
Molecular Detection of Anaplasma phagocytophilum and Ehrlichia Species in Ticks Removed from Humans in the Republic of Korea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Tick Collection and Identification
2.2. DNA Extraction
2.3. Polymerase Chain Reaction (PCR) Amplification
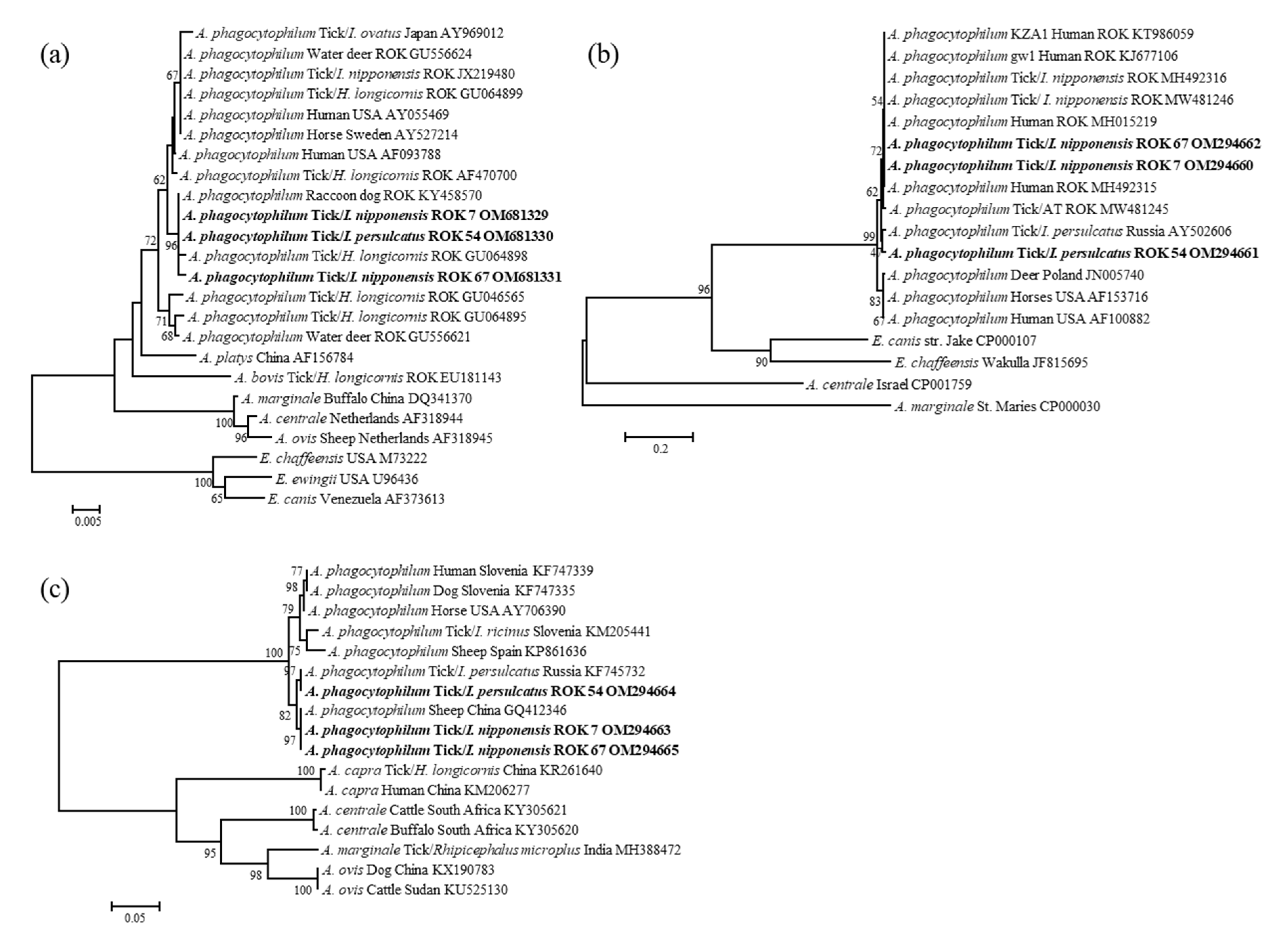
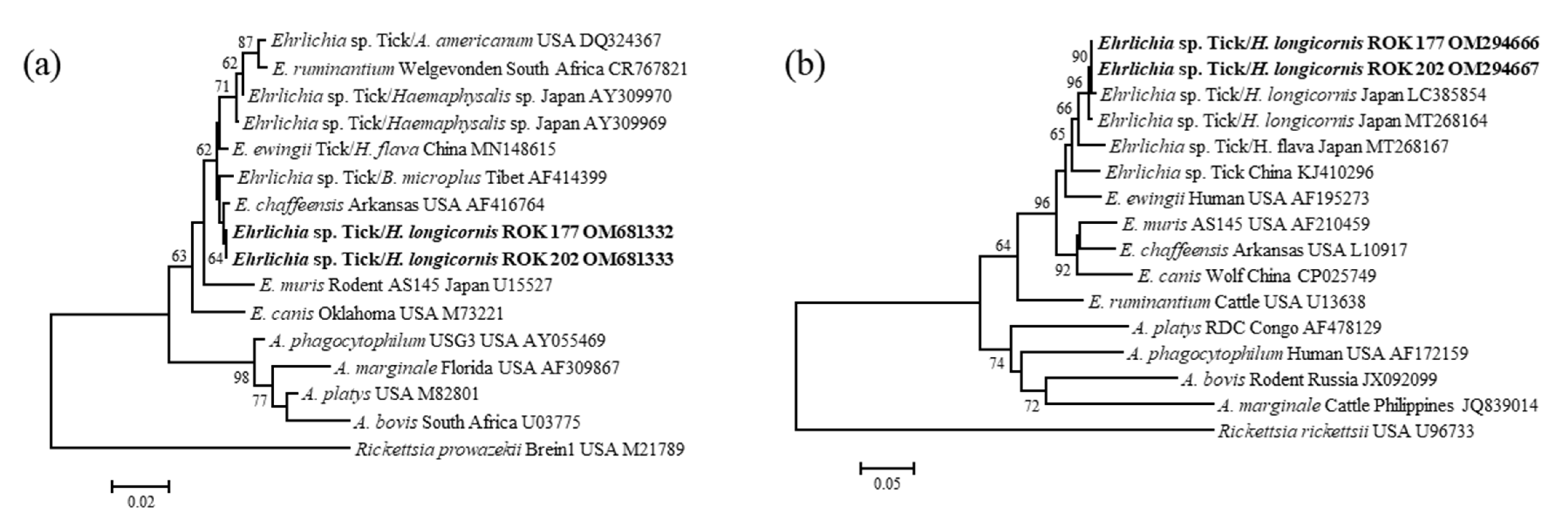
2.4. Nucleotide Sequencing and Phylogenetic Analysis
3. Results
3.1. Identification of Ticks
3.2. Detection of Anaplasma sp. and Ehrlichia sp.
3.3. Molecular and Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharifah, N.; Heo, C.C.; Ehlers, J.; Houssaini, J.; Tappe, D. Ticks and tick-borne pathogens in animals and humans in the island nations of Southeast Asia: A review. Acta Trop. 2020, 209, 105527. [Google Scholar] [CrossRef] [PubMed]
- Baneth, G. Tick-borne infections of animals and humans: A common ground. Int. J. Parasitol. 2014, 44, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Süss, J.; Klaus, C.; Gerstengarbe, F.W.; Werner, P.C. What makes ticks tick? Climate change, ticks, and tick-borne diseases. J. Travel Med. 2008, 15, 39–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.M.; Kim, M.S.; Park, M.S.; Park, J.H.; Chae, J.S. Identification of Ehrlichia chaffeensis, Anaplasma phagocytophilum, and A. bovis in Haemaphysalis longicornis and Ixodes persulcatus ticks from Korea. Vector-Borne Zoonotic Dis. 2003, 3, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.M.; Yi, Y.H.; Yu, D.H.; Lee, M.J.; Cho, M.R.; Desai, A.R.; Shringi, S.; Klein, T.A.; Kim, H.C.; Song, J.W.; et al. Tick-borne rickettsial pathogens in ticks and small mammals in Korea. Appl. Environ. Microbiol. 2006, 72, 5766–5776. [Google Scholar] [CrossRef] [Green Version]
- Adelson, M.E.; Rao, R.V.S.; Tilton, R.C.; Cabets, K.; Eskow, E.; Fein, L.; Occi, J.L.; Mordechai, E. Prevalence of Borrelia burgdorferi, Bartonella spp., Babesia microti, and Anaplasma phagocytophila in Ixodes scapularis ticks collected in Northern New Jersey. J. Clin. Microbiol. 2004, 42, 2799–2801. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.G.; Noh, B.E.; Lee, H.S.; Kim, T.K.; Song, B.G.; Lee, H.I. Nationwide temporal and geographical distribution of tick populations and phylogenetic analysis of severe fever with thrombocytopenia syndrome virus in ticks in Korea, 2020. Microorganisms 2021, 9, 1630. [Google Scholar] [CrossRef]
- Dumler, J.S.; Madigan, J.E.; Pusterla, N.; Bakken, J.S. Ehrlichioses in humans: Epidemiology, clinical presentation, diagnosis, and treatment. Clin. Infect. Dis. 2007, 45, S45–S51. [Google Scholar] [CrossRef]
- Ismail, N.; Walker, D.H. Balancing protective immunity and immunopathology: A unifying model of monocytotropic ehrlichiosis. Ann. N. Y. Acad. Sci. 2005, 1063, 383–394. [Google Scholar] [CrossRef]
- Chen, S.M.; Dumler, J.S.; Bakken, J.S.; Walker, D.H. Identification of a granulocytotropic Ehrlichia species as the etiologic agent of human disease. J. Clin. Microbiol. 1994, 32, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Markowitz, N.; Hawley, R.C.; Ristic, M.; Cox, D.; McDade, J.E. Human infection with Ehrlichia canis, a leukocytic rickettsia. N. Engl. J. Med. 1987, 316, 853–856. [Google Scholar] [CrossRef]
- Centers of Disease Control and Prevention. Epidemiology and Statistics. Number of Reported Cases of Anaplasmosis and Ehrlichiosis in US. 2000–2019. Available online: https://www.cdc.gov (accessed on 4 August 2021).
- Petrovec, M.; Lotric Furlan, S.; Zupanc, T.A.; Strle, F.; Brouqui, P.; Roux, V.; Dumler, J.S. Human disease in Europe caused by a granulocytic Ehrlichia species. J. Clin. Microbiol. 1997, 35, 1556–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Liu, H.; Xu, B.; Zhang, Z.; Jin, Y.; Li, W.; Lu, Q.; Li, L.; Chang, L.; Zhang, X.; et al. Rural residents in China are at increased risk of exposure to tick-borne pathogens Anaplasma phagocytophilum and Ehrlichia chaffeensis. BioMed Res. Int. 2014, 2014, 313867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, A.; Ando, S.; Kishimoto, T.; Fujita, H.; Kadosaka, T.; Nitta, Y.; Kawabata, H.; Watanabe, H. Presence of a novel Ehrlichia sp. in Ixodes granulatus found in Okinawa, Japan. Microbiol. Immunol. 2009, 53, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, K.; Matsuyama, Y.; Matsuda, H.; Sakamoto, L.; Matsumoto, K.; Yokoyama, N.; Inokuma, H. Detection of Anaplasma bovis and Anaplasma phagocytophilum DNA from Haemaphysalis megaspinosa in Hokkaido, Japan. Vet. Parasitol. 2010, 168, 170–172. [Google Scholar] [CrossRef]
- Heo, E.J.; Park, J.H.; Koo, J.R.; Park, M.S.; Park, M.Y.; Dumler, J.S.; Chae, J.S. Serologic and molecular detection of Ehrlichia chaffeensis and Anaplasma phagocytophila (human granulocytic ehrlichiosis agent) in Korean patients. J. Clin. Microbiol. 2002, 40, 3082–3085. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Yi, J.; Oh, W.S.; Kim, N.H.; Choi, S.J.; Choe, P.G.; Kim, N.J.; Lee, J.K.; Oh, M.D. Human granulocytic anaplasmosis, South Korea, 2013. Emerg. Infect. Dis. 2014, 20, 1708–1711. [Google Scholar] [CrossRef]
- Sachar, D.S. Ehrlichia chaffeensis infection in an active duty soldier stationed in Korea. Med. Surveill. Mon. Rep. 2000, 6, 9–11. [Google Scholar]
- Lee, S.J.; Kim, H.H.; Kim, J.Y.; Yoo, J.E.; Gill, B.C. Laboratory-based diagnostic test results for human granulocytic anaplasmosis in 2020. Public Health Wkly. Rep. PHWR 2021, 14, 2773–2780. [Google Scholar]
- Schouls, L.M.; Van De Pol, I.; Rijpkema, S.G.T.; Schot, C.S. Detection and identification of Ehrlichia, Borrelia burgdorferi sensu lato, and Bartonella species in Dutch Ixodes ricinus ticks. J. Clin. Microbiol. 1999, 37, 2215–2222. [Google Scholar] [CrossRef] [Green Version]
- Im, J.H.; Baek, J.; Durey, A.; Kwon, H.Y.; Chung, M.H.; Lee, J.S. Current status of tick-borne diseases in South Korea. Vector-Borne Zoonotic Dis. 2019, 19, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.J.; Jin, B.C.; Kim, K.H.; Yoo, M.S.; Seong, K.W.; Jeong, S.J.; Hyun, B.H.; Cho, Y.S. Molecular detection and phylogenetic analysis of Anaplasma spp. in Korean native goats from Ulsan Metropolitan City, Korea. Vector-Borne Zoonotic Dis. 2019, 19, 773–776. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.U.; Park, Y.J.; Ryu, J.H.; Jang, D.H.; Hwang, S.; Cho, H.C.; Park, J.; Han, J.I.; Choi, K.S. Identification of zoonotic tick-borne pathogens from Korean water deer (Hydropotes inermis argyropus). Vector-Borne Zoonotic Dis. 2020, 20, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Han, D.G.; Ryu, J.H.; Chae, J.B.; Chae, J.S.; Yu, D.H.; Park, B.K.; Kim, H.C.; Choi, K.S. Molecular detection of Anaplasma bovis in Holstein cattle in the Republic of Korea. Acta Vet. Scand. 2018, 60, 15. [Google Scholar] [CrossRef]
- Bang, M.S.; Kim, C.M.; Pyun, S.H.; Kim, D.M.; Yun, N.R. Molecular investigation of tick-borne pathogens in ticks removed from tick-bitten humans in the southwestern region of the Republic of Korea. PLoS ONE 2021, 16, e0252992. [Google Scholar] [CrossRef]
- Lee, S.H.; Shin, N.R.; Kim, C.M.; Park, S.; Yun, N.R.; Kim, D.M.; Jung, D.S. First identification of Anaplasma phagocytophilum in both a biting tick Ixodes nipponensis and a patient in Korea: A case report. BMC Infect. Dis. 2020, 20, 826. [Google Scholar] [CrossRef]
- Medlock, J.M.; Leach, S.A. Effect of climate change on vector-borne disease risk in the UK. Lancet Infect. Dis. 2015, 15, 721–730. [Google Scholar] [CrossRef]
- Diuk-Wasser, M.A.; Liu, Y.; Steeves, T.K.; Folsom-O’Keefe, C.; Dardick, K.R.; Lepore, T.; Bent, S.J.; Usmani-Brown, S.; Telford, S.R., 3rd; Fish, D.; et al. Monitoring human babesiosis emergence through vector surveillance New England, USA. Emerg. Infect. Dis. 2014, 20, 225–231. [Google Scholar] [CrossRef]
- Hai, V.V.; Almeras, L.; Socolovschi, C.; Raoult, D.; Parola, P.; Pagès, F. Monitoring human tick-borne disease risk and tick bite exposure in Europe: Available tools and promising future methods. Ticks Tick Borne Dis. 2014, 5, 607–619. [Google Scholar] [CrossRef]
- Yamaguti, N.; Tipton, V.J.; Keegan, H.L.; Toshioka, S. Ticks of Japan, Korea, and the Ryukyu Islands. Brigh. Young Univ. Sci. Bull. Biol. Ser. 1971, 15, 1. [Google Scholar]
- Barlough, J.E.; Madigan, J.E.; Derock, E.; Bigornia, L. Nested Polymerase Chain Reaction for Detection of Ehrlichia equi Genomic DNA in Horses and Ticks (Ixodes pacificus). Vet. Parasitol. 1996, 63, 319–329. [Google Scholar] [CrossRef]
- Massung, R.F.; Levin, M.L.; Munderloh, U.G.; Silverman, D.J.; Lynch, M.J.; Gaywee, J.K.; Kurtti, T.J. Isolation and Propagation of the Ap-Variant 1 Strain of Anaplasma phagocytophilum in a Tick Cell Line. J. Clin. Microbiol. 2007, 45, 2138–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Liu, Z.; Niu, Q.; Liu, J.; Xie, J.; Chen, Q.; Chen, Z.; Guan, G.; Liu, G.; Luo, J.; et al. Evaluation of Different Nested PCRs for Detection of Anaplasma phagocytophilum in Ruminants and Ticks. BMC Vet. Res. 2016, 12, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.Y.; Moon, B.C.; Bae, B.K.; Shin, E.-H.; Ko, Y.H.; Kim, Y.-J.; Park, Y.H.; Chae, J.-S. Genetic identification and phylogenetic analysis for Anaplasma and Ehrlichia spiecies in Haemaphysalis longicornis collected from Jeju Island, Korea. J. Bacteriol. Virol. 2009, 39, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.O.; Na, D.K.; Kim, C.M.; Li, Y.H.; Cho, Y.H.; Park, J.H.; Lee, J.H.; Eo, S.K.; Klein, T.A.; Chae, J.S. Identification and Prevalence of Ehrlichia chaffeensis Infection in Haemaphysalis longicornis Ticks from Korea by PCR, Sequencing and Phylogenetic Analysis Based on 16S RRNA Gene. J. Vet. Sci. 2005, 6, 151–155. [Google Scholar] [CrossRef]
- Pritt, B.S.; Sloan, L.M.; Johnson, D.K.; Munderloh, U.G.; Paskewitz, S.M.; McElroy, K.M.; McFadden, J.D.; Binnicker, M.J.; Neitzel, D.F.; Liu, G.; et al. Emergence of a New Pathogenic Ehrlichia species, Wisconsin and Minnesota. N. Engl. J. Med. 2011, 365, 422–429. [Google Scholar] [CrossRef] [Green Version]
- Jahfari, S.; Hofhuis, A.; Fonville, M.; van der Giessen, J.; van Pelt, W.; Sprong, H. Molecular detection of tick-borne pathogens in humans with tick bites and erythema migrans, in the Netherlands. PLoS Negl. Trop. Dis. 2016, 10, e0005042. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Pearson, P.; Dykstra, E.; Andrews, E.S.; Rich, S.M. Human-biting Ixodes ticks and pathogen prevalence from California, Oregon, and Washington. Vector-Borne Zoonotic Dis. 2019, 19, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Moon, K.; Kim, M.; Lee, W.G.; Lee, H.I.; Park, J.K.; Kim, Y.H. Seasonal distribution of Haemaphysalis longicornis (Acari: Ixodidae) and detection of SFTS virus in Gyeongbuk Province, Republic of Korea, 2018. Acta Trop. 2021, 221, 106012. [Google Scholar] [CrossRef]
- Kim-Jeon, M.D.; Jegal, S.; Jun, H.; Jung, H.; Park, S.H.; Ahn, S.K.; Lee, J.; Gong, Y.W.; Joo, K.; Kwon, M.J.; et al. Four year surveillance of the vector hard ticks for SFTS, Ganghwa-do, Republic of Korea. Korean J. Parasitol. 2019, 57, 691–698. [Google Scholar] [CrossRef]
- Park, J.W.; Lee, S.H.; Lee, G.S.; Seo, J.J.; Chung, J.K. Epidemiological characteristics of field tick-borne pathogens in Gwang-ju metropolitan area, South Korea, from 2014 to 2018. Osong Public Health Res. Perspect. 2020, 11, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.B.; Kang, J.G.; Kim, H.C.; Chong, S.T.; Lee, I.Y.; Shin, N.S.; Chae, J.S. Identification of tick species collected from wild boars and habitats of wild boars and domestic pigs in the Republic of Korea. Korean J. Parasitol. 2017, 55, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.S.; Kang, J.G.; Chae, J.B.; Cho, Y.K.; Shin, J.H.; Jheong, W.H.; Chae, J.S. Prevalence of severe fever with thrombocytopenia syndrome virus in ticks collected from national parks in Korea. Vector-Borne Zoonotic Dis. 2019, 19, 284–289. [Google Scholar] [CrossRef]
- Han, S.Y.; Sung, S.H.; Seo, J.W.; Kim, J.H.; Lee, S.J.; Lee, S.J.; Yoo, S.S. Isolation and Identification of Tick-Borne Pathogens in Hard Ticks Collected in Daejeon. Korean J. Vet. Serv. 2021, 44, 93–102. [Google Scholar] [CrossRef]
- Kang, S.W.; Doan, H.T.; Choe, S.E.; Noh, J.H.; Yoo, M.S.; Reddy, K.E.; Kim, Y.H.; Kweon, C.H.; Jung, S.C.; Chang, K.Y. Molecular investigation of tick-borne pathogens in ticks from grazing cattle in Korea. Parasitol. Int. 2013, 62, 276–282. [Google Scholar] [CrossRef]
- Kang, J.G.; Ko, S.; Kim, H.C.; Chong, S.T.; Klein, T.A.; Chae, J.B.; Jo, Y.S.; Choi, K.S.; Yu, D.H.; Park, B.K.; et al. Prevalence of Anaplasma and Bartonella spp. in ticks collected from Korean water deer (Hydropotes inermis argyropus). Korean J. Parasitol. 2016, 54, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.J.; Truong, A.T.; Kim, K.H.; Lim, J.Y.; Min, S.; Kim, H.C.; Yoo, M.S.; Yoon, S.S.; Klein, T.A.; Cho, Y.S. Molecular detection and phylogenetic analysis of tick-borne pathogens in ticks collected from horses in the Republic of Korea. Pathogens 2021, 10, 1069. [Google Scholar] [CrossRef]
- Kang, J.G.; Kim, H.C.; Choi, C.Y.; Nam, H.Y.; Chae, H.Y.; Chong, S.T.; Klein, T.A.; Ko, S.; Chae, J.S. Molecular detection of Anaplasma, Bartonella, and Borrelia species in ticks collected from migratory birds from Hong-do Island, Republic of Korea. Vector-Borne Zoonotic Dis. 2013, 13, 215–225. [Google Scholar] [CrossRef]
- Paddock, C.D.; Childs, J.E. Ehrlichia chaffeensis: A prototypical emerging pathogen. Clin. Microbiol. Rev. 2003, 16, 37–64. [Google Scholar] [CrossRef] [Green Version]
- Chae, J.S.; Yu, D.H.; Shringi, S.; Klein, T.A.; Kim, H.C.; Chong, S.T.; Lee, I.Y.; Foley, J. Microbial pathogens in ticks, rodents and a shrew in Northern Gyeonggi-do near the DMZ, Korea. J. Vet. Sci. 2008, 9, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.J.; Chae, J.S. Molecular detection of Ehrlichia chaffeensis and Anaplasma bovis in the salivary glands from Haemaphysalis longicornis ticks. Vector-Borne Zoonotic Dis. 2010, 10, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.B. 16S rRNA gene sequencing for bacterial pathogen identification in the clinical laboratory. Mol. Diagn. 2001, 6, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Sumner, J.W.; Nicholson, W.L.; Massung, R.F. PCR amplification and comparison of nucleotide sequences from the groESL heat shock operon of Ehrlichia species. J. Clin. Microbiol. 1997, 35, 2087–2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taira, M.; Ando, S.; Kawabata, H.; Fujita, H.; Kadosaka, T.; Sato, H.; Monma, N.; Ohashi, N.; Saijo, M. Isolation and molecular detection of Ehrlichia species from ticks in western, central, and eastern Japan. Ticks Tick Borne Dis. 2019, 10, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, C.; Dibernardo, A.; Koffi, J.; Wood, H.; Leighton, P.A.; Lindsay, L.R. Increased risk of tick-borne diseases with climate and environmental changes. Can. Commun. Dis. Rep. 2019, 45, 83–89. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.; Kamel, M. Climatic changes and their role in emergence and re-emergence of diseases. Environ. Sci. Pollut. Res. Int. 2020, 27, 22336–22352. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Target Gene | Primers | Sequence (5′ to 3′) | Amplicon Size (bp) | PCR Conditions | References | |
|---|---|---|---|---|---|---|
| Anaplasma 16s rRNA | EE1 | 1st | TCCTGGCTCAGAACGAACGCTGGCGGC | 1433 | 94 °C/5 min; 35 cycles: 94 °C/60 s, 50 °C/30 s, 72 °C/1.5 min; 72 °C/10 min | [32] |
| EE2 | AGTCACTGACCCAACCTTAAATGGCTG | |||||
| EE3 | 2nd | GTCGAACGGATTATTCTTTATAGCTTGC | 926 | 94 °C/5 min; 35 cycles: 94 °C/30 s, 50 °C/30 s, 72 °C/60 s; 72 °C/10 min | ||
| EE4 | CCCTTCCGTTAAGAAGGATCTAATCTCC | |||||
| Anaplasma ankA | ANK-F1 | 1st | GAAGAAATTACAACTCCTGAAG | 705 | 94 ℃/2 min; 40 cycles: 94 °C/30 s, 55 °C/30 s, 72 ℃/60 s; 72 °C/5 min | [33] |
| ANK-R1 | CAGCCAGATGCAGTAACGTG | |||||
| ANK-F2 | 2nd | TTGACCGCTGAAGCACTAAC | 664 | 94 °C/2 min; 30 cycles: 94 °C/30 s, 55 °C/30 s, 72 °C/60 s; 72 °C/5 min | ||
| ANK-R2 | ACCATTTGCTTCTTGAGGAG | |||||
| Anaplasma msp4 | MSP4AP5 | 1st | ATGAATTACAGAGAATTGCTTGTAGG | 849 | 94 °C/5 min; 35 cycles: 94 °C/60 s, 54 °C/60 s, 72 °C/60 s; 72 °C/10 min | [34] |
| MSP4AP3 | TTAATTGAAAGCAAATCTTGCTCCTATG | |||||
| MSP4f | 2nd | CTATTGGYGGNGCYAGAGT | 381 | 94 °C/5 min; 30 cycles: 94 °C/30 s, 55 °C/30 s, 72 °C/30 s; 72 °C/10 min | ||
| MSP4r | GTTCATCGAAAATTCCGTGGTA | |||||
| Ehrlichia 16s rRNA | AE1-F | 1st | AAGCTTAACACATGCAAGTCGAA | 1406 | 94 °C/5 min; 40 cycles: 94 °C/60 s, 59 °C/60 s, 72 °C/1.5 min; 72 °C/10 min | [35] |
| AE1-R | AGTCACTGACCCAACCTTAAATG | |||||
| HE1 | 2nd | CAATTGCTTATAACCTTTTGGTTATAAAT | 390 | 94 °C/3 min; 3 cycles: 94 °C/60 s, 55 °C/2 min, 72 °C/1.5 min; 92 °C/60 s; 37 cycles: 92 °C/60 s, 55 °C/2 min, 72 °C/60 s; 72 °C/10 min | [36] | |
| HE3 | TATAGGTACCGTCATTATCTTCCCTAT | |||||
| Ehrlichia groEL | GR0607F | 1st | GAAGATGCWGTWGGWTGTACKGC | 664 | 95 °C/5 min; 35 cycles: 95 °C/30 s, 54 °C/30 s, 72 °C/60 s; 72 °C/10 min | [37] |
| GR01294R | AGMGCTTCWCCTTCWACRTCYTC | |||||
| GR0677F | 2nd | ATTACTCAGAGTGCTTCTCARTG | 315 | 95 °C/5 min; 30 cycles: 94 °C/30 s, 57 °C/30 s, 72 °C/60 s; 72 °C/10 min | ||
| GR01121R | TGCATACCRTCAGTYTTTTCAAC |
| Species | Stage | No. of Collected Ticks | Anaplasma phagocytophilum | Ehrlichia sp. | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| March | April | May | June | July | August | September | October | Total (%) | Positive (%) | 95% CI | Positive (%) | 95% CI | ||
| Amblyomma testudinarium | Female | - | - | 1 | 5 | 1 | 1 | 2 | 1 | 11 (6.1) | 0 | 0 | 0 | 0 |
| Larva | - | - | - | - | - | - | - | 1 | 1 (0.6) | 0 | 0 | 0 | 0 | |
| Male | - | - | 1 | 1 | - | - | - | 1 | 3 (1.7) | 0 | 0 | 0 | 0 | |
| Nymph | - | 2 | 3 | 4 | 6 | 1 | - | 1 | 17 (9.4) | 0 | 0 | 0 | 0 | |
| Haemaphysalis flava | Female | - | - | - | - | - | - | - | 2 | 2 (1.1) | 0 | 0 | 0 | 0 |
| Male | - | - | 1 | - | - | - | - | 1 | 2 (1.1) | 0 | 0 | 0 | 0 | |
| Nymph | - | 2 | 2 | - | - | - | - | - | 4 (2.2) | 0 | 0 | 0 | 0 | |
| H. longicornis | Female | 1 | - | 7 | 6 | 21 | 34 (2 ‡) | 8 | - | 77 (42.8) | 0 | 0 | 2 (2.6) | 0.3–9.3 |
| Male | - | - | - | 1 | - | 1 | - | - | 2 (1.1) | 0 | 0 | 0 | 0 | |
| Nymph | - | 3 | 17 | 8 | 11 | 3 | 4 | 1 | 47 (26.1) | 0 | 0 | 0 | 0 | |
| Ixodes nipponensis | Female | - | 1 (1 †) | 4 | 4 (1 †) | 1 | - | - | - | 10 (5.6) | 2 (20.0) | 2.4–72.3 | 0 | 0 |
| Nymph | - | - | 1 | - | - | - | - | - | 1 (0.6) | 0 | 0 | 0 | 0 | |
| I. persulcatus | Female | - | - | 3 (1 †) | - | - | - | - | - | 3 (1.7) | 1 (33.3) | 0.8–185.7 | 0 | 0 |
| Total | Female | 1 | 1 | 15 | 15 | 23 | 35 | 10 | 3 | 103 (57.2) | 3 (2.9) | 0.6–8.5 | 2 (1.9) | 0.2–7.0 |
| Larva | - | - | - | - | - | - | - | 1 | 1 (0.6) | 0 | 0 | 0 | 0 | |
| Male | - | - | 2 | 2 | - | 1 | - | 2 | 7 (3.9) | 0 | 0 | 0 | 0 | |
| Nymph | - | 7 | 23 | 12 | 17 | 4 | 4 | 2 | 69 (38.3) | 0 | 0 | 0 | 0 | |
| Total | 1 | 8 | 40 | 29 | 40 | 40 | 14 | 8 | 180 (100.0) | 3 (1.7) | 0.3–4.9 | 2 (1.1) | 0.1–4.0 | |
| Region | Species | Total (%) | ||||
|---|---|---|---|---|---|---|
| Amblyomma testudinarium | Haemaphysalis flava | Haemaphysalis longicornis | Ixodes nipponensis | Ixodes persulcatus | ||
| Seoul Special City | 1 | 1 | 2 (1.1) | |||
| Gyeonggi-do Province | 0 | 2 | 29 | 2 | 33 (18.3) | |
| Gwangwon-do Province | 0 | 5 | 2 | 7 (3.9) | ||
| Chungcheongbuk-do Province | 0 | 9 | 1 | 1 | 11 (6.1) | |
| Chungcheongnam-do Province | 2 | 16 | 5 | 23 (12.8) | ||
| Jeollanam-do Province | 2 | 4 | 6 (3.3) | |||
| Gyeongsangbuk-do Province | 5 | 1 | 31 | 1 | 38 (21.1) | |
| Gyeongsangnam-do Province | 18 | 3 | 16 | 37 (20.6) | ||
| Jeju special self-governing Province | 0 | 2 | 2 (1.1) | |||
| Metropolitan area * | 4 | 2 | 12 | 2 | 20 (11.1) | |
| Unknown | 1 | 1 (0.6) | ||||
| Total | 32 | 8 | 126 | 11 | 3 | 180 (100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.-J.; Seo, J.Y.; Kim, S.Y.; Lee, H.I. Molecular Detection of Anaplasma phagocytophilum and Ehrlichia Species in Ticks Removed from Humans in the Republic of Korea. Microorganisms 2022, 10, 1224. https://doi.org/10.3390/microorganisms10061224
Kim Y-J, Seo JY, Kim SY, Lee HI. Molecular Detection of Anaplasma phagocytophilum and Ehrlichia Species in Ticks Removed from Humans in the Republic of Korea. Microorganisms. 2022; 10(6):1224. https://doi.org/10.3390/microorganisms10061224
Chicago/Turabian StyleKim, Yu-Jung, Ji Ye Seo, Seong Yoon Kim, and Hee Il Lee. 2022. "Molecular Detection of Anaplasma phagocytophilum and Ehrlichia Species in Ticks Removed from Humans in the Republic of Korea" Microorganisms 10, no. 6: 1224. https://doi.org/10.3390/microorganisms10061224