
Biosynthesis of Long-Chain Polyunsaturated Fatty Acids in Marine Gammarids: Molecular Cloning and Functional Characterisation of Three Fatty Acyl Elongases

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
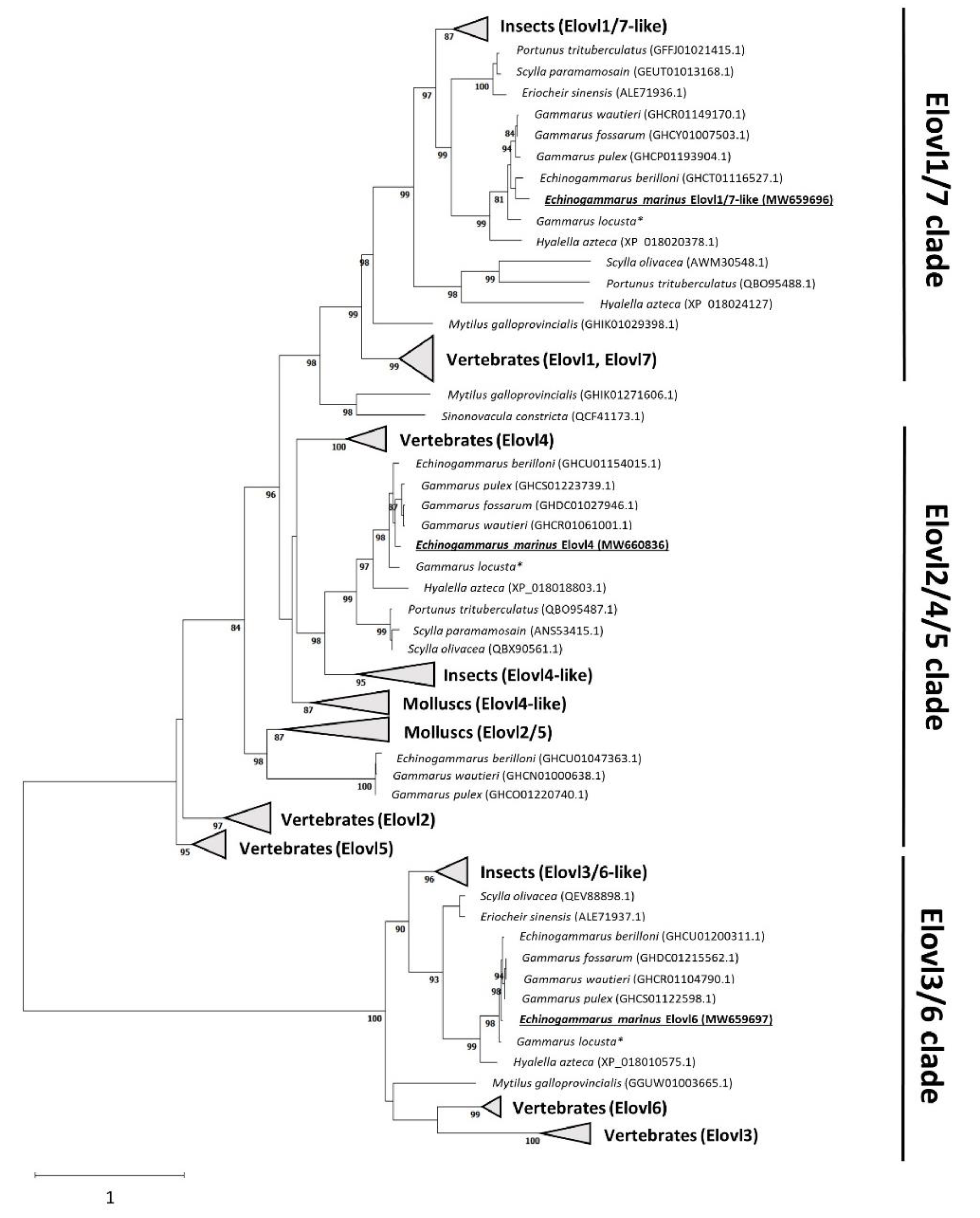
2.1. Phylogeny of the E. marinus Elovl
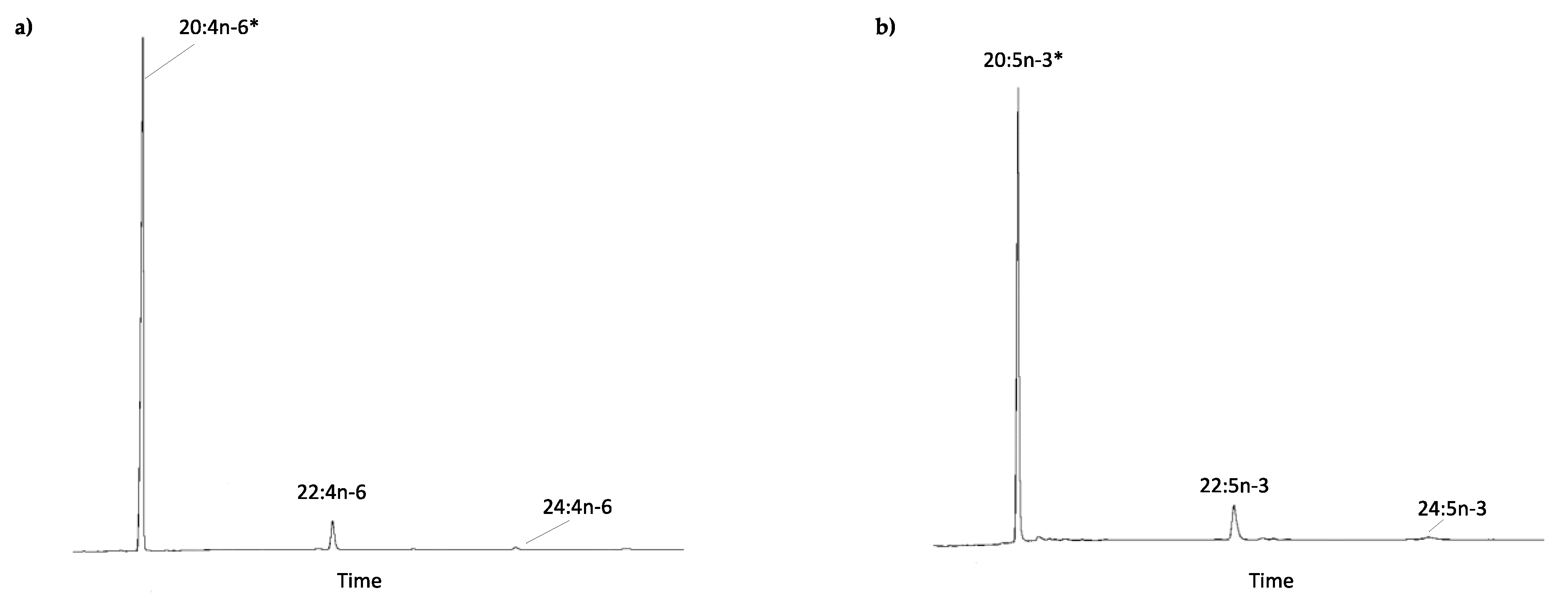
2.2. Roles of the E. marinus Elovl4, Elovl6 and Elovl1/7-like in LC-PUFA Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Molecular Cloning of Full-Length cDNAs of Three Elovls from E. marinus
4.2. Sequence and Phylogenetic Analysis of the E. marinus Elongases
4.3. Functional Characterisation of the E. marinus Elovl by Heterologous Expression in Yeast
4.4. Fatty Acid Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Guillou, H.; Zadravec, D.; Martin, P.G.P.; Jacobsson, A. The key roles of elongases and desaturases in mammalian fatty acid metabolism: Insights from transgenic mice. Prog. Lipid Res. 2010, 49, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef]
- Nichols, D.S. Prokaryotes and the input of polyunsaturated fatty acids to the marine food web. FEMS Microbiol. Lett. 2003, 219, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Khozin-Goldberg, I.; Iskandarov, U.; Cohen, Z. LC-PUFA from photosynthetic microalgae: Occurrence, biosynthesis, and prospects in biotechnology. Appl. Microbiol. Biotechnol. 2011, 91, 905–915. [Google Scholar] [CrossRef]
- Pereira, S.L.; Leonard, A.E.; Mukerji, P. Recent advances in the study of fatty acid desaturases from animals and lower eukaryotes. Prostaglandins Leukot. Essent. Fat. Acids 2003, 68, 97–106. [Google Scholar] [CrossRef]
- Monroig, Ó.; Kabeya, N. Desaturases and elongases involved in polyunsaturated fatty acid biosynthesis in aquatic invertebrates: A comprehensive review. Fish. Sci. 2018, 84, 911–928. [Google Scholar] [CrossRef] [Green Version]
- Kabeya, N.; Fonseca, M.M.; Ferrier, D.E.K.; Navarro, J.C.; Bay, L.K.; Francis, D.S.; Tocher, D.R.; Castro, L.F.C.; Monroig, Ó. Genes for de novo biosynthesis of omega-3 polyunsaturated fatty acids are widespread in animals. Sci. Adv. 2018, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Gladyshev, M.I.; Sushchik, N.N.; Makhutova, O.N. Production of EPA and DHA in aquatic ecosystems and their transfer to the land. Prostaglandins Lipid Mediat. 2013, 107, 117–126. [Google Scholar] [CrossRef]
- Tocher, D.R. Omega-3 long-chain polyunsaturated fatty acids and aquaculture in perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- Shepherd, C.J.; Monroig, O.; Tocher, D.R. Future availability of raw materials for salmon feeds and supply chain implications: The case of Scottish farmed salmon. Aquaculture 2017, 467, 49–62. [Google Scholar] [CrossRef]
- Galkanda-Arachchige, H.S.C.; Wilson, A.E.; Davis, D.A. Success of fishmeal replacement through poultry by-product meal in aquaculture feed formulations: A meta-analysis. Rev. Aquac. 2020, 12, 1624–1636. [Google Scholar] [CrossRef]
- Jannathulla, R.; Rajaram, V.; Kalanjiam, R.; Ambasankar, K.; Muralidhar, M.; Dayal, J.S. Fishmeal availability in the scenarios of climate change: Inevitability of fishmeal replacement in aquafeeds and approaches for the utilization of plant protein sources. Aquac. Res. 2019, 50, 3493–3506. [Google Scholar] [CrossRef]
- Turchini, G.M.; Torstensen, B.E.; Ng, W.K. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Sprague, M.; Betancor, M.B.; Tocher, D.R. Microbial and genetically engineered oils as replacements for fish oil in aquaculture feeds. Biotechnol. Lett. 2017, 39, 1599–1609. [Google Scholar] [CrossRef]
- Tocher, D.R.; Betancor, M.B.; Sprague, M.; Olsen, R.E.; Napier, J.A. Omega-3 long-chain polyunsaturated fatty acids, EPA and DHA: Bridging the gap between supply and demand. Nutrients 2019, 11, 89. [Google Scholar] [CrossRef] [Green Version]
- Napier, J.A.; Haslam, R.P.; Olsen, R.; Tocher, D.R.; Betancor, M.B. Agriculture can help aquaculture become greener. Nat. Food 2020, 1, 680–683. [Google Scholar] [CrossRef]
- Turchini, G.M.; Francis, D.S.; Senadheera, S.P.S.D.; Thanuthong, T.; De Silva, S.S. Fish oil replacement with different vegetable oils in Murray cod: Evidence of an “omega-3 sparing effect” by other dietary fatty acids. Aquaculture 2011, 315, 250–259. [Google Scholar] [CrossRef]
- Aas, T.S.; Ytrestøyl, T.; Åsgård, T. Utilization of feed resources in the production of Atlantic salmon (Salmo salar) in Norway: An update for 2016. Aquac. Rep. 2019, 15, 100216. [Google Scholar] [CrossRef]
- Naylor, R.L.; Hardy, R.W.; Bureau, D.P.; Chiu, A.; Elliott, M.; Farrell, A.P.; Forster, I.; Gatlin, D.M.; Goldburg, R.J.; Hua, K.; et al. Feeding aquaculture in an era of finite resources. Proc. Natl. Acad. Sci. USA 2009, 106, 15103–15110. [Google Scholar] [CrossRef] [Green Version]
- Dhont, J.; Dierckens, K.; Støttrup, J.; Van Stappen, G.; Wille, M.; Sorgeloos, P. Rotifers, Artemia and copepods as live feeds for fish larvae in aquaculture. In Advances in Aquaculture Hatchery Technology; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 157–202. ISBN 9780857091192. [Google Scholar]
- McKinnon, A.D.; Duggan, S.; Nichols, P.D.; Rimmer, M.A.; Semmens, G.; Robino, B. The potential of tropical paracalanid copepods as live feeds in aquaculture. Aquaculture 2003, 223, 89–106. [Google Scholar] [CrossRef]
- Odum, W.E.; Heald, E.J. Trophic analyses of an estuarine mangrove community. Bull. Mar. Sci. 1972, 22, 671–738. [Google Scholar]
- Dauby, P.; Scailteur, Y.; De Broyer, C. Trophic diversity within the eastern Weddell Sea amphipod community. Hydrobiologia 2001, 443, 69–86. [Google Scholar] [CrossRef]
- Baeza-Rojano, E.; Hachero-Cruzado, I.; Guerra-García, J.M. Nutritional analysis of freshwater and marine amphipods from the Strait of Gibraltar and potential aquaculture applications. J. Sea Res. 2014, 85, 29–36. [Google Scholar] [CrossRef]
- Jiménez-Prada, P.; Hachero-Cruzado, I.; Giráldez, I.; Fernández-Diaz, C.; Vilas, C.; Cañavate, J.P.; Guerra-García, J.M. Crustacean amphipods from marsh ponds: A nutritious feed resource with potential for application in Integrated Multi-Trophic Aquaculture. PeerJ 2018, 6, e4194. [Google Scholar] [CrossRef] [Green Version]
- Kolanowski, W.; Stolyhwo, A.; Grabowski, M. Fatty acid composition of selected freshwater gammarids (Amphipoda, Crustacea): A potentially innovative source of omega-3 LC PUFA. J. Am. Oil Chem. Soc. 2007, 84, 827–833. [Google Scholar] [CrossRef]
- Köprücü, K.; Özdemir, Y. Apparent digestibility of selected feed ingredients for Nile tilapia (Oreochromis niloticus). Aquaculture 2005, 250, 308–316. [Google Scholar] [CrossRef]
- Jiménez-Prada, P.; Hachero-Cruzado, I.; Guerra-García, J.M. Aquaculture waste as food for amphipods: The case of Gammarus insensibilis in marsh ponds from southern Spain. Aquac. Int. 2021, 29, 139–153. [Google Scholar] [CrossRef]
- Makhutova, O.N.; Shulepina, S.P.; Sharapova, T.A.; Kolmakova, A.A.; Glushchenko, L.A.; Kravchuk, E.S.; Gladyshev, M.I. Intraspecies variability of fatty acid content and composition of a cosmopolitan benthic invertebrate, Gammarus lacustris. Inl. Waters 2018, 8, 356–367. [Google Scholar] [CrossRef] [Green Version]
- Evjemo, J.O. Pilot Production of Gammarids (Original Title in Norwegian "Pilotproduksjon av Gammaridaer"); SINTEF Internal Report F18977; SINTEF Industrial Management: Trondheim, Norway, 2011. [Google Scholar]
- Evjemo, J.O. Mapping the Possibilities of Using Sludge for the Cultivation of Marine Biomass (Original Title in Norwegian "Kartlegging av Mulighetene for å Utnytte Slam til Dyrkning av Marin Biomasse"); SINTEF Internal Report F27561; SINTEF Industrial Management: Trondheim, Norway, 2016. [Google Scholar]
- Harlıoğlu, M.M.; Farhadi, A. Importance of Gammarus in aquaculture. Aquac. Int. 2018, 26, 1327–1338. [Google Scholar] [CrossRef]
- Alberts-Hubatsch, H.; Slater, M.J.; Beermann, J. Effect of diet on growth, survival and fatty acid profile of marine amphipods: Implications for utilisation as a feed ingredient for sustainable aquaculture. Aquac. Environ. Interact. 2019, 11, 481–491. [Google Scholar] [CrossRef]
- Monroig, Ó.; Tocher, D.R.; Navarro, J.C. Biosynthesis of polyunsaturated fatty acids in marine invertebrates: Recent advances in molecular mechanisms. Mar. Drugs 2013, 11, 3998–4018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, L.F.C.; Tocher, D.R.; Monroig, O. Long-chain polyunsaturated fatty acid biosynthesis in chordates: Insights into the evolution of Fads and Elovl gene repertoire. Prog. Lipid Res. 2016, 62, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Hao, M.; Huang, Y.; Zou, W.; Rong, H.; Wen, X. Cloning, tissue distribution and nutritional regulation of a fatty acyl Elovl4-like elongase in mud crab, Scylla paramamosain (Estampador, 1949). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2018, 217, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Mah, M.Q.; Kuah, M.K.; Ting, S.Y.; Merosha, P.; Janaranjani, M.; Goh, P.T.; Jaya-Ram, A.; Shu-Chien, A.C. Molecular cloning, phylogenetic analysis and functional characterisation of an Elovl7-like elongase from a marine crustacean, the orange mud crab (Scylla olivacea). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2019, 232, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Ting, S.Y.; Janaranjani, M.; Merosha, P.; Sam, K.; Wong, S.C.; Goh, P.; Mah, M.; Kuah, M.; Chong Shu-Chien, A. Two elongases, Elovl4 and Elovl6, fulfill the elongation routes of the LC-PUFA biosynthesis pathway in the orange mud crab (Scylla olivacea). J. Agric. Food Chem. 2020, 68, 4116–4130. [Google Scholar] [CrossRef]
- Sun, P.; Zhou, Q.; Monroig, Ó.; Navarro, J.C.; Jin, M.; Yuan, Y.; Wang, X.; Jiao, L. Cloning and functional characterization of an elovl4-like gene involved in the biosynthesis of long-chain polyunsaturated fatty acids in the swimming crab Portunus trituberculatus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2020, 242, 110408. [Google Scholar] [CrossRef]
- Poynton, H.C.; Hasenbein, S.; Benoit, J.B.; Sepulveda, M.S.; Poelchau, M.F.; Hughes, D.S.T.; Murali, S.C.; Chen, S.; Glastad, K.M.; Goodisman, M.A.D.; et al. The toxicogenome of Hyalella azteca: A model for sediment ecotoxicology and evolutionary toxicology. Environ. Sci. Technol. 2018, 52, 6009–6022. [Google Scholar] [CrossRef]
- Jakobsson, A.; Westerberg, R.; Jacobsson, A. Fatty acid elongases in mammals: Their regulation and roles in metabolism. Prog. Lipid Res. 2006, 45, 237–249. [Google Scholar] [CrossRef]
- Hashimoto, K.; Yoshizawa, A.C.; Okuda, S.; Kuma, K.; Goto, S.; Kanehisa, M. The repertoire of desaturases and elongases reveals fatty acid variations in 56 eukaryotic genomes. J. Lipid Res. 2008, 49, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Ran, Z.; Xu, J.; Liao, K.; Monroig, Ó.; Navarro, J.C.; Oboh, A.; Jin, M.; Zhou, Q.; Zhou, C.; Tocher, D.R.; et al. Biosynthesis of long-chain polyunsaturated fatty acids in the razor clam Sinonovacula constricta: Characterization of four fatty acyl elongases and a novel desaturase capacity. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 1083–1090. [Google Scholar] [CrossRef]
- Le, S.Q.; Dang, C.C.; Gascuel, O. Modeling protein evolution with several amino acid replacement matrices depending on site rates. Mol. Biol. Evol. 2012, 29, 2921–2936. [Google Scholar] [CrossRef] [Green Version]
- Neuparth, T.; Machado, A.M.; Montes, R.; Rodil, R.; Barros, S.; Alves, N.; Ruivo, R.; Castro, L.F.C.; Quintana, J.B.; Santos, M.M. Transgenerational inheritance of chemical-induced signature: A case study with simvastatin. Environ. Int. 2020, 144, 106020. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, H.; Zheng, H.; Wang, S.; Guo, Z.; Zhang, G. PUFA biosynthesis pathway in marine scallop Chlamys nobilis Reeve. J. Agric. Food Chem. 2014, 62, 12384–12391. [Google Scholar] [CrossRef]
- Monroig, Ó.; Guinot, D.; Hontoria, F.; Tocher, D.R.; Navarro, J.C. Biosynthesis of essential fatty acids in Octopus vulgaris (Cuvier, 1797): Molecular cloning, functional characterisation and tissue distribution of a fatty acyl elongase. Aquaculture 2012, 360–361, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Monroig, Ó.; Hontoria, F.; Varó, I.; Tocher, D.R.; Navarro, J.C. Investigating the essential fatty acids in the common cuttlefish Sepia officinalis (Mollusca, Cephalopoda): Molecular cloning and functional characterisation of fatty acyl desaturase and elongase. Aquaculture 2016, 450, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Monroig, Ó.; Lopes-Marques, M.; Navarro, J.C.; Hontoria, F.; Ruivo, R.; Santos, M.M.; Venkatesh, B.; Tocher, D.R.; Castro, L.F. Evolutionary functional elaboration of the Elovl2/5 gene family in chordates. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Zerangue, N.; Schwappach, B.; Jan, Y.N.; Jan, L.Y. A new ER trafficking signal regulates the subunit stoichiometry of plasma membrane K(ATP) channels. Neuron 1999, 22, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Sprecher, H. Metabolism of highly unsaturated n-3 and n-6 fatty acids. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2000, 1486, 219–231. [Google Scholar] [CrossRef]
- Monroig, O.; Tocher, D.R.; Castro, L.F.C. Polyunsaturated Fatty acid biosynthesis and metabolism in fish. In Polyunsaturated Fatty Acid Metabolism; Elsevier Inc.: Cambridge, MA, USA, 2018; pp. 31–60. ISBN 9780128112304. [Google Scholar]
- Deák, F.; Anderson, R.E.; Fessler, J.L.; Sherry, D.M. Novel cellular functions of very long chain-fatty acids: Insight from ELOVL4 mutations. Front. Cell. Neurosci. 2019, 13, 428. [Google Scholar] [CrossRef]
- Monroig, Ó.; De Llanos, R.; Varó, I.; Hontoria, F.; Tocher, D.R.; Puig, S.; Navarro, J.C. Biosynthesis of polyunsaturated fatty acids in Octopus vulgaris: Molecular cloning and functional characterisation of a stearoyl-CoA desaturase and an elongation of very long-chain fatty acid 4 protein. Mar. Drugs 2017, 15, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monroig, O.; Rotllant, J.; Cerdá-Reverter, J.M.; Dick, J.R.; Figueras, A.; Tocher, D.R. Expression and role of Elovl4 elongases in biosynthesis of very long-chain fatty acids during zebrafish Danio rerio early embryonic development. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2010, 1801, 1145–1154. [Google Scholar] [CrossRef] [Green Version]
- Agbaga, M.P.; Mandal, N.A.; Anderson, R.E. Retinal very long-chain PUFAs: New insights from studies on ELOVL4 protein. J. Lipid Res. 2010, 51, 1624–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, R.; Navarro, J.C.; Portolés, T.; Sales, C.; Beltrán, J.; Monroig, Ó.; Hernández, F. Identification of new, very long-chain polyunsaturated fatty acids in fish by gas chromatography coupled to quadrupole/time-of-flight mass spectrometry with atmospheric pressure chemical ionization. Anal. Bioanal. Chem. 2020, 413, 1039–1046. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Horton, T.; Kroh, A.; Ahyong, S.; Bailly, N.; Boyko, C.B.; Brandão, S.N.; Gofas, S.; Hooper, J.N.A.; Hernandez, F.; Holovachov, O.; et al. World Register of Marine Species (WoRMS). Available online: http://www.marinespecies.org (accessed on 8 August 2020).
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. Proc. Gatew. Comput. Environ. Work. 2010, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, F.; Domelevo Entfellner, J.B.; Wilkinson, E.; Correia, D.; Dávila Felipe, M.; De Oliveira, T.; Gascuel, O. Renewing Felsenstein’s phylogenetic bootstrap in the era of big data. Nature 2018, 556, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Lutteropp, S.; Kozlov, A.M.; Stamatakis, A. A fast and memory-efficient implementation of the transfer bootstrap. Bioinformatics 2020, 36, 2280–2281. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Monroig, Ó.; Navarro, J.C.; Tocher, D.R.; Zhou, Q.C. Molecular and functional characterisation of two elovl4 elongases involved in the biosynthesis of very long-chain (>C24) polyunsaturated fatty acids in black seabream Acanthopagrus schlegelii. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2017, 212, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Marques, M.; Ozório, R.; Amaral, R.; Tocher, D.R.; Monroig, Ó.; Castro, L.F.C. Molecular and functional characterization of a fads2 orthologue in the Amazonian teleost, Arapaima gigas. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2017, 203, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Monroig, Ó.; Tocher, D.; Hontoria, F.; Navarro, J. Functional characterisation of a Fads2 fatty acyl desaturase with Δ6/Δ8 activity and an Elovl5 with C16, C18 and C20 elongase activity in the anadromous teleost meagre (Argyrosomus regius). Aquaculture 2013, 412–413, 14–22. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| FA Substrate | FA Product | Elovl4 | Elovl6 | Elovl1/7-Like | Activity |
|---|---|---|---|---|---|
| 18:3n-3 | 20:3n-3 | 2.67 | 0.22 | 0.29 | C18→C20 |
| 18:2n-6 | 20:2n-6 | 1.32 | 0.11 | 0.19 | C18→C20 |
| 18:4n-3 | 20:4n-3 | 1.82 | 0.33 | 1.46 | C18→C20 |
| 22:4n-3 | 0.02 | nd | 0.02 | C20→C22 | |
| 18:3n-6 | 20:3n-6 | 1.21 | 0.35 | 0.87 | C18→C20 |
| 20:5n-3 | 22:5n-3 | 2.52 | nd | 13.17 | C20→C22 |
| 24:5n-3 | 0.05 | nd | 0.17 | C22→C24 | |
| 20:4n-6 | 22:4n-6 | 1.61 | nd | 5.75 | C20→C22 |
| 24:4n-6 | 0.02 | nd | 0.09 | C22→C24 | |
| 22:5n-3 | 24:5n-3 | 0.92 | nd | 2.16 | C22→C24 |
| 26:5n-3 | 0.09 | nd | nd | C24→C26 | |
| 22:4n-6 | 24:4n-6 | 0.46 | nd | 0.72 | C22→C24 |
| 22:6n-3 | 24:6n-3 | 0.38 | nd | 0.54 | C22→C24 |
| Gene | Aim | Primer Name | Primer Sequence | Cycles | Tm | Extension |
|---|---|---|---|---|---|---|
| ELOVL4 | 1st fragment generation | EM_ELOVL4_ F1 | TCTACAACCTTGCTGTCATG | 35 | 55 °C | 72 °C (1 min) |
| EM_ELOVL4_ R2 | TGCACAAAGCTGTTCATCAT | |||||
| 3’RACE PCR | 3’Outer_Primer | GCGAGCACAGAATTAATACGACTCACTATAGGT12 | 35 | 55 °C | 72 °C (75 s) | |
| 3’Inner_Primer | CGCGGATCCGAATTAATACGACTCACTATAGGT12 | |||||
| EM_ELOVL4_F3 | CACGTGTATCACCACTCGAC | |||||
| EM_ELOVL4_F4 | GGATTGGAGTCAAGTTTGTGG | |||||
| EM_ELOVL4_F5 | CCTGGCGGCAATGATGAACA | |||||
| Full ORF | EM_ELOVL4_ORF_U5F | ATGAATTTAGTGAACAACAA | 35 | 62 °C (10 cycles) 58 °C (25 cycles) | 72 °C (1 min) | |
| EM_ELOVL4_ORF_U3R | CAAGATGCCTGAACTCCCGGT | |||||
| Functional characterisation | EM_FW_ELOVL4_HindIII EM_RV_ELOVL4_XbaI | CCCAAGCTTACAATGGCTGCCTCTGTT CCGTCTAGACTACATGTCCTTTCGAGG | 35 | 62 °C (10 cycles) 58 °C (25 cycles) | 72 °C (45 s) | |
| ELOVL6 | 1st fragment generation | EM_ELOVL6_ F1 EM_ELOVL6_ R2 | GGCTTCTGGAACTGGATGTT TTCATGTAGGCACGACGGAA | 35 | 55 °C | 72 °C (1 min) |
| Full ORF | EM_ELOVL6_ORF_U5F EM_ELOVL6_ORF_U3R | CTTTACCACGTTTTACTGGG ACTGGTAGTTTTGTATGCAT | 35 | 62 °C (10 cycles) 58 °C (25 cycles) | 72 °C (1 min) | |
| Functional characterisation | EM_FW_ELOVL_HindIII EM_RV_ELOVL_XbaI | CCCAAGCTTACGATGGCCCTCTCGGAC CCGTCTAGATTAATCGAACTTCCCTCCCT | 35 | 62 °C (10 cycles) 58 °C (25 cycles) | 72 °C (45 s) | |
| ELOVL1/7-LIKE | 1st fragment generation | EM_ELOVL_ F1 | GTCATCCACCACGGATGCATG | 35 | 55 °C | 72 °C (1 min) |
| EM_ELOVL_ R1 | GCCTTCACGTAGAAGTTGGAG | |||||
| Full ORF | EM_ELOVL_ORF_U5F | AAAAACGTGTTCTCGGCCAG | 35 | 62 °C (10 cycles) 58 °C (25 cycles) | 72 °C (1 min) | |
| EM_ELOVL_ORF_U3R | GAGGCTTAACTAAAACGAAC | |||||
| Functional characterisation | EM_FW_ELOVL_HindIII EM_RV_ELOVL_XbaI | CCCAAGCTTAAGATGGCGGGTACAGCA CCGTCTAGATCAGTCGTCCTTCCGAGG | 35 | 62 °C (10 cycles) 58 °C (25 cycles) | 72 °C (45 s) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribes-Navarro, A.; Navarro, J.C.; Hontoria, F.; Kabeya, N.; Standal, I.B.; Evjemo, J.O.; Monroig, Ó. Biosynthesis of Long-Chain Polyunsaturated Fatty Acids in Marine Gammarids: Molecular Cloning and Functional Characterisation of Three Fatty Acyl Elongases. Mar. Drugs 2021, 19, 226. https://doi.org/10.3390/md19040226
Ribes-Navarro A, Navarro JC, Hontoria F, Kabeya N, Standal IB, Evjemo JO, Monroig Ó. Biosynthesis of Long-Chain Polyunsaturated Fatty Acids in Marine Gammarids: Molecular Cloning and Functional Characterisation of Three Fatty Acyl Elongases. Marine Drugs. 2021; 19(4):226. https://doi.org/10.3390/md19040226
Chicago/Turabian StyleRibes-Navarro, Alberto, Juan C. Navarro, Francisco Hontoria, Naoki Kabeya, Inger B. Standal, Jan O. Evjemo, and Óscar Monroig. 2021. "Biosynthesis of Long-Chain Polyunsaturated Fatty Acids in Marine Gammarids: Molecular Cloning and Functional Characterisation of Three Fatty Acyl Elongases" Marine Drugs 19, no. 4: 226. https://doi.org/10.3390/md19040226