Tylenchus; Boleodorus | |
| 28 | Aksu | Helicotylenchus; Filenchus; Ditylenchus | |
| 29 | Hacıyakup | Pratylenchus; Merlinius; Ditylenchus; Scutylenchus Filenchus | |
| 30 | Hacıyakup | Dorylaimida | |
| 31 | Söğütlü | Mahsudiye | Pratylenchus; Aphelenchoides; Filenchus; Boleodorus; Dorylaimida |
| 32 | Town center | Ditylenchus; Dorylaimida | |
| 33 | Akyazı | Town center | Helicotylenchus; Amplimerlinius; Merlinius; Ditylenchus; Filenchus; Dorylaimida |
| 34 | Şerefiye | Helicotylenchus; Ditylenchus; Monachus; Dorylaimida | |
| 35 | Şerefiye | Aphelenchoides; Ditylenchus; Dorylaimida | |
| 36 | Kocaali | Town center | Ditylenchus; Amplimerlinius; Scutylenchus; Monachus; Scutylenchus |
| 37 | Town center | Helicotylenchus; Ditylenchus; Scutylenchus; Filenchus Dorylaimida | |
| 38 | Pamukova | Mekece | Merlinius; Ditylenchus; Filenchus; Dorylaimida |
| 39 | Mekece | Helicotylenchus; Dorylaimida | |
| 40 | Mekece | Merlinius; Ditylenchus; Tylenchus |
| No. | Nematode Genera | Infested Fields (%) * | Nematode Abundance ** |
|---|---|---|---|
| 1 | Merlinus | 28 | 960 ± 120 (220–1100) |
| 2 | Trophurus | 16 | 240 ± 20 (200–360) |
| 3 | Paratrophurus | 14 | 140 ± 40 (80–980) |
| 4 | Pratylenchus | 26 | 240 ± 40 (400–980) |
| 5 | Amplimerlinus | 32 | 340 ± 60 (200–880) |
| 6 | Helicotylenchus | 40 | 420 ± 50 (280–780) |
| 7 | Tylenchus | 34 | 320 ± 40 (200–480) |
| 8 | Pratylenchoides | 30 | 640 ± 160 (200–980) |
| 9 | Scutylenchus | 28 | 550 ± 60 (100–860) |
| 10 | Filenchus | 24 | 280 ± 20 (100–200) |
| 11 | Boleodorus | 20 | 120 ± 160 (140–360) |
| 12 | Basiria | 16 | 240 ± 30 (160–900) |
| No. | District | Number of Fields Surveyed | Infestation (%) | Average Number of Nematodes/100 g * |
|---|---|---|---|---|
| 1 | Hendek | 5 | 3 | 40 |
| 2 | Pamukova | 4 | 4 | 20 |
| 3 | Geyve | 7 | 2 | 15 |
| 4 | Akyazı | 5 | 5 | 25 |
| Total | 22 | 35 * | 25 | |
| No. | District | Location | Latitude | Longitude |
|---|---|---|---|---|
| 1 | Kaynarca | Kızılcaali | 40°98′66″ | 30°40′71″ |
| 2 | Kızılcaali | 40°98′27″ | 30°40′58″ | |
| 3 | Merkez | 41°03′33″ | 30°30′77″ | |
| 4 | Küçükkaynarca | 41°03′38″ | 30°32′48″ | |
| 5 | Geyve | Merkez-I | 40°50′41″ | 30°30′55″ |
| 6 | Merkez-II | 40°51′74″ | 30°31′06″ | |
| 7 | Çamlık | 40°45′24″ | 30°37′93″ | |
| 8 | Ilıca | 40°45′25″ | 30°38′10″ | |
| 9 | Taraklı | Hacıyakup-I | 40°44′69″ | 30°37′94″ |
| 10 | Hacıyakup-II | 40°43′95″ | 30°43′85″ | |
| 11 | Aksu | 40°44′64″ | 30°45′37″ | |
| 12 | Söğütlü | Merkez | 40°89′69″ | 30°46′43″ |
| Morphological Characters | Geyve | Taraklı | Kaynarca | Söğütlü |
|---|---|---|---|---|
| Body length | 521.3 ± 4.84 b,* (482.8–542.5) | 548.3 ± 3.74 a (471.8–588.6) | 518.9 ± 6.62 b (467.8–548.5) | 556.3 ± 7.52 a (481.5–589.8) |
| Stylet length | 23.2 ± 0.45 b (22.2–25.6) | 24.9 ± 0.32 a (22.5–27.6) | 23.5 ± 0.49 b (21.6–24.8) | 24.2 ± 0.38 a,b (21.6–25.9) |
| Tail length | 50.45 ± 1.86 c (43.4–62.6) | 52.66 ± 3.35 b (41.6–64.6) | 54.68 ± 3.94 a (45.6–66.8) | 51.24 ± 4.52 b (42.5–67.6) |
| Hyaline tail tip length | 25.6 ± 1.32 b (21.4–34.5) | 26.6 ± 2.57 a (22.4–32.3) | 25.8 ± 1.45 b (21.9–37.6) | 25.4 ± 1.88 b (20.4–36.4) |
| Fenestral length | 51.32 ± 2.32 b (42.1–63.4) | 58.34 ± 3.44 a (43.8–68.4) | 54.54 ± 2.32 a,b (45.2–63.2) | 48.34 ± 1.64 b (40.2–59.8) |
| Semifenestral width | 20.4 ± 2.43 c (15.2–26.6) | 24.4 ± 2 a (16.7–24.6) | 23.2 ± 1.04 a,b (16.8–27.5) | 22.4 ± 2.09 b (15.2–28.6) |
| Vulval bridge width | 10.56 ± 1.32 b (6.8–17.2) | 14.44 ± 2.43 a (9.2–18.6) | 12.09 ± 2.33 a,b (7.88–16.54) | 15.65 ± 2.33 a (9.2–17.5) |
| Vulval slit length | 19.5 ± 1.02 b (12.6–25.8) | 22.6 ± 1.58 a (10.3–26.2) | 20.2 ± 2.34 a,b (13.4–25.6) | 18.2 ± 3.32 b (13.3–25.5) |
| Nematode Genus | Average Number of Nematodes/100 g Soil (Frequency) | pi | ln(pi) | pi × ln(pi) |
|---|---|---|---|---|
| Merlinus | 960 | 0.213808 | −1.54267 | −0.32984 |
| Trophurus | 240 | 0.053452 | −2.92897 | −0.15656 |
| Paratrophurus | 140 | 0.031180 | −3.46797 | −0.10813 |
| Pratylenchus | 240 | 0.053452 | −2.92897 | −0.15656 |
| Amplimerlinus | 340 | 0.075724 | −2.58066 | −0.19542 |
| Helicotylenchus | 420 | 0.093541 | −2.36935 | −0.22163 |
| Tylenchus | 320 | 0.071269 | −2.64129 | −0.18824 |
| Pratylenchoides | 640 | 0.142539 | −1.94814 | −0.27769 |
| Scutylenchus | 550 | 0.122494 | −2.09969 | −0.25720 |
| Filenchus | 280 | 0.062361 | −2.77482 | −0.17304 |
| Boleodorus | 120 | 0.026726 | −3.62212 | −0.09680 |
| Basiria | 240 | 0.053452 | −2.92897 | −0.15656 |
| Total | 4490 | 1 | −31.8336 | −2.31767 |
| H = 2.31767 | ||||
| EH = 0.932699 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keçici, A.İ.; Bozbuğa, R.; Öcal, A.; Yüksel, E.; Özer, G.; Yildiz, Ş.; Lahlali, R.; Slaats, B.; Dababat, A.A.; İmren, M. Diversity and Identification of Plant-Parasitic Nematodes in Wheat-Growing Ecosystems. Microorganisms 2022, 10, 1534. https://doi.org/10.3390/microorganisms10081534
Keçici Aİ, Bozbuğa R, Öcal A, Yüksel E, Özer G, Yildiz Ş, Lahlali R, Slaats B, Dababat AA, İmren M. Diversity and Identification of Plant-Parasitic Nematodes in Wheat-Growing Ecosystems. Microorganisms. 2022; 10(8):1534. https://doi.org/10.3390/microorganisms10081534
Chicago/Turabian StyleKeçici, Ayça İrem, Refik Bozbuğa, Atilla Öcal, Ebubekir Yüksel, Göksel Özer, Şenol Yildiz, Rachid Lahlali, Brigitte Slaats, Abdelfattah A. Dababat, and Mustafa İmren. 2022. "Diversity and Identification of Plant-Parasitic Nematodes in Wheat-Growing Ecosystems" Microorganisms 10, no. 8: 1534. https://doi.org/10.3390/microorganisms10081534