
The Role of the Immune Checkpoint Molecules PD-1/PD-L1 and TIM-3/Gal-9 in the Pathogenesis of Preeclampsia—A Narrative Review

 and
and 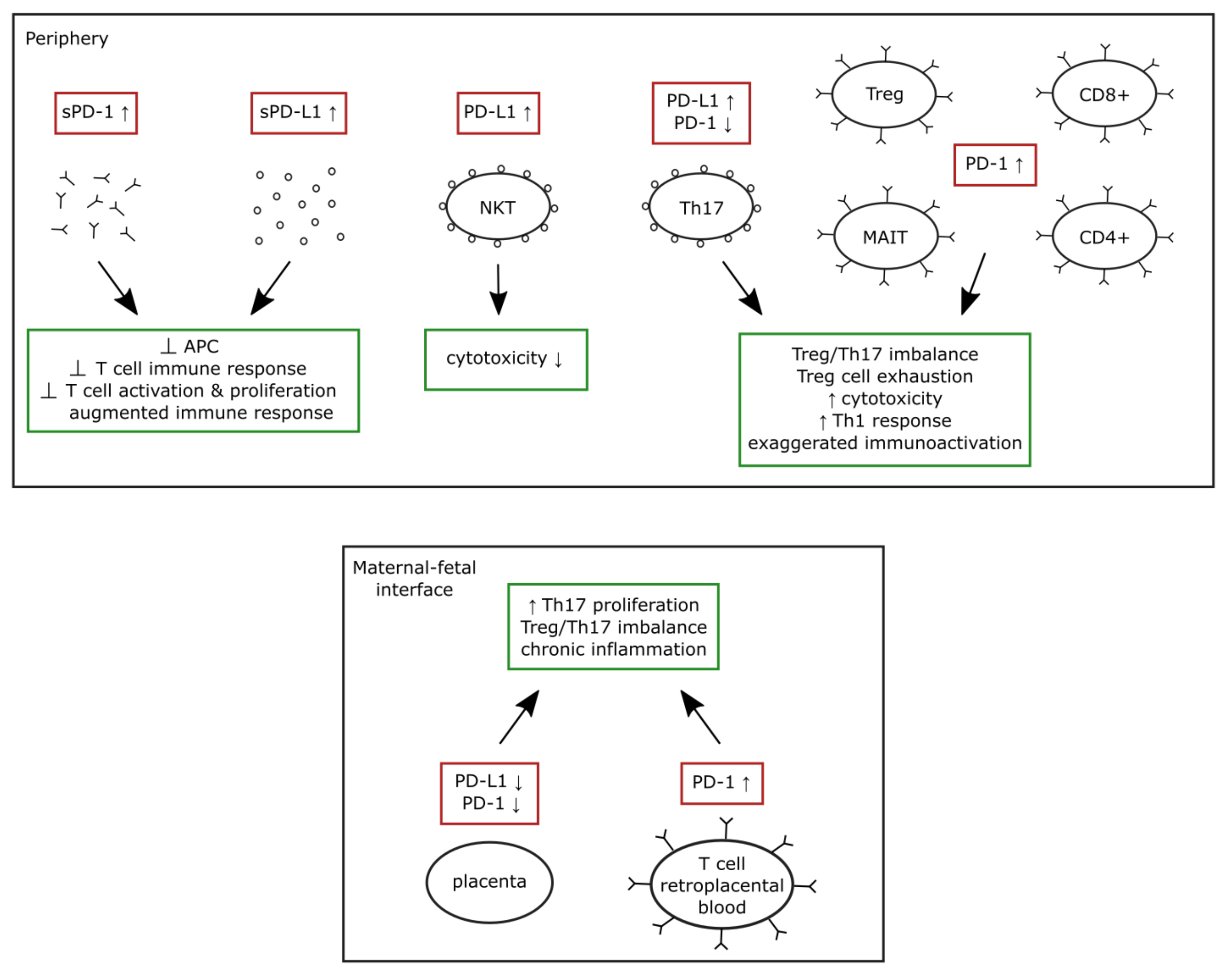{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. The PD-1/PD-L1 System in Normal Pregnancy and Preeclampsia
3.1. The PD-1/PD-L1 System in Normal Pregnancy
3.2. The PD-1/PD-L1 System in the Human Placenta
3.3. The PD-1/PD-L1 System in Preeclampsia
4. The TIM-3/Gal-9 System in Normal Pregnancy and Preeclampsia
4.1. The TIM-3/Gal-9 System in Normal Pregnancy
4.2. The TIM-3/Gal-9 System in the Human Placenta
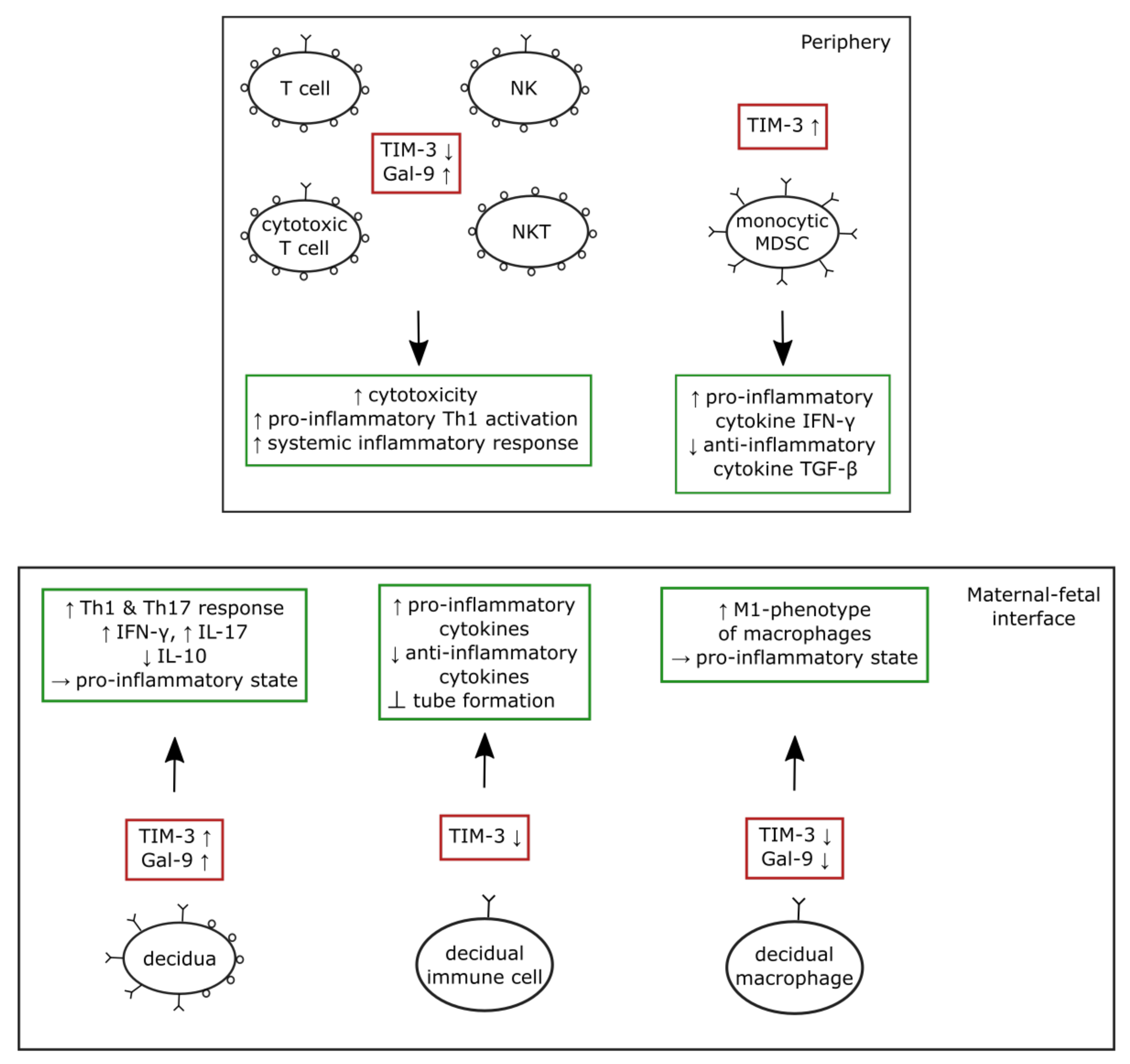
4.3. The TIM-3/Gal-9 System in Preeclampsia
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rana, S.; Lemoine, E.; Granger, J.; Karumanchi, S.A. Preeclampsia: Pathophysiology, challenges, and perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.C.; Chao, K.C.; Huang, S.J. Innate immunity, decidual cells, and preeclampsia. Reprod. Sci. 2013, 20, 339–353. [Google Scholar] [CrossRef]
- Mor, G.; Cardenas, I. The immune system in pregnancy: A unique complexity. Am. J. Reprod. Immunol. 2010, 63, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Meggyes, M.; Szanto, J.; Lajko, A.; Farkas, B.; Varnagy, A.; Tamas, P.; Hantosi, E.; Miko, E.; Szereday, L. The possible role of CD8+/Vα7.2+/CD161++ T (MAIT) and CD8+/Vα7.2+/CD161lo T (MAIT-like) cells in the pathogenesis of early-onset pre-eclampsia. Am. J. Reprod. Immunol. 2018, 79, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurieva, R.; Thomas, S.; Nguyen, T.; Martin-Orozco, N.; Wang, Y.; Kaja, M.-K.; Yu, X.-Z.; Dong, C. T-cell tolerance or function is determined by combinatorial costimulatory signals. EMBO J. 2006, 25, 2623–2633. [Google Scholar] [CrossRef] [Green Version]
- Petroff, M.G.; Chen, L.; Phillips, T.A.; Azzola, D.; Sedlmayr, P.; Hunt, J.S. B7 family molecules are favorably positioned at the human maternal-fetal interface. Biol. Reprod. 2003, 68, 1496–1504. [Google Scholar] [CrossRef] [Green Version]
- Veras, E.; Kurman, R.J.; Wang, T.-L.; Shih, I.-M. PD-L1 expression in human placentas and gestational trophoblastic diseases. Int. J. Gynecol. Pathol. 2017, 36, 146–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miko, E.; Meggyes, M.; Doba, K.; Barakonyi, A.; Szereday, L. Immune Checkpoint Molecules in Reproductive Immunology. Front. Immunol. 2019, 10, 846. [Google Scholar] [CrossRef]
- Chikuma, S. Basics of PD-1 in self-tolerance, infection, and cancer immunity. Int. J. Clin. Oncol. 2016, 21, 448–455. [Google Scholar] [CrossRef]
- Riley, J.L. PD-1 signaling in primary T cells. Immunol. Rev. 2009, 229, 114–125. [Google Scholar] [CrossRef]
- Francisco, L.M.; Sage, P.T.; Sharpe, A.H. The PD-1 pathway in tolerance and autoimmunity. Immunol. Rev. 2010, 236, 219–242. [Google Scholar] [CrossRef] [PubMed]
- Guleria, I.; Khosroshahi, A.; Ansari, M.J.; Habicht, A.; Azuma, M.; Yagita, H.; Noelle, R.J.; Coyle, A.; Mellor, A.L.; Khoury, S.; et al. A critical role for the programmed death ligand 1 in fetomaternal tolerance. J. Exp. Med. 2005, 202, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.C.; Latchman, Y.E.; Buhlmann, J.E.; Tomczak, M.F.; Horwitz, B.H.; Freeman, G.J.; Sharpe, A.H. Regulation of PD-1, PD-L1, and PD-L2 expression during normal and autoimmune responses. Eur. J. Immunol. 2003, 33, 2706–2716. [Google Scholar] [CrossRef] [PubMed]
- Habicht, A.; Dada, S.; Jurewicz, M.; Fife, B.; Yagita, H.; Azuma, M.; Sayegh, M.H.; Guleria, I. A Link between PDL1 and T Regulatory Cells in Fetomaternal Tolerance. J. Immunol. 2007, 179, 5211–5219. [Google Scholar] [CrossRef]
- D’Addio, F.; Riella, L.V.; Mfarrej, B.G.; Chabtini, L.; Adams, L.T.; Yeung, M.; Yagita, H.; Azuma, M.; Sayegh, M.H.; Guleria, I. The Link between the PDL1 Costimulatory Pathway and Th17 in Fetomaternal Tolerance. J. Immunol. 2011, 187, 4530–4541. [Google Scholar] [CrossRef] [Green Version]
- Taglauer, E.S.; Yankee, T.; Petroff, M.G. Maternal PD-1 regulates accumulation of fetal antigen-specific CD8+ T cells in pregnancy. J. Reprod. Immunol. 2009, 80, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enninga, E.A.L.; Harrington, S.M.; Creedon, D.J.; Ruano, R.; Markovic, S.N.; Dong, H.; Dronca, R.S. Immune checkpoint molecules soluble program death ligand 1 and galectin-9 are increased in pregnancy. Am. J. Reprod. Immunol. 2017, 79, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuyama, M.; Mezawa, H.; Kawai, T.; Urashima, M. Elevated Soluble PD-L1 in Pregnant Women’s Serum Suppresses the Immune Reaction. Front. Immunol. 2019, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Meggyes, M.; Miko, E.; Szigeti, B.; Farkas, N.; Szereday, L. The importance of the PD-1/PD-L1 pathway at the maternal-fetal interface. BMC Pregnancy Childbirth 2019, 19, 74. [Google Scholar] [CrossRef] [Green Version]
- Meggyes, M.; Nagy, D.U.; Szereday, L. Investigation of the PD-1 and PD-L1 Immune Checkpoint Molecules Throughout Healthy Human Pregnancy and in Nonpregnant Women. J. Clin. Med. 2020, 9, 2536. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Teng, X.; Fu, G.; Bao, L.; Tang, J.; Shi, H.; Lu, W.; Lu, Y. Analysis of PD-L1 expression in trophoblastic tissues and tumors. Hum. Pathol. 2018, 84, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Mincheva-Nilsson, L. Immunosuppressive Protein Signatures Carried by Syncytiotrophoblast-Derived Exosomes and Their Role in Human Pregnancy. Front. Immunol. 2021, 12, 1–10. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Aldo, P.; You, Y.; Ding, J.; Kaislasuo, J.; Petersen, J.F.; Lokkegaard, E.; Peng, G.; Paidas, M.J.; Simpson, S. Trophoblast-secreted soluble-PD-L1 modulates macrophage polarization and function. J. Leukoc. Biol. 2020, 108, 983–998. [Google Scholar] [CrossRef]
- van ’t Hof, L.J.; Dijkstra, K.L.; van der Keur, C.; Eikmans, M.; Baelde, H.J.; Bos, M.; van der Hoorn, M.L.P. Decreased expression of ligands of placental immune checkpoint inhibitors in uncomplicated and preeclamptic oocyte donation pregnancies. J. Reprod. Immunol. 2020, 142, 103194. [Google Scholar] [CrossRef]
- Gu, Y.; Morgan, J.; Lewis, D.F.; Cooper, D.B.; McCathran, C.E.; Wang, Y. Maternal soluble PD-1 levels are significantly increased in women with preeclampsia. Am. J. Reprod. Immunol. 2019, 83, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Lang, J. Soluble PD-1 and PD-L1: Predictive and prognostic significance in cancer. Oncotarget 2017, 8, 97671–97682. [Google Scholar] [CrossRef] [Green Version]
- Meggyes, M.; Miko, E.; Lajko, A.; Csiszar, B.; Sandor, B.; Matrai, P.; Tamas, P.; Szereday, L. Involvement of the PD-1/PD-L1 co-inhibitory pathway in the pathogenesis of the inflammatory stage of early-onset preeclampsia. Int. J. Mol. Sci. 2019, 20, 583. [Google Scholar] [CrossRef] [Green Version]
- Morita, K.; Tsuda, S.; Kobayashi, E.; Hamana, H.; Tsuda, K.; Shima, T.; Nakashima, A.; Ushijima, A.; Kishi, H.; Saito, S. Analysis of TCR Repertoire and PD-1 Expression in Decidual and Peripheral CD8+ T Cells Reveals Distinct Immune Mechanisms in Miscarriage and Preeclampsia. Front. Immunol. 2020, 11, 1082. [Google Scholar] [CrossRef]
- Tian, M.; Zhang, Y.; Liu, Z.; Sun, G.; Mor, G.; Liao, A. The PD-1/PD-L1 inhibitory pathway is altered in pre-eclampsia and regulates T cell responses in pre-eclamptic rats. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Daraei, N.; Ghafourian, M.; Ghadiri, A.; Amari, A.; Najafian, M.; Rokhafrooz, S. Evaluation of exhausted regulatory t cells in preeclampsia. Iran. J. Immunol. 2019, 16, 163–169. [Google Scholar] [PubMed]
- Zhang, Y.; Liu, Z.; Tian, M.; Hu, X.; Wang, L.; Ji, J.; Liao, A. The altered PD-1/PD-L1 pathway delivers the ‘one-two punch’ effects to promote the Treg/Th17 imbalance in pre-eclampsia. Cell Mol. Immunol. 2018, 15, 710–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zhang, X.; Du, N.; Sun, H.; Chen, L.; Bao, H.; Zhao, Q.; Qu, Q.; Ma, D.; Kwak-Kim, J.; et al. Immune checkpoint molecules on T cell subsets of pregnancies with preeclampsia and gestational diabetes mellitus. J. Reprod. Immunol. 2020, 142, 103208. [Google Scholar] [CrossRef]
- Pianta, S.; Magatti, M.; Vertua, E.; Signoroni, P.B.; Muradore, I.; Nuzzo, A.M.; Rolfo, A.; Silini, A.R.; Quaglia, F.; Todros, T.; et al. Amniotic mesenchymal cells from pre-eclamptic placentae maintain immunomodulatory features as healthy controls. J. Cell. Mol. Med. 2015, 20, 157–169. [Google Scholar] [CrossRef]
- Kandel, S.; Adhikary, P.; Li, G.; Cheng, K. The TIM3/Gal9 signaling pathway: An emerging target for cancer immunotherapy. Cancer Lett. 2021, 510, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Lei, Z.; Liu, Y.; Li, B.; Zhang, L.; Fang, H.; Song, C.; Wang, X.; Zhang, G.-M.; Feng, Z.-H.; et al. Human Pregnancy Up-Regulates Tim-3 in Innate Immune Cells for Systemic Immunity. J. Immunol. 2009, 182, 6618–6624. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, J.; Zhang, D.; Hong, X.; Tao, Y.; Wang, S.; Xu, Y.; Piao, H.; Yin, W.; Yu, M.; et al. Tim-3 signaling in peripheral NK cells promotes maternal-fetal immune tolerance and alleviates pregnancy loss. Sci. Signal. 2017, 10, eaah4323. [Google Scholar] [CrossRef] [Green Version]
- Meggyes, M.; Miko, E.; Polgar, B.; Bogar, B.; Farkas, B.; Illes, Z.; Szereday, L. Peripheral blood TIM-3 Positive NK and CD8+ T cells throughout pregnancy: TIM-3/Galectin-9 interaction and its possible role during pregnancy. PLoS ONE 2014, 9, e92371. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Chen, C.; Li, M.; Qian, J.; Sun, F.; Li, Y.; Yu, M.; Wang, M.; Zang, X.; Zhu, R.; et al. Blockade of CTLA-4 and Tim-3 pathways induces fetal loss with altered cytokine profiles by decidual CD4+T cells. Cell Death Dis. 2019, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhu, X.; Xu, Y.; Zhang, D.; Li, Y.; Tao, Y.; Piao, H.; Li, D.; Du, M. Programmed cell death-1 (PD-1) and T-cell immunoglobulin mucin-3 (Tim-3) regulate CD4+T cells to induce Type 2 helper T cell (Th2) bias at the maternal–fetal interface. Hum. Reprod. 2016, 31, 700–711. [Google Scholar] [CrossRef]
- Wang, S.-C.; Li, Y.-H.; Piao, H.-L.; Hong, X.-W.; Zhang, D.; Xu, Y.-Y.; Tao, Y.; Wang, Y.; Yuan, M.-M.; Li, D.-J.; et al. PD-1 and Tim-3 pathways are associated with regulatory CD8+ T-cell function in decidua and maintenance of normal pregnancy. Cell Death Dis. 2015, 6, e1738. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Sun, F.; Li, M.; Qian, J.; Chen, C.; Wang, M.; Zang, X.; Li, D.; Yu, M.; Du, M. The appropriate frequency and function of decidual Tim-3+CTLA-4+CD8+ T cells are important in maintaining normal pregnancy. Cell Death Dis. 2019, 10, 407. [Google Scholar] [CrossRef] [PubMed]
- Chabtini, L.; Mfarrej, B.; Mounayar, M.; Zhu, B.; Batal, I.; Dakle, P.J.; Smith, B.D.; Boenisch, O.; Najafian, N.; Akiba, H.; et al. TIM-3 regulates innate immune cells to induce fetomaternal tolerance. Bone 2014, 23, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sisti, G.; Minis, E.; Di Tommaso, M.; Malosso, E.R.M.; Parenti, A.; Paccosi, S.; Romero, M.I.P.; Witkin, S.S. T-cell immunoglobulin mucin-3 and galectin-9 levels in peripheral blood mononuclear cells predict fetal acidemia in twin pregnancies. J. Matern. Neonatal. Med. 2020, 33, 1683–1687. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Zhou, W.H.; Tao, Y.; Wang, S.C.; Jiang, Y.L.; Zhang, D.; Piao, H.-L.; Fu, Q.; Li, D.-J.; Du, M.-R. The Galectin-9/Tim-3 pathway is involved in the regulation of NK cell function at the maternal-fetal interface in early pregnancy. Cell Mol. Immunol. 2016, 13, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Yang, M.; Ban, Y.; Gao, W.; Song, B.; Wang, Y.; Zhang, Y.; Shao, Q.; Kong, B.; Qu, X. Tim-3 Is Upregulated in NK Cells during Early Pregnancy and Inhibits NK Cytotoxicity toward Trophoblast in Galectin-9 Dependent Pathway. PLoS ONE 2016, 11, e147186. [Google Scholar] [CrossRef]
- Wang, S.; Cao, C.; Piao, H.; Li, Y.; Tao, Y.; Zhang, X.; Zhang, D.; Sun, C.; Zhu, R.; Wang, Y.; et al. Tim-3 protects decidual stromal cells from toll-like receptor-mediated apoptosis and inflammatory reactions and promotes Th2 bias at the maternal-fetal interface. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Zhu, Q.; Wang, Y.; Wang, L.; Li, Z.; Mor, G.; Liao, A. Newly characterized decidual Tim-3+Treg cells are abundant during early pregnancy and driven by IL-27 coordinately with Gal-9 from trophoblasts. Hum. Reprod. 2020, 35, 2454–2466. [Google Scholar] [CrossRef] [PubMed]
- Hutter, S.; Knabl, J.; Andergassen, U.; Mayr, D.; Hofmann, S.; Kuhn, C.; Mahner, S.; Arck, P.; Jeschke, U. Fetal gender specific expression of tandem-repeat galectins in placental tissue from normally progressed human pregnancies and intrauterine growth restriction (IUGR). Placenta 2015, 36, 1352–1361. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Sun, F.; Qian, J.; Chen, L.; Li, D.; Wang, S.; Du, M. Tim-3/CTLA-4 pathways regulate decidual immune cells-extravillous trophoblasts interaction by IL-4 and IL-10. FASEB J. 2021, 35, e21754. [Google Scholar] [CrossRef] [PubMed]
- Miko, E.; Meggyes, M.; Bogar, B.; Schmitz, N.; Barakonyi, A.; Varnagy, A.; Farkas, B.; Tamas, P.; Bodis, J.; Szekeres-Bartho, J.; et al. Involvement of Galectin-9/TIM-3 Pathway in the Systemic Inflammatory Response in Early-Onset Preeclampsia. PLoS ONE 2013, 8, e71811. [Google Scholar] [CrossRef]
- Hao, H.; He, M.; Li, J.; Zhou, Y.; Dang, J.; Li, F.; Yang, M.; Deng, D. Upregulation of the Tim-3/Gal-9 pathway and correlation with the development of preeclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 194, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, C.; Sun, F.; Li, M.; Du, M.; Li, X.; Zhang, Y. Involvement of the Tim-3 Pathway in the Pathogenesis of Pre-Eclampsia. Reprod. Sci. 2021, 28, 3331–3340. [Google Scholar] [CrossRef]
- Dong, S.; Shah, N.K.; He, J.; Han, S.; Xie, M.; Wang, Y.; Cheng, T.; Liu, Z.; Shu, C. The abnormal expression of Tim-3 is involved in the regulation of myeloid-derived suppressor cells and its correlation with preeclampsia. Placenta 2021, 114, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-H.; Wang, L.-L.; Liu, H.; Muyayalo, K.P.; Huang, X.-B.; Mor, G.; Liao, A.-H. Galectin-9 Alleviates LPS-Induced Preeclampsia-Like Impairment in Rats via Switching Decidual Macrophage Polarization to M2 Subtype. Front. Immunol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mittelberger, J.; Seefried, M.; Franitza, M.; Garrido, F.; Ditsch, N.; Jeschke, U.; Dannecker, C. The Role of the Immune Checkpoint Molecules PD-1/PD-L1 and TIM-3/Gal-9 in the Pathogenesis of Preeclampsia—A Narrative Review. Medicina 2022, 58, 157. https://doi.org/10.3390/medicina58020157
Mittelberger J, Seefried M, Franitza M, Garrido F, Ditsch N, Jeschke U, Dannecker C. The Role of the Immune Checkpoint Molecules PD-1/PD-L1 and TIM-3/Gal-9 in the Pathogenesis of Preeclampsia—A Narrative Review. Medicina. 2022; 58(2):157. https://doi.org/10.3390/medicina58020157
Chicago/Turabian StyleMittelberger, Johanna, Marina Seefried, Manuela Franitza, Fabian Garrido, Nina Ditsch, Udo Jeschke, and Christian Dannecker. 2022. "The Role of the Immune Checkpoint Molecules PD-1/PD-L1 and TIM-3/Gal-9 in the Pathogenesis of Preeclampsia—A Narrative Review" Medicina 58, no. 2: 157. https://doi.org/10.3390/medicina58020157