
National Surveillance of Acute Flaccid Paralysis Cases in Senegal during 2017 Uncovers the Circulation of Enterovirus Species A, B and C

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Data Collection
2.3. Sample Preparation
2.4. RNA Extraction and Amplification
2.5. Sequencing and Phylogenetic Analysis
2.6. Recombination Analysis
2.7. Statistical Analysis
3. Results
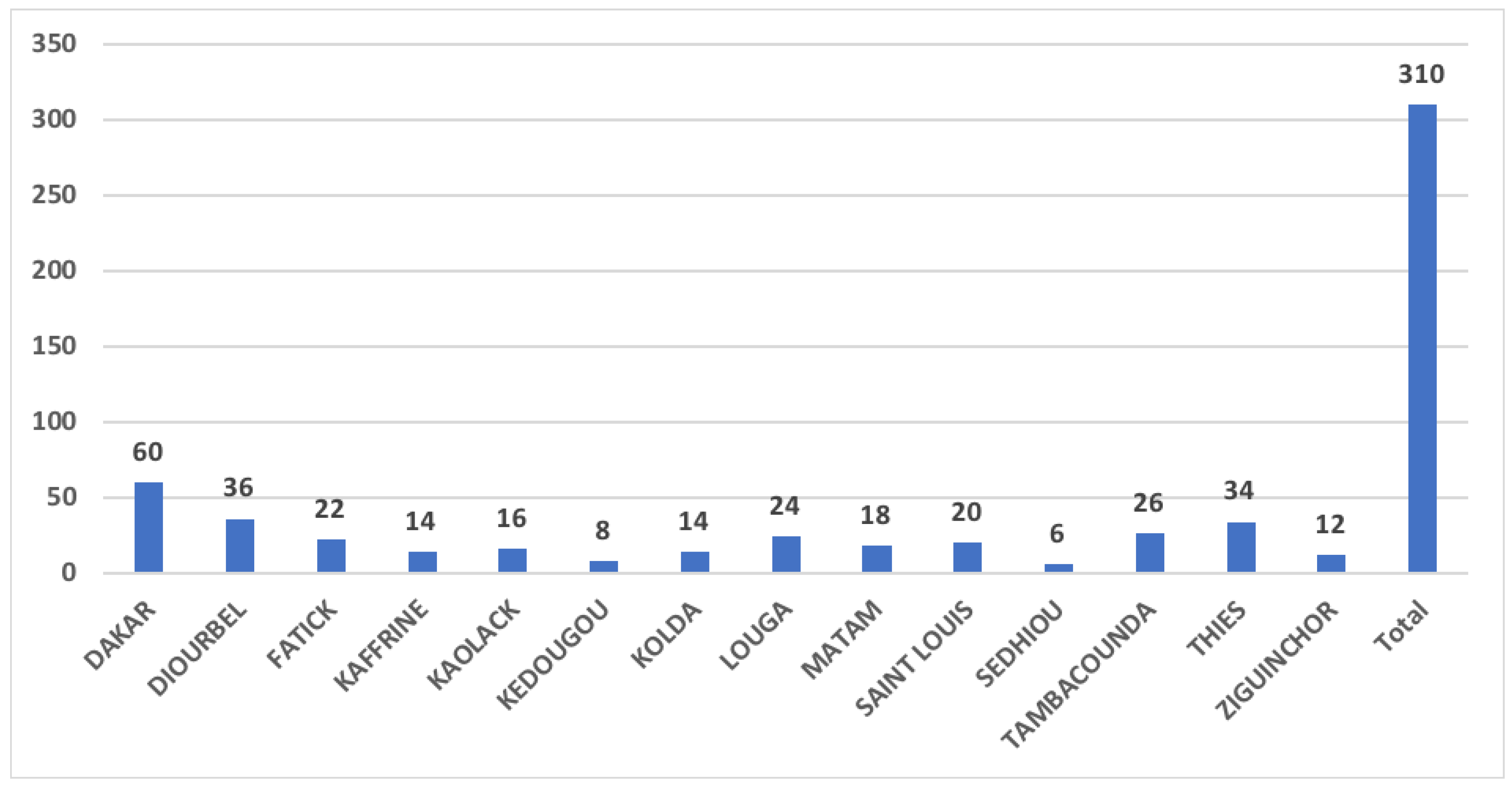
3.1. Isolation Data
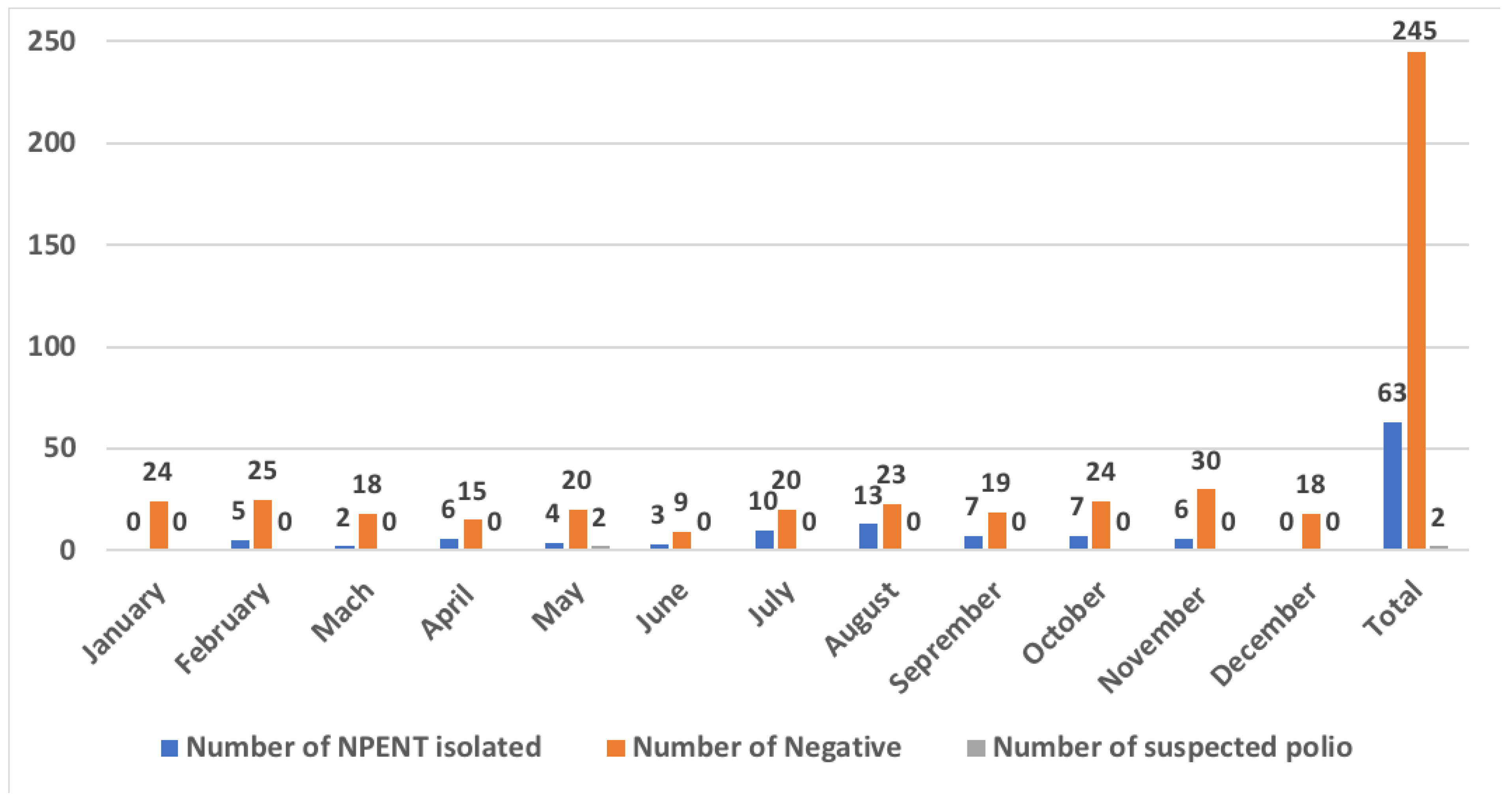
3.2. Real Time RT-PCR Analysis
3.3. Virus Diversity
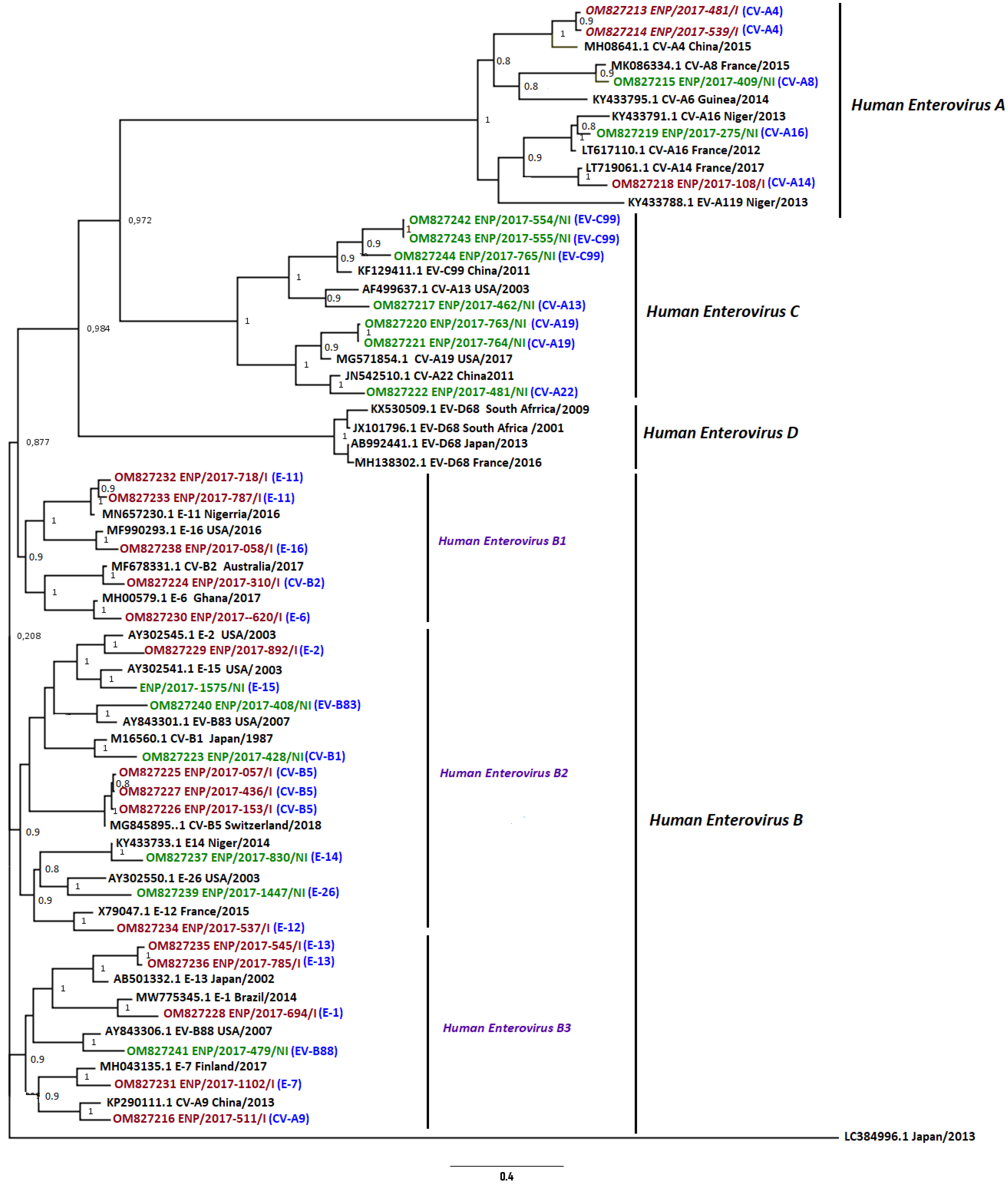
3.4. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Sequence_ID | Collection_Date | Isolation Source | Country | Host | Strain | GenBank Accession Number |
|---|---|---|---|---|---|---|
| ENP/2017-481/I | 2017 | ZIGUINCHOR | Senegal | Human | coxsackievirus A4 | OM827213 |
| ENP/2017-539/I | 2017 | DAKAR | Senegal | Human | coxsackievirus A4 | OM827214 |
| ENP/2017-409/NI | 2017 | SEDHIOU | Senegal | Human | coxsackievirus A8 | OM827215 |
| ENP/2017-511/I | 2017 | LOUGA | Senegal | Human | coxsackievirus A9 | OM827216 |
| ENP/2017-462/NI | 2017 | DAKAR | Senegal | Human | coxsackievirus A13 | OM827217 |
| ENP/2017-108/I | 2017 | THIES | Senegal | Human | coxsackievirus A14 | OM827218 |
| ENP/2017-275/NI | 2017 | MATAM | Senegal | Human | coxsackievirus A16 | OM827219 |
| ENP/2017-763/NI | 2017 | KAFFRINE | Senegal | Human | coxsackievirus A19 | OM827220 |
| ENP/2017-764/NI | 2017 | THIES | Senegal | Human | coxsackievirus A19 | OM827221 |
| ENP/2017-481/NI | 2017 | DAKAR | Senegal | Human | coxsackievirus A22 | OM827222 |
| ENP/2017-428/NI | 2017 | DAKAR | Senegal | Human | coxsackievirus B1 | OM827223 |
| ENP/2017-310/I | 2017 | SAINT LOUIS | Senegal | Human | coxsackievirus B2 | OM827224 |
| ENP/2017-057/I | 2017 | DAKAR | Senegal | Human | coxsackievirus B5 | OM827225 |
| ENP/2017-153/I | 2017 | DIOURBEL | Senegal | Human | coxsackievirus B5 | OM827226 |
| ENP/2017-436/I | 2017 | DIOURBEL | Senegal | Human | coxsackievirus B5 | OM827227 |
| ENP/2017-694/I | 2017 | DIOURBEL | Senegal | Human | echovirus E1 | OM827228 |
| ENP/2017-892/I | 2017 | KAOLACK | Senegal | Human | echovirus E2 | OM827229 |
| ENP/2017-620/I | 2017 | DIOURBEL | Senegal | Human | echovirus E6 | OM827230 |
| ENP/2017-1102/I | 2017 | SAINT LOUIS | Senegal | Human | echovirus E7 | OM827231 |
| ENP/2017-718/I | 2017 | FATICK | Senegal | Human | echovirus E11 | OM827232 |
| ENP/2017-787/I | 2017 | MATAM | Senegal | Human | echovirus E11 | OM827233 |
| ENP/2017-537/I | 2017 | DAKAR | Senegal | Human | echovirus E12 | OM827234 |
| ENP/2017-545/I | 2017 | LOUGA | Senegal | Human | echovirus E13 | OM827235 |
| ENP/2017-785/I | 2017 | MATAM | Senegal | Human | echovirus E13 | OM827236 |
| ENP/2017-830/NI | 2017 | TAMBACOUNDA | Senegal | Human | echovirus E14 | OM827237 |
| ENP/2017-058/I | 2017 | DAKAR | Senegal | Human | echovirus E16 | OM827238 |
| ENP/2017-1447/NI | 2017 | TAMBACOUNDA | Senegal | Human | echovirus E26 | OM827239 |
| ENP/2017-408/NI | 2017 | DAKAR | Senegal | Human | enterovirus B83 | OM827240 |
| ENP/2017-479/NI | 2017 | THIES | Senegal | Human | enterovirus B88 | OM827241 |
| ENP/2017-554/NI | 2017 | THIES | Senegal | Human | enterovirus C99 | OM827242 |
| ENP/2017-555/NI | 2017 | THIES | Senegal | Human | enterovirus C99 | OM827243 |
| ENP/2017-765/NI | 2017 | THIES | Senegal | Human | enterovirus C99 | OM827244 |
| ENP/2017-1575/NI | 2017 | DAKAR | Senegal | Human | echovirus E15 | OM867579 |
| ENP/2017-408/I | 2017 | DAKAR | Senegal | Human | echovirus E7 | ON502380 |
| ENP/2017-1125/I | 2017 | MATAM | Senegal | Human | echovirus E2 | ON502381 |
| ENP/2017-1409/I | 2017 | TAMBACOUNDA | Senegal | Human | echovirus E33 | ON502382 |
| ENP/2017-1446/I | 2017 | TAMBACOUNDA | Senegal | Human | echovirus E13 | ON502383 |
References
- Tapparel, C.S.; Petty, T.J.; Kaiser, L. Picornavirus and enterovirus diversity with associated human diseases. Infect. Genet. Evol. 2013, 14, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Oberste, M.S.M.; Kilpatrick, D.R.; Pallansch, M.A. Molecular evolution of the human enteroviruses: Correlation of serotype with VP1 sequence and application to picornavirus classification. J. Virol. 1999, 73, 1941–1948. [Google Scholar] [CrossRef] [Green Version]
- Tangermann, R.H.A.; Birmingham, M.; Horner, R.; Olivé, J.M.; Nkowane, B.M.; Hull, H.F. Current status of the global eradication of poliomyelitis. World Health Stat. Q. 1997, 50, 188–194. [Google Scholar] [PubMed]
- Sawyer, M.H. Enterovirus infections: Diagnosis and treatment. Curr. Opin. Pediatr. 2001, 13, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Abzug, M.J. diagnosis, and management of enterovirus infections in neonates. Paediatr. Drugs 2004, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Senegal—GPEI. Available online: polioeradication.org (accessed on 15 February 2022).
- World Health Organization (WHO). Available online: https://apps.who.int/iris/bitstream/handle/10665/345661/WER9639-481-484-eng-fre.pdf (accessed on 15 February 2022).
- Available online: https://www.nytimes.com/2020/08/24/world/africa/polio-africa-eradicated.html (accessed on 15 February 2022).
- Crom, S.C.M.O.; van Loon, A.M.; Argilagos-Alvarez, A.A.; Peeters, M.F.; van Furth, A.M.; Rossen, J.W.A. Detection of enterovirus RNA in cerebrospinal fluid: Comparison of two molecular assays. J. Virol. Methods. 2012, 179, 104–107. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Guidelines for Environmental Surveillance of Poliovirus Circulation; WHO: Geneva, Switzerland, 2003; pp. 9–10. Available online: https://polioeradication.org/wp-content/uploads/2016/07/WHO_V-B_03.03_eng.pdf (accessed on 22 October 2021).
- Adeniji, J.A.O.; George, U.E.; Ibok, U.I.; Faleye, T.O.C.; Adewumi, M.O. Preponderance of enterovirus C in RD-L20B-cell-culture-negative stool samples from children diagnosed with acute flaccid paralysis in Nigeria. Arch. Virol. 2017, 162, 3089–3101. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Surveillance Indicators. Available online: https://polioeradication.org/polio-today/polio-now/surveillance-indicators/ (accessed on 22 October 2021).
- World Health Organization (WHO). Polio Laboratory Manual teWG, Switzerland. 2004. Available online: https://apps.who.int/iris/handle/10665/68762 (accessed on 22 October 2021).
- Available online: https://apps.who.int/iris/bitstream/handle/10665/208873/WHO_POLIO_15.05_fre.pdf (accessed on 7 January 2022).
- Nix, W.A.O.; Pallansch, M.A. Sensitive, seminested PCR amplification of VP1 sequences for direct identification of all enterovirus serotypes from original clinical specimens. J. Clin. Microbiol. 2006, 44, 2698–2704. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.S.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Paramvir, S.D.; Arkin, A.P. FastTree 2: Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Garcia, M.D.; kebe, O.; Fall, A.D.; Ndiaye, K. Identification and molecular characterization of non-polio enteroviruses from children with acute flaccid paralysis in West Africa, 2013–2014. Sci. Rep. 2017, 7, 3808. [Google Scholar] [CrossRef] [PubMed]
- Fall, A.; Ndiaye, N.; Jallow, M.M.; Barry, M.A.; Touré, C.S.B.; Kebe, O.; Kiori, D.E.; Sy, S.; Dia, M.; Goudiaby, D.; et al. Enterovirus D68 Subclade B3 Circulation in Senegal, 2016: Detection from Infuenza-like Illness and Acute Flaccid Paralysis Surveillance. Sci. Rep. 2019, 9, 13881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzezutka, A.; Cook, N. Survival of human enteric viruses in the environment and food. FEMS Microbiol. Rev. 2004, 28, 441–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apostol, L.N.; Suzuki, A.; Bautista, A.; Galang, H.; Paladin, F.J.; Fuji, N.; Lupisan, S.; Olveda, S.; Oshitani, H. Detection of non-polio enteroviruses from 17 years of virological surveillance of acute flaccid paralysi in the Philippines. J. Med. Virol. 2012, 84, 624–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessaud, M.; Pillet, S.; Ibrahim, W.; Joffret, M.L.; Pozzetto, B.; Delpeyroux, F.; Gouandjika-Vasilache, L. Molecular characterization of human enteroviruses in the Central African Republic: Uncovering wide diversity and identification of a new human enterovirus A71 genogroup. J. Clin. Micro. 2012, 50, 1650–1658. [Google Scholar] [CrossRef] [Green Version]
- Cavé, C.A.; Bièche, D.; Brault, F.; de Fraipont, F.; Fina, S.; Loric, L.; Maisonneuve, F.; Namour, S.T. RT-PCR in clinical diagnosis. Ann. Biol. Clin. 2003, 61, 635–644. [Google Scholar]
- Shan Teo, F.M.; Chu, J.J.H. Diagnosis of human enteroviruses that cause hand, foot and mouthdisease. Expert Rev. Anti. Infect. Ther. 2016, 14, 443–445. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; wang, C.; Yang, M.; Yang, G.; Ma, X. Etiology of Multiple Non-EV71 and Non-CVA16 Enteroviruses Associated with Hand, Foot and Mouth Disease in Jinan, China, 2009-June 2013. PLoS ONE 2015, 10, e0142733. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Si, L.; Ciao, L.; Zhong, H.; Dong, N.; Xu, J. Enterovirus genotypes causing hand foot and mouth disease in Shanghai, China: A molecular epidemiological analysis. BMC Inf. Dis. 2013, 13, 489. [Google Scholar] [CrossRef] [Green Version]
- Faleye, T.O.C.; Adewumi, M.O.; Japhet, M.O.; David, O.M.; Oluyege, A.O.; Adeniji, J.A.; Famurewa, O. Non-polio enteroviruses in faeces of children diagnosed with acute flaccid paralysis in Nigeria. Virol. J. 2017, 14, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouwer, L.; van der Sanden, S.M.G.; Calis, J.C.J.; Bruning, A.H.L.; van Hensbroek, M.B.; Pajkrt, D.; Wolthers, K.C. High frequency of Polio-like Enterovirus C strains with differential clustering of CVA-13 and EV-C99 subgenotypes in a cohort of Malawian children. Arch Virol. 2018, 163, 2645–2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Z. Non-polio enteroviruses from acute flaccid paralysis surveillance in Shandong Province, China, 1988–2013. Sci. Rep. 2014, 4, 6167. [Google Scholar] [CrossRef] [Green Version]
- Odoom, J.K. Human Enteroviruses isolated during acute flaccid paralysis surveillance in Ghana: Implications for the posteradication era. Pan Afr. Med. J. 2012, 12, 74. [Google Scholar] [PubMed]
- Oyero, O.G.; Adu, F.D. Non-polio enteroviruses serotypes circulating in Nigeria. Afri. J. Med. Med. Sci. 2010, 39, 201–208. [Google Scholar]
- Mullins, J. A Emergence of echovirus type 13 as a prominent enterovirus. Clin. Infect. Dis. 2004, 38, 70–77. [Google Scholar] [CrossRef]
- Maan, H.S.; Chowdhary, R.; Shakya, A.K.; Dhole, T.N. Genetic variants of echovirus 13, northern India, 2010. Emerg. Infect. Dis. 2013, 19, 293–296. [Google Scholar] [CrossRef]
- Cheon, D.S.; Lee, J.; Lee, K.; Yoon, J.; Cho, H. Isolation and molecular identification of echovirus 13 isolated from patients of aseptic meningitis in Korea, 2002. J. Med. Virol. 2004, 73, 439–442. [Google Scholar] [CrossRef]
- Sadeuh-Mba, S.A.; Hugo Kavunga, M.; Joffret, M.L.; Yogolelo, R.; Endegue-Zanga, M.C.; Delpeyroux, F. Genetic landscape and macro-evolution of cocirculating Coxsackieviruses A and Vaccinederived Polioviruses in the Democratic Republic of Congo, 2008-2013. PLoS Negl Trop Dis. 2019, 19, 13. [Google Scholar]
- Sadeuh-Mba, S.A.; Bessaud, M.; Massenet, D.; Joffret, M.-L.; Endegue, M.-C.; Njouom, R.; Reynes, J.-M.; Rousset, D.; Delpeyroux, F. High frequency and diversity of species C enteroviruses in Cameroon and neighboring countries. J. Clin. Microbiol. 2013, 51, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Mombo, I.M. First Detection of an Enterovirus C99 in a Captive Chimpanzee with Acute Flaccid Paralysis, from the Tchimpounga Chimpanzee Rehabilitation Center, Republic of Congo. PLoS ONE 2005, 10, e0136700. [Google Scholar] [CrossRef]
- Schmidt, N.; Lennette, E.H. Propagation and isolation of group A coxsackieviruses in RD cells. J. Clin. Microbiol. 1975, 2, 183–185. [Google Scholar] [CrossRef]
- Lipson, S.; Costello, P.; Szabo, K. Sensitivity of rhabdomyosarcoma and guinea pig embryo cell cultures to field isolates of difficult-to-cultivate group A coxsackieviruses. J. Clin. Microbiol. 1988, 26, 1298–1303. [Google Scholar] [CrossRef] [Green Version]
- Combelas, N.; Holmblat, B.; Joffret, M.L.; Colbère-Garapin, F.; Delpeyroux, F. Recombination between poliovirus and coxsackie A viruses of species C: A model of viral genetic plasticity and emergence. Viruses 2011, 3, 1460–1484. [Google Scholar] [CrossRef] [Green Version]
- Lafolie, L. Optimisation du diagnostic des infections à entérovirus et étude de leur pouvoir pathogène. Ph.D. Thesis, Université Clermont Auvergne, Clermont-Ferrand, France, 2018. [Google Scholar]
- Van Leer-Buter, C.C.R.; Borger, E.; Hubert, G.; Niesters, M. Newly Identified Enterovirus C Genotypes, Identified in the Netherlands through Routine Sequencing of All Enteroviruses Detected in Clinical Materials from 2008 to 2015. J. Clin. Microbiol. 2016, 54, 2306–2314. [Google Scholar] [CrossRef] [Green Version]
- Tokarz, R.; Haq, S.; Luna, G.; Andrew, J.; Bennett, A.J.; Silva, M.; Leguia, M.; Kasper, M.; Bausch, D.G.; Lipkin, W.L. Genomic analysis of two novel human enterovirus C genotypes found in respiratory samples from Peru. J. Gen. Virol. 2013, 94, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.P.; Costa, E.V.; Oliveira, S.S.; Souza, M.C.; Edson, E.; Silva, D. RT-PCR based analysis of cell culture negative stool samples from poliomyelitis suspected cases. J. Clin. Virol. 2002, 23, 149–152. [Google Scholar] [CrossRef]
- Johnson, A.A.; Faleye, T.O.C. Impact of cell lines included in enterovirus isolation protocol on perception of non polio enterovirus species C diversity. J. Virol. Methods. 2014, 207, 238–247. [Google Scholar] [CrossRef]
- Oyero, O.G.; Adu, F.D.; Ayukekbong, J.A. Molecular characterization of diverse species enterovirus-B types from children with acute flaccid paralysis and asymptomatic children in Nigeria. Virus Res. 2014, 30, 189–193. [Google Scholar] [CrossRef]
- Abbasian, F.; Tabatabaie, H.; Sarijloo, M.; Shahmahmoodi, S.; Yousefi, A.; Saberbaghi, T.; Mokhtari, A.T.; Nategh, R. A comparative analysis of routine techniques: Reverse transcriptase polymerase chain reaction (RT-PCR) and five cell lines for detection of enteroviruses in stool samples. Iran J. Microbiol. 2011, 3, 75–79. [Google Scholar]
- Brown, D.M.; Zhang, Y.; Scheuermann, R.H. Epidemiology and Sequence-Based Evolutionary Analysis of Circulating Non-Polio Enteroviruses. Microorganisms 2020, 8, 1856. [Google Scholar] [CrossRef] [PubMed]



| Species | Type | Specimen Name | Total | Type | Specimen Name | Total |
|---|---|---|---|---|---|---|
| Isolated (I) | Unisolated (NI) | |||||
| Enterovirus A | coxsackievirus A4 | ENP/17-481 ENP/17-539 | 2 | coxsackievirus A8 | ENP/17-409 | 1 |
| coxsackievirus A14 | ENP/17-108 | 1 | coxsackievirus A16 | ENP/17-275 ENP/17-408 | 2 | |
| Enterovirus B | coxsackievirus A9 | ENP/17-511 | 1 | coxsackievirus B1 | ENP/17-428 | 1 |
| coxsackievirus B2 | ENP/17-310 | 1 | enterovirus B83 | ENP/17-408 | 1 | |
| coxsackievirus B5 | ENP/17-057 ENP/17-153 ENP/17-436 | 3 | enterovirus B88 | ENP/17-479 | 1 | |
| echovirus 1 | ENP/17-694 | 1 | echovirus 13 | ENP/17-785 | 1 | |
| echovirus 2 | ENP/17-892 ENP/17-1125 | 2 | echovirus 14 | ENP/17-830 | 1 | |
| echovirus 6 | ENP/17-620 | 1 | echovirus 15 | ENP/17-1575 | 1 | |
| echovirus 7 | ENP/17-1102 ENP/17-408 | 2 | echovirus 26 | ENP/17-1447 | 1 | |
| echovirus 11 | ENP/17-718 ENP/17-787 | 2 | ||||
| echovirus 12 | ENP/17-537 | 1 | ||||
| echovirus 13 | ENP/17-545 ENP/17-785 ENP/17-1446 | 3 | ||||
| echovirus 16 | ENP/17-058 | 1 | ||||
| echovirus 33 | ENP/17-1409 | 1 | ||||
| Enterovirus C | coxsackievirus A21 | ENP/17-832 | 1 | coxsackievirus A13 | ENP/17-462 | 1 |
| coxsackievirus A19 | ENP/17-763 ENP/17-764 | 2 | ||||
| coxsackievirus A22 | ENP/17-481 | 1 | ||||
| enterovirus C99 | ENP/17-554 ENP/17-555 ENP/17-765 | 3 | ||||
| Total | 23 | 17 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndiaye, N.; Fall, A.; Kébé, O.; Kiory, D.; Dia, H.; Fall, M.; Dia, N.; Sall, A.A.; Faye, M.; Faye, O. National Surveillance of Acute Flaccid Paralysis Cases in Senegal during 2017 Uncovers the Circulation of Enterovirus Species A, B and C. Microorganisms 2022, 10, 1296. https://doi.org/10.3390/microorganisms10071296
Ndiaye N, Fall A, Kébé O, Kiory D, Dia H, Fall M, Dia N, Sall AA, Faye M, Faye O. National Surveillance of Acute Flaccid Paralysis Cases in Senegal during 2017 Uncovers the Circulation of Enterovirus Species A, B and C. Microorganisms. 2022; 10(7):1296. https://doi.org/10.3390/microorganisms10071296
Chicago/Turabian StyleNdiaye, Ndack, Amary Fall, Ousmane Kébé, Davy Kiory, Hamet Dia, Malick Fall, Ndongo Dia, Amadou Alpha Sall, Martin Faye, and Ousmane Faye. 2022. "National Surveillance of Acute Flaccid Paralysis Cases in Senegal during 2017 Uncovers the Circulation of Enterovirus Species A, B and C" Microorganisms 10, no. 7: 1296. https://doi.org/10.3390/microorganisms10071296