
Assessment of Psyllid Handling and DNA Extraction Methods in the Detection of ‘Candidatus Liberibacter Solanacearum’ by qPCR


Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Insects
2.2. Insect Preparations
2.3. DNA Extraction Methods
2.4. DNA Yield and Purity
2.5. Amplification Conditions by Conventional and qPCR
2.6. Morphological State of Psyllids after DNA Extraction
2.7. Time Consumed, Cost Estimation, and Residues Generated
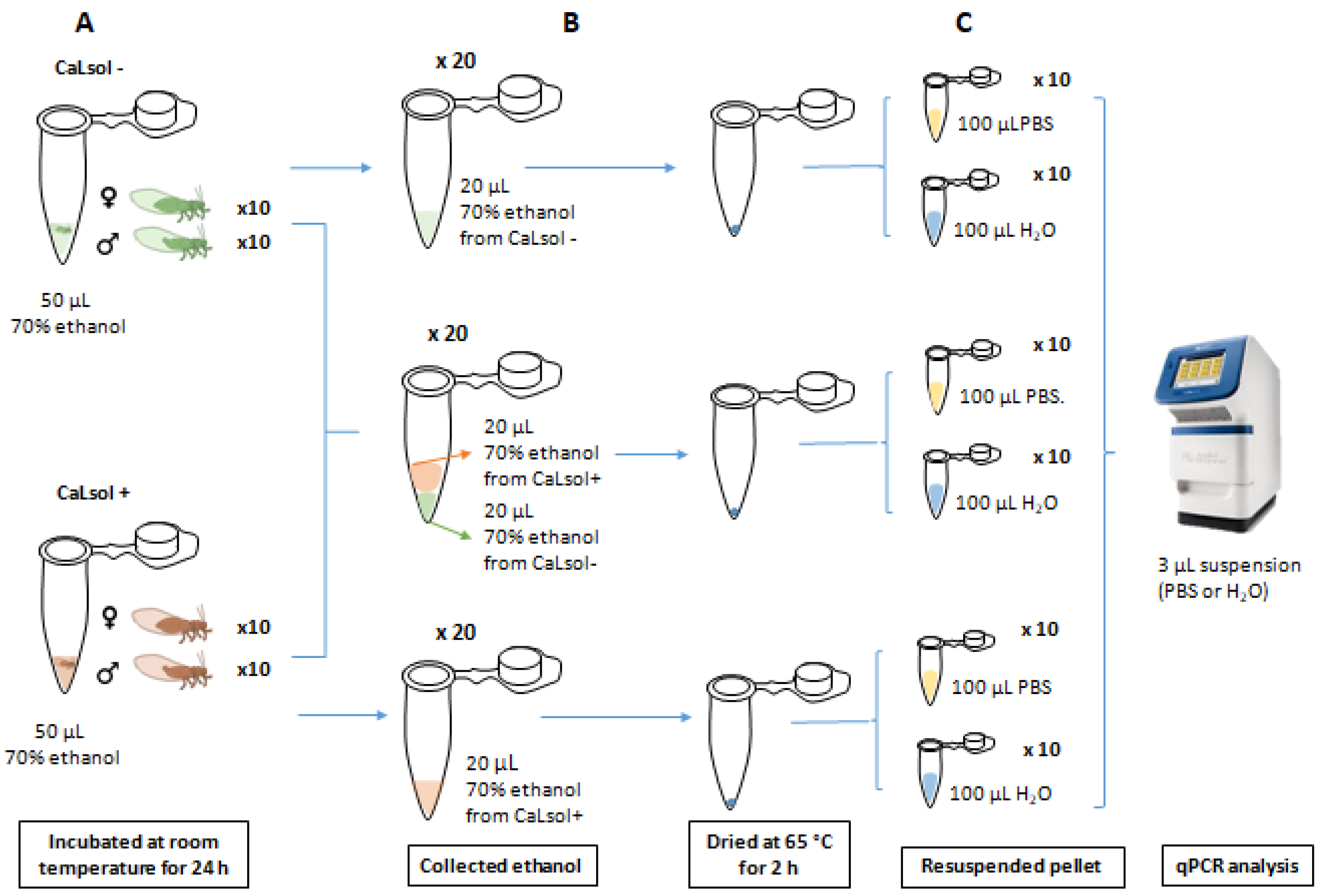
2.8. Cross-Contamination Assays
2.8.1. Assay 1
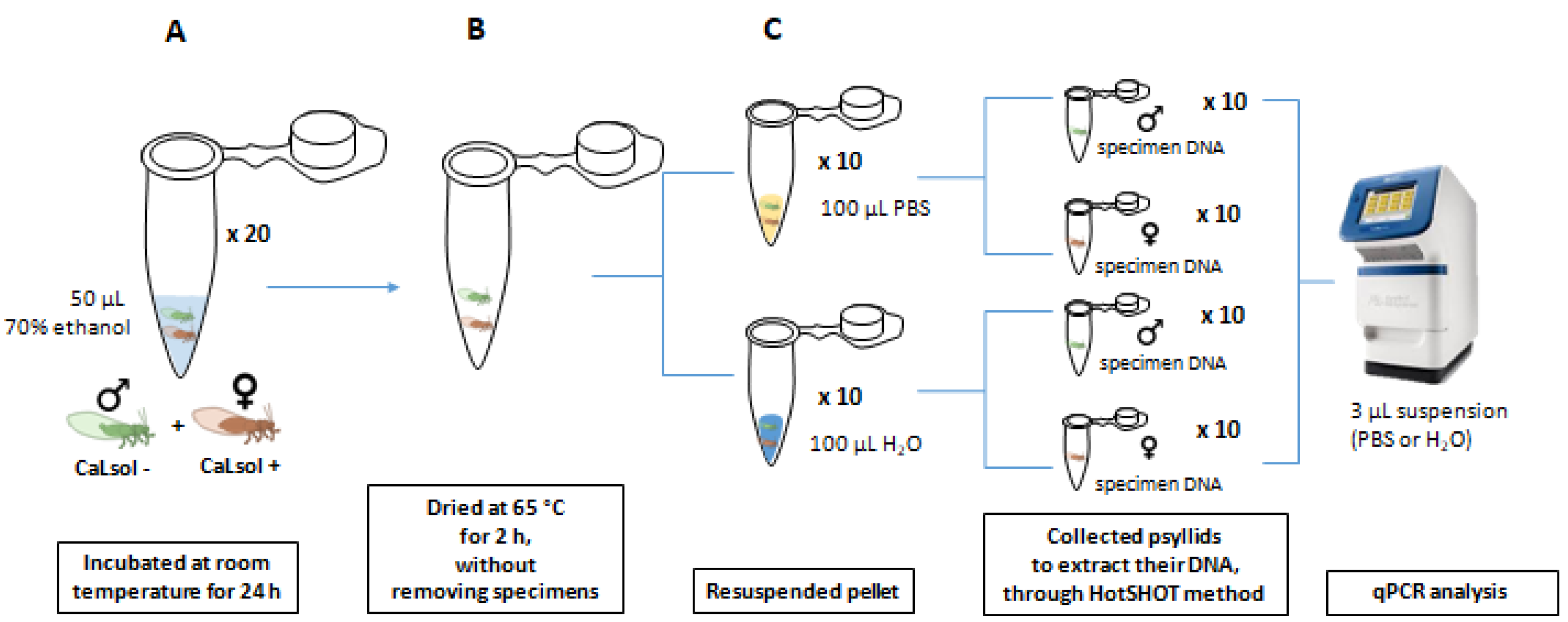
2.8.2. Assay 2
2.8.3. Assay 3
2.9. Data Analysis
3. Results
3.1. DNA Yield and Purity
3.2. Conventional PCR and qPCR
3.3. Morphological State of Psyllids after DNA Extraction
3.4. Time Consumed, Cost Estimation, and Residues Generated
3.5. Cross Contamination Assay
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jagoueix, S.; Bové, J.M.; Garnier, M. The phloem-limited bacterium of greening disease of citrus is a member of the subdivision of the proteobacteria. Int. J. Syst. Bacteriol. 1994, 44, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, A.K.; Trumble, J.T.; Stouthamer, R.; Paine, T.D. A new huanglongbing species, ‘Candidatus Liberibacter psyllaurous’, found to infect tomato and potato, is vectored by the psyllid Bactericera cockerelli (Sulc). Appl. Environ. Microbiol. 2008, 74, 5862–5865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liefting, L.W.; Perez-Egusquiza, Z.C.; Clover, G.R.G. A New ‘Candidatus Liberibacter species’ in Solanum tuberosum in New Zealand. Plant Dis. 2009, 92, 1474. [Google Scholar] [CrossRef] [Green Version]
- Liefting, L.W.; Weir, B.S.; Pennycook, S.R.; Clover, G.R.G. ‘Candidatus Liberibacter solanacearum’, associated with plants in the family Solanaceae. Int. J. Syst. Evol. Microbiol. 2009, 59, 2274–2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munyaneza, J.E. Zebra Chip Disease of potato: Biology, epidemiology and Management. Am. J. Potato Res. 2012, 89, 329–350. [Google Scholar] [CrossRef] [Green Version]
- Haapalainen, M. Biology and epidemics of ‘Candidatus Liberibacter specie’, psyllid-transmitted plant-pathogenic bacteria. Ann. Appl. Biol. 2014, 165, 172–198. [Google Scholar] [CrossRef]
- Teresani, G.R.; Bertolini, E.; Alfaro-Fernández, A.; Martínez, C.; Tanaka, F.A.O.; Kitajima, E.W.; Roselló, M.; Sanjuan, S.; Ferrándiz, J.C.; López, M.M.; et al. Association of ‘Candidatus Liberibacter solanacearum’ with a vegetative disorder of celery in Spain and development of a real-time PCR method for its detection. Phytopathology 2014, 104, 804–811. [Google Scholar] [CrossRef] [Green Version]
- Nelson, W.R.; Fisher, T.W.; Munyaneza, J.E. Haplotypes of ‘Candidatus Liberibacter solanacearum’ suggest long-standing separation. Eur. J. Plant Pathol. 2011, 130, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Nelson, W.R.; Sengoda, V.G.; Alfaro-Fernández, A.O.; Font, M.I.; Crosslin, J.M.; Munyaneza, J.E. A new haplotype of ‘Candidatus Liberibacter solanacearum’ identified in the Mediterranean region. Eur. J. Plant Pathol. 2013, 135, 633–639. [Google Scholar] [CrossRef]
- Haapalainen, M.L.; Wang, J.; Latvala, S.; Lehtonen, M.T.; Pirhonen, M.; Nissinen, A. Genetic variation of ‘Candidatus Liberibacter solanacearum’ haplotype C and identification of a novel haplotype from Trioza urticae and stinging nettle. Phytopathology 2018, 108, 925–934. [Google Scholar] [CrossRef] [Green Version]
- Swisher-Grimm, K.D.; Garczynski, S.F. Identification of a new haplotype of ‘Candidatus Liberibacter solanacearum’ in Solanum tuberosum. Plant Dis. 2019, 103, 468–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haapalainen, M.; Latvala, S.; Wickström, A.; Wang, J.; Pirhonen, M.; Nissinen, A.I. A novel haplotype of ‘Candidatus Liberibacter solanacearum’ found in Apiaceae and Polygonaceae family plants. Eur. J. Plant Pathol. 2020, 156, 413–423. [Google Scholar] [CrossRef] [Green Version]
- EPPO, European and Mediterranean Plant Protection. PM 7/143 (1) ‘Candidatus Liberibacter solanacearum’. EPPO Bull. 2020, 50, 49–68. [Google Scholar] [CrossRef] [Green Version]
- Munyaneza, J.E.; Fisher, T.W.; Sengoda, V.G.; Garczynski, S.F.; Nissinen, A.; Lemmetty, A. Association of ‘Candidatus Liberibacter solanacearum’ with the psyllid, Trioza apicalis (Hemiptera: Triozidae) in Europe. J. Econ. Entomol. 2010, 103, 1060–1070. [Google Scholar] [CrossRef] [PubMed]
- Munyaneza, J.E.; Lemmetty, A.; Nissinen, A.; Sengoda, V.G.; Fisher, T.W. Molecular detection of aster yellows phytoplasma and ‘Candidatus Liberibacter solanacearum’ in carrots affected by the psyllid Trioza apicalis (Hemiptera: Triozidae) in Finland. J. Plant Pathol. 2011, 93, 697–700. [Google Scholar]
- Munyaneza, J.E. Zebra chip disease, ‘Candidatus Liberibacter’, and potato psyllid: A global threat to the potato industry. Am. J. Potato Res. 2015, 92, 230–235. [Google Scholar] [CrossRef]
- Alfaro-Fernández, A.; Siverio, F.; Cebrián, M.C.; Villaescusa, F.J.; Font, M.I. ‘Candidatus Liberibacter solanacearum’ associated with Bactericera trigonica-affected carrots in the Canary Islands. Plant Dis. 2012, 96, 581. [Google Scholar] [CrossRef]
- Alfaro-Fernández, A.; Cebrián, M.C.; Villaescusa, F.J.; Hermoso de Mendoza, A.; Ferrándiz, J.C.; Sanjuán, S.; Font, M.I. First report of ‘Candidatus Liberibacter solanacearum’ in carrots in mainland Spain. Plant Dis. 2012, 96, 582. [Google Scholar] [CrossRef]
- Loiseau, M.; Garnier, S.; Boirin, V.; Merieae, M.; Leguay, A.; Renaudin, I.; Renvoisé, J.P.; Gentit, P. First report of ‘Candidatus Liberibacter solanacearum’ in carrot in France. Plant Dis. 2014, 98, 839. [Google Scholar] [CrossRef]
- Tahzima, R.; Maes, M.; Achbani, E.H.; Swisher, K.D.; Munyaneza, J.E.; De Jonghe, K. First Report of ‘Candidatus Liberibacter solanacearum’ on carrot in Africa. Plant Dis. 2014, 98, 1426. [Google Scholar] [CrossRef]
- Holeva, M.C.; Glynos, P.E.; Karafla, C.D. First report of ‘Candidatus Liberibacter solanacearum’ on carrot in Greece. Plant Dis. 2017, 101, 1654. [Google Scholar] [CrossRef]
- Ben Othmen, S.; Morán, F.E.; Navarro, I.; Barbé, S.; Martínez, C.; Marco-Noales, E.; Chermiti, B.; López, M.M. ‘Candidatus Liberibacter solanacearum’ haplotypes D and E in carrot plants and seeds in Tunisia. J. Plant Pathol. 2018, 100, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Mawassi, M.; Dror, O.; Bar-Joseph, M.; Piasezky, A.; Sjölund, J.M.; Levitzky, N.; Shoshana, N.; Meslenin, L.; Haviv, S.; Porat, C.; et al. ‘Candidatus Liberibacter solanacearum’ is tightly associated with carrot yellows symptoms in Israel and transmitted by the prevalent psyllid vector Bactericera trigonica. Phytopathology 2018, 108, 1056–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haapalainen, M.; Latvala, S.; Rastas, M.; Wang, J.; Hannukkala, A.; Pirhonen, M.; Nissinen, A.I. A Carrot Pathogen ‘Candidatus Liberibacter solanacearum’ Haplotype C detected in symptomless potato plants in Finland. Potato Res. 2018, 61, 1–20. [Google Scholar] [CrossRef] [Green Version]
- MAPA. Programa Para La Aplicación De La Normativa Fitosanitaria De La Patata; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2019; Available online: https://www.mapa.gob.es/es/agricultura/temas/sanidad-vegetal/manualpatata2019_tcm30-135971.pdf (accessed on 26 July 2021).
- Ruiz-Padilla, A.; Redondo, C.; Asensio, A.; Garita-Cambronero, J.; Martínez, C.; Pérez-Padilla, V.; Marquínez, R.; Collar, J.; García-Méndez, E.; Alfaro-Fernández, A.; et al. Assessment of multilocus sequence analysis (MLSA) for identification of ‘Candidatus Liberibacter solanacearum’ from different host plants in Spain. Microorganisms 2020, 8, 1446. [Google Scholar] [CrossRef] [PubMed]
- Mauck, K.E.; Sun, P.; Meduri, V.; Hansen, A.K. New Ca. Liberibacter psyllaurous’ haplotype resurrected from a 49-year-old specimen of Solanum umbelliferum: A native host of the psyllid vector. Sci. Rep. 2019, 9, 9530. [Google Scholar] [CrossRef]
- Contreras-Rendón, A.; Sánchez-Pale, J.R.; Fuentes-Aragón, D.; Alanís-Martínez, I.; Silva-Rojas, H.V. Conventional and qPCR reveals the presence of ‘Candidatus Liberibacter solanacearum’ haplotypes A, and B in Physalis philadelphica plant, seed, and Βactericera cockerelli psyllids, with the assignment of a new haplotype H in Convolvulaceae. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2020, 113, 533–551. [Google Scholar] [CrossRef]
- Sumner-Kalkun, J.C.; Highet, F.; Arnsdorf, Y.M.; Back, E.; Carnegie, M.; Madden, S.; Carboni, S.; Billaud, W.; Lawrence, Z.; Kenyon, D. ‘Candidatus Liberibacter solanacearum’ distribution and diversity in Scotland and the characterisation of novel haplotypes from Craspedolepta spp. (Psyllidae: Aphalaridae). Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Antolínez, C.A.; Fereres, A.; Moreno, A. Risk assessment of ‘Candidatus Liberibacter solanacearum’ transmission by the psyllids Bactericera trigonica and B. tremblayi from Apiaceae crops to potato. Sci. Rep. 2017, 7, 45534. [Google Scholar] [CrossRef] [Green Version]
- Teresani, G.R.; Hernández, E.; Bertolini, E.; Siverio, F.; Moreno, A.; Fereres, A.; Cambra, M. Transmission of ‘Candidatus Liberibacter solanacearum’ by Bactericera trigonica Hodkinson to vegetable hosts. Span. J. Agric. Res. 2017, 15, e1011. [Google Scholar] [CrossRef]
- EPPO, European and Mediterranean Plant Protection. EPPO Global Database: Bactericera Trigonica. Available online: https://gd.eppo.int/taxon/BCTCTR (accessed on 6 October 2020).
- Mustafa, T.; Horton, D.R.; Cooper, W.R.; Swisher, K.D.; Zack, S.; Pappu, H.R.; Munyaneza, J.E. Use of electrical penetration graph technology to examine transmission of ‘Candidatus Liberibacter solanacearum’ to potato by three haplotypes of potato psyllid (Bactericera cockerelli; Hemiptera: Triozidae). PLoS ONE 2015, 10, e0138946. [Google Scholar] [CrossRef] [PubMed]
- Teresani, G.; Hernández, E.; Bertolini, E.; Siverio, F.; Marroquín, C.; Molina, J.; de Mendoza, A.H.; Cambra, M. Search for potential vectors of ‘Candidatus Liberibacter solanacearum’: Population dynamics in host crops. Span. J. Agric. Res. 2015, 13, e1002. [Google Scholar] [CrossRef] [Green Version]
- Antolínez, C.A.; Moreno, A.; Ontiveros, I.; Pla, S.; Plaza, M. Seasonal abundance of psyllid species associated with carrots and potato fields in Spain. Insects 2019, 10, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjölund, M.J.; Clark, M.; Carnegie, M.; Greenslade, A.F.C.; Ouvrard, D.; Highet, F.; Sigvald, R.; Bell, J.R.; Arnsdorf, Y.M.; Cairns, R.; et al. First report of ‘Candidatus Liberibacter solanacearum’ in the United Kingdom in the psyllid Trioza anthrisci. New Dis. Rep. 2017, 36, 4. [Google Scholar] [CrossRef] [Green Version]
- Tian, E.W.; Yu, H. A Simple and rapid DNA extraction protocol of small insects for PCR amplification. Entomol. News. 2014, 123, 303. [Google Scholar] [CrossRef]
- Zeh, D.W.; May, C.A.; Coffroth, M.A.; Bermingham, E. MboI and Macrohaltica—Quality of DNA fingerprints is strongly enzyme-dependent in an insect (Coleoptera). Mol. Ecol. 1993, 2, 61–63. [Google Scholar] [CrossRef]
- Ball, S.L.; Armstrong, K.F. Rapid, One-Step DNA Extraction for insect pest identification by using DNA barcodes. J. Econ. Entomol. 2008, 101, 523–532. [Google Scholar] [CrossRef]
- Calderón-Cortés, N.; Quesada, M.; Cano-Camacho, H.; Zavala-Páramo, G. A simple and rapid method for DNA isolation from xylophagous insects. Int. J. Mol. Sci. 2010, 11, 5056–5064. [Google Scholar] [CrossRef]
- Chen, H.; Rangasamy, M.; Tan, S.Y.; Wang, H.; Siegfried, B.D. Evaluation of five methods for total DNA extraction from western corn rootworm beetles. PLoS ONE 2010, 5, e11963. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, N.; Luís, M.I.; Revers, F. Developing a rapid, efficient and low cost method for rapid DNA extraction from arthropods. Ciênc. Rural 2011, 419419, 1563–1570. [Google Scholar]
- Musapa, M.; Kumwenda, T.; Mkulama, M.; Chishimba, S.; Norris, D.E.; Thuma, P.E.; Mharakurwa, S. A Simple Chelex Protocol for DNA Extraction from Anopheles spp. J. Vis. Exp. 2013, 71, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, M.T.S.; Gomes-Júnior, P.P.; Batista, M.V.A.; Leal-Balbino, T.C.; Araujo, A.L.; Balbino, V.Q. Novel DNA extraction assay for molecular idenification of Aedes spp. eggs. Genet. Mol. Res. 2014, 13, 8776–8782. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.J.; Simon, C. Protocol for Arthropods. Ann. Entomol. Soc. Am. 1995, 88, 281–283. [Google Scholar] [CrossRef]
- Gilbert, M.T.P.; Moore, W.; Melchior, L.; Worebey, M. DNA extraction from dry museum beetles without conferring external morphological damage. PLoS ONE 2007, 2, e272. [Google Scholar] [CrossRef] [Green Version]
- Castalanelli, M.A.; Severtson, D.L.; Brumley, C.J.; Szito, A.; Foottit, R.G.; Grimm, M.; Munyard, K.; Groth, D.M. A rapid non-destructive DNA extraction method for insects and other arthropods. J. Asia Pac. Entomol. 2010, 13, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Miura, K.; Higashiura, Y.; Maeto, K. Evaluation of easy, non-destructive methods of DNA extraction from minute insects. Appl. Entomol. Zool. 2017, 52, 349–352. [Google Scholar] [CrossRef] [Green Version]
- Sjölund, M.J. Non-destructive DNA extraction from Psyllids. POnTE, Pest Organisms Threatening Europe. 2017. Available online: https://www.ponteproject.eu/protocols-calsol/non-destructive-dna-extraction-psyllids/ (accessed on 5 March 2021).
- Asghar, U.; Malik, M.F.; Anwar, F.; Javed, A.; Raza, A. DNA extraction from insects by using different techniques: A review. Adv. Entomol. 2015, 3, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Abad, J.A.; French-Monar, R.D.; Rascoe, J.; Wen, A.; Gudmestad, N.C.; Secor, G.A.; Lee, M.; Duan, Y.; Levy, L. Multiplex real-time PCR for detection, identification and quantification of ‘Candidatus Liberibacter solanacearum’ in potato plants with zebra chip. J. Microbiol. Methods. 2009, 78, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Casquet, J.; Thebaud, C.; Gillespie, R.G. Chelex without boiling, a rapid and easy technique to obtain stable amplifiable DNA from small amounts of ethanol-stored spiders. Mol. Ecol. Resour. 2012, 12, 136–141. [Google Scholar] [CrossRef]
- Bertolini, E.; Felipe, R.T.A.; Sauer, A.V.; Lopes, S.A.; Arilla, A.; Vidal, E.; Mourão Filho, F.D.A.A.; Nunes, W.M.C.; Bové, J.M.; López, M.M.; et al. Tissue-print and squash real-time PCR for direct detection of ‘Candidatus Liberibacter’ species in citrus plants and psyllid vectors. Plant Pathol. 2014, 63, 1149–1158. [Google Scholar] [CrossRef]
- Collado-Romero, M.; Mercado-Blanco, J.; Olivares-García, C.; Valverde-Corredor, A.; Jiménez-Díaz, R.M. Molecular variability within and among Verticillium dahliae vegetative compatibility groups determined by fluorescent amplified fragment length polymorphism and polymerase chain reaction markers. Phytopathology 2006, 96, 485–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truett, G.E.; Heeger, P.; Mynatt, R.L.; Walker, J.A.; Warman, M.L. Myocardial atrophy and chronic mechanical unloading of the failing human heart: Implications for cardiac assist device-induced myocardial recovery. J. Am. Coll. Cardiol. 2014, 64, 1602–1612. [Google Scholar]
- Bioline. Meridian Bioscience. TRIsureTM. 2020. Available online: https://www.bioline.com/trisure.html (accessed on 20 November 2020).
- Ravindran, A.; Levy, J.; Pierson, E.; Gross, D.C. Development of primers for improved PCR detection of the potato zebra chip pathogen, ‘Candidatus Liberibacter solanacearum’. Plant Dis. 2011, 95, 1542–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.S.; Dai, T.M.; Tian, H.; Wan, F.H.; Zhang, G.F. Comparative analysis of eight DNA extraction methods for molecular research in mealybugs. PLoS ONE 2019, 14, e0226818. [Google Scholar] [CrossRef] [PubMed]
- Hajibabaei, M.; DeWaard, J.R.; Ivanova, N.V.; Ratnasingham, S.; Dooh, R.T.; Kirk, S.L.; Mackie, P.M.; Hebert, P.D. Critical factors for assembling a high volume of DNA barcodes. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1959–1967. [Google Scholar] [CrossRef] [Green Version]
- Dittrich-Schröder, G.; Wingfield, M.J.; Klein, H.; Slippers, B. DNA extraction techniques for DNA barcoding of minute gall-inhabiting wasps. Mol. Ecol. Resour. 2012, 12, 109–115. [Google Scholar] [CrossRef]
- Olson, N.D.; Morrow, J.B. DNA extract characterization process for microbial detection methods development and validation. BMC Res. Notes 2012, 5, 668. [Google Scholar] [CrossRef] [Green Version]
- Konakandla, B.; Park, Y.; Margolies, D. Whole genome amplification of Chelex-extracted DNA from a single mite: A method for studying genetics of the predatory mite Phytoseiulus persimilis. Exp. Appl. Acarol. 2006, 40, 241–247. [Google Scholar] [CrossRef]
- Martín-Platero, A.M.; Peralta-Sánchez, J.M.; Soler, J.J.; Martínez-Bueno, M. Chelex-based DNA isolation procedure for the identification of microbial communities of eggshell surfaces. Anal. Biochem. 2010, 397, 253–255. [Google Scholar] [CrossRef]
- Stephen, C.Y.; Lin, S.; Lai, K. An evaluation of the performance of five extraction methods: Chelex® 100, QIAamp® DNA Blood Mini Kit, QIAamp® DNA Investigator Kit, QIAsymphony® DNA Investigator® Kit and DNA IQTM. Sci. Justice 2015, 55, 200–208. [Google Scholar]
- Turan, C.; Nanni, I.M.; Brunelli, A.; Collina, M. New rapid DNA extraction method with Chelex from Venturia inaequalis spores. J. Microbiol. Methods. 2015, 115, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J. The Condensed Protocols from Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2006. [Google Scholar]
- Fang, G.; Hammar, S.; Grumet, R. A quick and inexpensive method for removing polysaccharides from plant genomic DNA. Biotechniques 1992, 13, 52–54. [Google Scholar]
- Chakraborty, S.; Saha, A.; Neelavar Ananthram, A. Comparison of DNA extraction methods for non-marine molluscs: Is modified CTAB DNA extraction method more efficient than DNA extraction kits? 3 Biotech 2020, 10, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Qiagen. Supplementary Protocol: Purification of Total DNA from Insects Using the DNeasy® Blood &Tissue Kit. Available online: https://www.qiagen.com/br/resources/faq?id=58ddc6c7-d1f4-45d8-9156-8174fed15bf3&lang=en (accessed on 6 May 2020).
- Merck. Safety Data Sheet: Hexadecyltrimethylammonium Bromide Version 6.0. Available online: https://www.sigmaaldrich.com/MSDS/MSDS/DisplayMSDSPage.do?country=ES&language=es&productNumber=H9151&brand=SIGMA&PageToGoToURL=https%3A%2F%2Fwww.sigmaaldrich.com%2Fcatalog%2Fproduct%2Fsigma%2Fh9151%3Flang%3Des (accessed on 6 May 2020).
- Ashfaq, M.; Hebert, P.D.N. DNA barcodes for bio-surveillance: Regulated and economically important arthropod plant pests. Genome 2016, 59, 933–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, D.C.; Saillard, C.; Eveillard, S.; Danet, J.L.; Ayres, A.J.; Bové, J.M. ‘Candidatus Liberibacter americanus’, associated with citrus Huanglongbing (greening disease) in São Paulo State, Brazil. Int. J. Syst. Evol. Microbiol. 2005, 55, 1857–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capoor, D.P.; Rao, D.G.; Viawanath, S.M. Diaphoria citri Kuway., a vector of the greening disease of citrus in India. Indian Agric. Res. Inst. 1967, 376, 572–579. [Google Scholar]
- McClean, A.P.D.; Oberholzer, P.C.J. Citrus psylla, a vector of the greening disease of Sweet Orange. S. Afr. J. Agric. Sci. 1965, 81, 297–298. [Google Scholar]
- Manjunath, K.L.; Halbert, S.E.; Ramadugu, C.; Webb, S.; Lee, R.F. Detection of ‘Candidatus Liberibacter asiaticus’ in Diaphorina citri and its importance in the management of citrus huanglongbing in Florida. Phytopathology 2008, 98, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Tabachnick, W.J. Diaphorina citri (Hemiptera: Liviidae) Vector Competence for the Citrus Greening Pathogen ‘Candidatus Liberibacter Asiaticus’. J. Econ. Entomol. 2015, 108, 839–848. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen Preparation | ||||
|---|---|---|---|---|
| Methods * | Whole | Grinding | Cut-Off Head | Punctured Abdomen |
| CTAB | N/A | 7 ♀ + 7 ♂ | N/A | N/A |
| Chelex | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ |
| TRIsureTM | N/A | 7 ♀ + 7 ♂ | N/A | N/A |
| Squashed on membrane | N/A | 7 ♀ + 7 ♂ | N/A | N/A |
| HotSHOT | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ |
| PBS | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ |
| DNeasy® | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ | 7 ♀ + 7 ♂ |
| Methods | DNA Yield (ng·mg−1) * | DNA Purity (A260/A280) |
|---|---|---|
| CTAB | 1757 ± 296 d | 1.89 ± 0.11 a |
| Chelex | 24,328 ± 1638 a | 1.15 ± 0.03 b |
| TRIsureTM | 20,746 ± 4511 ab | 1.59 ± 0.07 a |
| Squashed on membrane | 6471 ± 713 cd | 0.67 ± 0.05 c |
| HotSHOT | 11,964 ± 1187 bc | 1.03 ± 0.04 b |
| PBS | 3850 ± 489 cd | 0.62 ± 0.06 c |
| DNeasy® | 3271 ± 380 d | 1.85 ± 0.07 a |
| Specimen Preparations | DNA Extraction Methods | |||
|---|---|---|---|---|
| Chelex | HotSHOT | PBS | DNeasy® | |
| Whole | 31,164 ± 3261 a | 8078 ± 1271 a | 1582 ± 211 b | 2386 ± 256 b |
| Grinding | 24,328 ± 1638 a | 11,964 ± 1187 a | 3850 ± 489 a | 3271 ± 380 ab |
| Cut off head | 32,936 ± 2816 a | 8664 ± 1021 a | 2750 ± 388 ab | 3121 ± 320 ab |
| Punctured abdomen | 31,814 ± 1956 a | 10,643 ± 1523 a | 2661 ± 357 ab | 3786 ± 373 a |
| Specimen Preparations | DNA Extraction Methods | |||
|---|---|---|---|---|
| Chelex | HotSHOT | PBS | DNeasy® | |
| Whole | 1.17 ± 0.02 a | 0.80 ± 0.06 b | 1.38 ± 0.16 a | 1.62 ± 0.09 a |
| Grinding | 1.15 ± 0.03 a | 1.03 ± 0.04 a | 0.62 ± 0.06 b | 1.85 ± 0.07 a |
| Cut off head | 1.19 ± 0.02 a | 0.98 ± 0.04 ab | 1.15 ± 0.07 a | 1.78 ± 0.06 a |
| Punctured abdomen | 1.10 ± 0.03 a | 0.79 ± 0.06 b | 1.23 ± 0.11 a | 1.85 ± 0.07 a |
| DNA Extraction Methods | qPCR Protocols * | |||
|---|---|---|---|---|
| CaLsol | Lso | |||
| PP | Cq | PP | Cq | |
| CTAB | 100 | 21.9 ± 1.0 ab | 100 | 22.4 ± 1.1 c |
| Chelex | 100 | 20.1 ± 0.5 ab | 100 | 18.7 ± 0.4 ab |
| TRIsureTM | 100 | 29.8 ± 0.7 c | 92.8 | 27.3 ± 0.9 c |
| Squashed on membrane | 100 | 29.8 ± 0.8 c | 100 | 24.8 ± 1.0 c |
| HotSHOT | 100 | 18.1 ± 0.4 a | 100 | 16.6 ± 0.2 a |
| PBS | 64.3 | 27.5 ± 1.2 bc | 64.3 | 21.6 ± 0.7 bc |
| DNeasy® | 100 | 20.2 ± 0.2 ab | 100 | 18.1 ± 0.2 ab |
| qPCR Protocols * | DNA Extraction Methods | Specimen Preparation | |||
|---|---|---|---|---|---|
| Whole | Ground | Cut-Off Head | Punctured Abdomen | ||
| CaLsol | Chelex | 23.4 ± 0.8 b | 20.1 ± 0.5 a | 23.5 ± 1.1 b | 23.9 ± 0.7 b |
| HotSHOT | 18.9 ± 0.7 a | 18.1 ± 0.4 a | 18.6 ± 0.6 a | 20.1 ± 0.8 a | |
| PBS | 27.4 ± 1.4 a | 27.5 ± 1.2 a | 26.3 ± 0.7 a | 28.1 ± 1.1 a | |
| DNeasy® | 25.4 ± 1.1 a | 20.2 ± 0.2 a | 22.4 ± 0.7 a | 23.6 ± 1.0 a | |
| Lso | Chelex | 21.9 ± 0.7 b | 18.7 ± 0.3 a | 21.9 ± 0.9 b | 21.6 ± 0.6 b |
| HotSHOT | 18.0 ± 0.5 a | 16.6 ± 0.2 a | 17.8 ± 0.5 a | 18.4 ± 0.8 a | |
| PBS | 23.9 ± 1.5 a | 21.6 ± 0.7 a | 21.7 ± 0.8 a | 24.0 ± 1.2 a | |
| DNeasy® | 25.1 ± 0.7 c | 18.1 ± 0.1 a | 20.9 ± 0.9 ab | 21.5 ± 0.9 bc | |
| DNA Extraction Methods | qPCR Protocols * | |||||
|---|---|---|---|---|---|---|
| CaLsol | Lso | |||||
| PP | Cq (t = 0) | Cq (t = 130) | PP | Cq (t = 0) | Cq (t = 130) | |
| CTAB | 100 | 22.5 ± 1.7 | 28.2 ± 1.3 | 100 | 22.5 ± 1.3 | 27.4 ± 1.2 |
| Chelex | 100 | 21.0 ± 1.2 | 23.6 ± 0.6 | 100 | 19.4 ± 0.6 | 23.5 ± 0.6 |
| Squashed on membrane | 66.7 | 29.7 ± 1.3 | 32.4 ± 1.7 | 100 | 25.2 ± 1.5 | 31.2 ± 1.1 |
| HotSHOT | 100 | 18.0 ± 0.8 | 22.8 ± 0.4 | 83.3 | 17.4 ± 0.7 | 25.3 ± 1.5 |
| DNeasy® | 100 | 23.6 ± 2.4 | 25.4 ± 3.4 | 100 | 20.3 ± 1.9 | 24.3 ± 2.0 |
| DNA Extraction Methods | Time Consumed * | Cost Consumables (€/Sample) | Hazardous Reagent Residues * | Non-Hazardous Reagent Residues * |
|---|---|---|---|---|
| CTAB | 2 h 20 min ** | 0.33 | 4 mL buffer CTAB + 4 mL chloroform:isoamyl alcohol (24:1) | 1.2 mL isopropanol+ 10 mL 70% ethanol |
| Chelex | 30 min *** | 0.26 | - | - |
| TRIsureTM | 2 h ** | 1.16 | 10 mL TRIsureTM + 2 mL chloroform | 10 mL 0.1 M sodium citrate with 10% ethanol + 3.5 mL 100% ethanol + 15 mL 75% ethanol |
| Squashed on membrane | 15 min | 0.24 | - | - |
| HotSHOT | 30 min | 0.07 | - | - |
| PBS | 35 min | 0.07 | - | - |
| DNeasy® | 30 min *** | 2.56 | - | 0.2 mL proteinase K, 2 mL ethanol 100% and buffers: 1.8 mL ATL, 2 mL AL, 5 mL AW1 and 5 mL AW2. |
| Assay No. | Description | Sample Analyzed | PP ** | Cq, Mean ± SE, n = 10 *** |
|---|---|---|---|---|
| 1 | 10 specimens CaLsol- (5♀ and 5♂) (A) | H2O | 0% | nd |
| 10 specimens CaLsol+ (5♀ and 5♂) (B) | H2O | 100% | 30.4 ± 3.6 | |
| 10 mixtures of ethanol (A + B) | H2O | 0% | nd | |
| 10 specimens CaLsol- (5♀ and 5♂) (A) | PBS | 0% | nd | |
| 10 specimens CaLsol+ (5♀ and 5♂) (B) | PBS | 100% | 29.7 ± 0.6 | |
| 10 ethanol mixture (A + B) | PBS | 100% | 29.0 ± 0.6 | |
| 2 | 10 same-sex couples (5 ♀ CaLsol+ ♀ CaLsol- and 5 ♂ CaLsol+ ♂ CaLsol−) | H2O | 100% | 31.8 ± 1.0 |
| 10 same-sex couples (5 ♀ CaLsol+ ♀ CaLsol- and 5 ♂ CaLsol + ♂ CaLsol−) | PBS | 100% | 28.9 ± 0.4 | |
| 3 * | 10 ♂ specimens CaLsol- | Insect | 100% | 33.5 ± 0.4 |
| 10 ♀ specimens CaLsol+ | Insect | 100% | 17.2 ± 0.8 | |
| 10 ♂ specimens CaLsol- | Insect | 100% | 30.9 ± 2.3 | |
| 10 ♀ specimens CaLsol+ | Insect | 100% | 20.2 ± 2.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quintana, M.; de-León, L.; Cubero, J.; Siverio, F. Assessment of Psyllid Handling and DNA Extraction Methods in the Detection of ‘Candidatus Liberibacter Solanacearum’ by qPCR. Microorganisms 2022, 10, 1104. https://doi.org/10.3390/microorganisms10061104
Quintana M, de-León L, Cubero J, Siverio F. Assessment of Psyllid Handling and DNA Extraction Methods in the Detection of ‘Candidatus Liberibacter Solanacearum’ by qPCR. Microorganisms. 2022; 10(6):1104. https://doi.org/10.3390/microorganisms10061104
Chicago/Turabian StyleQuintana, María, Leandro de-León, Jaime Cubero, and Felipe Siverio. 2022. "Assessment of Psyllid Handling and DNA Extraction Methods in the Detection of ‘Candidatus Liberibacter Solanacearum’ by qPCR" Microorganisms 10, no. 6: 1104. https://doi.org/10.3390/microorganisms10061104