
Antimicrobial Action of Compounds from Marine Seaweed


1
Departamento de Biología Funcional y Ciencias de la Salud, Facultad de Ciencias, Universidade de Vigo, As Lagoas, Ourense 32004, Spain
2
Departamento de Química Analítica, Facultad de Ciencias, Universidade de Vigo, As Lagoas, Ourense 32004, Spain
3
Departamento de Enxeñería Química, Facultad de Ciencias. Universidade de Vigo, As Lagoas, Ourense 32004, Spain
*
Author to whom correspondence should be addressed.
Mar. Drugs 2016, 14(3), 52; https://doi.org/10.3390/md14030052
Submission received: 18 December 2015
/
Revised: 18 January 2016
/
Accepted: 25 February 2016
/
Published: 9 March 2016
(This article belongs to the Special Issue Antibacterial Marine Pharmacology)
Abstract
:Seaweed produces metabolites aiding in the protection against different environmental stresses. These compounds show antiviral, antiprotozoal, antifungal, and antibacterial properties. Macroalgae can be cultured in high volumes and would represent an attractive source of potential compounds useful for unconventional drugs able to control new diseases or multiresistant strains of pathogenic microorganisms. The substances isolated from green, brown and red algae showing potent antimicrobial activity belong to polysaccharides, fatty acids, phlorotannins, pigments, lectins, alkaloids, terpenoids and halogenated compounds. This review presents the major compounds found in macroalga showing antimicrobial activities and their most promising applications.
1. Introduction
Marine organisms produce a variety of compounds with pharmacological activities, including anticancer, antimicrobial, antifungal, antiviral, antiinflammatory and others, and are potential sources of new therapeutic agents. Marine organisms survive and live within complex communities and in close association with others in a competitive and hostile environment. They produce complex secondary metabolites as a response to ecological pressure, such as competition for space, predation and tide variations. Some of these compounds are antimicrobials that inhibit or limit the development and growth of other competitive microorganisms.
Marine sessile organisms, such as algae, sponges and corals, have developed physiological adaptations, including the synthesis of bioactives which confer defense against grazers and/or the installation of epiphytes and fouling organisms [1,2,3]. Metabolites from green, brown and red marine algae may be useful for inhibiting bacteria, viruses, fungi and other epibionts (e.g., cytostatic, antiviral, antihelmintic, antibacterial, antifungal activity). Algae crude extracts and their fractionated or purified components also exhibit, for example, anticoagulant [4], antiviral [5], antioxidant [6], anticancer [7], and antiinflammatory [8] activities.
Microorganisms have developed new strategies to evade the action of antibiotics, leading to multiple drug-resistant bacterial strains. With increasing resistance of pathogens to antibiotics, there is a public health priority for exploring and developing cheaper and effective natural antimicrobial agents with better potential, less side effects than antibiotics, good bioavailability, and minimal toxicity [9]. It is also worthwhile to test the marine antimicrobials for possible synergism with existing drugs [10].
Updated information on this research area has been compiled recently, including the antiviral properties of marine organisms [11], the seaweed-associated bacterial and fungal communities [12], and the diversity and bioactives production of actinobacteria associated with the marine organisms [13]. Based on published studies, Vatsos and Rebours [14] reviewed the antimicrobial properties of seaweed extracts related with aquaculture. Eom et al. [15] reviewed the antimicrobial effects of phlorotannins from brown algae, in relation to the food and pharmaceutical industries. Abu-Ghannam and Rajauria [16] reviewed the algal antimicrobials with potential food applications. The studies showing biological activities of extracts from native and some non-native Brazilian seaweed [17] and the research progress concerning the isolation and structural elucidation of the secondary metabolites from the genus Cystoseira [18] were overviewed. The objective of this work is to gather the recently published information on the antimicrobial properties of compounds from seaweed, their extraction and the major applications.
2. Bioactive Compounds
In the food, pharmaceutical, cosmetic, cosmeceutical, nutraceutical and biomedicine industries, seaweed/macroalgae are used as a valuable source of bioactive compounds. Many compounds, being antiparasitic, antiviral or antibacterial, are effective [19]. The influence of some natural factors, such as the environmental conditions, including light, temperature or salinity, the life stage, reproductive state and age of the seaweed, and the geographical location and seasonality, allowed for the consideration that this antimicrobial activity was not attributed to a single compound, but it could be related to some of them and to a combination of metabolites.
Seaweed or macroalgae provide a great variety of metabolites and natural bioactive compounds with antimicrobial activity, such as polysaccharides, polyunsaturated fatty acids, phlorotannins and other phenolic compounds, and carotenoids.
2.1. Polysaccharides and Derived Oligosaccharides
The main components of green, brown and red seaweed are usually polysaccharides, which may have storage and structural functions. Cell walls of algae are composed of a variety of polysaccharides including alginic acid and alginates, carrageenans and agar, laminarans, fucoidans, ulvans and derivatives [20,21].
Their antimicrobial activity depends on some factors, such as their distribution, molecular weight, charge density, sulphate content (in sulphated polysaccharides), and structural and conformation aspects. In addition, oligosaccharides obtained by depolymerization of seaweed polysaccharides also induce protection against viral, fungal and bacterial infections in plants [22].
These polymeric carbohydrates structures are usually composed of various monosaccharides linked with different glucosidic bonds. Some algal polysaccharides, such as sulphated galactans of the red algae or ulvans of the green algae, have linear backbones containing dissacharide repeating units. Otherwise, alginic acids have linear molecules built up of different blocks of two monomerics units. Algal macromolecules include sulfated polysaccharides such as: carrageenan and agar from red algae; alginate, fucan and laminarinan from brown alga; and cellulose and ulvan from green algae.
2.1.1. Alginates
Algins/alginates are available in both acid and salt forms. Alginic acids are linear copolymers of two uronic acids, β-d-mannuronic acid (M) and α-l-guluronic acid (G) linked in position 1→4. The salt forms (alginates), with several cations (Na+, K+, Mg2+ and Ca2+), are the major components of brown seaweed cell walls and also of the intracellular matrix [21]. Alginates are anionic polysaccharides and are made up of β-d-mannuronic acid (M) and α-l-guluronic acid (G), and alternating blocks of d-guluronic and d-mannuronic (M-M, G-G or M-G blocks) [22,23] (Figure 1). The molecular weight of alginate ranges generally between 500 and 1000 kDa.
2.1.2. Carrageenans
Carrageenans are the major components of red seaweed cell walls, and this group of molecules is composed of linear polysaccharide chains with sulphate half-esters attached to the sugar unit. There are three general forms: kappa, lambda and iota, according to the degree of sulphation (Figure 1). Kappa carrageenan and iota carrageenan have one or two, respectively, sulphate groups per disaccharide unit and anhydrogalactose residue, and lambda carrageenan has three sulphate groups per disaccharide unit [22].
2.1.3. Agar
Agar is a mixture of at least two polysaccharides, i.e., agarose and agaropectin, extracted also from red seaweed with similar structural and functional properties as carrageenans. Agarose is the predominant fraction of agar, and it consists of high molecular weight polysaccharides composed of repeating units of (1→3)-β-d-galactopyranosyl-(1→4)-3,6-anhydro-α-l-galactopyranose (Figure 1). The structure of agaropectin, with a lower molecular weight than agarose, is essentially made up of alternating (1→3)-β-d-galactopyranose and of (1→4)-3,6-anhydro-α-l-galacto-pyranose residues.
2.1.4. Galactans
Sulphated galactans are the main extracellular polysaccharides of red algae (but are also found in brown and green algae). Typical structure is a linear chain of galactoses; a chain of alternating 3-β-d-galactopyranose (G units) and 4-α-d-galactopyranose residues, or 4-3,6-anhydrogalactopyranose residues complete their structural backbone with presence of d-series (d unit) in carrageenans and l-series (l unit) in agarans. Other exceptional galactans present the dl-hybrids that enclose G unit attached to both d and l units [24].
2.1.5. Laminarans
Laminaran is the principal storage polysaccharide of brown seaweed (e.g., Laminaria or Saccharina spp.) and their content can represent up to 32%–35% (d.w.). Laminarans are small glucans and are a linear polysaccharide composed of β-(1→3)-linked glucose, containing randomly β-(1→6) intra-chain branching, with a ratio around 3:1 [23]. The degree of polymerization varying between 20 to 50 units and the polymeric chains can be of two types according to their reducing end: M chains end with a d-mannitol residue, whereas G chains end with a glucose residue (Figure 1). The molecular weight is approximately 5000 Da depending on the degree of polymerization (usually 25) [25].
2.1.6. Fucoidans/Fucans
Fucoidans and laminarans are considered as the main water-soluble polysaccharides of brown algae. Fucoidans are a complex and heterogeneous group of polysaccharides, which contribute to intercellular mucilage and are sulfated polysaccharides composed of l-fucose and sulphate ester groups with minor amounts of different molecules, which can vary from monosaccharides (i.e., mannose, arabinose, glucose, galactose, xylose, etc.), acidic monosaccharides, acetyl groups to proteins [26].
Terms such as fucans, fucosans, fucose containing polymers or sulfated fucans have also been adopted for this group; however, finally, according to the IUPAC terminology, fucoidans is retained for polysaccharides of algal origin, and fucan sulphates (or fucans) to the similar polymers from marine invertebrates [21].
Various molecular weights, from 100 to 1600 kDa, have been reported in the literature for fucoidans [25] because they may differ considerably in their composition and chemical structure (degree of branching, substituents, sulphation and type of linkages).
Fucoidan composition varies with species and geographical origin, even within the same species. It appears that the prevalent core backbone structures are primarily the (1→3)-linked α-l-fucopyranosyl backbone structure, and secondarily the backbone structure composed of alternating α (1→3) and α (1→4)-linked l-fucopyranosyls [27]. The first group includes the fucoidans from Laminaria spp., Analipus japonicus, Cladosiphon okamuranus, and Chorda filum and the second group included fucoidans isolated from Ascophyllum nodosum and Fucus sp. However, sulphate- and acetyl-groups and some sugar residues may occur at C2 and/or at C4 positions.
2.1.7. Ulvans
Ulvan designates a water-soluble sulphated polysaccharides extracted from the intercellular space and in the fibrillar wall of green seaweed (mainly Ulva sp.) and accounts from 18% to 29% of the algal dry weight [28]. These polysaccharides are mainly composed of glucuronic acid and iduronic acid units together with rhamnose and xylose sulfates, connected by α- and β-1→4 bonds, with an average molecular weight of ulvans ranging from 189 to 8200 kDa.
The main repeating disaccharide units reported are ulvanobiouronic acid 3-sulphate types containing either glucuronic or iduronic acid (Figure 1). Additionally, minor repeating units have been reported that contain sulfated xylose replacing the uronic acid or glucuronic acid as a branch on O-2 of the rhamnose-3-sulphate [29].
2.2. Lipids, Fatty Acids Ans Sterols
Algal lipids content in seaweed ranges from 0.12% to 6.73% (dry weight), and are composed mainly of phospholipids, glycolipids and non-polar glycerolipids (neutral lipids) [30]:
- Phospholipids are located in extra-chloroplast membranes and account for 10%–20% of total lipids in algae. They are characterized by higher contents of n-6 fatty acids, and the major fatty acids present are oleic, palmitic, stearic, arachidonic and eicosapentanoic acids. The most dominant phospholipid in algae is phosphatidylglycerol in green algae, phosphatidylcholine in red algae, and phosphatidylcholine and phosphatidylethanolamine in brown algae.
- Glycolipids are located in photosynthetic membranes and constitute more than half of the lipids in the main algal groups. They are characterized by high n-3 polyunsaturated fatty acids. Three major types of glycolipids are monogalactosyldiacylglycerides, digalactosyldiacylglycerides, and sulfoquinovosyldiacylglycerides [31].
- Triacylglicerol is the most prevalent neutral lipid, their content ranging from 1% to 97% with a function of storage and energy reservoir.
Fatty acids are carboxylic acids with aliphatic chains and prevalent even carbon numbers (C4-C28) that may be straight or branched, saturated or unsaturated. According to the double bond, fatty acids are classified as monounsaturated (MUFA) or polyunsaturated (PUFA) fatty acids, and the last one could be classified as n-3 or n-6 depending on the position of the first double bond from the methyl end. Green algae are rich in C18 PUFAs, mainly α-linolenic (C18:3 n-3), stearidonic (C18:4 n-3) and linoleic (C18:2 n-6) acids; red algae is rich in C20 PUFAs, mainly arachidonic (C20:4 n-6) and eicosapentaenoic (C20:5 n-3) acids, and brown algae exhibit both.
Oxylipins are the oxygenated products of fatty acids and are mainly derived from C16, C18, C20 and C22 PUFAs and confer innate immunity in response to biotic and abiotic stress, such as pathogenic bacteria and herbivores. [30].
Sterols are structural components of cell membrane and regulate membrane fluidity and permeability. They are composed of four rings (A–D) with a hydroxyl group in carbon-3, two methyl groups at C18 and C19 carbons and a side chain at C17 (see Figure 2). The main sterols in macroalgae are cholesterol, fucosterol, isofucosterol, clionasterol [30].
2.3. Phenolic Compounds
Phenolic compounds are secondary metabolites because they are not directly involved in primary processes such as photosynthesis, cell division or reproduction of algae. They are characterized by an aromatic ring with one or more hydroxyl groups and the antimicrobial action is due to the alteration of microbial cell permeability and the loss of internal macromolecules or by the interference with the membrane function and loss of cellular integrity and eventual cell death [16].
Chemically, structures ranging from simple phenolic molecules to complex polymers with a wide range of molecular sizes (126–650 kDa) have been described [26]. Polyphenols could be divided into phloroglucinols and phlorotannins. Phloroglucinol contains an aromatic phenyl ring with three hydroxyl groups (Figure 2). Phlorotannins are oligomers or polymers of phloroglucinol with additional halogen or hydroxyl groups; and, according to the inter-linkage, phlorotannins can be subdivided into six specific groups: (i) phlorethols (with aryl-ether linkage); (ii) fucols (with aryl-aryl bonds); (iii) fucophlorethols (with ether or phenyl linkage); (iv) eckols (with dibenzo[1,4]dioxin linkages); the less frequent (v) fuhalols (with ortho-/para-arranged ether bridges containing an additional hydroxyl on one unit); and (vi) carmalols (with dibenzodioxin moiety) [32]. The presence of simple phenols, such as hydroxycinnamic and benzoic acids and derivates, and flavonoids were reported in the green seaweed [33], but brown seaweed has higher contents of phenolic compounds than green and red macroalgae. The typical phlorotannin profile from brown algal with antimicrobial activity mainly consists of phloroglucinol, eckol and dieckol [19].
2.4. Pigments
Algae as photosynthetic organisms can synthezise the three basic classes of pigments found in marine algae:chlorophylls, carotenoids and phycobiliproteins, allowing classification of seaweed into Chlorophyceae (green algae), Phaeophyceae (brown algae) and Rhodophyceae (red algae). The green color is due to the presence of chlorophylls a and b, the greenish brown color is attributed to the fucoxanthin, chlorophylls a and c, and responsible for the red color are the phycobilins, such as phycoerythrin and phycocyanin [34].
The antimicrobial mechanism proposed for carotenoids could lead to the accumulation of lysozime, an immune enzyme that digests bacterial cell walls [16]. Carotenoids are present in all algae and are lipid-soluble, natural pigments composed of eight units of five carbons, namely tetraterpenoids, with up to 15 conjugated double bonds. Carotenoids are usually divided in two classes: carotenes (when the chain end with a cyclic group, containing only carbon and hydrogen atoms) and xantophylls or oxycarotenoids (which have at least one oxygen atom as a hydroxyl group, as an oxy-group or as a combination of both). β-Carotene is the most common carotene (Figure 2), whereas lutein, fucoxanthin (Figure 2) and violaxanthin belong to the xanthophylls class [35]. β-Carotene, lutein, violaxanthin, neoxanthin and zeaxanthin are found in green seaweed species; α- and β-carotene, lutein and zeaxanthin are present in red seaweed, and β-carotene, violaxanthin and fucoxanthin are found in brown algae [34].
2.5. Other Compounds
Seaweed is also able to produce other secondary metabolites with a broad range of antifungal, antiviral, antibacterial, antimacrofouling and antiprotozoan activities, such as terpenes, alkaloids, lectins or halogenated compounds.
2.5.1. Lectins
Lectins are natural bioactive ubiquitous proteins or glycoproteins of non-immune response that bind reversibly to glycans of glycoproteins, glycolipids and polysaccharides possessing at least one non-catalytic domain causing agglutination.
Algal lectins differ from terrestrial lectins because they are monomeric, low molecular weight proteins, exhibiting high content of acidic amino acids, with isoelectric point in the range of 4–6. They do not require metal ions for their biological activities, and most of them show higher specificity for oligosaccharides and/or glycoproteins than for monosaccharides. Based on the binding properties to glycoproteins, algal lectins are categorized into three major categories: complex type N-glycan specific lectins, high mannose (HM) type N-glycan specific lectins and lectins with specificity to both the above types of N-glycans [36]. Lectins from marine organisms are also classified into C-type lectins, F-type lectins, galectins, intelectins, and rhamnose-binding lectins [37].
2.5.2. Alkaloids
An alkaloid is a compound that has nitrogen atom(s) in a cyclic ring. Numerous biological amines and halogenated cyclic nitrogen-containing substances are included in the term alkaloid. The latter is specific to marine organisms and marine algae. They could not be found in terrestrial plants. Alkaloids in marine algae were classified in three groups as follows [38]: (i) phenylethylamine alkaloids; (ii) indole and halogenated indole alkaloids; and (iii) other alkaloids, such as 2,7-naphthyridine derivatives. Alkaloids isolated from marine algae mostly belong to 2-phenylethylamine and indole groups. Halogenated alkaloids are specific for algae, being bromine- and chloride-containing alkaloids particularly dominant in Chlorophyta. Most of the alkaloids of the indole group are concentrated in Rhodophyta [39].
2.5.3. Terpenes
Terpenes represent one of the major classes of metabolites produced by marine algae. Chemically, they are derived from the five-carbon precursor isopentenyl pyrophosphate, and are classified into hemiterpenes (C5), monoterpenes (C10), sesquiterpenes (C15), diterpenes (C20), sesterterpenes (C25), triterpenes (C30) and polyterpenes (>C30). Chlorophyceae contain cyclic and linear sesqui-, di-, and triterpenes, while Rhodophyceae are characterized by a high structural diversity of halogenated secondary metabolites whose polyhalogenated monoterpenes exhibit a wide range of antimicrobial activities [40].
Some terpenes were isolated from seaweed [41], such as the diterpene neophytadiene (Figure 2); the sesquiterpenes cartilagineol, obtusol and elatol; the diterpene aldehyde halitunal which shows antiviral activity; the dolastane diterpenes 4-hydroxy-9,14-dihydroxydolasta-1(15),7-diene (1) and 4,7,14-trihydroxydolasta-1(15),8-diene (2); the diterpenoid halimediatrial and halimedalactone; the antibiotic cyclopropane containing sesquiterpene cycloeudesmol; the sesquiterpenoid puupehenone and its derivatives; etc. Many of them present antiviral activity; others, such as the sesquiterpene (−)-elatol are antifouling agents, and capisterones, which are triterpene sulphate esters (Figure 2), exhibit potent antifungal action.
2.5.4. Halogenated Compounds
Other compounds found in macroalgae are halogenated metabolites, mainly brominated [1,33]: furanones 4-bromo-3-butyl-5-(dibromoethylene)-2(5H)-furanone and 4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone; bromoditerpenes, such as 12 S-hydroxybromospha-erodiol, bromosphaerone and isoparguerol and their derivatives; bromophenols, such as bis(2,3-dibromo-4,5-dihydroxybenzyl) ether, 5-bromo-3,4-dihydroxybenzaldehyde and 3,3′,5,5′-tetrabromo-2,2′,4,4′tetrahydroxydiphenylmethane (Figure 2); and polar compounds, such as 2,3-dibromo-4,5-dihydroxyphenyl-ethylamine and 3,4-dihydroxyphenyl-ethylamine.
3. Assesment of Antimicrobial Activity
Several methods are widely used by researchers to detect and to measure the antimicrobial activity of algal extracts or their metabolites. Authors refer most often to in vitro and sometimes in vivo assays, but different algal extracts quantities and microorganisms are tested, making it difficult to unify results. In some cases, an initial in vitro screening is followed by an in vivo study, but most studies on the antimicrobial effects of seaweed are either only in vitro or only in vivo. Vatsos and Rebours [14] report the different assays published about antimicrobial activity of seaweed extracts in aquaculture. Here, we will exhibit the in vitro and in vivo methods more commonly referred to by authors in different fields.
3.1. In Vitro Assays
3.1.1. Diffusion Agar Tests
The method widely used by authors to evaluate antibacterial and antifungical activity is the agar disk diffusion test. Briefly, it is based on the use of sterile paper disks impregnated with the algal extract to be evaluated (concentrations varies greatly from one author to another [42,43,44,45,46,47]) and are deposited in an agar plate with a particular bacterial or fungal culture (the suspensions of microorganisms ranging between 105 cells/mL [48] and 108 colony forming units (cfu)/mL [42,49]. Soluble extracts diffuse into the culture medium (generally Mueller-Hinton Agar used for bacteria and Saboureaud Agar plates, with or without chloramphenicol for fungal strains) and after the incubation time suitable for each microorganism, and the halos of inhibition of microbial growth around the disks are recorded and measured. Bansemir et al. [47] refer to the use of p-iodo nitrotetrazolium violet, 5% in 50% aqueous ethanol to detect the inhibition zone because living bacteria produce a red-colored compound by reaction with the coloring solution and the inhibition zone appears colorless.
The diffusion agar test can be performed also preparing wells in agar. Thanigaivel et al. [50,51] prepared Petri plates containing the culture medium and inoculated them by swabbing the microorganism chosen. Once dried under sterile conditions, wells 6–8 mm in diameter were made with a punch, and the algal extracts were placed into them. Shanmughapriya et al. [52] performed the agar well diffusion method using porcelain beans. The plates were incubated under the optimal conditions for each microorganism, and the presence or not of an inhibition zone was determined.
3.1.2. Growth Inhibition Assay
García Bueno et al. [53] tested the antibacterial activity of water-soluble seaweed extracts in 96-well plates. Bacterial growth in the presence of algal extracts was monitored by measuring optical density (OD) at 490 nm every 30 min for 24 h. After incubation, the intensity of growth in the presence of the tested compounds and controls was compared. Cox et al. [54] assessed the antibacterial activity of varying concentrations of a hydrophilic extract from Himanthalia elongata in carbohydrate and protein model food systems, and Dubber and Harder [55] used a highly sensitive growth inhibition assay that recorded the fluorescence intensity of stained DNA.
Pinteus et al. [43] determined the antifungal activity by following the yeast growth in yeast extract peptone dextrose (YPD) broth in the presence or absence of extracts. The growth was monitored spectrophotometrically (OD 600 nm) after 20 h of incubation. The growth inhibition was assayed as the percentage of change in turbidity in the presence or absence of algal extracts.
3.1.3. Minimun Inhibitory Concentration (MIC) Determination
MIC represents the lowest concentration of crude or purified algal extracts that inhibits the bacterial or fungal growth. The concentration serial broth (micro) dilution assay has been used in several studies [56,57,58].
Boisvert et al. [56] used a microdilution method using 96-well flat bottom microplate. Two-fold serial dilutions of extracts samples were made by repeated transfers of 125-μL volumes of tryptic soy broth (TSB) in a 96-well flat-bottom microplate. Each well was inoculated with 50 μL of bacterial suspension adjusted to 105 cfu/mL. Optical density was monitored every 10 min with a microplate spectrophotometer, and percentage of inhibition was calculated. Bazes et al. [57] assayed different concentrations of algal extracts incubated with 2 × 108 cells/mL for 48 h at 20 °C; after, that growth was monitored by measuring OD600.
Hellio et al. [58], determined the MIC of crude algal extracts via the macrodilution method. They assayed the extract concentration between 96 and 4 μg/mL, and 2 × 108 cfu/mL were placed in a liquid medium suitable for microorganisms tested with algal extracts.
The MTT (methylthiazol tetrazolium) cell proliferation assay was used by Spavieri et al. [59] to evaluate the antitubercular activity against Mycobacterium tuberculosis. MICs values were measured using aproppiated bacterial media in a 96-well flat bottom plate. Serial extract dilutions (final concentrations of 1–256 μg/mL) and 100 μL with 2.5 × 107 Mycobacterium tuberculosis strain H37Rv were added to the wells. The plate was incubated with gentle rocking for 7 days at 37 °C. Ten μL of MTT were added to each well, and the plates were then incubated for a further 24 h. MICs were recorded as the lowest concentration at which a purple precipitate of formazin did not appear in the wells.
3.1.4. Other Studies
The antiprotoozal activity of 90 to 123 µg/mL of brown algae extracts against 104/50 µL bloodstream forms of Tripanosoma brucei rhodesiense, 5000/100 µL trypomastigote forms of T. cruzi, and 105 amastigotes of Leishmania donovani were measured by Spavieri et al. [59] using 96-well microtiter plates; after incubation, microplates were read and data analyzed using a microplate reader.
Bazes et al. [57] assayed inhibition of phytoplankton growth by algal extracts. The effect of different concentrations of algal extracts on phytoplanctonic strains was assessed after 72 h by estimating the chlorophylla-a content. The percentage of growth inhibition was calculated. Additionally, they determined the percentage of inhibition of the germination of spores of Ulva sp. (600/mL) in plastic Petri dishes after incubation for 5 days at 20 °C under 24 h light.
Bouhlal et al. [60] determined the antiviral activity against Herpes simplex virus tipo1of algal extracts. Vero cells (African green monkey kidney cell line) were infected with HSV-1 suspension without or in the presence of different dilutions of the extracts. After incubation, antiviral activity was evaluated by the neutral red dye method. The 50% effective antiviral extract concentration (EC50) was expressed as the concentration that achieved a protection of 50% of virus-infected cells. OD measured at 540 nm was directly related to the percentage of viable cells, which was inversely related to the cytopathic effect.
Wang et al. [61] reviewed most of the studies on antiviral effects of marine polysaccharides carried out both in vitro or in mouse model systems. Therefore, further studies are needed in order to investigate their antiviral activities in human subjects.
3.2. In Vivo Assays
In the literature related to the assessment of antimicrobial activity of the crude extracts or fractions of algae, the in vivo assays are less numerous. In vivo assays depend critically on the target organism or substrate varying between one another. Vatsos and Rebours [14] refer to farmacodynamic, pharmacokinetic studies and artificial challenges like survival rate, progress of disease and severity of signs or lessions in the studies applied to aquaculture. For possible use in humans, they are much more restricted, so the detection assays in vitro antimicrobial activity will be fundamental. In this section, some of the in vivo assays based on published studies are collected.
Manilal et al. [62] assayed in vivo the therapeutic potential of crude algal extracts, which were rationalized with commercial shrimp feed and orally administered for different durations, followed by the artificial bacterial challenge experiment. The determination of the effective dose of medicated feed is necessary to perform the in vivo assays; after three weeks of feeding trial, ten shrimps from each group (medicated and control groups) were individually challenged with four species of live Vibrio. The challenged animals were monitored for a period of two weeks for mortality and infections. During this period, the shrimps were fed continuously with their respective feeds. The cause of death/infection was ascertained by reisolating the respective organism from the shrimp body and subjecting the isolates to standard biochemical tests. The morbidity and mortality were recorded daily and final mortality was evaluated after two weeks of postchallenge, and percentage of mortality was calculated.
Thanigaivel et al. [9,50,51] performed assays to demonstrate the antibacterial protection for fish by inmersion method. Therefore, for the experimentally induced infection of fish, different concentrations (101 to 105) of pathogenic bacterial cell suspension were added to the water where healthy fish were maintained. For treatment different concentrations varying from 50 mg/L to 500 mg/L of prepared seaweed extract were also added. Syntomatic or dead fish were recorded.
4. Production of Antimicrobials from Seaweed
Due to the different chemical natures of potential antimicrobials, the conditions leading to their maximal production, extraction and recovery should be optimized for each particular case. This section summarizes the information for the production of compounds with antimicrobial action, including an overview of the biotic and abiotic factors influencing their content in seaweed and the effect of processing stages. Most studies refer to crude extracts, with different components that may act separately or synergistically to exert antimicrobial activity.
4.1. Factors Affecting the Content of Antimicrobials in Seaweed
The chemical composition of the algae and the antimicrobial activities vary with species, physiological status, the region of the thallus, environmental aspects (climate, location, salinity, temperature), pollution, growth conditions, collection time and epiphytic organisms [63,64,65].
Different studies confirmed the variation in chemical compositions and the antimicrobial action according to season. Most authors detected maximal antimicrobial potential in spring, probably due to the predominance of some active compounds in this period [53,66,67,68]. However, a different variation of the phenolic composition and antibacterial/antioxidant activities was observed, with the highest inhibitory activity in spring and summer, and phenolic content and antioxidant activity in late winter [64]. Other studies have shown a lack of activity during some seasons [68], no clear seasonal variation of antifungal [66] and antibacterial activities [55], or a different pattern among algal classes, whereas Phaeophyceae showed an absence of activity in certain seasons, Rhodophyceae showed variations along seasons and Chlorophyceae were active throughout the year [69]. The effect of the latitude and related environmental factors on the phenolic content and phlorethol type in Sargassum muticum collected in different European coasts was reported [70].
Seaweed-associated microbial communities are highly diverse and rich sources of exceptional molecular structures [71]. Over a long evolutionary period, the marine organisms sharing a common environment have established associations [13]. Bacteria present either outside or inside the algal cells protect the host against pathogens by producing bioactive compounds. Despite epiphytic microorganisms showing high antimicrobial activity [71,72,73], they are not the object of this work. In most mentioned studies, epiphytic bacteria have been removed by washing.
4.2. Extraction of Antimicrobials from Seaweed
The bioactives yield can be affected by the method of conditioning and extraction. The drying stage is important, since loss of volatile antimicrobials present in fresh algae (hydrogen peroxide, terpenoid and bromo-ether compounds and volatile fatty-acids) could occur at high temperatures [74]. These authors observed increased permeability of cell membranes and reported that extracts from dried algae processed at high temperatures presented wider inhibition zones for Salmonella enteritidis, Pseudomonas aeruginosa, Listeria innocua, and on both clinical and food isolates of Staphylococcus aureus. Similarly, Chambers et al. [75] reported that crude ethanol extracts of dried Chondrus crispus had a lower MIC against the growth of bacteria and phytoplankton species than the extracts from the fresh seaweed. Cox et al. [76] reported a reduction of the total phenolic content and antioxidants from H. elongata during hydrothermal processing and drying, respectively. Drying followed by boiling increased the phytochemicals content and enhanced the inhibitory potential.
The fermentation of an Eisenia bicyclis broth with Candida utilis YM-1 during one day enhanced the phenolic content (eckol, dieckol, dioxinodehydroeckol, and phlorofucofuroeckol-A), and the antimicrobial activity against methicillin-resistant S. aureus and food-borne pathogenic bacteria [77].
Cell wall and storage polysaccharides from green, brown and red seaweed, including ulvans, alginates, fucans, laminarin and carrageenans can trigger defense responses in plants against pathogens. The oligosaccharides obtained by depolymerization of seaweed polysaccharides also induce protection against viral, fungal and bacterial infections in plants, leading to the accumulation of proteins and compounds with antimicrobial activities [22].
Choi et al. [67] reported that irradiation with Co-60 gamma rays at 1–30 kGy increased the Hunter color L value of Ecklonia cava ethanol extracts in a dose-dependent manner, without altering the antimicrobial activity, but the a and b values and the antioxidant properties decreased with irradiation. An acute toxicity test using mice and a cytotoxicity assay on the murine macrophage cell line RAW264.7 suggested that irradiated E. cava extracts can be safely used by humans at moderate doses.
4.2.1. Solvent Extraction
Various methods have been proposed to extract the bioactive compounds from seaweed, using organic solvents. The yield of extractables and antimicrobials from the different species of seaweed is solvent dependent. Systematical evaluation and optimization of the solvent is necessary for accurate and reproducible preparation of extracts. Several studies aimed at selecting the best solvent, which was usually one of the following: water, methanol, ethanol, acetone, ethyl acetate, dichloromethane, chloroform, diethyl ether and hexane [58,68,78,79,80,81]. Different authors have compared the different families of seaweed regarding antibacterial activity [6,69,82,83,84]. Table 1, Table 2 and Table 3 present some examples of the microorganisms inhibited by solvent extracts from red, green and brown seaweed, respectively.
In most cases, the extracts produced using organic solvents appear more efficient [14,79], probably because the inhibition mechanisms are due in part to the hydrophobic nature of some components, such as fatty products [3,106]. Other works confirmed that polar extracts have higher antibacterial activity [98,113,114,115]. Abundant studies have confirmed that alcoholic solutions and/or hydrophilic solvent mixtures provided better activity, i.e., methanol and acetone extracts were more active than those in lipophilic solvents as chloroform/methanol [68,74]. Krish and Das [68] reported that the ethanol and methanol extracts from Cladophora rupestris were more active than the ethyl acetate ones, but all of them were less potent than ampicillin. Methanol was found to be more effective in a number of studies [55,66,84,98,113,116,117], but also ethanol [80], acetone [78,82,101], water [98], ethyl ether [3] and ethyl acetate [74,118] were tried. Rajauria et al. [107] found considerable variations in the extraction yield and antimicrobial activity among different concentrations of methanol, 60% methanol providing optimal performance from H. elongata. However, the optimal solvent depends on many factors, particularly on the target solutes and microorganisms, and a general recommendation is not possible, i.e., Cox et al. [6] selected methanol for brown and acetone for red and green seaweed species, whereas Shanmugam et al. [119] selected methanol for red and for brown seaweed, and acetone was better for green species.
The intensification of the extraction stage can reduce the time and energy demand. Patra et al. [120] proposed microwave-assisted hydrodistillation to extract the essential oil from Enteromorpha linza. The major active compounds, such as n-hexanal, 2-chloro-5-methyl-4-(2-thienyl) pyrimidine, tetradecanoic acid, pentadecanoic acid, (z,z)-6,9-cis-3,4-epoxy-nonadecadiene, 13-octadecenal and azetidine, individually or synergistically inhibited the growth of pathogenic bacteria. Kadam et al. [121] reported the beneficial effect of ultrasound assisted extraction on the laminarin yields from Ascophyllum nodosum and Laminarina hyperborea. The purified extracts have shown inhibition of Staphylococcus aureus, Listeria monocytogenes, E.coli and Salmonella typhimurium.
4.2.2. Alternative Solvents
In addition to conventional solvents, alternative green technologies have been proposed for the extraction of algal components [122]. Pressurized solvents, i.e., supercritical carbon dioxide (SC-CO2) or subcritical water extraction, are considered green, environmentally friendly methods. Plaza et al. [106] proposed the use of pressurized solvent extraction of H. elongata with hexane, ethanol and water and did not observe that increased temperatures significantly chaged the antimicrobial activity of the extracts, mainly consisting of fatty acids (palmitoleic and oleic acids), sterols, alkanes and phenols. Boisvert et al. [56] used ethanol under pressurized liquid conditions to extract antimicrobials form brown and green algae.
Sivagnanam et al. [112] proposed the supercritical (SC)-CO2 (45 °C, 250 bar) extraction using ethanol as a co-solvent to produce extracts with phenolic content and antioxidant properties higher than those with conventional solvents from brown seaweed (Saccharina japonica and Sargassum horneri). However, the acetone-methanol extracts exhibited better antimicrobial activities.
Meillisa et al. [111] proposed subcritical water hydrolysis (200–280 °C, 1.3–6.0 MPa) to extract antibacterial compounds from S. japonica, previously de-oiled by SC-CO2 extraction. Strong antibacterial activity against two Gram-negative (E. coli and S. typhimurium) and two Gram-positive bacteria (S. aureus and B. cereus) was found using acetic acid to aid in the hydrolysis. The MIC values ranged from 1.60 to 3.50 mg/mL, and the optimal hydrolysis temperatures were 240 °C against B. cereus and S. typhimurium and 280 °C against E. coli and S. aureus. Since acetic acid had antibacterial activity at 1%, and only 0.48%–0.6% acetic acid remained in the hydrolysate, the addition of acetic acid in the hydrolysis process can improve the extraction of antibacterial substances from the alga.
4.2.3. Sequential Extraction and Purification
A series of extraction stages has been proposed either from the algal material or as a purification strategy of crude extracts. Examples of the first alternative include a sequential extraction of H. elongata with a mixture of low polarity solvents (chloroform, diethyl ether and n-hexane) followed by the extraction of the residue with 60% aqueous methanol and the pooled crude extract was further partitioned using water and ethyl acetate [54]. Amorim et al. [90] repeated hot aqueous extraction on the raffinate to obtain a crude sulfated polysaccharide from G. ornate.
Crude extracts have been purified or refined in bioactive-guided fractionation processes, including sequential solvent extraction and/or chromatography. Águila-Ramírez et al. [3] purified crude acetone/methanol extracts with ethyl ether and with butanol. Jaswir et al. [81] fractionated a methanolic extract of Sargassum plagyophillum using solvents of increasing polarity. Kim et al. [123] fractionated an 80% ethanol crude extract of S. muticum with n-hexane, dichloromethane, ethylacetate, and butanol.
De Felício et al. [124] partitioned Bostrychia tenella extracts using n-hexane and dichloromethane and further choromatographic fractionation. Park et al. [102] reported the fractionation of the 4:1 methanol:water extract from E. linza, using chloroform, Sephadex LH-20 gels and reverse-phase HPLC (high-performance liquid chromatography) using a C18 column to yield pure compounds. Ismail et al. [125] fractionationated aqueous extracts from Zonaria tournefortii by flash chromatography in a C18 column. Kavita et al. [83] fractionated a L. papillosa extract using silica gel chromatography with a solvent mixture of increasing polarity. Rodrigues et al. [96] reported the fractionation of dichloromethane extracts of Sphaerococcus coronopifolius, using eluents of increasing polarities in normal phase vacuum liquid chromatography on silica. Bromoditerpenes were the active compounds in fractions showing growth inhibition against S. aureus and C. albicans.
5. Applications
5.1. Biofouling
All surfaces immersed in the marine environment are susceptible to being colonized by biofouling communities, becoming one of the biggest problems faced by the shipping companies. Since marine seaweed thalli contain less epibionts compared to biofilms on inanimate substrata, probably because they produce secondary metabolites which prevent bacterial attachment and growth. Several species of seaweed exhibit different patterns of production of bioactive compounds depending on the seasonal variation of the fouling pressure, and a higher production in spring and summer was reported, corresponding to the algae and invertebrates spawning season, and maximal values for water temperature and light intensity [2,3]. In the past, paints were formulated with toxic compounds such as As, Hg or TBT (tributyltin). Consciousness of the marine pollution, and its effects on both the food-chain and the genetic mutations of exposed animals, led to increased restrictions on the use of toxic biocides in the manufacture of antifouling products. Alternative solutions based on antifoulants with broad-spectrum activity and low toxicity to non-target organisms, stability in a paint formulation, and commercial availability are sought [3,75]. Marine macroalgae could be an interesting source, since fatty acids, lipopeptides, amides, alkaloids, terpenoids, lactones, pyrroles and steroids could be active antifoulants [31]. Barreto and Meyer [126] reported the isolation of lanosol ethyl ether from Osmundaria serrata and its antimicrobial activity against various terrestrial bacteria and fungi, and marine bacteria isolated from the surface of the seaweed. Vairappan et al. [127] isolated brominated sesquiterpenes, acethylmajapolenes, halogenated sesquiterpenes, some halogenated acetogenins and bromoalenes from Laurencia sp. These compounds displayed antibacterial activity against some marine bacterial strains. Sesquiterpene-type compounds could be the active compounds in ethyl ether extract of Laurencia johnstonii, which showed inhibition zones comparable to those obtained with erythromycin [3].
The antifouling activity can be evaluated against bacteria, fungi, diatoms, macroalgal spores, mussel phenoloxidase activity, and barnacles and testing under real world conditions to predict performance, i.e., in static docks, is recommended. The seasonal and geographical variations of shipping paths should be considered for the formulation of antifoulants effective against a broad spectrum of organisms and marine environments.
Hellio et al. [58] found high levels of in vitro antifouling activity of aqueous, ethanolic and dichloromethane extracts from 30 marine algae from Brittany coast (France) against 35 isolates of marine bacteria. About 20% of the extracts were found to be active and non-toxic to oyster and sea urchin larvae and mouse fibroblasts. Hellio et al. [2] reported that dichloromethane extracts of Rhodophyceae were the most active among antifouling agents against different macroalgal species from the Bay of Concarneau (France).
Águila-Ramírez et al. [3] tested six seaweed extracts against the growth of selected species of marine colonisers (bacteria, fungi and microalgae). The ether extracts of Laurencia johnstonii, Ulva lactuca and Dictyota flabellata were more active in comparison with buthanol extracts against S. aureus. The most potent against the marine microalgae Rhodosorus magnei, Neorhodella cyanea and Prymnesium calathiferum were U. lactuca and L. johnstonii (MIC: 0.1–1 µg/mL) and C. fragile (MIC: 1–10 µg/mL).
Ben Redjem et al. [128] compared the organic extracts of eleven species of brown macroalgae collected from northern coasts of Tunisia and reported the highest antibacterial activity of the Cutleriales and Dictyotales, and the best inhibition against the diatom Chaetoceros calcarans for Scytosiphonales. Mathan et al. [129] reported that acetone/methanol extracts further fractionated with ethyl ether; with butanol, they were active against Halomonas marina, Polaribacter irgensii, Pseudoalteromonas elyakovii, Rosevarius tolerans, Vibrio aestuarianus, V. anguillarum and V. pomeroyri.
Some studies confirmed the lack of toxicity of algal extracts. Bazes et al. [57] observed antifouling activity of extracts from Ceramium botryocarpum on well plates against bacteria associated with immersed surfaces: Vibrio sp. and Pseudovibrio denitrificans. Chambers et al. [75] evaluated the antifoulant potential of Chondrus crispus on marine bacterial, phytoplankton and macroalgae strains. The extract from dried seaweed was more active than the fresh extract, and the longer shelf life of a dry product might be preferable. Macroalgae tests confirmed the anti-germination action against U. pinnatifida and U. intestinalis spores and a test panel sea exposure at docks, and confirmed its activity for six weeks and better performance than the same paint containing chlorothalonil. However, in open sea trials the extract showed less fouling coverage than chlorothalonil.
5.2. Aquaculture
Diseases of microbial origin are responsible for high mortality rates and lesions on fish skin, with important economic losses in aquaculture. According to the review by Vatsos and Rebours [14], in the last 20 years, there has been an increasing interest in various seaweed extracts as prophylactic and/or therapeutic agents, namely, red and brown seaweed, those exhibiting significant antimicrobial properties against fish and shrimp.
Vibriosis is a common disease affecting warm and cold water fish and seafood species caused by bacteria of the genus Vibrio, such as Vibrio anguillarum, V. ordalii, V. harveyi, V. vulnificus, V. parahaemolyticus, V. alginolyticus, V. salmonicida, which can accumulate in the fish flesh. Treatment with commercial antibiotics can cause toxicity to the animals and the release of chemical residues into the environment. Some Vibrio strains, including V. harveyi, V. parahaemolyticus and V. splendidus, are resistant to several antibiotics. Cavallo et al. [45] observed different susceptibilities of the chloroform/methanol extracts of red and green algae on Vibrio species using the disc diffusion method. G. longissima offered the broadest antibacterial spectrum, showing activity against V. ordalii, V. salmonicida, V. alginolyticus and V. vulnificus, followed by Cladophora rupestris extract, active against three species, and Chaetomorpha linum and G. dura extracts against two species, but none of the extracts tested demonstrated inhibition against V. splendidus and V. harveyi.
Mendes et al. [74] characterized the antimicrobial activity of solvent extracts from Gracilaria vermiculophylla, Porphyra dioica and C. crispus, both from wild and from integrated multi-trophic aquaculture. The higher potency in extracts from aquaculture species, when compared with the wild ones may be due to the environmental conditions, such as the presence of larger concentration of compounds from the breeding tanks, the constant water motion and aeration, and to the exposure to higher light intensities during longer periods of time. Aquaculture extracts of G. vermiculophylla and P. dioica presented a higher content of fatty acids. The ethyl acetate extracts predominated saturated fatty acids, especially palmitic acid, followed by polyunsaturated and monounsaturated fatty acids.
Bansemir et al. [47] screened, via agar diffusion disk assay, dichloromethane, methanol and water extracts of cultivated seaweed for their activities against Aeromonas salmonicida ssp. salmonicida, A. hydrophila ssp. hydrophila, Yersinia ruckeri, Pseudomonas anguilliseptica and Vibrio anguillarum. These latter two were the more susceptible strains to dichloromethane extracts of Asparagopsis armata, Ceramium rubrum, Drachiella minuta, Falkenbergia rufolanosa, Gracilaria cornea and Halopitys incurvus. Crude extract of Asparagopsis sp. also showed in vivo antibacterial activity against shrimp Vibrio pathogens [62].
Dubber and Harder [55] investigated antibacterial effects of hexane and methanol extracts of Mastocarpus stellatus, Laminaria digitata and Ceramium rubrum on fish pathogenic bacteria. Gram-positive Bacillaceae were generally more susceptible than Gram-negative Vibrionaceae, Listonella anguillarum, Pseudomonas anguilliseptica and Aeromonas salmonicida.
Cavallo et al. [45] confirmed the activity of chloroform/methanol extracts against some pathogenic Vibrio species using the disk diffusion method. The best candidates were G. longissima and C. linum, active against V. alginolyticus, V. vulnificus and V. ordalii.
Cortés et al. [130] evaluated the antimicrobial activity of Ceramium rubrum from Chile on the bacteria Yersinia ruckeri and the oomycete Saprolegnia parasitica, causing enteric red mouth disease and saprolegniasis, respectively. The ethanol and dichloromethane extracts were effective against S. parasitica using the agar dilution method, and MIC values were determined by the broth dilution method. The whole extract was more active than the individual components, suggesting a synergistic effect among the components.
García-Bueno et al. [53] observed by the microplate method that water extracts of Grateloupia turuturu, from the French Atlantic coast, were active against the abalone pathogen V. harveyi strain ORM4. However, extracts from Palmaria palmata collected in the same coast were inactive.
Krish and Das [68] reported that the crude methanol, ethanol and ethyl acetate extracts of Cladophora rupestris collected in a Mediterranean area, showed good antimicrobial activity against V. harveyii, V. parahaemolytical and V. alginolyticus, which were measured with the agar diffusion method and the zone of inhibition. The fatty acid profile showed palmitic, myristic, oleic, alpha linolenic, palmitoleic and linoleic acids. Although scarcer, studies with other algal components have been reported. Tanniou et al. [70] tested the antibacterial activity of phenolic products from Sargasum against Vibrio aestuarianus, V. anguillarum and V. parahaemolyticus.
In addition to in vitro screenings, studies have considered the antimicrobial effects incorporating the seaweed extracts either in the feeds or added into the water in which the fish and shrimp were reared. It has been suggested that the direct addition of antimicrobial seaweed substances into the water culture led to interferences between pathogens, rather than their growth. In their recent review, Vatsos and Rebours [14] emphasized on the need of pharmacodynamics and pharmacokinetics studies and also on the need of studies confirming their applicability on a commercial scale.
Thanigaivel et al. [50] confirmed that the ethanol extracts of Chaetomorpha antennina in vitro by the agar well diffusion method and in vivo on shrimps infected with Vibrio parahaemolyticus. The ethanol extract of C. antennina was very effective in controlling V. parahaemolyticus, which is resistant to ampicillin and sensitive to erythromycin. Thanigaivel et al. [9,50,51,131] confirmed in vitro the antibacterial activity of aqueous and ethanol extracts of Padina gymnospora, Gracilaria folifera, Sargassum cinereum and S. longifolium via the agar well diffusion method and in vivo in fingerlings of Oreochromis mossambicus (tilapia) infected with Pseudomonas aeruginosa and detection by PCR.
Oliveira et al. [132] performed a 40-day experiment consisting of the inclusion of Ascophyllum nodosum meals at a dose of 20 g/kg on the diet of Nile tilapia inoculated with Aeromonas hydrophila. The width was greater for the treatment with the algal meal, but there was no influence on the performance parameters of the fingerlings. The occurrence of lesions in animals inoculated with A. hydrophila and fed with the alga was lower and declined in a shorter period of time than in the control group; prevention of hepatopancreatic congestion in infected animals was also observed.
5.3. Human Health
5.3.1. Antibacterial and Antifungical
The potential of seaweed as a source of compounds active against pathogenic microorganisms has been confirmed in different studies. Padmakumar and Ayyakkannu [69] screened 80 species against bacterial and fungal pathogens. Of the algae, 70% exhibited antibacterial activity but only 27.5% showed antifungal activity. Among the species tested, S. aureus, Vibrio spp and Trichophyton mentagrophytes were the most susceptible, whereas P. aeruginosa and Aspergillus flavus were the most resistant.
Freile-Peligrin and Morales [63] evaluated the activity against Gram-positive, Gram-negative and fungus of the ethanolic and lipid-soluble extracts from seaweed species from the coast of Yucatan, Mexico, and found that more than 80% species were active against the bacteria Gram-positive tested. Ely et al. [42] tested methanolic extracts of Indian seaweed against clinical bacteria and fungi, using paper disk assays. Stoechospermum marginatum extract was effective only against Vibrio cholerae and Cladophora prolifera against V. cholerae, Klebsiella sp., S. aureus and Aspergillus niger.
Kim et al. [123] reported antimicrobial activitiy of crude extracts and solvent fractions from Sargassum muticum, especially the dichloromethane fraction against B. subtilis, Listeria monocytogenes, S. aureus, E. coli, Salmonella enteritidis and P. aeruginosa.
Dhanya et al. [133] reported the antimicrobial activity of extracts from Ulva reticulata against Staphylococcus aureus, Escherichia coli, Pseudomonas aeruginosa, Salmonella typhi, and Bacillus subtilis.
Kim et al. [134] reported the in vitro broad-spectrum antimicrobial activity of ethyl-ether extracts of U. lactuca against methicillin-resistant S. aureus (MRSA), but not antifungal activity against C. albicans. Muñoz-Ochoa et al. [86] screened sixty ethanol extracts of marine flora of Baja California Sur (Mexico) to evaluate the reversing effect of the bacterial resistance to antibiotics in combination with a sublethal concentration of ampicillin or erythromycin. Thirthy five of the assayed extracts showed inhibitory activity against S. aureus, 48 were active against Streptococcus pyogenes, but none were active against E. coli. The most active extracts were from Laurencia spp., Gelidium robustum, Chnoospora implexa, Padina mexicana, Gracilaria subsecundata and Dictyopteris undulata.
Oh et al. [135] tested for antimicrobial activity against bacteria and fungi human pathogens the bromophenols isolated from the red alga Odonthalia corymbifera, and found that natural products were more active against fungi and synthetic bromophenols against bacteria. Shanmughapriya et al. [52] tested 40 different seaweeds; of these, seven species showed high antimicrobial activity against multirresistant pathogens.
Osman et al. [82] observed the antimicrobial activities of crude extracts from the species of Rhodophyta, Chlorophyta and Phaeophyta against B. subtilis, S. aureus, E. coli, Klebsiella pneumoniae and Candida albicans. Taskin et al. [92] reported inhibition of C. albicans growth by a methanolic extract from Cystoseira mediterranea.
Manilal et al. [88] tested the crude methanolic extracts of marine organisms against multiresistant human pathogens. They found that the Falkenbergia-phase of Asparagopsis taxiformis was highly active, and the most abundant metabolites were oleic acid and n-hexadecanoic acid.
Pierre et al. [98] reported Chaetomorpha aerea extracts in methanol and in water exhibited in vitro antimicrobial activity against three Gram-positive bacteria. Salem et al. [118] screened methanolic and ethyl acetate extracts from eight different seaweeds collected from Hurghada, a city on the Red Sea coast in Egypt, for their antibacterial activities, evaluating the zone of inhibition and MIC values. Higher susceptibility of Gram-positive bacteria to the algal extracts was observed, and ethyl acetate extract of C. racemosa was the most inhibitory extract.
Adaikalaraj et al. [113] evaluated the antibacterial activity of methanol and aqueous extracts from Gracilaria verrucosa, G. ferugosoni, Hypnea musciformis, Enatiocladia prolifera and Gelidium species against selected bacterial pathogens and one yeast by using the disc diffusion method. In most of the seaweeds, methanol extract was found to be more effective. Salmonella typhi, S. aureus, B. subtilis and C. albicans were resistant to all aqueous extracts. Águila-Ramírez et al. [3] confirmed that ether extracts of L. johnstonii, D. flabellata and U. lactuca from the Gulf of California showed activity against Staphylococcus aureus, but only the buthanol fraction from D. flabellata was active. No activity was observed against E. coli, Bacillus cereus, B. subtilis and Staphylococcus epidimidis. Amorim et al. [90] determined the effect on bacterial growth of a crude sulfated polysaccharide from G. ornate and fractions extracted at different temperatures, showing low contents of sulfate (5.88%–10.3%) and proteins (0.1%–3.7%). Go-3 was tested on the growth of bacteria (B.subtilis, S.aureus, Enterobacter aerogens, E. coli, P. aeruginosa, Salmonela choleraesuis and S. typhi), but only E. coli was inhibited. Rosaline et al. [78] screened the antibacterial efficacy of extracts in hexane, ethyl acetate, acetone and methanol of Sargassum wightii, Chaetomorpha linum and Padina gymnospora against selected Gram-positive and Gram-negative human pathogenic bacteria. The acetone extracts of the three algae showed higher inhibitory activity for the selected bacterial species than other solvent extracts. P. gymnospora and S. wightii were more active than C. linum against human pathogenic bacteria using the disc diffusion method. Águila-Ramírez et al. [3] reported antibiosis of the extracts from L. johnstonii, U. lactuca and D. flabellata. The ether extracts were more active in comparison with butanol extracts against S. aureus. L. johnstonii extracts showed good activity against E. coli. Mhadhebi et al. [46] screened 24 organic fractions of 6 seaweeds from the Tunisian Mediterranean coast against 8 human pathogenic bacteria and 5 Candida pathogenic strains. All the extracts exhibited moderate to weak activity against S. aureus; S. epidermis, E. coli and Micrococcus luteus. The chloroform and the ethyl acetate extracts from Cystoseira crinita and C. sedoides showed a higher antifungal activity against four Candida strains.
Bianco et al. [94] evaluated the in vitro anti-infective potential of organic extracts from different marine organisms, including seaweed collected along the Brazilian coast, and found that Osmundaria obtusiloba showed moderate activity against P. aeruginosa. Genovese et al. [136] confirmed the antifungal activity of Asparagopsis taxiformis, from the Straits of Messina (Italy), against Aspergillus fumigatus, A. terreus and A. flavus. The lowest MIC observed were <0.15 mg/mL, and the highest were >5 mg/mL. Jassbi et al. [114] reported antioxidant and antifungal activity in the water extracts of a red algae, Hypnea flagelliformis, and two brown algae, Cystoseira myrica and Sargassum boveanum, from the Persian Gulf. The active substances against S. aureus and B. subtilis using the agar disc diffusion and nutrient broth micro-dilution bioassays were identified as free fatty acids, fucosterol, cholesterol and 22-dehydroxychlosterol. Mathan et al. [129] assayed acetone/methanol extracts further fractionated with ethyl ether and with butanol, in an agar diffusion test against human pathogenic bacteria: E. coli, S. aureus, B. cereus, B. subtilis and S. epidermidis. Mohandass et al. [137] proposed the use of Sargassum cinereum extracts as a reducing agent in the extracellular synthesis of silver nanoparticles. The MIC against Staphylococcus aureus was 2.5 μL (25 μg/disc), and against Enterobacter aerogenes, S. typhi and Proteus vulgaris were 100 μg/disc. Osman et al. [138] tested macroalgae collected from Egypt against pathogenic Gram-positive and Gram-negative bacteria, and one clinical yeast strain, Candida albicans. The tested species of Chlorophyta were more potent inhibitors than those from Rhodophyta and Phaeophyta. The extract of U. fasciata was the most active (phthalate esters derivatives being the active components), followed by E. compressa, U. lactuca and E. linza; K. pneumoniae was the most sensitive microorganism. Saritha et al. [79] reported antibacterial activity of U. lactuca extracts against Shigella sonnei, B. subtilis, E. feacalis, S. typhimurium, E. coli, S. aureus, S. pyogenes and Staphylococcus epidermis.
Al Hazzani et al. [84] reported higher in vitro antimicrobial activity of methanol and acetone extracts from the brown algae (L. japonica, U. pinnatifida, E. bicyclis) than from the red alga P. tenera against Gram-positive and Gram-negative bacteria, some were antibiotic-resistant such as methicillin-resistant S. aureus and P. aeruginosa, and, resistant against yeast, C. albicans. Horincar et al. [85] studied macroalgae from the Romanian Black Sea coast for their volatile compounds content. The major components in the extract of Ceramium virgatum were 3-hexen-2-one, acetone, hexanal and o-cymene, in that of Cladophora vagabunda were hexanal, octane, nonanal, octanal, 2,5,5-trimethyl-2-hexene, 3-hexen-2-one, and o-cymene and in that of U. intestinalis were hexanal, trichloromethane, nonanal, 3-hexen-2-one, and octanal, and had a greater content of mono- and polyunsaturated fatty acids, palmitic acid, arachidonic acid, and oleic acid. The lipid fatty acids affected the biological activity against S. enteritidis, E. coli, Listeria monocytogenes, B. cereus, and differences among algal families were observed, i.e., the total lipids from Laminariaceae were more active antimicrobial than those from Alariaceae [139]. Ismail et al. [125] reported the antibacterial and antifungal activities of aqueous extracts and fractions of Zonaria tournefortii against C. albicans, Cryptococcus neoformans, and several multi-resistant S. aureus. Jaswir et al. [81] produced crude extracts from four species of brown seaweed (Sargassum plagyophillum, S. flavellum, S. binderi and Padina australis) from Malaysia. Crude extracts showed no antimicrobial activity against the Gram-negative bacteria (P. aeruginosa and E. coli), whereas the Gram-positive bacteria (B. subtilis and S. aureus) were inhibited. Kavita et al. [83] screened the methanol extract of 38 seaweed samples against human pathogenic bacteria, and Gram-positive (S. aureus and B. subtilis) and Gram-negative (E. coli and P. aeruginosa) bacteria. Laurencia papillosa showed maximum antimicrobial activity, and the active fraction was identified as a cholesterol derivative, 24-propylidene cholest-5-en-3 beta-ol. The MIC values against clinical isolates ranged from 1.2 to 1.7 µg/mL. Krish and Das [68] reported that the crude methanol, ethanol and ethyl acetate extract of Cladophora rupestris at 250–1000 µg/mL showed concentration-dependent antimicrobial activity against several human pathogenic bacteria, including E. coli, P. aeruginosa and S. aureus. The activity was evaluated with the zone of inhibition, which was compared to that of amphicilin. Whereas the methanol extract was more active against E. coli, the ethyl acetate was a greater inhibitor of S. aureus and the ethanol extract against S. aureus and P. aeruginosa. Nogueira et al. [110] reported modulation of antibiotic activity between the Padina sanctae-crucis ethanolic and methanolic extracts and E. coli and S. aureus, and a moderate modulatory effect against these microorganisms and P. aeruginosa, Candida tropicalis and C. kruseim. Stabili et al. [65] confirmed the activity against human pathogenic bacteria and yeasts of lipid extracts of one Mediterranean alga with the difussion disk method. Tanniou et al. [70] observed the antibacterial activity of phenolics from Sargasum collected in different countries against E. coli, S. aureus and P. aeruginosa.
El Wahidi et al. [140] reported the antimicrobial activity of dichloromethane and ethanol extracts from seaweeds collected from the Moroccan’s Atlantic coast against 2 Gram-positive (B. subtilis and S. aureus) and 2 Gram-negative (E. coli and P. aeruginosa) bacteria, and against two pathogenic yeasts (Candida albicans and Cryptococcus neoformans) using the agar disk-diffusion method. Cystoseira brachycarpa, C. compressa, Fucus vesiculosus, and Gelidium sesquipedale showed better antimicrobial activity with a broad spectrum.
Karthikeyan et al. [80] tested the antibacterial activity for organic solvent extracts from seaweeds from Kodinar coast in India: Enteromorpha sp., Cystoseria indica, Sargassum swartzii, Gracilaria corticata, Caulerpa taxifolia and C. racemosa against E. coli, Proteus sp., P. aeruginosa, Klebsiella pneumoniae and S. aureus. The maximum antibacterial activity was observed in the ethanol extracts of all the seaweeds except C. racemosa. Kolsi et al. [44] evaluated the antibacterial and antifungical activities of organic extracts (hexane, ethylacetate and methanol) of 13 seaweed species from the Tunisian coastline against 8 Gram-negative and Gram-positive human pathogens, yeast and a fungi in an agar disk diffusion test. They found that brown algae showed higher inhibitory activity on bacteria than on fungi. Kosanic et al. [101] reported that the acetone extracts of U. lactuca and E. intestinalis showed antimicrobial and cytotoxic activity on four human cancer cell lines. The extracts of U. lactuca showed MIC values ranging from 0.156 to 5 mg/mL, but it was relatively weak in comparison to standard antibiotics. Bacillus mycoides and B. subtilis were the most susceptible to the tested extracts. Lopes et al. [141] proposed phlorotannin extracts due to their antifungal activity against several yeast and dermatophyte strains, using a micromethod for the evaluation of the MIC and the MLC. Pinteus et al. [43] evaluated the methanol, n-hexane and dichloromethane extracts of macroalgae (Rhodophyta, Chlorophyta and Heterokontophyta divisions) from Peniche coast (Portugal) for antibacterial activity, evaluated by the disc diffusion method against B. subtilis and E. coli. The antifungal potency of the Sphaerococcus coronopifolius extracts, evaluated against Saccharomyces cerevisiae as a model, was similar to the standard amphotericin B. Asparagopsis armata and Sphaerococcus coronopifolius revealed the most potent antimicrobials. Wei et al. [142] found that an anthraquinone dihydroxy derivative, 1,8-dihydroxy-anthraquinone, first isolated from Porphyra haitanensis, showed antibacterial activity against S. aureus, inhibiting cell growth at logarithmic phase due to its interaction with the cell wall and cell membrane, leading to increased permeability, leakage of cytoplasm and the deconstruction of cell.
Patra et al. [103] reported that Enteromorpha linza L. essential oil, containing high amounts of acids (54.6%) and alkenes (21.1%), was effective against both E. coli and S. typhimurium. MIC and MBC values for both pathogens were 12.5 mg/mL and 25.0 mg/mL, respectively. Essential oil induced a bactericidal effect via structural membrane damage caused by its deposition in the cytosol or through enzymatic degradation of bacterial intracellular enzymes that resulted in cellular lysis.
Spavieri et al. [59] tested the antimycobacterial activity of crude extracts of 21 species of brown algae from British and Irish waters but only the Bifurcaria bifurcata extract was weakly active against Mycobacterium tuberculosis.
Marine algae could be a source of therapeutic agents for chronic gastritis and peptic ulceration. Lee et al. [143] screened 27 Korean species of seaweed for potential anti-Helicobacter pylori activity, and seven showed strong inhibitory activity based on the agar diffusion method. The strongest activity was observed for ethanol extracts from Ishige okamurae. The inhibition zone of this extract was 9.0 mm at 1 mg/disk, the MIC was 12 µg/mL based on the broth microdilution assay, and the free urease assay confirmed that the 80% methanol extracts had 75.4% inhibition at 0.1 mg/mL. Both the I. okamurae phenolic compounds and the nitrogen compounds of the extract significantly inhibited H. pylori. No toxicity was observed in a study with BALB/c mice at a dose of 5 g/kg body weight.
Ha et al. [100] screened 44 seaweed species for anti-Gardnerella vaginalis activity and 27 of them showed antimicrobial activity by the agar disk-diffusion method. Ulva pertusa exhibited the highest activity against the strains of G. vaginalis causing vaginosis (MIC = 312 µg/mL), and against Candida albicans, the main cause of candidiasis (MIC = 2.5 mg/mL). Nitrogenous compounds were the main active agents against G. vaginalis. No inhibitory effect against Lactobacillus gasseri and Lactobacillus jensenii, present in the normal vaginal microflora, were observed at 10 mg/mL.
Vedhagiri et al. [144] evaluated compounds isolated from Asparagopsis taxiformis for their inhibitory action against Leptospira javanica isolated from rodent carriers and found MICs and MBC in the range of 100–1600 μg/mL, whereas those for penicillin and doxycyline were in the range of 25–200 μg/mL. The GC-MS analysis revealed the presence of 4,5-dimethyl-1H-pyrrole-2-carboxylic acid ethyl ester, fatty acids, 14-methyl-pentadecanoic acid methyl ester, octadecanoic acid methyl ester, octadec-9-enoic acid 2,3-dihydroxy-propyl ester, 9-octadecanoic acid, methyl ester and trace amount of chlorobenzene.
Acne
Effective and safe acne vulgaris therapies could be derived from algal extracts. Amiguet et al. [105] observed that the crude ethyl acetate extracts from Fucus evanescens showed strong antibacterial activity against Propionibacterium acnes (culture collection and clinical isolate), and also against Hemophilus influenzae, Legionella pneumophila, and Streptococcus pyogenes, Clostridium difficile and methicillin-resistant S. aureus, whereas B. cereus, E. coli, K. pneumoniae, and P. aeruginosa were not significantly affected. The main active compound was identified as a β-d-galactosyl O-linked glycolipid.
Lee et al. [104] confirmed the efficacy of a crude methanolic extract from Eisenia bicyclis. The ethyl acetate fraction showed the strongest antibacterial activity, the most active compound among the six isolated from this fraction was fucofuroeckol-A, with a MIC value ranging from 32 to 128 µg/mL. This compound also reversed the erythromycin and lincomycin resistance of P. acnes. A weak synergistic effect of fucofuroeckol with erythromycin and with lincomycin was observed.
Oral Microbes
Periodontitis is a chronic inflammatory disease initiated by Gram-negative pathogens, such as Prevotella intermedia and Porphyromonas gingivalis. Traditional antibiotic therapies for periodontitis target the bacterial infection, but identification of new therapeutic agents with few or no side effects and potent antimicrobial activity is desirable.
Enteromorpha linza extracts displayed antimicrobial activity against P. intermedia and P. gingivalis without side effects at a moderate dose [145] and in a mouth rinse has shown effects similar to those of Listerine® against gingivitis. Park et al. [102] reported that the main active compounds, isolated by Sephadex LH-20 gel and reverse-phase HPLC, were unsaturated fatty acids, stearidonic acid (SA) and gamma-linolenic acid (GLA). Their MIC values were 39.06 µg/mL against P. intermedia and 9.76 µg/mL against P. gingivalis, respectively. The MIC values of pure compounds against Candida albicans were 312.50 μg SA/mL, 78.12 μg GLA/mL, 312.50 μg triclosan/mL and 4.88 μg chlorhexidine/mL, and against Streptococcus mutans were 1250 SA μg/mL, 625 μg GLA/mL and 9.76 μg chlorhexidine/mL [145].
Kim et al. [146] confirmed the antimicrobial activity of ethanol extracts of L. japonica against oral microbials. The MICs of ethanol extracts were 250 and 62.5 µg/mL against Actinomyces naeslundii and Actinomyces odontolyticus, respectively, and 250 and 62.5 µg/mL for Fusobacterium nucleatum and Porphyromonas gingivalis, respectively. The MBCs of A. naeslundii and A. odontolyticus were 500 and 250 µg/mL, respectively. A dose dependent effect and a change in the cell surface texture of Streptococcus mutans, A. odontolyticus and P. gingivalis was observed.
The topical application of products derived from n-3 fatty acids can protect against inflammation-induced tissue and bone loss associated with periodontitis in experimental models [147]. The safety of stearidonic acid was confirmed in rat and in human studies [102]. Lee et al. [148] reported, via the broth dilution method and checkerboard and time-kill assay, that fucoidan was efficient against oral bacteria, either alone or with antibiotics MIC/MBC values for the fucoidan against oral bacteria ranging between 0.125–0.50/0.25–1.00 mg/mL, for ampicillin 0.125–64/0.5–64 μg/mL and for gentamicin 2–256/4–512 μg/mL, respectively. The MIC and MBC values were reduced to one sixteenth and the rate of killing was increased when fucoidan was combined with antibiotics.
5.3.2. Antiprotozoals
De Felício et al. [124] reported that some fractions from Bostrychia tenella were more active than gentian violet and amphotericin B as antiprotozoal agents against Trypanosoma cruzi and Leishmania amazonensis. Among the volatile compounds identified were fatty acids, low molecular mass hydrocarbons, esters and steroids and also uncommon substances, such as neophytadiene. Crude extracts of 21 brown algae were screened by Spavieri et al. [59] against Trypanosoma cruzi and T. brucei rhodesiensi and Leishmania donovani. All algal extracts showed significant activity against T. brucei rhodesiensi and L. donovani, and two Sargassaceae strains were the most potent. Bianco et al. [94] reported that the organic extracts from Anadyomene saldanhae, Caulerpa cupressoides, Canistrocarpus cervicornis, Dictyota sp., Ochtodes secundiramea, and Padina sp. collected along the Brazilian coast showed promising antiprotozoal results against Leishmania braziliensis, and only Dictyota sp. was effective against T.cruzi.
5.3.3. Antivirals
Yasuhara-Bell and Lu [11] reviewed the antiviral properties of marine organisms, including those from algae, active against a wide range of viruses, including herpes viruses (HSV-1, HSV-2, HCMV), togaviruses (Sindbis virus, Semliki Forest virus), paramyxoviruses (RSV), rhabdoviruses (VSV), and both human and simian immune deficiency viruses (HIV and SIV). Extracts and different compounds from algae might be promising for future antiviral design. Extracts of Gigartina atropurpurea, Plocamium cartilagineum, Splachnidium rugosum, and Undaria pinnatifida inhibited Herpes simplex virus types 1 (HSV-1) and 2 (HSV-2) [149]. Bouhlal et al. [60] screened fifty-five aqueous, methanolic, chloroform-methanolic and dichloromethanolic extracts of Rhodophyta from the coast of Morocco for inhibitory compounds against HSV-1 by the cell viability method. The aqueous extracts of 10 seaweeds were capable of inhibiting the replication of HSV-1 in vitro at an EC50 ranging from <2.5 to 75.9 µg/mL, without any cytotoxic effect on the Vero cells in the range of the concentrations assayed.
Different chemical structures were responsible for this activity. The glycolipid sulfoquinovosyldiacylglycerol from Osmundaria obtusiloba collected from Southeastern Brazilian coast demonstrated antiviral activity against HSV-1 and HSV-2, with EC50 values of 42 μg/mL and and 12 μg/mL, respectively [10,150]. Koishi et al. [151] reported that extracts from Caulerpa racemosa, Canistrocarpus cervicornis, Padina gymnospora and Palisada perforate attenuated dengue virus infection probably by acting at an early stage of the cycle of infection, like binding or internalization.
The diterpenes 8,10,18-trihydroxy-2,6-dolabelladiene and (6R)-6-hydroxydichotoma-4,14-diene-1,17-dial, isolated from Dictyota pfaffii and D. menstrualis, respectively, inhibited HSV-1 infection in Vero cells [152]. Mendes et al. [153] reported that the extract of the marine algae Stypopodium zonale, two meroditerpenoids (atomaric acid and epitaondiol) derived from it, and a methyl ester derivative of vatomaric acid exerted antiviral activity against human metapneumovirus replication with a selectivity index of 20.78, >56.81, 49.26, and 12.82, respectively.
Lectins, polysaccharides, and tannins with anti-HIV-1 activity have been isolated from marine macroalgae [154]. The antiviral activities of sulfated polysaccharides from seaweed, their structural chemistry and their potential for therapeutic application, have been reviewed [29,61,115,155]. Marine polysaccharides can inhibit the replication of viruses through interfering with a few steps in the virus’ life cycle or improving the host immune responses to accelerate the process of viral clearance. Naesens et al. [156] reported that a 26 kDa xylogalactofucan from Sphacelaria indica, containing (1→3)-linked l-fucopyranosyl and d-galactopyranosyl residues and rich in sulfated polysaccharides, showed strong activity against the human herpevirus-6 (HHV-6). Bandyopadhyay et al. [157] reported that the 21 kDa alginate from S. indica (41% guluronic and 59% mannuronic acid) displayed IC50 values of 0.6–10 μg/mL against HSV-1, and attributed the inhibition to direct interaction of polysaccharides with viral particles. Fucoidan from Cladosiphon okamuranus was stronger than ribavirin against Newcastle disease virus in the Vero cell line at the early stages of infection, by inhibition of viral penetration [158]. Polysaccharides from Rhodophytas had major antiviral activity, whereas those from Phaeophytas showed major antitumoral and other activities diversity.
Carragenans of the red algae were active against both enveloped and non-enveloped viruses. The specific antiviral effect was related to the molecular weight and degree of sulfation, and different viruses depended on the type of carrageenan, viral serotype and the host cell. The number of necrotic lesions on Xanthinc tobacco leaves was lessened by concurrent inoculation with tobacco mosaic virus and kappa/beta-carrageenan from Tichocarpus crinitus, compared to virus-inoculated leaves [159]. Carrageenans from Meristiella gelidium were effective inhibitors of dengue virus DENV-2 if compared to G. griffithsiae samples and reference polysaccharides [160].
Jiang et al. [89] reported the in vitro antioxidant, antimicrobial and antischistosomal activities of the pethroleum ether fraction of a Grateloupia livida ethanolic extract. This fraction showed antimicrobial activity against the human pathogenic trematode Schistosoma japonicum adult worm. Neither cellular cytotoxicity at concentrations up to 100 μg/mL nor acute oral toxicity in mice at doses up to 2000 mg/kg was observed. Most of the 25 components identified have known antioxidant and antimicrobial activities.
5.4. Food
Microbial food spoilage is responsible for important economic losses due to the ubiquitous presence of microorganisms and their impact on human health. Food products also undergo autoxidation, leading to the development of rancid flavors and odors, and a reduction in the sensory attributes, nutritional quality and food safety. Replacement of synthetic additives and exploitation of new natural antioxidant and antibacterial substances with a possible role as nutraceutical agents is encouraged. Multiple functionality seaweed extracts could be incorporated into foods as natural preservatives to enhance the food quality, safety and stability [33,54,161].
Different studies confirmed the antimicrobial action of seaweed against food spoilage microorganisms. Phlorotannins are potent antioxidants and showed a strong bactericidal effect against food-borne bacteria [15]. The highest antimicrobial action is not necesarily coincident with the highest antioxidant action. Devi et al. [162] evaluated ten edible seaweeds from India for antioxidant and antimicrobial activity against food-borne pathogens. Gelidella acerosa and Haligra sp have high phenolic content and exhibited antibacterial activity against S. aureus, but G. acerosa has the highest antioxidant activity. Krish and Das [68] reported that the antioxidant and antimicrobial activity of Cladophora rupestris solvent extracts and Scytosiphon lomentaria methanol extract inhibited the Gram-negative bacterium S. typhimurium [92].
Cox et al. [6] tested Laminaria digitata, L. saccharina, Himanthalia elongata, Palmaria palmata, C. crispus and Enteromorpha spirulina from the Irish coast using a microtiter method against food spoilage and food pathogenic bacteria: L. monocytogenes, S. abony, E. faecalis and P. aeruginosa. All methanolic extracts inhibited the bacterial growth except C. crispus extracts. Brown algal extracts showed more antibacterial activity than red and green extracts, and H. elongata was the most efficient. Cox et al. [54] found that the hydrophilic extract from H. elongata provided up to 100% inhibition of S. abony and L. monocytogenes. Concentration-dependent antibacterial activity was observed in the range of 0.125–8 mg/mL, but no inhibition was detected under 1 mg/mL. The extracts showed bactericidal effect in carbohydrate model food systems and bacteriostatic effect in protein model food systems (at 1%–10%). Sodium benzoate exhibited greater inhibition at 2–0.5 mg/mL and showed bacteriostatic effect in carbohydrate medium, whereas the extract had a bactericidal effect.
Gupta et al. [117] reported that extracts from H. elongata at 6% inhibited the growth of food spoilage microorganisms (P. aeruginosa and E. faecalis) and food pathogens (L. monocytogenes and S. abony). Lower concentrations of extract prolonged the lag phase and reduced both the exponential growth rate and final population densities of the culture.
Beaulieu et al. [163] measured the antibacterial activity of a protein extract from Saccharina longicruris and from the <10 kDa and >10 kDa fractions via liquid growth inhibition assays using an optimized critical microdilution method against E. coli, Micrococcus luteus, Brochotrix thermosphacta, E. faecalis, Mycobacterium smegmatis, S. aureus, Streptococcus, P. aeruginosa, Vibrio splendidus, and V. gigantis. No activity was detected when each peptide was tested separately at 2.50 mg/mL, but significant activity was observed when mixed all together at this total peptide concentration.
Boisvert et al. [56] tested S. longicruris, Ascophyllum nodosum and U. lactuca from the St. Lawrence Estuary against E. coli, M. luteus and B. thermosphacta, determining the MIC values using a broth microdilution assay. U. lactuca extracts obtained with pressurized ethanol showed the highest antibacterial activity against the strains tested, and was therefore more effective towards Gram-negative bacteria, whereas A. nodosum was more efficient towards Gram-positive strains. The higher metal ions content in U. lactuca extracts (Al, Fe, Co, Ni, Cu, Zn, Cd, Hg, Pb) than in the other extracts could be determinant for the activity.
Moronery et al. [164] added a spray-dried Laminaria digitata extract (9.3% laminarin, 7.8% fucoidan) to minced pork patties at levels of 0.01%–0.5% (w/w). After storage in a modified atmosphere for 14 days at 4 °C, at 0.5% the extract exerted the greatest lipid pro-oxidant activity in fresh patties, whereas the lipid oxidation was decreased in cooked patties, but the microbiological status, pH, and water-holding capacity were not influenced.
Shiahaan et al. [165] prepared S. japonica as a powder (500–900 µm) for the physical adsorption and further release of allyl isothiocyanate, an inhibitor of food-borne bacteria. When the alga was deoiled, higher adsorption of allyl isothiocyanate was observed in comparison to raw alga. No loss of activity was detected against E. coli, S. typhimurium or B. cereus, and only a nominal activity against S. aureus was observed.
5.5. Other Uses
Other novel or less common applications have also been proposed based on the antimicrobial properties of seaweed extracts and compounds. Macroalgae contain compounds which could be suited for controlling specific plant pathogens. Jiménez et al. [166] studied in vitro and in vivo aqueous and ethanolic extracts obtained from nine Chilean marine macro-algae collected at different seasons for properties that reduce the growth of plant pathogens or decrease the injury severity of plant foliar tissues following pathogen infection. Manilal et al. [109] prepared methanol extracts from Lobophora variegata of the Kollam coast and confirmed that the in vitro antimicrobial activity of algal extract was investigated in Petri dishes. The extracts showed higher toxicity against bacterial pathogens than against mosquito pupae, nematodes and plant seeds.
Hierholtzer et al. [167] studied the effect of phloroglucinol and phenolic compounds extracted from Laminaria digitata on mixed microbial anaerobic organisms used for anaerobic digestion of algae. The bactericidal activity of phlorotannins is a function of their level of polymerization. Phlorotannins induce significant extra- and intra-cellular effects, with the disruption of cell membranes observed with most microorganisms.
Marine macroalgae polysaccharides could be prebiotic ingredients for both human and animal health applications. Prebiotics are non-digestible compounds that stimulate the growth and/or activity of beneficial gut microbiota which, in turn, confer health benefits on the host. Feeding with algal polysaccharides can modify the fecal and colonic population of Lactobacillus spp., Bifidobacterium and Enterobacterium [168].
Tan et al. [95] developed an antimicrobial wound dressing, consisting in a hydrogel prepared with polyvinyl alcohol, polyvinylpyrrolidone and 1% seaweed extract. It was effective against nine clinically-relevant wound pathogens as the commonly used silver-based dressing, Acticoat. The dressing, evaluated using agar diffusion tests, gel leaching and gel transfer assays, proved effective up to 97 h, without leaving a residue following five subsequent transfers. Antimicrobial activity was stable for up to 6 months of storage at 4 °C, but activity was slightly reduced after 15 weeks.
A scheme of the different stages considered in this review and proposed for the utilization of antimicrobial compounds for a variety of applications is presented in Figure 3. The advantages related to the reduction or elimination of some problems are also indicated.
6. Conclusions and Future Trends
Marine algae are one of the largest producers of biomass in the marine environment and represent a potential source of new diverse and unique compounds. Many of the substances obtained from seaweed, such as alginates, carrageenan and agar have been used for decades in traditional medicine, pharmacology and food. Other compounds have bacteriostatic or antibacterial, antiviral, antitumor, anti-inflammatory and antifouling activities. Therefore, seaweed could provide promising bioactives that can be used in the treatment of human diseases, or new antimicrobial agents to replace synthetic antibacterial agents used in agriculture and in the food industry.
Much attention has been paid to the development of innovative projects for the pharmaceutical applications of seaweed, particularly in the design of novel antimicrobial drugs. Further investigations for the identification of promising algal species, standardization of analytical methods, isolation of compounds through bioassay guided fractionation, detailed chemical characterization and evaluation of their safety, as well as the evaluation of synergistic effects among the components, and efforts to enhance the yields and lower the extraction costs, are needed.
Acknowledgments
The authors are grateful to the Ministry of Science and Innovation of Spain (Research Project Ref. CTM2012–38095, partially funded by the FEDER Program of the European Union) for the financial support of this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bhadury, P.; Wright, P.C. Exploitation of marine algae: Biogenic compounds for potential antifouling applications. Planta 2004, 219, 561–578. [Google Scholar] [CrossRef] [PubMed]
- Hellio, C.; Marechal, J.P.; Veron, B.; Bremer, G.; Clare, A.S.; Le Gal, Y. Seasonal variation of antifouling activities of marine algae from the Brittany coast (France). Mar. Biotechnol. 2004, 6, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Águila-Ramírez, R.N.; Arenas-González, A.; Hernández-Guerrero, C.J.; González-Acosta, B.; Borges-Souza, J.M.; Veron, B.; Pope, J.; Hellio, C. Antimicrobial and antifouling activities achieved by extracts of seaweeds from Gulf of California, Mexico. Hidrobiologica 2012, 22, 8–15. [Google Scholar]
- Wijesinghe, W.A.J.P.; Athukorala, Y.; Jeon, Y.J. Effect of anticoagulative sulfated polysaccharide purified from enzyme-assistant extract of a brown seaweed Ecklonia cava on Wistar rats. Carbohydr. Polym. 2011, 86, 917–921. [Google Scholar] [CrossRef]
- Damonte, E.B.; Matulewicz, M.C.; Cerezo, A.S. Sulfated seaweed polysaccharides as antiviral agents. Curr. Med. Chem. 2004, 11, 2399–2419. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar]
- Namvar, F.; Tahir, P.M.; Mohamad, R.; Mahdavi, M.; Abedi, P.; Najafi, T.F.; Rahman, H.S.; Jawaid, M. Biomedical Properties of Edible Seaweed in Cancer Therapy and Chemoprevention Trials: A Review. Nat. Prod. Commun. 2013, 8, 1811–1820. [Google Scholar] [PubMed]
- Kazlowska, K.; Hsu, T.; Hou, C.C.; Yang, W.C.; Tsai, G.J. Anti-inflammatory properties of phenolic compounds and crude extract from Porphyra dentate. J. Ethnopharmacol. 2010, 128, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Thanigaivel, S.; Hindu Vidhya, S.; Vijayakumar, S.; Mukherjee, A.; Chandrasekaran, N.; Thomas, J. Differential solvent extraction of two seaweeds and their efficacy in controlling Aeromonas salmonicida infection in Oreochromis mossambicus a novel therapeutic approach. Aquaculture 2015, 433, 56–64. [Google Scholar] [CrossRef]
- Cheung, R.C.F.; Wong, J.H.; Pan, W.L.; Chan, Y.S.; Yin, C.M.; Dan, X.L.; Wang, H.X.; Fang, E.F.; Lam, S.K.; Ngai, P.H.K.; et al. Antifungal and antiviral products of marine organisms. Appl. Microbiol. Biotechnol. 2014, 98, 3475–3494. [Google Scholar] [CrossRef] [PubMed]
- Yasuhara-Bell, J.; Lu, Y. Marine compounds and their antiviral activities. Antivir. Res. 2010, 86, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Kumari, P.; Reddy, C.R.K. Antimicrobial compounds from seaweed-associated bacteria and fungi. Appl. Microbiol. Biotechnol. 2015, 99, 1571–1586. [Google Scholar] [CrossRef] [PubMed]
- Valliappan, K.; Sun, W.; Li, Z. Marine actinobacteria associated with marine organisms and their potentials in producing pharmaceutical natural products. Appl. Microbiol. Biotechnol. 2014, 98, 7365–7377. [Google Scholar] [CrossRef] [PubMed]
- Vatsos, I.N.; Rebours, C. Seaweed extracts as antimicrobial agents in aquaculture. J. Appl. Phycol. 2015, 27, 2017–2035. [Google Scholar] [CrossRef] [Green Version]
- Eom, S.H.; Kim, Y.M.; Kim, S.K. Antimicrobial effect of phlorotannins from marine brown algae. Food Chem. Toxicol. 2012, 50, 3251–3255. [Google Scholar] [CrossRef] [PubMed]
- Abu-Ghannam, N.; Rajauria, G. Antimicrobial activity of compounds isolated from algae. In Functional Ingredients from Algae for Foods and Nutraceuticals; Domínguez, H., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 287–306. [Google Scholar]
- Fernandes, D.R.P.; de Oliveira, V.P.; Valentin, Y.Y. Seaweed biotechnology in Brazil: Six decades of studies on natural products and their antibiotic and other biological activities. J. Appl. Phycol. 2014, 26, 1923–1937. [Google Scholar] [CrossRef]
- Gouveia, V.; Seca, A.M.L.; Barreto, M.C.; Pinto, D.C.G.A. Di- and sesquiterpenoids from Cystoseira genus: Structure, intra-molecular transformations and biological activity. Mini Rev. Med. Chem. 2013, 13, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Suleria, H.A.R.; Osborne, S.; Masci, P.; Gobe, G. Marine-based nutraceuticals: An innovative trend in the food and supplement industries. Mar. Drugs 2015, 13, 6336–6351. [Google Scholar] [CrossRef] [PubMed]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Dominguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef] [PubMed]
- Usov, A.I. Chemical structures of algal polysaccharides. In Functional Ingredients from Algae for Foods and Nutraceuticals; Domínguez, H., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 23–86. [Google Scholar]
- Vera, J.; Castro, J.; González, A.; Moenne, A. Review: Seaweed polysaccharides and derived oligosaccharides stimulate defense responses and protection against pathogens in plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Kraan, S. Algal polysaccharides, novel applications and outlook. In Carbohydrates—Comprehensive Studies on Glycobiology and Glycotechnology; Chang, C.-F., Ed.; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Ahmadi, A.; Moghadamtousi, S.Z.; Abubakar, S.; Zandi, K. Antiviral potential of algae polysaccharides isolated from marine sources: A review. BioMed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr. Polym. 2007, 69, 530–537. [Google Scholar] [CrossRef]
- Cardoso, M.S.; Carvalho, G.L.; Silva, J.P.; Rodrigues, S.M.; Pereira, R.O.; Pereira, L. Bioproducts from seaweeds: A review with special focus on the Iberian Peninsula. Curr. Org. Chem. 2014, 18, 896–917. [Google Scholar] [CrossRef]
- Tutor, M.A.; Meyer, A.S. Fucoidans from brown seaweeds: An update on structures, extraction techniques and use of enzymes as tools for structural elucidation. RSC Adv. 2013, 3, 8131–8141. [Google Scholar]
- Alves, A.; Sousa, R.A.; Reis, R.L. A practical perspective on ulvan extracted from green algae. J. Appl. Phycol. 2013, 25, 407–424. [Google Scholar] [CrossRef] [Green Version]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Kumar, M.; Reddy, C.R.K.; Jha, B. Algal lipids, fatty acids and sterols. In Functional Ingredients from Algae for Foods and Nutraceuticals; Domínguez, H., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 87–134. [Google Scholar]
- Plouguerné, E.; da Gama, B.A.P.; Pereira, R.C.; Barreto-Bergter, E. Glycolipids from seaweeds and their potential biotechnological applications. Front. Cell. Infect. Microbiol. 2014, 4, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.P.; Sidana, J. Phlorotannins. In Functional Ingredients from Algae for Foods and Nutraceuticals; Domínguez, H., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 181–204. [Google Scholar]
- Gupta, S.; Abu-Ghannam, N. Recent developments in the application of seaweeds or seaweed extracts as a means for enhancing the safety and quality attributes of foods. Innov. Food Sci. Emerg. Technol. 2011, 12, 600–609. [Google Scholar] [CrossRef]
- Kraan, S. Pigments and minor compounds in algae. In Functional Ingredients from Algae for Foods and Nutraceuticals; Domínguez, H., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 205–251. [Google Scholar]
- Christaki, E.; Bonos, E.; Giannenas, I.; Florou-Paneri, P. Functional properties of carotenoids originating from algae. J. Sci. Food Agric. 2013, 93, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.S.; Thakur, S.R.; Bansal, P. Algal lectins as promising biomolecules for biomedical research. Crit. Rev. Microbiol. 2015, 41, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Cheung, R.C.F.; Wong, J.H.; Pan, W.; Chan, Y.S.; Yin, C.; Dan, X.; Ng, T.B. Marine lectins and their medicinal applications. Appl. Microbiol. Biotechnol. 2015, 99, 3755–3773. [Google Scholar] [CrossRef] [PubMed]
- Güven, K.S.; Percot, A.; Sezik, E. Alkaloids in Marine Algae. Mar. Drugs 2010, 8, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.; Valentão, P.; Andrade, P.B. Bioactive compounds from macroalgae in the new millennium: Implications for neurodegenerative diseases. Mar. Drugs 2014, 12, 4934–4972. [Google Scholar] [CrossRef] [PubMed]
- Bedoux, G.; Hardouin, K.; Burlot, A.S.; Bourgougnon, N. Bioactive compounds from seaweeds: Cosmetic applications and future development. Adv. Bot. Res. 2014, 71, 345–379. [Google Scholar]
- El Gamal, A.A. Biological importance of marine algae. Saudi Pharm. J. 2010, 18, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Ely, R.; Supriya, T.; Naik, C.G. Antimicrobial activity of marine organisms collected off the coast of South East India. J. Exp. Mar. Biol. Ecol. 2004, 309, 121–127. [Google Scholar] [CrossRef]
- Pinteus, S.; Alves, C.; Monteiro, H.; Araújo, E.; Horta, A.; Pedrosa, R. Asparagopsis armata and Sphaerococcus coronopifolius as a natural source of antimicrobial compounds. World J. Microbiol. Biotechnol. 2015, 31, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Kolsi, R.B.A.; Frikha, D.; Jribi, I.; Hamza, A.; Feki, L.; Belghith, K. Screening of antibacterial and antifongical activity in marine macroalgae and Magnoliophytea from coast of Tunisia. Int. J. Pharm. Pharm. Sci. 2015, 7, 47–51. [Google Scholar]
- Cavallo, R.A.; Acquaviva, M.I.; Stabili, L.; Cecere, E.; Petrocelli, A.; Narracci, M. Antibacterial activity of marine macroalgae against fish pathogenic Vibrio species. Cent. Eur. J. Biol. 2013, 8, 646–653. [Google Scholar] [CrossRef]
- Mhadhebi, L.; Chaiebb, K.; Bouraoui, A. Evaluation of antimicrobial activity of organic fractions of six marine algae from tunisian Mediterranean coasts. Int. J. Pharm. Pharm. Sci. 2012, 4, 534–537. [Google Scholar]
- Bansemir, A.; Blume, M.; Schröder, S.; Lindequist, U. Screening of cultivated seaweeds for antibacterial activity against fish pathogenic bacteria. Aquaculture 2006, 252, 79–84. [Google Scholar] [CrossRef]
- Beaulieu, L.; Thibodeau, J.; Desbiens, M.; Saint-Louis, R.; Zatylny-Gaudin, C.; Thibault, S. Evidence of antibacterial activities in peptide fractions originating from snow crab (Chionoecetes opilio) by-products. Probiotics Antimicrob. Proteins 2010, 2, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Bouhlal, R.; Riadi, H.; Martínez, J.; Bourgougnon, N. The antibacterial potential of the seaweeds (Rhodophyceae) of the Strait of Gibraltar and the Mediterranean coast of Morocco. Afr. J. Biotechnol. 2010, 9, 6365–6372. [Google Scholar]
- Thanigaivel, S.; Vijayakumar, S.; Mukherjee, A.; Chandrasekaran, N.; Thomas, J. Antioxidant and antibacterial activity of Chaetomorpha antennina against shrimp pathogen Vibrio parahaemolyticus. Aquaculture 2014, 433, 467–475. [Google Scholar] [CrossRef]
- Thanigaivel, S.; Vijayakumar, S.; Gopinath, S.; Mukherjee, A.; Chandrasekaran, N.; Thomas, J. In vivo and in vitro antimicrobial activity of Azadirachta indica (Lin) against Citrobacter freundii isolated from naturally infected Tilapia (Oreochromis mossambicus). Aquaculture 2015, 437, 252–255. [Google Scholar] [CrossRef]
- Shanmughapriya, S.; Manilal, A.; Sujith, S.; Selvin, J. Seghal Kiran, G.; Natarajaseenivasan, K. Antimicrobial activity of seaweeds extracts against multiresistant pathogens. Ann. Microbiol. 2008, 58, 535–541. [Google Scholar] [CrossRef]
- García-Bueno, N.; Decottignies, P.; Turpin, V.; Dumay, J.; Paillard, C.; Stiger-Pouvreau, V.; Kervarec, N.; Pouchus, Y.F.; Marín-Atucha, A.A.; Fleurence, J. Seasonal antibacterial activity of two red seaweeds, Palmaria palmata and Grateloupia turuturu, on European abalone pathogen Vibrio harveyi. Aquat. Living Resour. 2014, 27, 83–89. [Google Scholar] [CrossRef]
- Cox, S.; Turley, G.H.; Rajauria, G.; Abu-Ghannam, N.; Jaiswal, A.K. Antioxidant potential and antimicrobial efficacy of seaweed (Himanthalia elongata) extract in model food systems. J. Appl. Phycol. 2014, 26, 1823–1831. [Google Scholar] [CrossRef]
- Dubber, D.; Harder, T. Extracts of Ceramium rubrum, Mastocarpus stellatus and Laminaria digitata inhibit growth of marine and fish pathogenic bacteria at ecologically realistic concentrations. Aquaculture 2008, 274, 196–200. [Google Scholar] [CrossRef]
- Boisvert, C.; Beaulieu, L.; Bonnet, C.; Pelletier, E. Assessment of the antioxidant and antibacterial activities of three species of edible seaweeds. J. Food Biochem. 2015, 39, 377–387. [Google Scholar] [CrossRef]
- Bazes, S.; Silkina, A.; Defer, D.; Bernède-Bauduin, C.; Quéméner, E.; Braud, J.P.; Bourgougnon, N. Active substances from Ceramium botryocarpum used as antifouling products in aquaculture. Aquaculture 2006, 258, 664–674. [Google Scholar] [CrossRef]
- Hellio, C.; De La Broise, D.; Dufossé, L.; Le Gal, Y.; Bourgougnon, N. Inhibition of marine bacteria by extract of macroalgae: Potential use for environmentally friendly paints. Mar. Environ. Res. 2001, 52, 231–247. [Google Scholar] [CrossRef]
- Spavieri, J.; Allmendinger, A.; Kaiser, M.; Casey, R.; Hingley-Wilson, S.; Lalvani, A.; Guiry, M.D.; Blunden, G.; Tasdemir, D. Antimycobacterial, antiprotozoal and cytotoxic potential of twenty-one brown algae (Phaeophyceae) from British and Irish waters. Phytother. Res. 2010, 24, 1724–1729. [Google Scholar] [CrossRef] [PubMed]
- Bouhlal, R.; Riadi, H.; Bourgougnon, N. Antiviral activity of the extracts of Rhodophyceae from Morocco. Afr. J. Biotechnol. 2010, 9, 7968–7975. [Google Scholar]
- Wang, W.; Wang, S.X.; Guan, H.S. The antiviral activities and mechanisms of marine polysaccharides: An overview. Mar. Drugs 2012, 10, 2795–2816. [Google Scholar] [CrossRef] [PubMed]
- Manilal, A.; Selvin, J.; George, S. In vivo therapeutic potentiality of red seaweed, Asparagopsis (Bonnemaisoniales, Rhodophyta) in the treatment of vibriosis in Penaeus monodon, Fabricius. Saudi J. Biol. Sci. 2012, 19, 165–175. [Google Scholar]
- Freile-Pelegrín, Y.; Morales, J.L. Antibacterial activity in marine algae from the coast of Yucatan, Mexico. Bot. Mar. 2004, 47, 140–146. [Google Scholar] [CrossRef]
- Trigui, M.; Gasmi, L.; Zouari, I.; Tounsi, S. Seasonal variation in phenolic composition, antibacterial and antioxidant activities of Ulva rigida (Chlorophyta) and assessment of antiacetylcholinesterase potential. J. Appl. Phycol. 2013, 25, 319–328. [Google Scholar] [CrossRef]
- Stabili, L.; Acquaviva, M.I.; Biandolino, F.; Cavallo, R.A.; De Pascali, S.A.; Fanizzi, F.C.; Narracci, M.; Cecere, E.; Petrocelli, A. Biotechnological potential of the seaweed Cladophora rupestris (Chlorophyta, Cladophorales) lipidic extract. New Biotechnol. 2014, 31, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Stirk, W.A.; Reinecke, D.L.; van Staden, J. Seasonal variation in antifungal, antibacterial and acetylcholinesterase activity in seven South African seaweeds. J. Appl. Phycol. 2007, 19, 271–276. [Google Scholar] [CrossRef]
- Choi, J.S.; Ha, Y.M.; Lee, B.B.; Moon, H.E.; Cho, K.K.; Choi, I.S. Seasonal variation of antibacterial activities in the green alga Ulva pertusa Kjellman. J. Environ. Biol. 2014, 35, 341–344. [Google Scholar] [PubMed]
- Krish, S.; Das, A. In-vitro bioactivity of marine seaweed, Cladophora rupestris. Int. J. Pharm. Biol. Sci. 2014, 5, 898–908. [Google Scholar]
- Padmakumar, K.; Ayyakkannu, K. Seasonal variation of antibacterial and antifungal activities of the extracts of marine algae from southern coasts of India. Bot. Mar. 1997, 40, 507–515. [Google Scholar] [CrossRef]
- Tanniou, A.; Vandanjon, L.; Incera, M.; Serrano Leon, E.; Husa, V.; Le Grand, J.; Nicolas, J.L.; Poupart, N.; Kervarec, N.; Engelen, A.; et al. Assessment of the spatial variability of phenolic contents and associated bioactivities in the invasive alga Sargassum muticum sampled along its European range from Norway to Portugal. J. Appl. Phycol. 2014, 26, 1215–1230. [Google Scholar] [CrossRef]
- El Amraoui, B.; El Amraoui, M.; Cohen, N.; Fassouane, A. Anti-Candida and anti-Cryptococcus antifungal produced by marine microorganisms. J. Mycol. Med. 2014, 24, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Villarreal-Gómez, L.J.; Soria-Mercado, I.E.; Guerra-Rivas, G.; Ayala-Sánchez, N.E. Antibacterial and anticancer activity of seaweeds and bacteria associated with their surface. Rev. Biol. Mar. Oceanogr. 2010, 45, 267–275. [Google Scholar] [CrossRef]
- Horta, A.; Pinteus, S.; Alves, C.; Fino, N.; Silva, J.; Fernandez, S.; Rodrigues, A.; Pedrosa, R. Antioxidant and antimicrobial potential of the Bifurcaria bifurcata epiphytic bacteria. Mar. Drugs 2014, 12, 1676–1689. [Google Scholar] [CrossRef] [PubMed]
- Mendes, M.; Pereira, R.; Sousa Pinto, I.; Carvalho, A.P.; Gomes, A.M. Antimicrobial activity and lipid profile of seaweed extracts from the North Portuguese Coast. Int. Food Res. J. 2013, 20, 3337–3345. [Google Scholar]
- Chambers, L.D.; Hellio, C.; Stokes, K.R.; Dennington, S.P.; Goodes, L.R.; Wood, R.J.K.; Walsh, F.C. Investigation of Chondrus crispus as a potential source of new antifouling agents. Int. Biodeterior. Biodegrad. 2011, 65, 939–946. [Google Scholar] [CrossRef]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. Effect of processing conditions on phytochemical constituents of edible Irish seaweed Himanthalia elongate. J. Food Proc. Preserv. 2012, 36, 348–363. [Google Scholar] [CrossRef]
- Eom, S.H.; Lee, D.S.; Kang, Y.M.; Son, K.T.; Jeon, Y.J.; Kim, Y.M. Application of yeast Candida utilis to ferment Eisenia. bicyclis for enhanced antibacterial effect. Appl. Biochem. Biotechnol. 2013, 171, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Rosaline, S.D.; Sakthivelkumar, S.; Rajendran, K.; Janarthanan, S. Screening of selected marine algae from the coastal Tamil Nadu, South India for antibacterial activity. Asian Pac. J. Trop. Biomed. 2012, 2, S140–S146. [Google Scholar] [CrossRef]
- Saritha, K.; Mani, A.E.; Priyalaxmi, M.; Patterson, J. Antibacterial activity and biochemical constituents of seaweed Ulva lactuca. Glob. J. Pharmacol. 2013, 7, 276–282. [Google Scholar]
- Karthikeyan, K.; Shweta, K.; Jayanthi, G.; Prabhu, K.; Thirumaran, G. Antimicrobial and antioxidant potential of selected seaweeds from Kodinar, Southern Coast of Saurashtra, Gujarat, India. J. Appl. Pharm. Sci. 2015, 5, 35–40. [Google Scholar] [CrossRef]
- Jaswir, I.; Tope, A.H.T.; Raus, R.A.; Monsur, H.A.; Ramli, N. Study on anti-bacterial potentials of some Malaysian brown seaweeds. Food Hydrocoll. 2014, 42, 275–279. [Google Scholar] [CrossRef]
- Osman, M.E.H.; Abushady, A.M.; Elshobary, M.E. In vitro screening of antimicrobial activity of extracts of some macroalgae collected from Abu-Qir bay Alexandria, Egypt. Afr. J. Biotechnol. 2010, 9, 7203–7208. [Google Scholar]
- Kavita, K.; Singh, V.K.; Jha, B. 24-branched delta 5 sterols from Laurencia papillosa red seaweed with antibacterial activity against human pathogenic bacteria. Microbiol. Res. 2014, 169, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Al Hazzani, A.A.; Shehata, A.I.; Moubayed, N.M.S.; Al Houri, H.J. Antimicrobial and biochemical properties of selected edible brown and red marine macroalgae. J. Pure Appl. Microbiol. 2014, 8, 1275–1282. [Google Scholar]
- Horincar, V.B.; Parfene, G.; Tyagi, A.K.; Gottardi, D.; Dinica, R.; Guerzoni, M.E.; Bahrim, G. Extraction and characterization of volatile compounds and fatty acids from red and green macroalgae from the Romanian Black Sea in order to obtain valuable bioadditives and biopreservatives. J. Appl. Phycol. 2014, 26, 551–559. [Google Scholar] [CrossRef]
- Muñoz-Ochoa, M.; Murillo-Álvarez, J.I.; Zermeño-Cervantes, L.A.; Martínez-Díaz, S.; Rodríguez-Riosmena, R. Screening of extracts of algae from Baja California Sur, Mexico as reversers of the antibiotic resistance of some pathogenic bacteria. Eur. Rev. Med. Pharmacol. Sci. 2010, 14, 739–747. [Google Scholar] [PubMed]
- Salta, M.; Wharton, J.A.; Dennington, S.P.; Stoodley, P.; Stokes, K.R. Anti-biofilm performance of three natural products against initial bacterial attachment. Int. J. Mol. Sci. 2013, 14, 21757–21780. [Google Scholar] [CrossRef] [PubMed]
- Manilal, A.; Sujith, S.; Selvin, J.; Kiran, G.S.; Shakir, C.; Lipton, A.P. Antimicrobial potential of marine organisms collected from the southwest coast of India against multiresistant human and shrimp pathogens. Sci. Mar. 2010, 74, 287–296. [Google Scholar]
- Jiang, Z.B.; Chen, Y.C.; Yao, F.; Chen, W.Z.; Zhong, S.P.; Zheng, F.C.; Shi, G.G. Antioxidant, antibacterial and antischistosomal activities of extracts from Grateloupia livida (Harv). Yamada. PLoS. ONE 2013, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Amorim, R.D.D.; Rodrigues, J.A.G.; Holanda, M.L.; Quindere, A.L.G.; de Paula, R.C.M.; Melo, V.M.M.; Benevides, N.M.B. Antimicrobial effect of a crude sulfated polysaccharide from the red seaweed Gracilaria ornata. Braz. Arch. Biol. Technol. 2012, 55, 171–181. [Google Scholar] [CrossRef]
- Stabili, L.; Acquaviva, M.I.; Biandolino, F.; Cavallo, R.A.; De Pascali, S.A.; Fanizzi, F.P.; Narracini, M.; Petrocelli, A.; Cecere, E. The lipidic extract of the seaweed Gracilariopsis longissima (Rhodophyta, Gracilariales): A potential resource for biotechnological purposes? New Biotechnol. 2012, 29, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Taskin, E.; Caki, Z.; Ozturk, M.; Taskin, E. Assessment of in vitro antitumoral and antimicrobial activities of marine algae harvested from the eastern Mediterranean sea. Afr. J. Biotechnol. 2010, 9, 4272–4277. [Google Scholar]
- Kandhasamy, M.; Arunachalam, K.D. Evaluation of in vitro antibacterial property of seaweeds of southeast coast of India. Afr. J. Biotechnol. 2008, 7, 1958–1961. [Google Scholar]
- Bianco, E.M.; de Oliveira, S.Q.; Rigotto, C.; Tonini, M.L.; Guimaraes, T.D.; Bittencourt, F.; Gouvea, L.P.; Aresi, C.; de Almeida, M.T.R.; Moritz, M.I.G.; et al. Anti-Infective potential of marine invertebrates and seaweeds from the Brazilian coast. Molecules 2013, 18, 5761–5778. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.P.; McLoughlin, P.; O’Sullivan, L.; Prieto, M.L.; Gardiner, G.E.; Lawlor, P.G.; Hughes, H. Development of a novel antimicrobial seaweed extract-based hydrogel wound dressing. Int. J. Pharm. 2013, 456, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.; Alves, C.; Horta, A.; Pinteus, S.; Silva, J.; Culioli, G.; Thomas, O.P.; Pedrosa, R. Antitumor and antimicrobial potential of bromoditerpenes Isolated from the red alga, Sphaerococcus coronopifolius. Mar. Drugs 2015, 13, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Nagappan, T.; Vairappan, C.S. Nutritional and bioactive properties of three edible species of green algae, genus Caulerpa (Caulerpaceae). J. Appl. Phycol. 2014, 26, 1019–1027. [Google Scholar] [CrossRef]
- Pierre, G.; Sopena, V.; Juin, C.; Mastouri, A.; Graber, M.; Maugard, T. Antibacterial activity of a sulfated galactan extracted from the marine alga Chaetomorpha aerea against Staphylococcus aureus. Biotechnol. Bioprocess Eng. 2011, 16, 937–945. [Google Scholar] [CrossRef]
- Yuvaraj, N.; Kanmani, P.; Satishkumar, R.; Paari, K.A.; Pattukumar, V.; Arul, V. Extraction, purification and partial characterization of Cladophora glomerata against multidrug resistant human pathogen Acinetobacter baumannii and fish pathogens. World J. Fish Mar. Sci. 2011, 3, 51–57. [Google Scholar]
- Ha, Y.M.; Choi, J.S.; Lee, B.B.; Moon, H.E.; Cho, K.K.; Choi, I.S. Inhibitory effects of seaweed extracts on the growth of the vaginal bacterium Gardnerella vaginalis. J. Environ. Biol. 2014, 35, 537–542. [Google Scholar] [PubMed]
- Kosanic, M.; Rankovic, B.; Stanojkovic, T. Biological activities of two macroalgae from Adriatic coast of Montenegro. Saudi J. Biol. Sci. 2015, 22, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Park, N.H.; Choi, J.S.; Hwang, S.Y.; Kim, Y.C.; Hong, Y.K.; Cho, K.K.; Choi, I.S. Antimicrobial activities of stearidonic and gamma-linolenic acids from the green seaweed Enteromorpha linza against several oral pathogenic bacteria. Bot. Stud. 2013, 54, 39–47. [Google Scholar] [CrossRef]
- Patra, J.K.; Das, G.; Baek, K.H. Antibacterial mechanism of the action of Enteromorpha linza L. essential oil against Escherichia coli and Salmonella typhimurium. Bot. Stud. 2015, 56, 13–21. [Google Scholar]
- Lee, J.H.; Eom, S.H.; Lee, E.H.; Jung, Y.J.; Kim, H.J.; Jo, M.R.; Son, K.T.; Lee, H.J.; Kim, J.H.; Lee, M.S.; et al. In vitro antibacterial and synergistic effect of phlorotannins isolated from edible Brown seaweed Eisenia bicyclis against acne-related bacteria. Algae 2014, 29, 47–55. [Google Scholar] [CrossRef]
- Amiguet, V.T.; Jewell, L.E.; Mao, H.; Sharma, M.; Hudson, J.B.; Durst, T.; Allard, M.; Rochefort, G.; Arnason, J.T. Antibacterial properties of a glycolipid-rich extract and active principle from Nunavik collections of the macroalgae Fucus evanescens C. Agardh (Fucaceae). Can. J. Microbiol. 2011, 57, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Plaza, M.; Santoyo, S.; Jaime, L.; Reina, G.G.B.; Herrero, M.; Señoráns, F.J.; Ibáñez, E. Screening for bioactive compounds from algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Rajauria, G.; Jaiswal, A.K.; Abu-Gannam, N.; Gupta, S. Antimicrobial, antioxidant and free radical-scavenging capacity of brown seaweed Himanthalia. elongata from western coast of Ireland. J. Food Biochem. 2013, 37, 322–335. [Google Scholar] [CrossRef]
- Patra, J.K.; Das, G.; Baek, K.H. Chemical composition and antioxidant and antibacterial activities of an essential oil extracted from an edible seaweed, Laminaria japonica L. Molecules 2015, 20, 12093–12113. [Google Scholar] [CrossRef] [PubMed]
- Manilal, J.S.; Thajuddin, N.; Sujith, S.; Panikkar, M.V.N.; Idhayadhulla, A.; Kumar, R.S. Biopotentials of marine alga, Lobophora variegata collected from the shouth indian littoral. Thalassas 2012, 28, 47–54. [Google Scholar]
- Nogueira, L.F.; Morais, E.C.; Brito, M.A.; Santos, B.S.; Vale, D.L.; Lucena, B.F.; Figueredo, F.G.; Guedes, G.M.; Tintino, S.R.; Souza, C.E.; et al. Evaluation of antibacterial, antifungal and modulatory activity of methanol and ethanol extracts of Padina sanctae-crucis. Afr. Health Sci. 2014, 14, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Meillisa, A.; Siahaan, E.A.; Park, J.N.; Woo, H.C.; Chun, B.S. Effect of subcritical water hydrolysate in the brown seaweed Saccharina japonica as a potential antibacterial agent on food-borne pathogens. J. Appl. Phycol. 2013, 25, 763–769. [Google Scholar] [CrossRef]
- Sivagnanam, S.P.; Yin, S.; Choi, J.H.; Park, Y.B.; Woo, H.C.; Chun, B.S. Biological properties of fucoxanthin in oil recovered from two brown seaweeds using supercritical CO2 extraction. Mar. Drugs 2015, 13, 3422–3442. [Google Scholar] [CrossRef] [PubMed]
- Adaikalaraj, G.; Patric, R.D.; Johnson, M.; Janakiraman, N.; Babu, A. Antibacterial potential of selected red seaweeds from Manapad coastal areas, Thoothukudi, Tamil Nadu, India. Asian Pac. J. Trop. Biomed. 2012, 2, 1077–1080. [Google Scholar] [CrossRef]
- Jassbi, A.R.; Mohabati, M.; Eslami, S.; Sohrabipour, J.; Miri, R. Biological activity and chemical constituents of red and brown algae from the Persian Gulf. Iran. J. Pharm. Res. 2013, 12, 339–348. [Google Scholar] [PubMed]
- De Jesus Raposo, M.F.; de Morais, A.M.B.; de Morais, R.M.S.C. Marine polysaccharides from algae with potential biomedical applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef] [PubMed]
- Varahalarao, V.; Chandrasekhar, N.K. In vitro antimicrobial potentiality of some marine algae against selected phytopathogens. Biomed. Pharmacol. J. 2009, 2, 277–280. [Google Scholar]
- Gupta, S.; Cox, S.; Rajauria, G.; Jaiswal, A.K.; Abu-Ghannam, N. Growth inhibition of common food spoilage and pathogenic microorganisms in the presence of brown seaweed extracts. Food Bioprocess Technol. 2012, 5, 1907–1916. [Google Scholar] [CrossRef]
- Salem, W.M.; Galal, H.; Nasr El-deen, F. Screening for antibacterial activities in some marine algae from the red sea (Hurghada, Egypt). Afr. J. Microbiol. Res. 2011, 5, 2160–2167. [Google Scholar] [CrossRef]
- Shanmugam, J.; Devi Raman, K.; Viswanathan, S.; Nallamuthu, T. Antibacterial and antioxidant activity of red seaweeds from Kilakarai, Rameswaram, Tamilnadu, India. J. Pharm. Biomed. Sci. 2013, 32, 1386–1395. [Google Scholar]
- Patra, J.K.; Kim, S.H.; Baek, K.H. Antioxidant and free radical-scavenging potential of essential oil from Enteromorpha. linza L. prepared by microwave-assisted hydrodistillation. J. Food Biochem. 2015, 39, 80–90. [Google Scholar] [CrossRef]
- Kadam, S.U.; O’Donnell, C.P.; Rai, D.K.; Hossain, M.B.; Burgess, C.M.; Walsh, D.; Tiwari, B.K. Laminarin from Irish brown seaweeds Ascophyllum nodosum and Laminaria hyperborea: Ultrasound Assisted Extraction, characterization and bioactivity. Mar. Drugs 2015, 13, 4270–4280. [Google Scholar] [CrossRef] [PubMed]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Lee, J.-A.; Kim, K.-N.; Yoon, W.-J.; Lee, W.J.; Park, S.-Y. Antioxidative and Antimicrobial activities of Sargassum muticum extracts. J. Korean Soc. Food Sci. Nutr. 2007, 36, 663–669. [Google Scholar] [CrossRef]
- De Felício, R.; de Albuquerque, S.; Young, M.C.M.; Yokoya, N.S.; Debonsi, H.M. Trypanocidal, leishmanicidal and antifungal potential from marine red alga Bostrychia tenella J. Agardh (Rhodomelaceae, Ceramiales). J. Pharm. Biomed. Anal. 2010, 52, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Salah, K.B.H.; Ahmed, M.; Mastouri, M.; Bouraoui, A.; Aouni, M. Antibacterial and antifungal activities of brown alga Zonaria tournefortii (JV Lamouroux). Allelopathy J. 2014, 34, 143–153. [Google Scholar]
- Barreto, M.; Meyer, J.J.M. Isolation and antimicrobial activity of a lanosol derivative from Osmundaria serrata (Rhodophyta) and a visual exploration of its biofilm covering. S. Afr. J. Bot. 2006, 72, 521–528. [Google Scholar] [CrossRef]
- Vairappan, C.S.; Suzuki, M.; Ishii, T.; Okino, T.; Abe, T.; Masuda, M. Antibacterial activity of halogenated sesquiterpenes from Malaysian Laurencia spp. Phytochemistry 2008, 69, 2490–2494. [Google Scholar] [CrossRef] [PubMed]
- Ben Redjem, Y.; Ktari, L.; Medhioub, A.; Romdhane, M.S.; Langar, H.; El Bour, M. Antibacterial and algicidal properties of some brown seaweeds from Northern coasts of Tunisia. Life Environ. 2013, 63, 127–133. [Google Scholar]
- Mathan, S.; Subramanian, V.; Nagamony, S.; Ganapathy, K. Isolation of endophytic fungiform marine algae and its bioactivity. Int. J. Res. Pharm. Sci. 2013, 4, 45–49. [Google Scholar]
- Cortés, Y.; Hormazábal, E.; Leal, H.; Urzúa, A.; Mutis, A.; Parra, L.; Quiroz, A. Novel antimicrobial activity of a dichloromethane extract obtained from red seaweed Ceramium rubrum (Hudson) (Rhodophyta: Florideophyceae) against Yersinia ruckeri and Saprolegnia. parasitica, agents that cause diseases in salmonids. Electron. J. Biotechnol. 2014, 17, 126–131. [Google Scholar] [CrossRef]
- Thanigaivel, S.; Chandrasekaran, N.; Mukherjee, A.; Thomas, J. Investigation of seaweed extracts as a source of treatment against bacterial fish pathogen. Aquaculture 2015, 448, 82–86. [Google Scholar] [CrossRef]
- Oliveira, S.T.L.; Veneroni-Gouveia, G.; Santos, A.C.; Sousa, S.M.N.; Veiga, M.L.; Krewer, C.C.; Costa, M.M. Ascophyllum nodosum in the diet of tilapia (Oreochromis niloticus) and its effect after inoculation of Aeromonas hydrophila. Pesqui. Vet. Bras. 2014, 34, 403–408. [Google Scholar] [CrossRef]
- Dhanya, K.I.; Swati, V.I.; Vanka, K.S.; Osborne, W.J. Antimicrobial activity of Ulva reticulata and its endophytes. J. Ocean Univ. China, 2016, 15, 363–369. [Google Scholar] [CrossRef]
- Kim, I.H.; Lee, D.G.; Lee, S.H.; Ha, J.M.; Ha, B.J.; Kim, S.K.; Lee, J.H. Antibacterial activity of Ulva lactuca against methicillin-resistant Staphylococcus aureus (MRSA). Biotechnol. Bioprocess Eng. 2007, 12, 579–582. [Google Scholar] [CrossRef]
- Oh, K.; Lee, J.H.; Chung, S.C.; Shin, J.; Shin, H.J.; Kimd, H.K.; Lee, H.S. Antimicrobial activities of the bromophenols from the red alga Odonthalia corymbifera and some synthetic derivatives. Bioorg. Med. Chem. Lett. 2008, 18, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Genovese, G.; Leitner, S.; Minicante, S.A.; Lass-Florl, C. The Mediterranean red alga Asparagopsis taxiformis has antifungal activity against Aspergillus species. Mycoses 2013, 56, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Mohandass, C.; Vijayaraj, A.S.; Rajasabapathy, R.; Satheeshbabu, S.; Rao, S.V.; Shiva, C.; De-Mello, L. Biosynthesis of silver nanoparticles from marine seaweed Sargassum cinereum and their antibacterial activity. Indian J. Pharm. Sci. 2013, 75, 606–610. [Google Scholar] [PubMed]
- Osman, M.E.H.; Aboshady, A.M.; Elshobary, M.E. Production and characterization of antimicrobial active substance from some macroalgae collected from AbuQir bay (Alexandria) Egypt. Afr. J. Biotechnol. 2013, 12, 6847–6858. [Google Scholar]
- Gerasimenko, N.I.; Martyyas, E.A.; Busarova, N.G. Composition of lipids and biological activity of lipids and photosynthetic pigments from algae of the families Laminariaceae and Alariaceae. Chem. Nat. Compd. 2012, 48, 737–741. [Google Scholar] [CrossRef]
- El Wahidi, M.; El Amraoui, B.; El Amraoui, M.; Bamhaoud, T. Screening of antimicrobial activity of macroalgae extracts from the Moroccan Atlantic coast. Ann. Pharm. Fr. 2015, 73, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Andrade, P.B.; Valentão, P. Screening of a marine algal extract for antifungal activities. Methods Mol. Biol. 2015, 1308, 411–420. [Google Scholar] [PubMed]
- Wei, Y.X.; Liu, Q.; Yu, J.; Feng, Q.; Zhao, L.; Song, H.P.; Wang, W.X. Antibacterial mode of action of 1,8-dihydroxy-anthraquinone from Porphyra haitanensis against Staphylococcus aureus. Nat. Prod. Res. 2015, 29, 976–979. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.B.; Choi, J.S.; Moon, H.E.; Ha, Y.M.; Kim, M.S.; Cho, K.K.; Choi, I.S. Inhibition of growth and urease of Helicobacter pylori by Korean edible seaweed extracts. Bot. Sci. 2013, 91, 515–522. [Google Scholar] [CrossRef]
- Vedhagiri, K.; Manilal, A.; Valliyammai, T.; Shanmughapriya, S.; Sujith, S.; Selvin, J.; Natarajaseeniva, K. Antimicrobial potential of a marine seaweed Asparagopsis taxiformis against Leptospira javanica isolates of rodent reservoirs. Ann. Microbiol. 2009, 59, 431–437. [Google Scholar] [CrossRef]
- Choi, J.S.; Ha, Y.M.; Joo, C.U.; Cho, K.K.; Kim, S.J.; Choi, I.S. Inhibition of oral pathogens and collagenase activity by seaweed extracts. J. Environ. Biol. 2012, 33, 115–121. [Google Scholar] [PubMed]
- Kim, Y.H.; Kim, J.H.; Jin, H.J.; Lee, S.Y. Antimicrobial activity of ethanol extracts of Laminaria japonica against oral microorganisms. Anaerobe 2013, 21, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, A.Z.; Buettner, C.; Phillips, R.S.; Davis, R.B.; Mukamal, K.J. n-3 fatty acids and periodontitis in US adults. J. Am. Diet. Assoc. 2010, 110, 1669–1675. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Jeong, M.R.; Choi, S.M.; Na, S.S.; Cha, J.D. Synergistic effect of fucoidan with antibiotics against oral pathogenic bacteria. Arch. Oral Biol. 2013, 58, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Harden, E.A.; Falshaw, R.; Carnachan, S.M.; Kern, E.R.; Prichard, M.N. Virucidal activity of polysaccharide extracts from four algal species against herpes simplex virus. Antivir. Res. 2009, 83, 282–289. [Google Scholar] [CrossRef] [PubMed]
- De Souza, L.M.; Sassaki, G.L.; Romanos, M.T.V.; Barreto-Bergter, E. Structural characterization and anti-HSV-1 and HSV-2 activity of glycolipids from the marine algae Osmundaria obtusiloba isolated from Southeastern Brazilian coast. Mar. Drugs 2012, 10, 918–931. [Google Scholar] [CrossRef] [PubMed]
- Koishi, A.C.; Zanello, P.R.; Bianco, E.M.; Bordignon, J.; dos Santos, C.N.D. Screening of dengue virus antiviral activity of marine seaweeds by an in situ enzyme-linked immunosorbent assay. PLoS ONE 2012, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Abrantes, J.L.; Barbosa, J.; Cavalcanti, D.; Pereira, R.C.; Fontes, C.F.L.; Teixeira, V.L.; Souza, T.M.L.; Paixão, I.C.P. The effects of the diterpenes Isolated from the Brazilian brown algae Dictyota pfaffii and Dictyota menstrualis against the Herpes Simplex Type-1 Replicative Cycle. Planta Med. 2010, 76, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Mendes, G.; Soares, A.R.; Sigiliano, L.; Machado, F.; Kaiser, C.; Romeiro, N.; Gestinari, L.; Santos, N.; Romanos, M.T.V. In vitro anti-HMPV activity of meroditerpenoids from marine alga Stypopodium zonale (Dictyotales). Molecules 2011, 16, 8437–8450. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Karadeniz, F. Anti-HIV activity of extracts and compounds from marine algae. Adv. Food Nutr. Res. 2011, 64, 255–265. [Google Scholar] [PubMed]
- Morya, V.K.; Kim, J.; Kim, E.-K. Algal fucoidan: Structural and size-dependent bioactivities and their perspectives. Appl. Microb. Biotechnol. 2012, 93, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Naesens, L.; Stephens, C.E.; Andrei, G.; Loregian, A.; De Bolle, L.; Snoeck, R.; Sowell, J.W.; De Clercq, E. Antiviral properties of new arylsulfone derivatives with activity against human betaherpesviruses. Antivir. Res. 2006, 72, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.S.; Navid, M.H.; Ghosh, T.; Schnitzler, P.; Ray, B. Structural features and in vitro antiviral activities of sulfated polysaccharides from Sphacelaria indica. Phytochemistry 2011, 72, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Elizondo-González, R.; Cruz-Suárez, L.E.; Ricque-Marie, D.; Mendoza-Gamboa, E.; Rodríguez-Padilla, C.; Trejo-Ávila, L.M. In vitro characterization of the antiviral activity of fucoidan from Cladosiphon okamuranus against Newcastle Disease Virus. Virol. J. 2012, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nagorskaia, V.P.; Reunov, A.V.; Lapshina, L.A.; Ermak, I.M.; Barabanova, A.O. Influence of kappa/beta-carrageenan from red alga Tichocarpus crinitus on development of local infection induced by tobacco mosaic virus in Xanthinc tobacco leaves. Izv. Akad. Nauk. Ser. Biol. 2008, 3, 360–364. [Google Scholar] [PubMed]
- De S.f-Tischer, P.C.; Talarico, L.B.; Noseda, M.D.; Guimarães, S.M.; Damonte, E.B.; Duarte, M.E.R. Chemical structure and antiviral activity of carrageenans from Meristiella gelidium against herpes simplex and dengue virus. Carbohydr. Polym. 2006, 63, 459–465. [Google Scholar] [CrossRef]
- Vijayavel, K.; Martínez, J.A. In vitro antioxidant and antimicrobial activities of two Hawaiian marine Limu: Ulva fasciata (Chlorophyta) and Gracilaria salicornia (Rhodophyta). J. Med. Food. 2010, 13, 1494–1499. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.P.; Suganthy, N.; Kesika, P.; Karutha Pandian, S. Bioprotective properties of seaweeds: In vitro evaluation of antioxidant activity and antimicrobial activity against food-borne bacteria in relation to polyphenolic content. BMC Complement. Altern. Med. 2008, 8, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, L.; Bondu, S.; Doiron, K.; Rioux, L.E.; Turgeon, S.L. Characterization of antibacterial activity from protein hydrolysates of the macroalga Saccharina longicruris and identification of peptides implied in bioactivity. J. Funct. Foods 2015, 17, 685–697. [Google Scholar] [CrossRef]
- Moroney, N.C.; O’Grady, M.N.; O’Doherty, J.V.; Kerry, J.P. Effect of a brown seaweed (Laminaria digitata) extract containing laminarin and fucoidan on the quality and shelf-life of fresh and cooked minced pork patties. Meat Sci. 2013, 94, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Siahaan, E.A.; Pendleton, P.; Woo, H.C.; Chun, B.S. Brown seaweed (Saccharina japonica) as an edible natural delivery matrix for allyl isothiocyanate inhibiting food-borne bacteria. Food Chem. 2014, 152, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.; Dorta, F.; Medina, C.; Ramírez, A.; Ramírez, I.; Peña-Cortés, H. Anti-phytopathogenic activities of macro-algae extracts. Mar. Drugs 2011, 9, 739–756. [Google Scholar] [CrossRef] [PubMed]
- Hierholtzer, A.; Chatellard, L.; Kierans, M.; Akunna, J.C.; Collier, P.J. The impact and mode of action of phenolic compounds extracted from brown seaweed on mixed anaerobic microbial cultures. J. Appl. Microbiol. 2013, 114, 964–973. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, L.; Murphy, B.; McLoughlin, P.; Duggan, P.; Lawlor, P.G.; Hughes, H.; Gardiner, G.E. Review prebiotics from marine macroalgae for human and animal health applications. Mar. Drugs 2010, 8, 2038–2064. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Basic chemical structures of mainly polysaccharides in macroalgae: alginates, carrageenans, agar, galactans, laminarans, fucoidans and ulvans.
Figure 1.
Basic chemical structures of mainly polysaccharides in macroalgae: alginates, carrageenans, agar, galactans, laminarans, fucoidans and ulvans.

Figure 2.
Chemical structures of fatty acids, sterol, phloroglucinol, carotenoids (β-carotene and fucoxanthin), terpenes (neophytadiene and cycloeudesmol) and a brominated compound.
Figure 2.
Chemical structures of fatty acids, sterol, phloroglucinol, carotenoids (β-carotene and fucoxanthin), terpenes (neophytadiene and cycloeudesmol) and a brominated compound.

Figure 3.
Stages proposed for the study and application of antimicrobials from seaweed, crossed out are the problems minimized or avoided.
Figure 3.
Stages proposed for the study and application of antimicrobials from seaweed, crossed out are the problems minimized or avoided.


{kind=link}
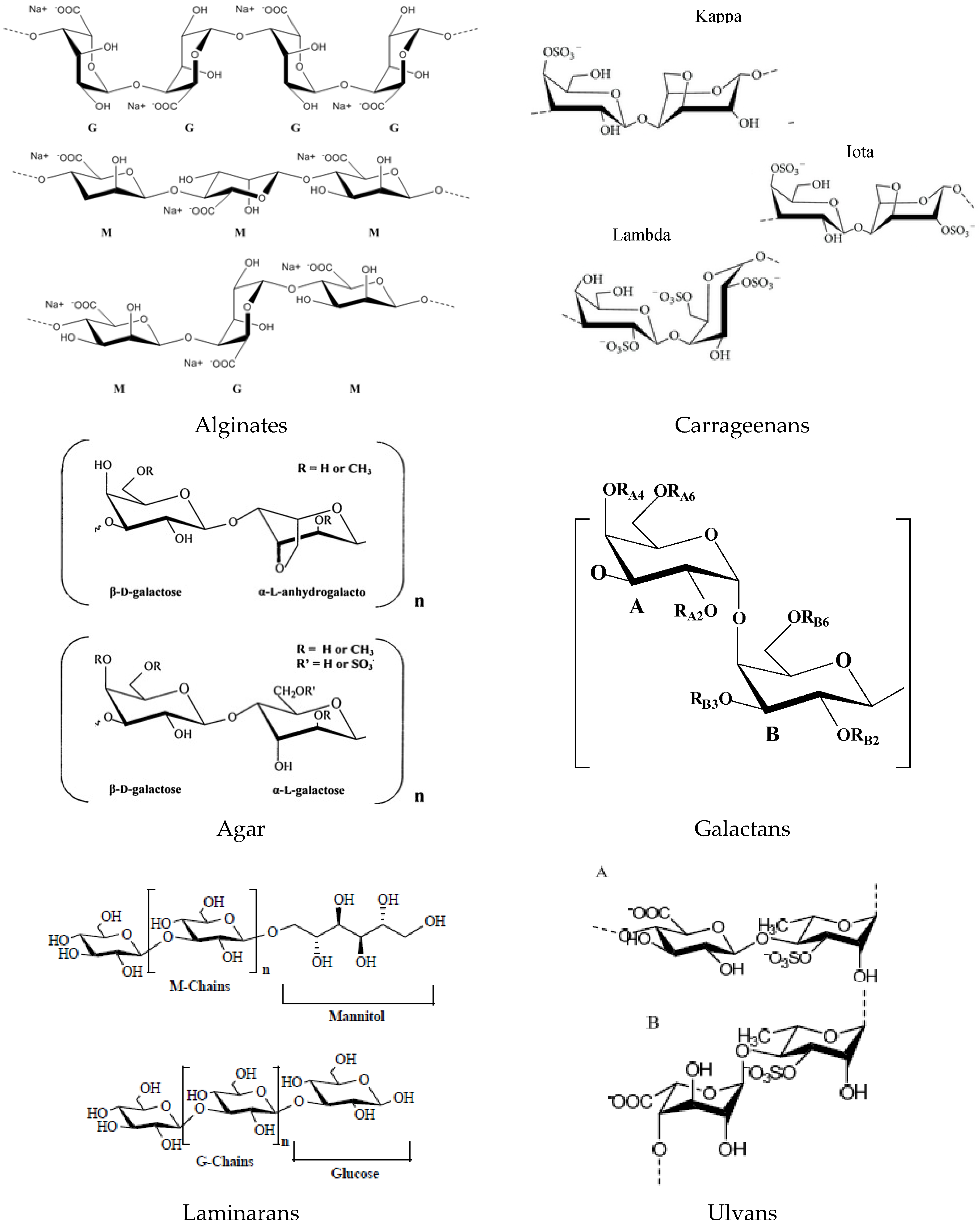{kind=link}
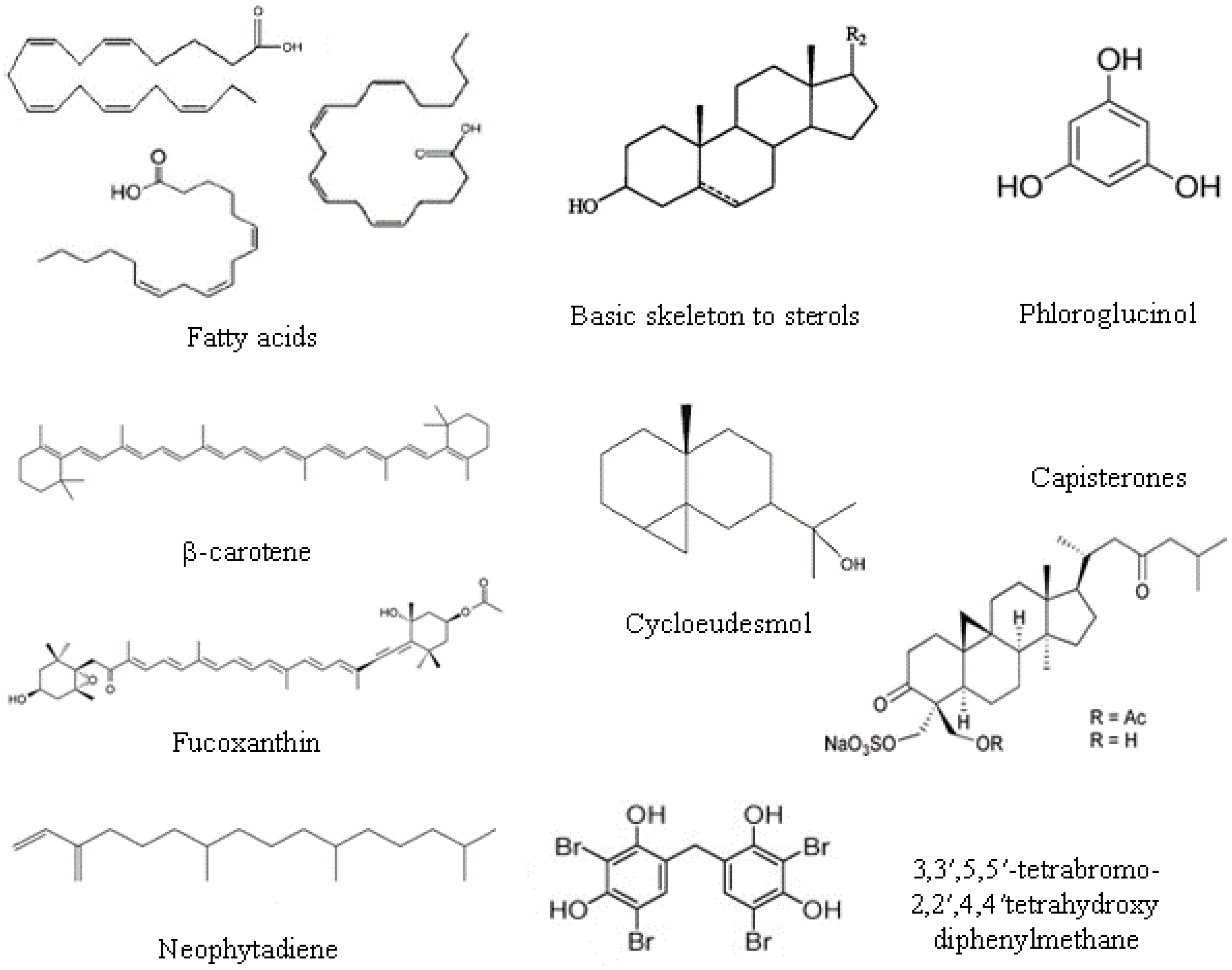{kind=link}
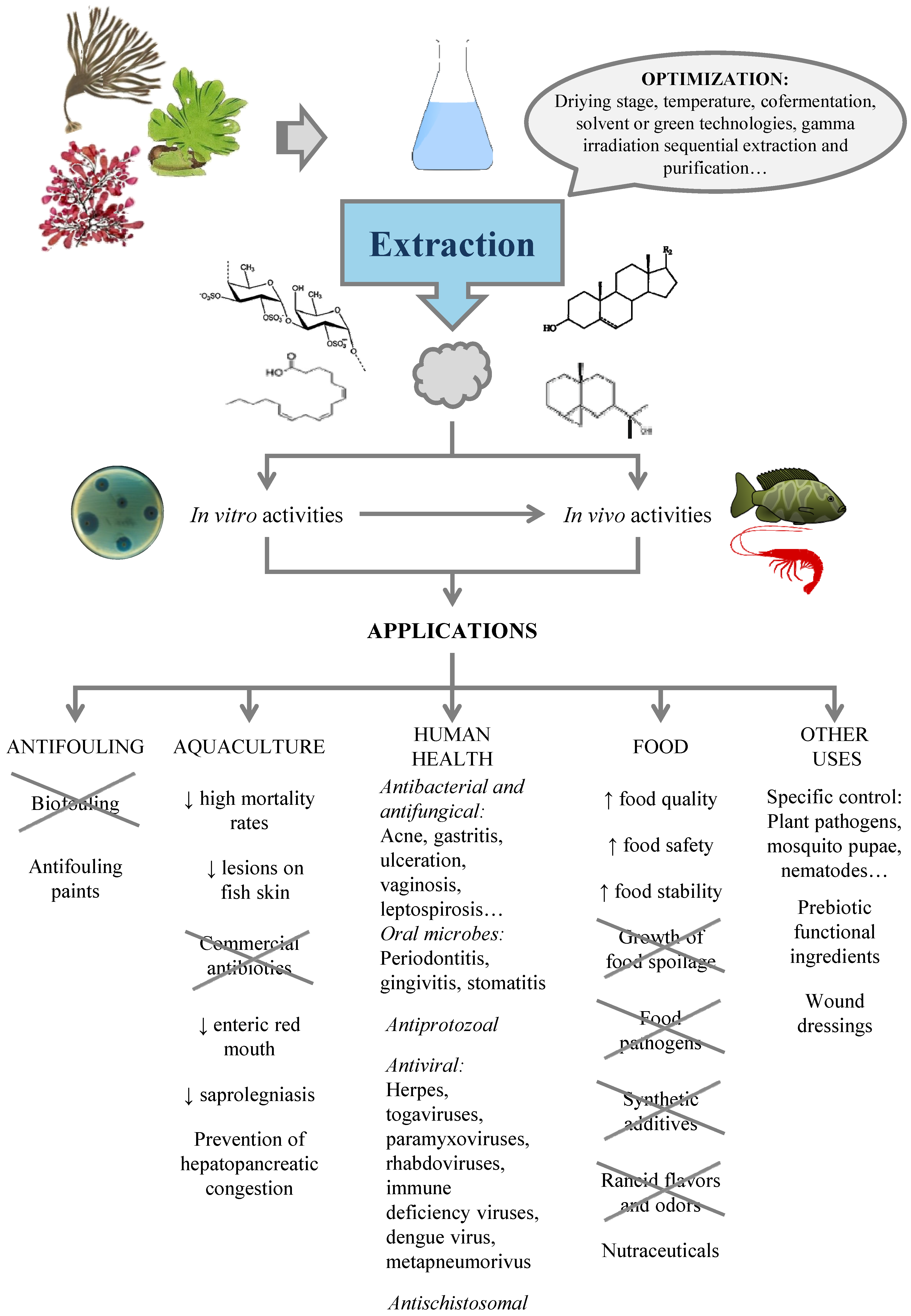{kind=link}
| Red Seaweed | Solvent | Organisms | Ref. |
|---|---|---|---|
| Alsidium corallinum | M | Escherichia coli, Klebsiella pneumoniae, Staphylococcus aureus | [49] |
| Ceramium rubrum | M | E. coli, Enterococcus faecalis, S. aureus | [49] |
| Ceramium virgatum | H/1:1 EE:Hp | Salmonella enteritidis, E.coli, Listeria monocytogenes, Bacillus cereus | [85] |
| Chondrocanthus acicularis | M | E. coli, K. pneumoniae, E. faecalis, S. aureus | [49] |
| Chondracanthus canaliculatus | E | S. aureus, Streptococcus pyogenes | [86] |
| Chondrus crispus | M | L. monocytogenes, Salmonella abony, E. faecalis, P. aeruginosa | [6] |
| C. crispus | 95% E | Pseudoalteromonas elyakovii, Vibrio aestuarianus, Polaribacter irgensii, Halomonas marina, Shewanella putrefaciens | [75] |
| C. crispus | EA | S. enteridis, E. coli, P. aeruginosa, L. innocua, B. cereus, S. aureus, L. brevis. E. faecalis, Candida sp. | [74] |
| DE | E. coli, P. aeruginosa, L. innocua, B. cereus, S. aureus, L. brevis, E. faecalis, Candida sp. | ||
| 1:1 M:W | E. coli, P. aeruginosa, L. innocua, B. cereus, S. aureus, L. brevis. E. faecalis, Candida sp. | ||
| C. crispus | 95% E/T | Cobetia marina, Marinobacter hydrocarbonoclasticus | [87] |
| Corallina elongata | E, A, M | B. subtilis, S. aureus, E. coli, Salmonella typhi, K. pneumoniae, Candida albicans | [82] |
| Corallina vancouverensis | E | S. aureus, S. pyogenes | [86] |
| Falkenbergia-phase of A. taxiformis | M | B. subtilis, K. pneumonia, P. aeruginosa, S aureus, S. epidermidis, Vibrio harveyi, V. alginolyticus, V. vulnificus, V. parahaemolyticus, V. alcaligenes | [88] |
| Ganonema farinosum | E | S. aureus, S. pyogenes | [86] |
| Gelidium attenatum | M | E. coli, K. pneumoniae, E. faecalis, S. aureus | [49] |
| Gelidium micropterum | M | V. parahaemolyticus, V. alcaligenes | [88] |
| Gelidium pulchellum | M | E. coli, E. faecalis, S. aureus | [49] |
| Gelidium pusillum | M | E. coli, K. pneumonia, E. faecalis, S. aureus | [49] |
| M | V. harveyi, V. alginolyticus, V. vulnificus, V. parahaemolyticus, V. alcaligenes | [88] | |
| Gelidium robustum | E | S. aureus, S. pyogenes | [86] |
| Gelidium spinulosum | M | E. coli, E. faecalis, S. aureus | [49] |
| Gracilaria dura | 2:1 C:M | V. ordalii, V. alginolyticus | [45] |
| Gracilaria gracilis | 2:1 C:M | V. salmonicida | [45] |
| Grateloupia livida | E/PE | S. aureus, E. coli, P. aeruginosa | [89] |
| Gracilaria ornata | W | E. coli | [90] |
| Gracilaria subsecundata | E | S. aureus, S. pyogenes | [86] |
| Gracilaria vermiculophylla | EA | S. enteridis, E. coli, P. aeruginosa, L. innocua, B. cereus, S. aureus, L. brevis. E. faecalis | [74] |
| DE | S. enteridis, E. coli, P. aeruginosa, L. innocua, B. cereus, S. aureus, L. brevis. E. faecalis, Candida sp. | ||
| 1:1 M:W | B. cereus, S. aureus | ||
| Gracilariopsis longissima | 2:1 C:M | V. alginolyticus, V. vulnificus, V. ordalii, V. salmonicida | [45] |
| M/2:1 C:M | V. alginolyticus, V. fluvialis, V. vulnificus, V. cholerae non O-1 | [91] | |
| Grateloupia filicina | M | V. harveyi, V. alginolyticus, V. vulnificus, V. parahaemolyticus, V. alcaligenes | [88] |
| G. filicina | M | S.aureus, B. subtilis, E. coli, P. aeruginosa | [83] |
| Halopitys incurvus | M | E. coli, K. pneumoniae, E. faecalis, S. aureus | [49] |
| Hypnea musciformis | M | C. albicans | [92] |
| H. musciformis | M | K. pneumonia, E. faecalis, S. aureus | [93] |
| M | E. coli, K. pneumoniae, E. faecalis, S. aureus | [49] | |
| Hypnea pannosa | M | S. aureus, B. subtilis, E. coli, P. aeruginosa | [83] |
| Hypnea valentiae | E | S. aureus, S. pyogenes | [86] |
| M | S. aureus, B. subtilis, E. coli, P. aeruginosa | [83] | |
| Janiarubens | E, A, M | B. subtilis, S. aureus, E. coli, S. typhi, K. pneumoniae, C. albicans | [82] |
| Laurencia dendroidea | A | S. aureus, E. faecalis | [94] |
| Laurencia jonhstonii | E | S. aureus, S. pyogenes | [86] |
| Laurencia pacifica | E | S. aureus, S. pyogenes | [86] |
| Laurencia papillosa | M | S. aureus, B. subtilis, E. coli, P. aeruginosa | [83] |
| M/SC | E. coli, P. aeruginosa, K. pneumoniae, Shigella flexineri | ||
| Laurencia sp. | M | P. aeruginosa, S.epidermidis, V. harveyi, V. alginolyticus, V. vulnificus, V. parahaemolyticus | [88] |
| Neorhodomela larix | E | S. aureus, S. pyogenes | [86] |
| Ochtodes secundiramea | A | S. aureus | [94] |
| Osmundaria obtusiloba | 2:1 DCM:M | P. aeruginosa | [94] |
| Plocamium cartilagineum | M | E. coli, E. faecalis, S. aureus | [49] |
| Polysiphonia lanosa | W | MRSA,S. aureus, E. cloacae, Clostridium perfringens | [95] |
| Polysphonia tuticorinensis | M | S. aureus, B. subtilis, E. coli, P. aeruginosa | [83] |
| Porphyra dioica | EA | E. coli, Bacillus cereus, Lactobacillus brevis. E. faecalis, Candida sp. | [74] |
| DE | E. coli, B. cereus, L. brevis. E. faecalis, Candida sp. | ||
| 1:1 M:W | S. aureus, E. faecalis | ||
| Portieria horemanii | M | V. harveyi, V. alginolyticus, V. vulnificus, | [88] |
| Pterocladia capillacea | E, A, M | B. subtilis, S. aureus, E. coli, S. typhi, K. pneumoniae, C. albicans | [82] |
| Pterosiphonia complanata | M | E. coli, E. faecalis, S. aureus | [49] |
| Rhodymenia californica | E | S. aureus, S. pyogenes | [86] |
| Sphaerococcus coronopifolius | M | S. aureus | [96] |
| DCM/SC | S. aureus, E. coli, P. aeruginosa, C. albicans | ||
| Spyridia filamentosa | M | S. aureus, E. coli, E. faecalis | [92] |
A: acetone; C: chloroform; DCM: dichloromethane; DE: diethylether; E: ethanol; EA: ethyl acetate; EE: ethyl ether; H: hexane; Hp: heptane; M: methanol; T: toluene; W: water; SC: silica chromatography.
| Green Seaweed | Extract | Organisms | Ref. |
|---|---|---|---|
| Boodlea composita | M | V. harveyi, V. alginolyticus, V. vulnificus, V. parahaemolyticus, V. alcaligenes | [88] |
| Bryopsis pennata | M | V. vulnificus, V. parahaemolyticus | [88] |
| Caulerpa lentillifera | M/EA | E. coli, Staphylococcus aureus, Streptococcus sp., Salmonella sp. | [97] |
| Caulerpa parvula | M | V. vulnificus, V. alcaligenes | [88] |
| Caulerpa racemosa | M; M/DE; M/W | E. coli, S. aureus, Streptococcus sp., Salmonella sp. | [97] |
| Chaetomorpha aerea | W | Bacilus subtilis, Micrococcus luteus, S. aureus | [98] |
| Chaetomorpha linum | 2:1 C:M | V. ordalii, V. vulnificus | [45] |
| Cladophora albida | M | V. harveyi, V. alginolyticus, V. vulnificus, V. parahaemolyticus, V. alcaligenes | [88] |
| Cladophora glomerata | M | V. fischeri, V. vulnificus, V. anguillarum, V. parahaemolyticus | [99] |
| Cladophora rupestris | M, EA, E 2:1 C:M 2:1 C:M | E. coli, Pseudomonas aeruginosa, S. aureus, V. harveyii, V. parahaemolyticus, V. alginolyticus Enterococcus sp., Streptococcus agalactiae, V. fluvialis; V. salmonicida; V. vulnificus, V. ordalii; V. cholera non-O1; V. metschnikovii V. ordalii, V. salmonicida, V. vulnificus | [68] [65] [45] |
| Cladophora sp. | E | S. aureus, Streptococcus pyogenes | [86] |
| Codium amplivesiculatum | E | S. aureus, S. pyogenes | [86] |
| Codium cuneatum | E | S. aureus, S. pyogenes | [86] |
| Codium simulans | E | S. aureus, S. pyogenes | [86] |
| Enteropmorpha compressa | M | K. pneumoniae, V. harveyi, V. alginolyticus, V. vulnificus, V. parahaemolyticus, V. alcaligenes | [88] |
| E. compressa | E | Gardnerella vaginalis | [100] |
| E. compressa | E, A, M | B. subtilis, S. aureus, E. coli, S. typhi, K. pneumoniae, C. albicans | [82] |
| Enteromorpha intestinalis | A | Bacillus mycoides, B. subtilis, E. coli. K. pneumonia, S. aureus, A. flavus, A. fumigatus. C. albicans, P. purpurescens, P. verrucosum | [101] |
| Enteromorpha linza | E, A, M | B. subtilis, S. aureus, E. coli, S. typhi, K. pneumoniae, C. albicans | [82] |
| E. linza | 4:1 M:W E W/DCM | Prevotella intermedia, Porphyromonas gingivalis G. vaginalis E. coli, Salmonella. typhimurium | [102] [100] [103] |
| Laurencia johnstonii | 1:1 A:M–Er | S. aureus | [3] |
| Ulva dactilifera | E | S. aureus, S. pyogenes | [86] |
| Ulva fasciata | E, A, M | B. subtilis, S. aureus, E. coli, S. typhi, K. pneumoniae, C. albicans | [82] |
| Ulva lactuca | E, A, M | B. subtilis, S. aureus, E. coli, S. typhi, K. pneumoniae, C. albicans | [82] |
| U. lactuca | 1:1 A:M–Er | S. aureus | [3] |
| U. lactuca | A | B. mycoides, B. subtilis, E. coli. K. pneumoniae, S. aureus, A. flavus, A. fumigatus. C. albicans, P. purpurescens, P. verrucosum | [101] |
| Ulva pertusa | E | G. vaginalis | [100] |
| U. pertusa | M | G. vaginalis | [67] |
| Ulva prolifera | 2:1 C:M | V. ordali | [45] |
A: acetone; C: chloroform; DCM: dichloromethane; DE: diethylether; E: ethanol; EA: ethyl acetate; Er: ether; M: methanol; T: toluene; W: water.
| Brown Seaweed | Solvent | Organisms | Ref. |
|---|---|---|---|
| Chnoospora implexa | E | S. aureus, S. pyogenes | [86] |
| Cladophora rupestris | M | E. coli, S. aureus, P. aeruginosa, V. harveyii, V. parahaemolyticus, V. alginolyticus | [68] |
| C. rupestris | E | E. coli, S. aureus, P. aeruginosa, V. harveyii, V. parahaemolyticus, V. alginolyticus | [68] |
| C. rupestris | EA | E. coli, S. aureus, P. aeruginosa, V. harveyii, V. parahaemolyticus | [68] |
| Colpomenia sinuosa | E E, A, M | S. aureus, S. pyogenes B. subtilis, S. aureus, E. coli, S. typhi, K. pneumoniae, C. albicans | [86] [82] |
| Colpomenia tuberculata | E | S. aureus, Sreptococcus pyogenes | [86] |
| Cystoseira osmundacea | E | S. pyogenes | [86] |
| Cystoseira trinodis | M | S. aureus, B. subtilis, E. coli, P. aeruginosa | [83] |
| Dictyopteris delicatula | E | S. aureus, S. pyogenes | [86] |
| Dictyopteris undulata | E | S. aureus, S. pyogenes | [86] |
| Dictyota dichotoma | M | S. aureus, B. subtilis, E. coli, P. aeruginosa | [83] |
| Dictyota flabellata | E | S. aureus, S. pyogenes | [86] |
| Dictyota indica | M | S. aureus, B. subtilis, E. coli, P. aeruginosa | [83] |
| Dictyota sp. | 2:1 DCM:M | S. aureus, Enterococcus faecalis, P. aeruginosa | [94] |
| Eisenia bicyclis | M; M/H; M/DCM; M/B | S. aureus, S. epidermidis, Propionibacterium acnes | [104] |
| E. bicyclis | M/EA | S. aureus, S. epidermidis, P. acnes, P. aeruginosa | [104] |
| Fucus evanescens | EA | Hemophilus influenzae, Legionella pneumophila, Propionibacterium acnes S. pyogenes, Clostridium difficile, methicillin-resistant S. aureus | [105] |
| Himanthalia elongata | H, E, W (PLE) | Aspergillus niger, C. albicans, E. coli, S. aureus | [106] |
| H. elongata | W, M and mixtures | L. monocytogenes, E. faecalis, P. aeruginosa, S. abony | [107] |
| H. elongata | C,DE,H/ 60% M:W/1:1 W:EA | L. monocytogenes, S. abony | [54] |
| Hydroclathrus clathratus | E | S. aureus, S. pyogenes | [86] |
| Laminaria japonica | W | E. coli, S. aureus, B. cereus | [108] |
| Lobophora variegata | M | V. alginolyticus, V. vulnificus, V. parahaemolyticus, B. subtilis B.cereus, Micrococcus luteus, S. typhimurium, Aeromonas hydrophila, E. coli | [88] [109] |
| Padina concrescens | M E | V. alginolyticus, V. vulnificus, V. parahaemolyticus, B. subtilis S. aureus, S. pyogenes | [88] [86] |
| Padina mexicana | E | S. aureus, S. pyogenes | [86] |
| Padina pavonica | M | E. coli | [92] |
| Padina sanctae crucis | M, E | E. coli, S. aureus, P. aeruginosa, Candida tropicalis, C. kruzei | [110] |
| Padina tetrastomatica | M | V. alginolyticus, V. vulnificus, V. parahaemolyticus, B. subtilis | [88] |
| Rosenvingea intrincata | E | S. aureus, S. pyogenes | [86] |
| Saccharina japonica | scW + AcH | E. coli, S. typhimurium, S. aureus, B. cereus | [111] |
| S. japonica | H, E | E. coli, L. monocytogenes, S. aureus | [112] |
| S. japonica | SC-CO2 + E, 1:1 A:E | E. coli, L. monocytogenes, B. cereus, S. aureus, C. albicans, A. brasiliensis | [112] |
| Sargassum binderi | M, A | S. aureus, B. subtilis | [81] |
| S. binderi | EA | B. subtilis | [81] |
| Sargassum flavellum | M, A, EA | B. subtilis | [81] |
| Sargassum horneri | SC-CO2 + E, 1:1 A:E | E. coli, L. monocytogenes, B. cereus, S. aureus, C. albicans, A. brasiliensis | [112] |
| S. horneri | H, E | E. coli, L. monocytogenes, S. aureus | [112] |
| Sargassum horridum | E | S. aureus, S. pyogenes | [86] |
| Sargassum myriocystum | M | S. aureus, B. subtilis, E. coli, P. aeruginosa | [83] |
| Sargassum plagyophillum | M | B.subtilis, S. aureus | [81] |
| Sargassum vulgare | E, A, M | B. subtilis, S. aureus, E. coli, S. typhi, K. pneumoniae, C. albicans | [82] |
| Scytosiphon lomentaria | M | S. aureus, S. typhimurium, E. coli | [92] |
| Stoe-chospermum marginatum | M | S. aureus, B. subtilis, E. coli, P. aeruginosa | [83] |
| Turbinaria ornata | M | S. aureus, B. subtilis, E. coli, P. aeruginosa | [83] |
AcH: acetic acid; A: acetone; B: butanol; C: chloroform; DCM: dichloromethane; DE: diethylether; E: ethanol; EA: ethyl acetate; H: hexane; M: methanol; scW: subcritical water; PLE: pressurized liquid extraction; W: water; Sequence os solvents; / fraction.
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial Action of Compounds from Marine Seaweed. Mar. Drugs 2016, 14, 52. https://doi.org/10.3390/md14030052
AMA Style
Pérez MJ, Falqué E, Domínguez H. Antimicrobial Action of Compounds from Marine Seaweed. Marine Drugs. 2016; 14(3):52. https://doi.org/10.3390/md14030052
Chicago/Turabian StylePérez, María José, Elena Falqué, and Herminia Domínguez. 2016. "Antimicrobial Action of Compounds from Marine Seaweed" Marine Drugs 14, no. 3: 52. https://doi.org/10.3390/md14030052
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.