

Cyclophosphamide Induces Lipid and Metabolite Perturbation in Amniotic Fluid during Rat Embryonic Development
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Embryo-Fetal Toxicity
2.2.1. Animals
2.2.2. Treatment and Sample Collection
2.2.3. Oxidative Stress Assessment
2.2.4. Statistical Analysis
2.3. Untargeted Lipidomics and Metabolomic Analysis
2.3.1. Sample Preparation
2.3.2. Chromatographic Separation and Mass Spectrometer Settings
2.3.3. Data Processing and Statistical Analysis
3. Results
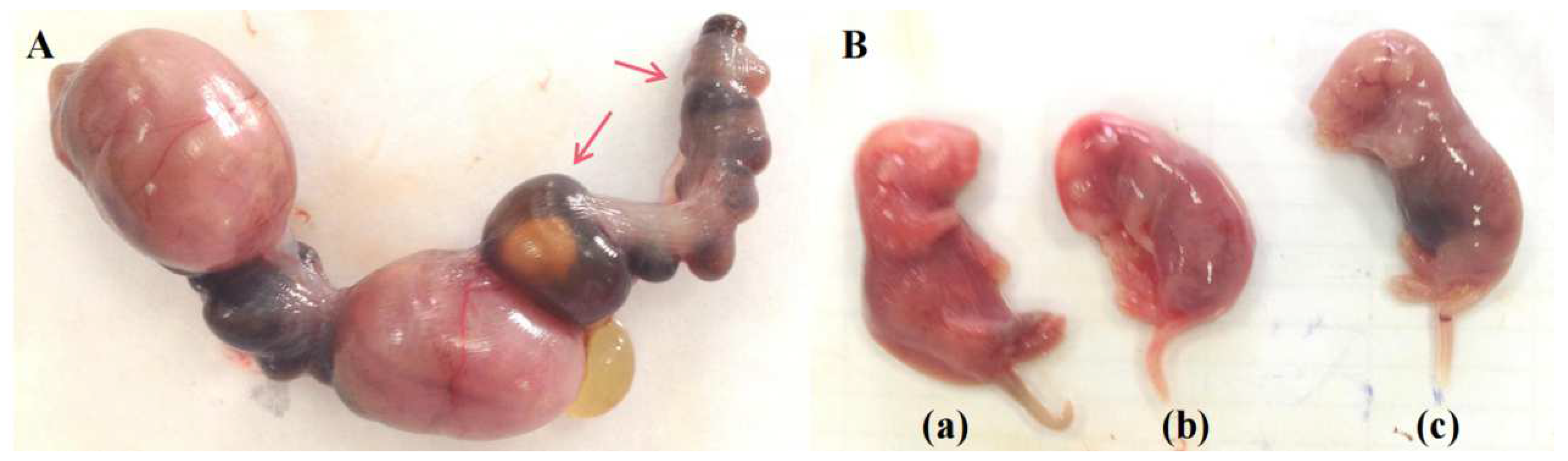
3.1. CP disturbs Embryonic Development
3.2. Lipidomic Profiling of Amniotic Fluid after CP Treatment
3.2.1. Lipid Components in the Amniotic Fluid
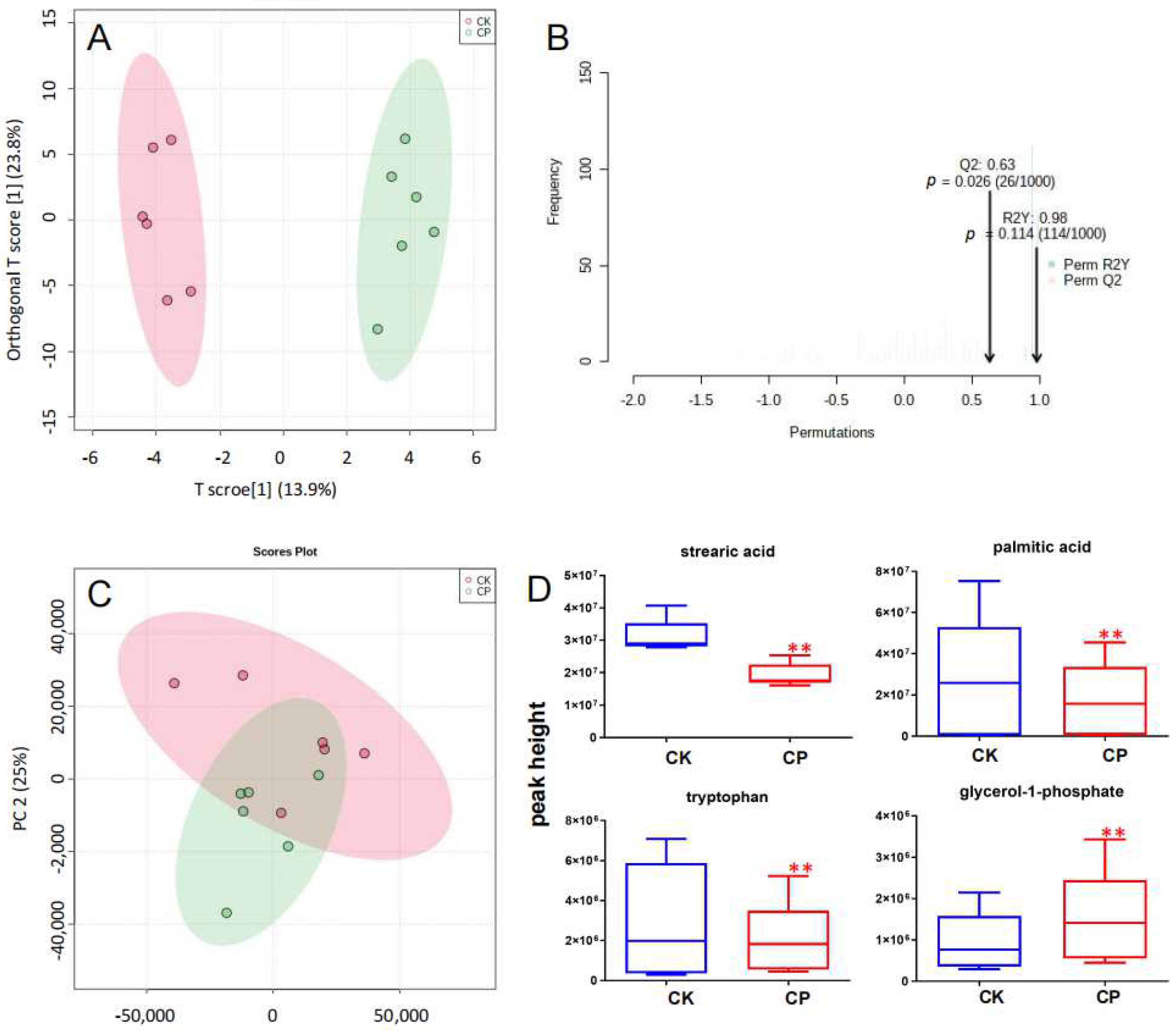
3.2.2. UHPLC-Q-Exactive Orbitrap MS Method Validation
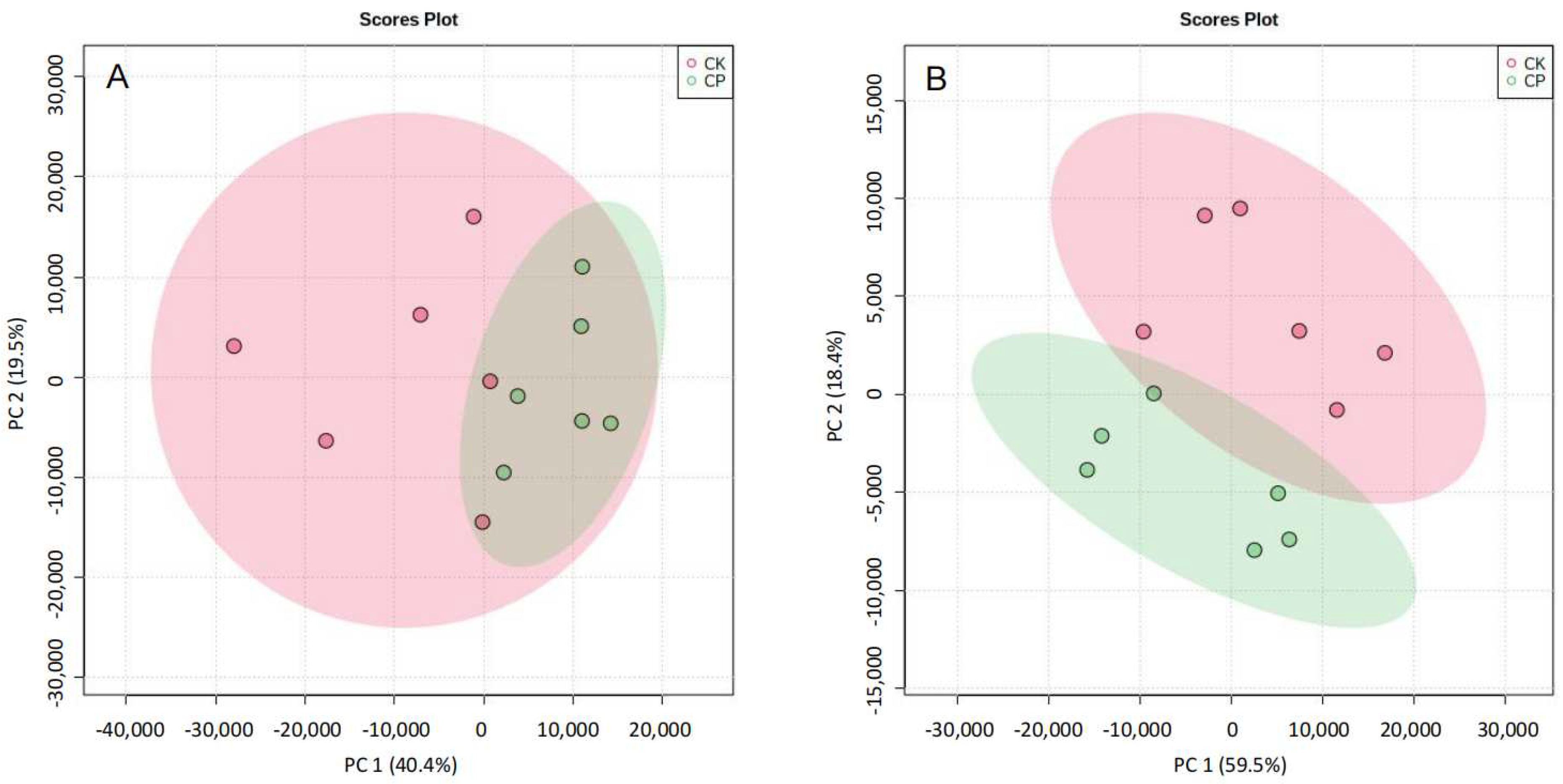
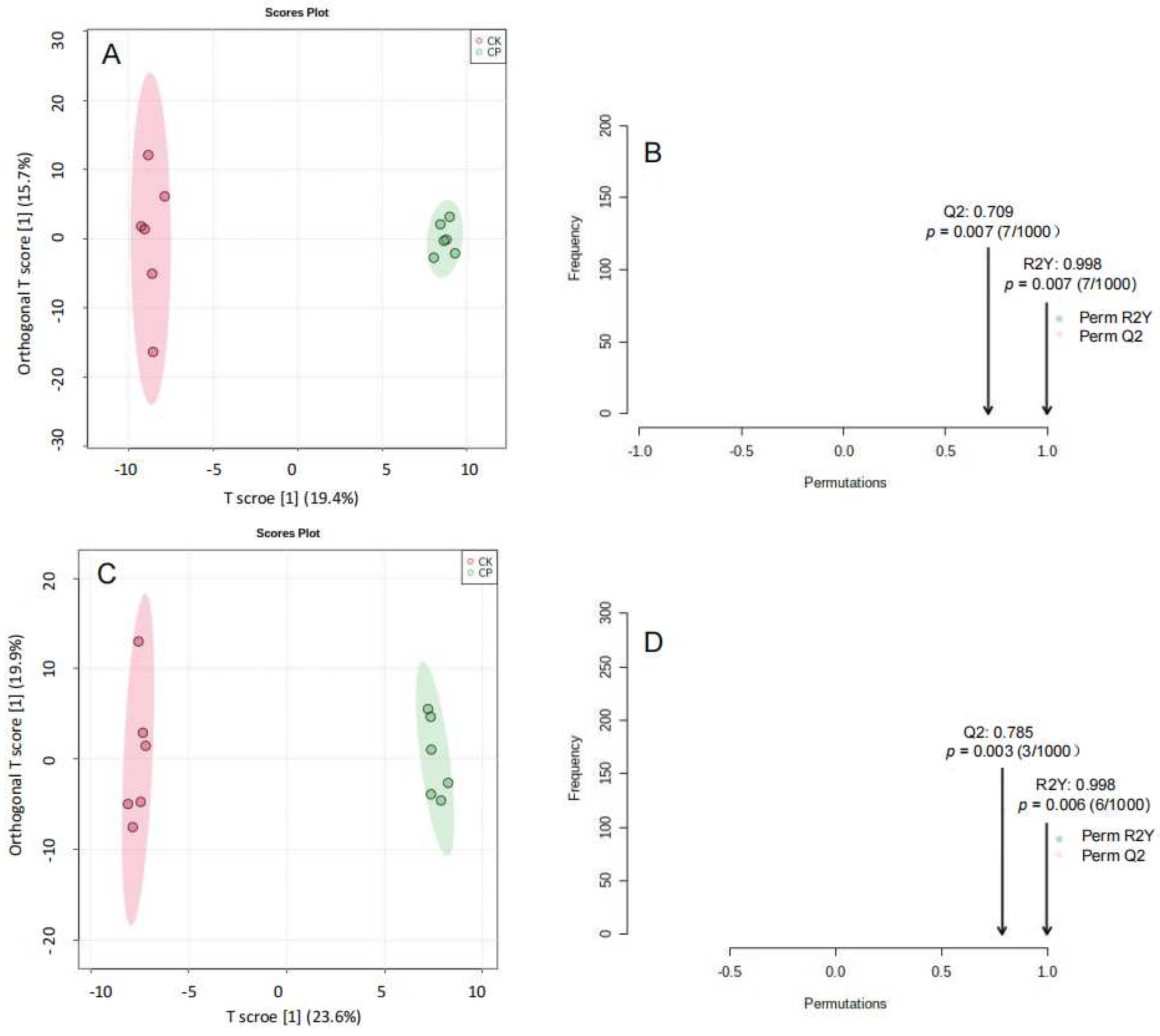
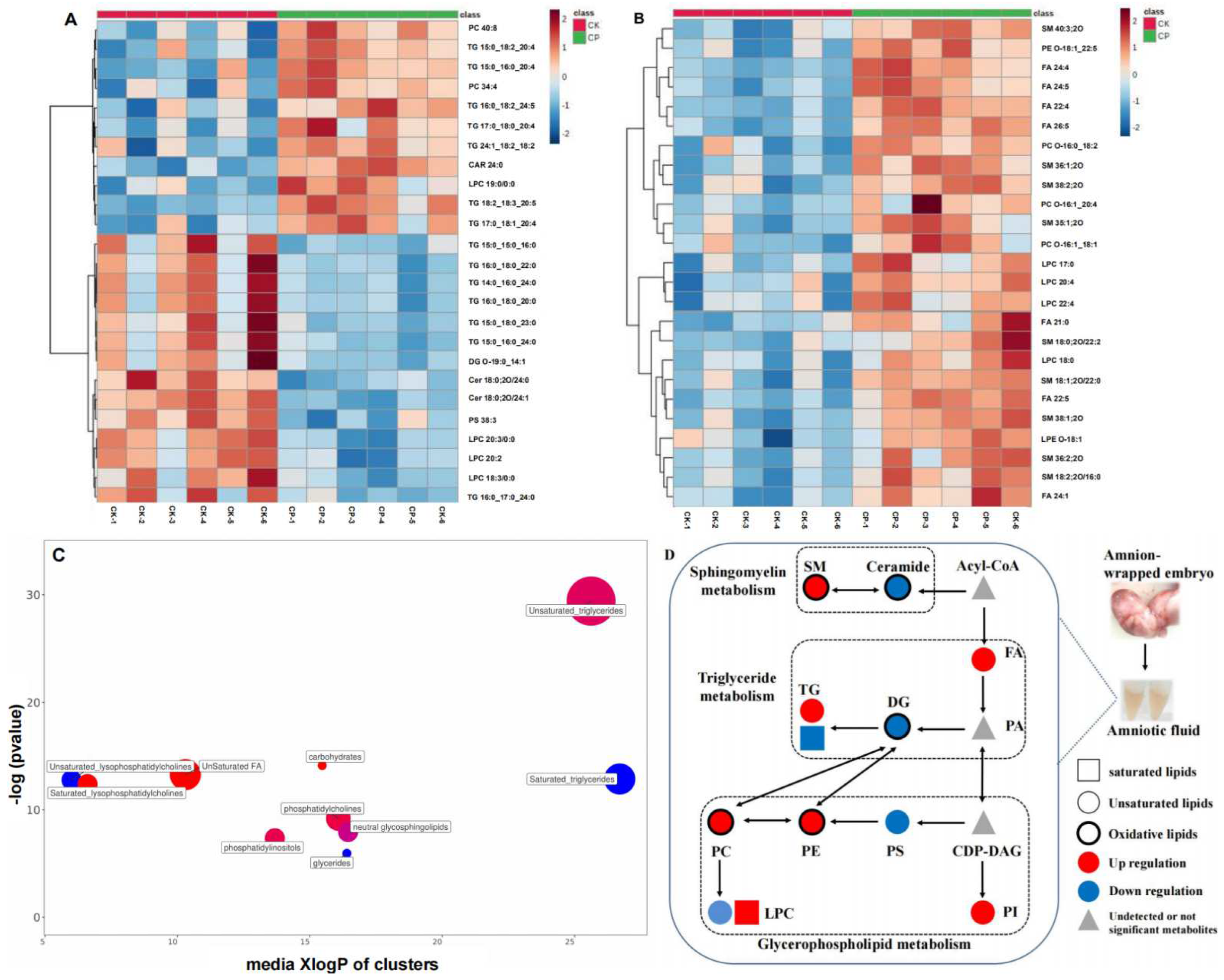
3.2.3. Non-Targeted Lipidomics Analysis
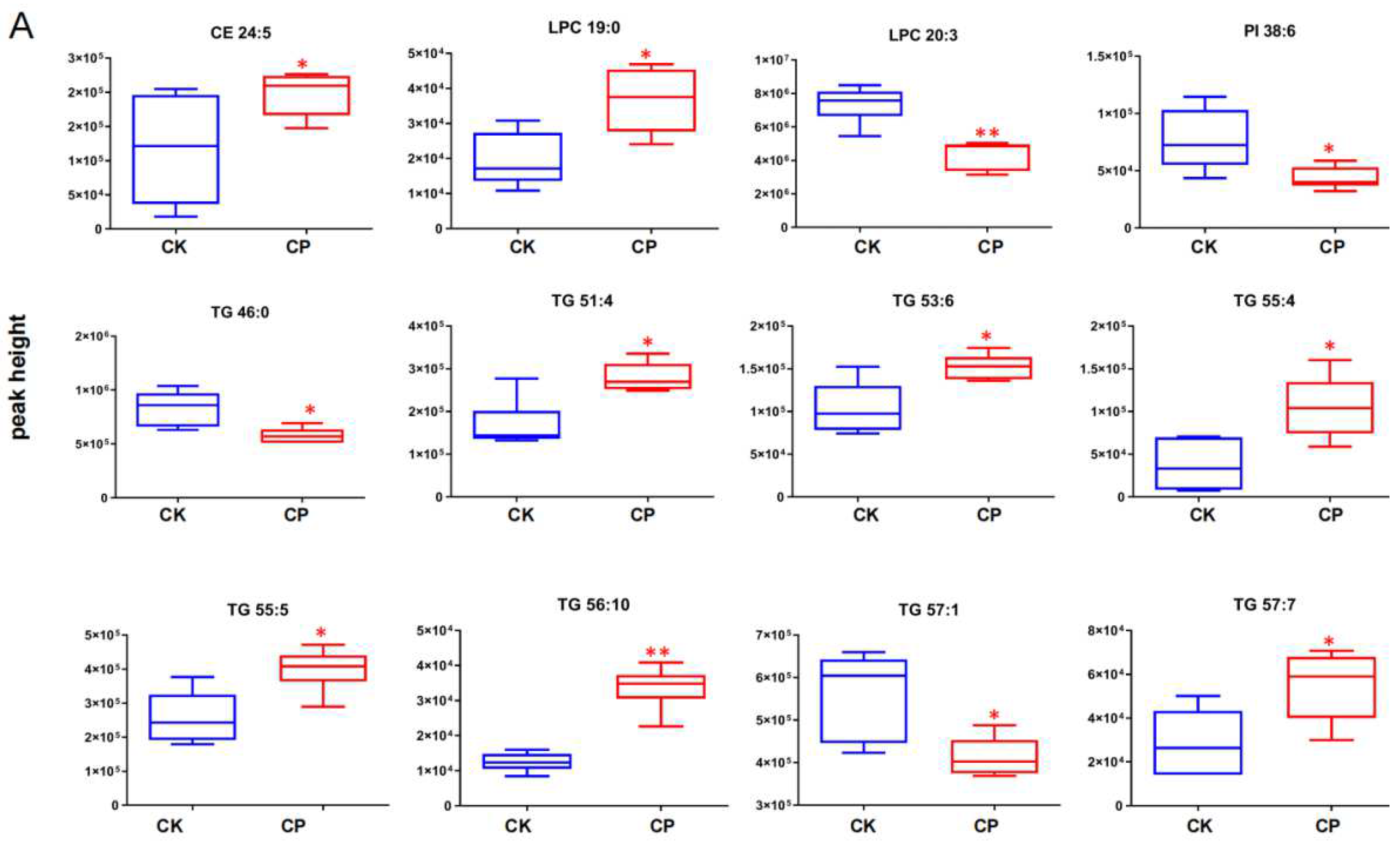
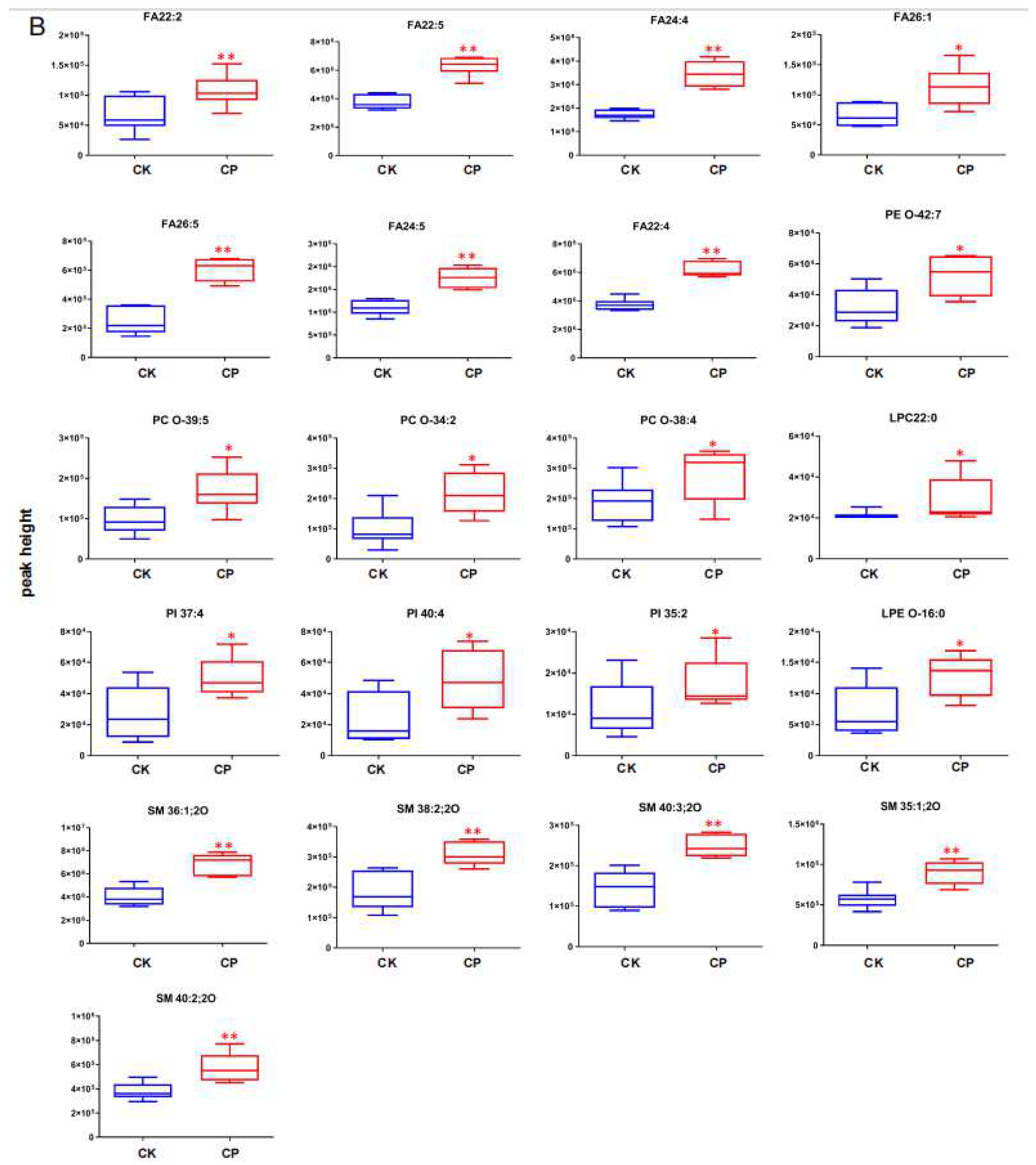
3.2.4. CP Induces Lipid Perturbation in the Amniotic Fluid
3.3. Metabolic Profiling of Amniotic Fluid
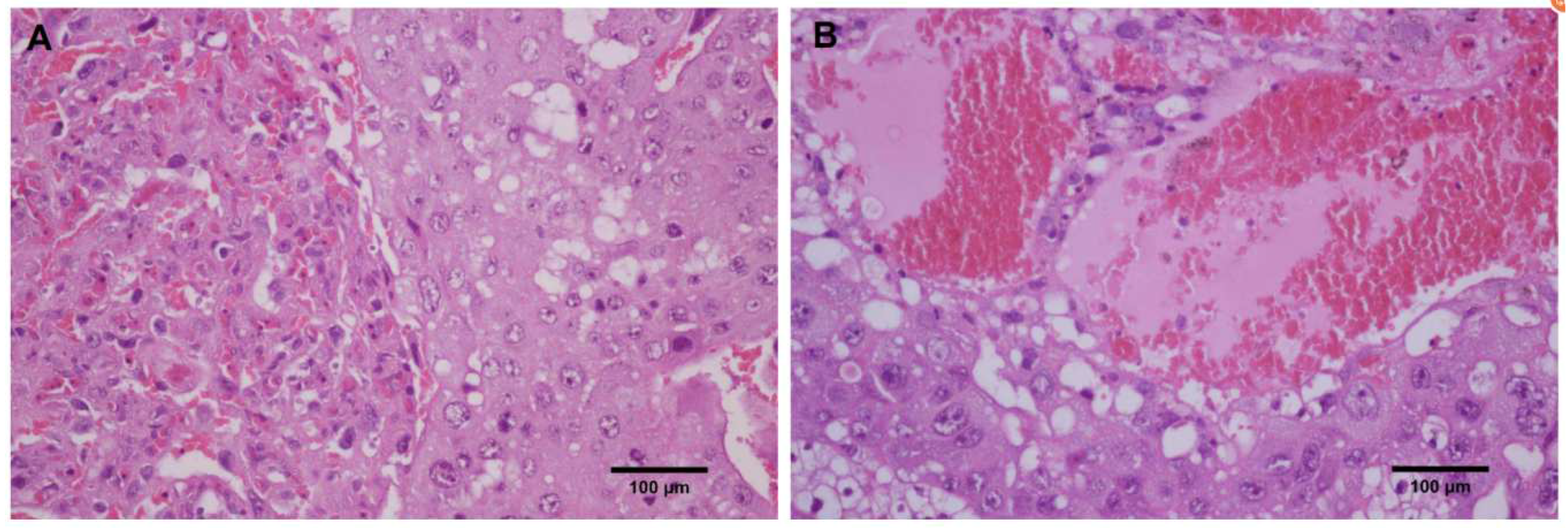
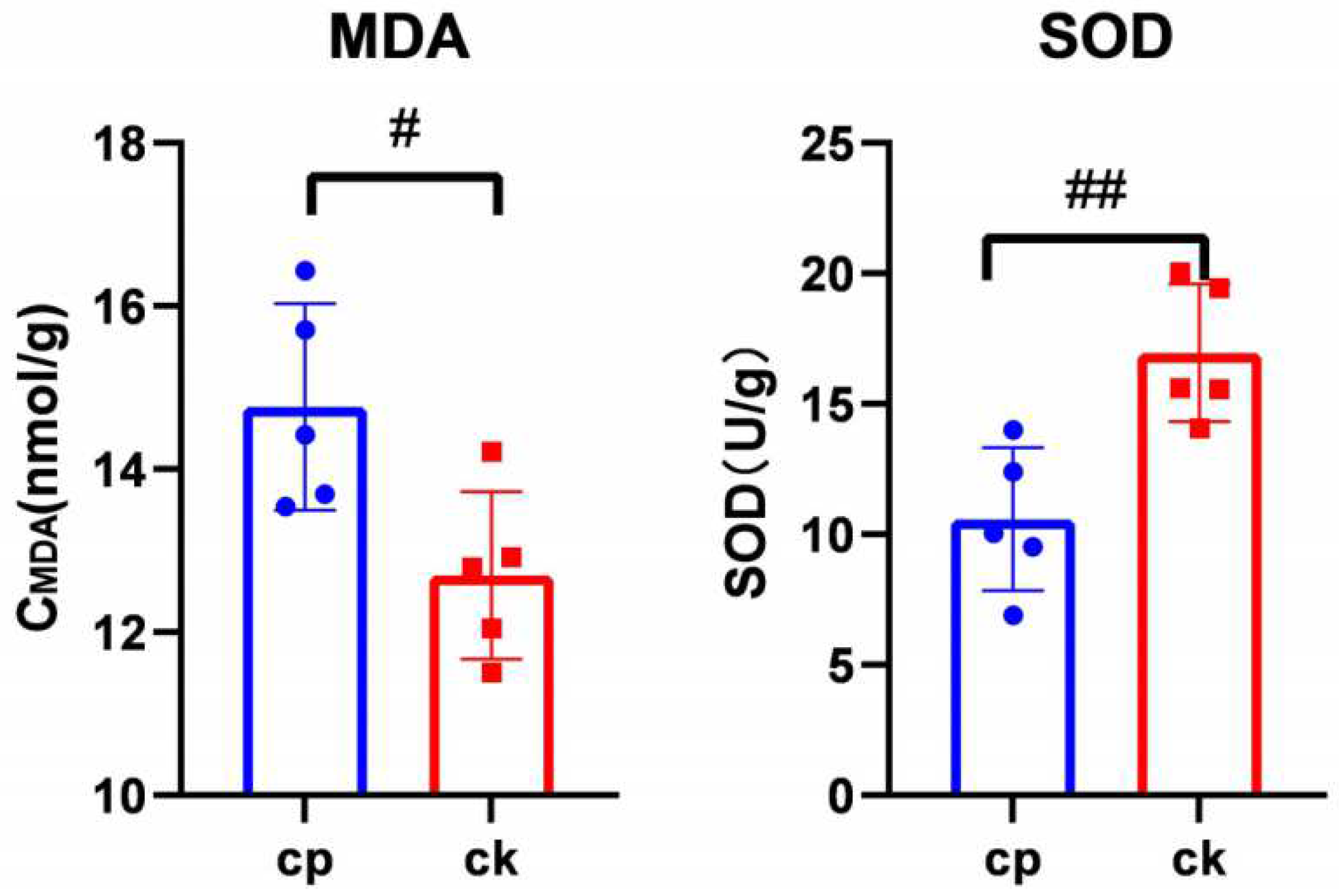
3.4. CP Triggers Placental Oxidative Stress
4. Discussion
4.1. FAs
4.2. SMs
4.3. PIs
4.4. TGs
4.5. SAs, PAs, and Tryptophan
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Acyl-CoA | acyl coenzyme A |
| AMN | adrenal medulla neuropathy |
| Arb | arbitrary unit |
| BSTFA | O-Bistrimethylsilyl) triflfluoroacetamide |
| CAR | cardiolipin |
| CCALD | childhood-onset cerebral adrenoleukodystrophy |
| CDP-DAG | cytidine diphosphate diacylglycerol |
| CE | cholesteryl ester |
| Cer | ceramide |
| CP | cyclophosphamide |
| DG | diacylglycerol |
| EFAs | essential fatty acids |
| EI | electron ionization |
| FA | fatty acid |
| GC-MS | gas chromatography–mass spectrometry |
| GD | gestation day |
| HMIT | H+-myo-inositol transporter |
| LC-MS | liquid chromatography–mass spectrometry |
| LCPUFAs | long-chain polyunsaturated fatty acids |
| LPL | lipoprotein lipase |
| lysoPC, LPC | lysophosphatidylcholine |
| lysoPE, LPE | lysophosphatidylethanolamine |
| MDA | malonaldehyde |
| MTBE | methyl tert-butyl ether |
| NTD | neural tube defects |
| PA | phosphatidic acid |
| PC | phosphatidylcholine |
| PE | phosphatidylethanolamine |
| PI | phosphatidylinositol |
| PIP | phosphatidylinositol 4-phosphate |
| PIP2 | 4, 5-diphosphate |
| PS | phosphatidylserine |
| PUFA | polyunsaturated fatty acid |
| QC | quality control |
| RSD | relative standard deviations |
| S1P | sphingosine-1-phosphate |
| SD | Sprague–Dawley |
| SD | standard deviation |
| SGA | small-for-gestational-age |
| SM | sphingomyelin |
| SMases | sphingomyelinases |
| SMIT | sodium–myo-inositol transporter |
| SOD | superoxide dismutase |
| Sph | sphingosine |
| TG | triglyceride |
| TIC | total ion current |
| VLCFAs | very-long-chain fatty acids |
| VLCUFAs | very-long-chain unsaturated fatty acids |
References
- Ghareeb, M.A.; Sobeh, M.; El-Maadawy, W.H.; Mohammed, H.S.; Khalil, H.; Botros, S.; Wink, M. Chemical Profiling of Polyphenolics in Eucalyptus globulus and Evaluation of Its Hepato-Renal Protective Potential Against Cyclophosphamide Induced Toxicity in Mice. Antioxidants 2019, 8, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakhem, G.A.; Motosko, C.C.; Mu, E.W.; Ho, R.S. Infertility and teratogenicity after paternal exposure to systemic dermatologic medications: A systematic review. J. Am. Acad. Dermatol. 2019, 80, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lee, I.C.; Lim, J.H.; Moon, C.; Bae, C.S.; Kim, S.H.; Shin, D.H.; Park, S.C.; Kim, H.C.; Kim, J.C. Protective effects of pine bark extract on developmental toxicity of cyclophosphamide in rats. Food Chem. Toxicol. 2012, 50, 109–115. [Google Scholar] [CrossRef]
- El-Dakdoky, M.H. Influence of mefloquine administration during early pregnancy on rat embryonic development. Toxicol. Mech. Methods 2015, 25, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.Y.; Zhang, K.; Xie, H.H.; Shan, J.J.; Xie, T. Effects of raw pinellia ternate and Ganjiang Renshen Banxia Pill on reproductive related toxicity of mice. China J. Tradit. Chin. Med. Pharm. 2017, 32, 3154–3157. [Google Scholar]
- Kim, S.H.; Lee, I.C.; Baek, H.S.; Moon, C.; Kim, S.H.; Yoo, J.C.; Shin, I.S.; Kim, J.C. Induction of cytochrome P450 3A1 expression by diallyl disulfide: Protective effects against cyclophosphamide-induced embryo-fetal developmental toxicity. Food Chem. Toxicol. 2014, 69, 312–319. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C. Systems biology: Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef]
- Shi, J.; Cao, B.; Wang, X.W.; Aa, J.Y.; Duan, J.A.; Zhu, X.X.; Wang, G.J.; Liu, C.X. Metabolomics and its application to the evaluation of the efficacy and toxicity of traditional Chinese herb medicines. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2016, 1026, 204–216. [Google Scholar] [CrossRef]
- Virgiliou, C.; Gika, H.G.; Witting, M.; Bletsou, A.A.; Athanasiadis, A.; Zafrakas, M.; Thomaidis, N.S.; Raikos, N.; Makrydimas, G.; Theodoridis, G.A. Amniotic Fluid and Maternal Serum Metabolic Signatures in the Second Trimester Associated with Preterm Delivery. J. Proteome Res. 2017, 16, 898–910. [Google Scholar] [CrossRef]
- Dasgupta, S.; Arya, S.; Choudhary, S.; Jain, S.K. Amniotic fluid: Source of trophic factors for the developing intestine. World J. Gastrointest. Pathophysiol. 2016, 7, 38–47. [Google Scholar] [CrossRef]
- Fanos, V.; Atzori, L.; Makarenko, K.; Melis, G.B.; Ferrazzi, E. Metabolomics application in maternal-fetal medicine. BioMed Res. Int. 2013, 2013, 720514. [Google Scholar] [CrossRef]
- Orczyk-Pawilowicz, M.; Jawien, E.; Deja, S.; Hirnle, L.; Zabek, A.; Mlynarz, P. Metabolomics of human amniotic fluid and maternal plasma during normal pregnancy. PLoS ONE 2016, 11, e0152740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, K.; Alexander, J.; Azuma, R.; Xiao, R.; Snyder, N.W.; Mesaros, C.A.; Blair, I.A.; Pinney, S.E. Gestational diabetes alters the metabolomic profile in 2nd trimester amniotic fluid in a sex-specific manner. Int. J. Mol. Sci. 2018, 19, 2696. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.H.; Yan, Y.E.; Liang, G.; Liu, Y.S.; Li, X.J.; Zhang, B.J.; Chen, L.B.; Yu, H.; He, X.H.; Wang, H. Maternal and fetal metabonomic alterations in prenatal nicotine exposure-induced rat intrauterine growth retardation. Mol. Cell. Endocrinol. 2014, 394, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Kang, A.; Peng, L.; Xie, T.; Ji, J.; Zhou, W.; Shan, J.; Di, L. Gas chromatography-mass spectrometry based plasma metabolomics of H1N1-induced inflammation in mice and intervention with Flos Lonicerae Japonica-Fructus Forsythiae herb pair. J. Chromatogr. B Analyt. Technol. Biomed Life Sci. 2018, 1092, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, K.; Singla, E.; Sahu, B.; Naura, A.S. PARP inhibitor, olaparib ameliorates acute lung and kidney injury upon intratracheal administration of LPS in mice. Mol. Cell. Biochem. 2015, 400, 153–162. [Google Scholar] [CrossRef]
- Chen, S.; Hoene, M.; Li, J.; Li, Y.; Zhao, X.; Häring, H.U.; Schleicher, E.D.; Weigert, C.; Xu, G.; Lehmann, R. Simultaneous extraction of metabolome and lipidome with methyl tert-butyl ether from a single small tissue sample for ultra-high performance liquid chromatography/mass spectrometry. J. Chromatogr. A 2013, 1298, 9–16. [Google Scholar] [CrossRef]
- Shan, J.; Qian, W.; Shen, C.; Lin, L.; Xie, T.; Peng, L.; Xu, J.; Yang, R.; Ji, J.; Zhao, X. High-resolution lipidomics reveals dysregulation of lipid metabolism in respiratory syncytial virus pneumonia mice. RSC Adv. 2018, 8, 29368–29377. [Google Scholar] [CrossRef] [Green Version]
- Barupal, D.K.; Fiehn, O. Chemical Similarity Enrichment Analysis (ChemRICH) as alternative to biochemical pathway mapping for metabolomic datasets. Sci. Rep. 2017, 7, 14567. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Zhang, Y.; Qian, W.; Peng, L.; Lin, L.; Xu, J.; Xie, T.; Ji, J.; Zhan, X.; Shan, J. Surfactant lipidomics of alveolar lavage fluid in mice based on altra-high-performance liquid chromatography coupled to hybrid quadrupole-exactive orbitrap mass spectrometry. Metabolites 2019, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Şengül, E.; Gelen, V.; Gedikli, S.; Özkanlar, S.; Gür, C.; Çelebi, F.; Çınar, A. The protective effect of quercetin on cyclophosphamide-Induced lung toxicity in rats. Biomed. Pharmacother. 2017, 92, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Pugh, D.; Farrah, T.E.; Gallacher, P.J.; Kluth, D.C.; Dhaun, N. Cyclophosphamide-Induced Lung Injury. Kidney Int. Rep. 2018, 4, 484–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tisoncik-Go, J.; Gasper, D.J.; Kyle, J.E.; Eisfeld, A.J.; Selinger, C.; Hatta, M.; Morrison, J.; Korth, M.J.; Zink, E.M.; Kim, Y.M.; et al. Integrated Omics Analysis of pathogenic host responses during pandemic H1N1 influenza virus infection: The crucial role of lipid metabolism. Cell Host Microbe 2016, 19, 254–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, M.M.; Karathia, H.; Pujolassos, M.; Troisi, J.; Valitutti, F.; Subramanian, P.; Camhi, S.; Kenyon, V.; Colucci, A.; Serena, G.; et al. Multi-omics analysis reveals the influence of genetic and environmental risk factors on developing gut microbiota in infants at risk of celiac disease. Microbiome 2020, 8, 130. [Google Scholar] [CrossRef]
- Nakamura, M.T.; Yudell, B.E.; Loor, J.J. Regulation of energy metabolism by long-chain fatty acids. Prog. Lipid Res. 2014, 53, 124–144. [Google Scholar] [CrossRef]
- Wan, J.Y.; Tu, X.H.; Wei, F.; Jiang, Z.B.; Xu, S.L.; Lv, X.; Dong, X.Y.; Xiang, J.Q.; Siew-y, Q.; Chen, H. Research progress of very long-chain fatty acids. Food Nutr. China 2019, 25, 5–11. [Google Scholar] [CrossRef]
- Glatz, J.F.; Luiken, J.J.; Bonen, A. Membrane fatty acid transporters as regulators of lipid metabolism: Implications for metabolic disease. Physiol. Rev. 2010, 90, 367–417. [Google Scholar] [CrossRef] [Green Version]
- Savary, S.; Trompier, D.; Andréoletti, P.; Le Borgne, F.; Demarquoy, J.; Lizard, G. Fatty acids—induced lipotoxicity and inflammation. Curr. Drug Metab. 2012, 13, 1358–1370. [Google Scholar] [CrossRef]
- Parisi, L.R.; Li, N.; Atilla-Gokcumen, G.E. Very long chain fatty acids are functionally involved in necroptosis. Cell Chem. Biol. 2017, 24, 1445–1454.e8. [Google Scholar] [CrossRef] [Green Version]
- Parisi, L.R.; Sowlati-Hashjin, S.; Berhane, I.A.; Galster, S.L.; Carter, K.A.; Lovell, J.F.; Chemler, S.R.; Karttunen, M.; Atilla-Gokcumen, G.E. Membrane disruption by very long chain fatty acids during necroptosis. ACS Chem. Biol. 2019, 14, 2286–2294. [Google Scholar] [CrossRef]
- Hama, K.; Fujiwara, Y.; Takashima, S.; Hayashi, Y.; Yamashita, A.; Shimozawa, N.; Yokoyama, K. Hexacosenoyl-CoA is the most abundant very long-chain acyl-CoA in ATP binding cassette transporter D1-deficient cells. J. Lipid Res. 2020, 61, 523–536. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, H.; Yoshida, T.; Sanaki, T.; Shigaki, S.; Morita, H.; Oyama, M.; Mitsui, M.; Tanaka, Y.; Nakano, T.; Mitsutake, S.; et al. Possible roles of long-chain sphingomyelines and sphingomyelin synthase 2 in mouse macrophage inflammatory response. Biochem. Biophys. Res. Commun. 2017, 482, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Okazaki, T. Ceramide/Sphingomyelin Rheostat regulated by sphingomyelin synthases and chronic diseases in murine models. J. Lipid Atheroscler. 2020, 9, 380–405. [Google Scholar] [CrossRef] [PubMed]
- Wegner, M.S.; Schiffmann, S.; Parnham, M.J.; Geisslinger, G.; Grösch, S. The enigma of ceramide synthase regulation in mammalian cells. Prog. Lipid Res. 2016, 63, 93–119. [Google Scholar] [CrossRef] [PubMed]
- Vanier, M.T. Niemann-Pick diseases. Handb. Clin. Neurol. 2013, 113, 1717–1721. [Google Scholar] [CrossRef]
- Astudillo, L.; Sabourdy, F.; Therville, N.; Bode, H.; Ségui, B.; Andrieu-Abadie, N.; Hornemann, T.; Levade, T. Human genetic disorders of sphingolipid biosynthesis. J. Inherit. Metab. Dis. 2015, 38, 65–76. [Google Scholar] [CrossRef]
- Spassieva, S.D.; Ji, X.; Liu, Y.; Gable, K.; Bielawski, J.; Dunn, T.M.; Bieberich, E.; Zhao, L. Ectopic expression of ceramide synthase 2 in neurons suppresses neurodegeneration induced by ceramide synthase 1 deficiency. Proc. Natl. Acad. Sci. USA 2016, 113, 5928–5933. [Google Scholar] [CrossRef] [Green Version]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Jennemann, R.; Rabionet, M.; Gorgas, K.; Epstein, S.; Dalpke, A.; Rothermel, U.; Bayerle, A.; van der Hoeven, F.; Imgrund, S.; Kirsch, J.; et al. Loss of ceramide synthase 3 causes lethal skin barrier disruption. Hum. Mol. Genet. 2012, 21, 586–608. [Google Scholar] [CrossRef] [Green Version]
- Carlomagno, G.; Nordio, M.; Chiu, T.T.; Unfer, V. Contribution of myo-inositol and melatonin to human reproduction. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 159, 267–272. [Google Scholar] [CrossRef]
- Bridges, D.; Saltiel, A.R. Phosphoinositides: Key modulators of energy metabolism. Biochim. Biophys. Acta 2015, 1851, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Kiani, A.K.; Paolacci, S.; Calogero, A.E.; Cannarella, R.; Di Renzo, G.C.; Gerli, S.; Della Morte, C.; Busetto, G.M.; De Berardinis, E.; Del Giudice, F.; et al. From Myo-inositol to D-chiro-inositol molecular pathways. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 2390–2402. [Google Scholar] [CrossRef] [PubMed]
- Jauniaux, E.; Hempstock, J.; Teng, C.; Battaglia, F.C.; Burton, G.J. Polyol concentrations in the fluid compartments of the human conceptus during the first trimester of pregnancy: Maintenance of redox potential in a low oxygen environment. J. Clin. Endocrinol. Metab. 2005, 90, 1171–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, S.W.; Copp, A.J.; Greene, N.; Glazier, J.D. Maternal inositol status and neural tube defects: A role for the human yolk sac in embryonic inositol delivery? Adv. Nutr. 2021, 12, 212–222. [Google Scholar] [CrossRef]
- Cárdenas, A.M.; Fernández-Olivares, P.; Díaz-Franulic, I.; González-Jamett, A.M.; Shimahara, T.; Segura-Aguilar, J.; Caviedes, R.; Caviedes, P. Knockdown of Myo-Inositol Transporter SMIT1 Normalizes Cholinergic and Glutamatergic Function in an Immortalized Cell Line Established from the Cerebral Cortex of a Trisomy 16 Fetal Mouse, an Animal Model of Human Trisomy 21 (Down Syndrome). Neurotox. Res. 2017, 32, 614–623. [Google Scholar] [CrossRef]
- Ghio, A.; Bertolotto, A.; Resi, V.; Volpe, L.; Di Cianni, G. Triglyceride metabolism in pregnancy. Adv. Clin. Chem. 2011, 55, 133–153. [Google Scholar] [CrossRef]
- Álvarez, D.; Muñoz, Y.; Ortiz, M.; Maliqueo, M.; Chouinard-Watkins, R.; Valenzuela, R. Impact of Maternal Obesity on the Metabolism and Bioavailability of Polyunsaturated Fatty Acids during Pregnancy and Breastfeeding. Nutrients 2020, 13, 19. [Google Scholar] [CrossRef]
- Wang, C.; Kong, L.; Yang, Y.; Wei, Y.; Zhu, W.; Su, R.; Lin, L.; Yang, H. Recommended reference values for serum lipids during early and middle pregnancy: A retrospective study from China. Lipids Health Dis. 2018, 17, 246. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Jia, Z.; Su, S.; Han, L.; Meng, L.; Tang, G.; Wang, J.; Zhang, C.; Xie, X.; Zhang, Y.; et al. Establishment of trimester-specific reference intervals of serum lipids and the associations with pregnancy complications and adverse perinatal outcomes: A population-based prospective study. Ann. Med. 2021, 53, 1632–1641. [Google Scholar] [CrossRef]
- Rodie, V.A.; Caslake, M.J.; Stewart, F.; Sattar, N.; Ramsay, J.E.; Greer, I.A.; Freeman, D.J. Fetal cord plasma lipoprotein status in uncomplicated human pregnancies and in pregnancies complicated by pre-eclampsia and intrauterine growth restriction. Atherosclerosis 2004, 176, 181–187. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.; Nie, H.; Li, G.; Su, J.; Cao, X.; Cao, Y.; Wei, F. Amniotic fluid metabolomic and lipidomic alterations associated with hemoglobin Bart’s diseases. Metabolomics 2021, 17, 82. [Google Scholar] [CrossRef] [PubMed]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical relevance of biomarkers of oxidative stress. Antioxid. Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Luo, Z.; He, Y.; Shan, J.; Guo, J.; Li, J. Application of untargeted lipidomics based on UHPLC-high resolution tandem MS analysis to profile the lipid metabolic disturbances in the heart of diabetic cardiomyopathy mice. J. Pharm. Biomed. Anal. 2020, 190, 113525. [Google Scholar] [CrossRef]
- Gonçalves, B.M.; Graceli, J.B.; da Rocha, P.B.; Tilli, H.P.; Vieira, E.M.; de Sibio, M.T.; Peghinelli, V.V.; Deprá, I.C.; Mathias, L.S.; Olímpio, R.; et al. Placental model as an important tool to study maternal-fetal interface. Reprod. Toxicol. 2022, 112, 7–13. [Google Scholar] [CrossRef]
- Xu, K.; Liu, G.; Fu, C. The Tryptophan Pathway Targeting Antioxidant Capacity in the Placenta. Oxidative Med. Cell. Longev. 2018, 2018, 1054797. [Google Scholar] [CrossRef] [PubMed]
- Li, L. Inffuence of the change of fatty acidsin yolk on embryo. China Poult. 2019, 20, 3–5. [Google Scholar] [CrossRef]
- Jiang, L.; Dang, W.; Zhou, J. Progress in stearic acid in rheumatic diseases. Chin. J. Rheumatol. 2021, 25, 762–765. [Google Scholar] [CrossRef]
- McKeegan, P.J.; Sturmey, R.G. The role of fatty acids in oocyte and early embryo development. Reprod. Ferti. Dev. 2011, 24, 59–67. [Google Scholar] [CrossRef]
- Aardema, H.; Vos, P.L.; Lolicato, F.; Roelen, B.A.; Knijn, H.M.; Vaandrager, A.B.; Helms, J.B.; Gadella, B.M. Oleic acid prevents detrimental effects of saturated fatty acids on bovine oocyte developmental competence. Biol. Reprod. 2011, 85, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Ma, W.; Chen, X.; Zeng, X. Tryptophan catabolism and its application in pig diets. Chin. J. Anim. Nutr. 2020, 32, 1019–1024. [Google Scholar] [CrossRef]
- Roth, W.; Zadeh, K.; Vekariya, R.; Ge, Y.; Mohamadzadeh, M. Tryptophan metabolism and gut-brain homeostasis. Int. J. Mol. Sci. 2021, 22, 2973. [Google Scholar] [CrossRef] [PubMed]
- Modoux, M.; Rolhion, N.; Mani, S.; Sokol, H. Tryptophan metabolism as a pharmacological target. Trends Pharmacol. Sci. 2021, 42, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Du, L.; Qi, R. Research progress of tryptophan and its metabolites in regulating muscle growth and development. Chin. J. Anim. Sci. 2022, 58, 21–25. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | CK | CP 15 mg·kg−1 |
|---|---|---|
| Pregnant rats (N) a | 7 | 9 |
| Maternal weight on GD17 (g; ± s) | 350.9 ± 25.4 | 319.3 ± 24.9 * |
| Fetal and placental weights (g; ± s) | 27.1 ± 11.3 | 20.1 ± 13.1 * |
| Implanted embryos (N) a | 87 | 77 |
| Viable embryos (N) a | 80 | 59 |
| Fetuses resorbed (N [%]) b | 6(6.9) | 7(9.1) |
| Late fetal death (N [%]) b | 1(1.1) | 11(14.3) * |
| Fetal weight (g; ± s) | 0.84 ± 0.04 | 0.55 ± 0.08 ** |
| Body length of fetuses (mm; ± s) | 20.33 ± 0.46 | 18.27 ± 1.06 ** |
| Tail length of fetuses (mm; ± s) | 6.68 ± 0.70 | 6.56 ± 0.56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Fang, H.; Chong, Y.; Lin, L.; Xie, T.; Ji, J.; Shen, C.; Shi, C.; Shan, J. Cyclophosphamide Induces Lipid and Metabolite Perturbation in Amniotic Fluid during Rat Embryonic Development. Metabolites 2022, 12, 1105. https://doi.org/10.3390/metabo12111105
Xu J, Fang H, Chong Y, Lin L, Xie T, Ji J, Shen C, Shi C, Shan J. Cyclophosphamide Induces Lipid and Metabolite Perturbation in Amniotic Fluid during Rat Embryonic Development. Metabolites. 2022; 12(11):1105. https://doi.org/10.3390/metabo12111105
Chicago/Turabian StyleXu, Jianya, Huafeng Fang, Ying Chong, Lili Lin, Tong Xie, Jianjian Ji, Cunsi Shen, Chen Shi, and Jinjun Shan. 2022. "Cyclophosphamide Induces Lipid and Metabolite Perturbation in Amniotic Fluid during Rat Embryonic Development" Metabolites 12, no. 11: 1105. https://doi.org/10.3390/metabo12111105