
New Species of Nectriaceae (Hypocreales) from China

State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101, China
*
Authors to whom correspondence should be addressed.
J. Fungi 2022, 8(10), 1075; https://doi.org/10.3390/jof8101075
Submission received: 14 September 2022
/
Revised: 6 October 2022
/
Accepted: 10 October 2022
/
Published: 13 October 2022
(This article belongs to the Special Issue Ascomycota: Diversity, Taxonomy and Phylogeny)
Abstract
:Species of Nectriaceae commonly occur on living and decaying woody substrates, soil, fruitbodies of other fungi, and insects. Some of them are reported as endophytes, opportunistic pathogens of crops and humans, or producers of mycotoxins. To explore the species diversity of the family, specimens from different regions of China were collected and examined. Four novel taxa of Penicillifer, Pseudocosmospora, and Thelonectria were introduced on the basis of morphological characteristics and DNA sequence analyses of combined datasets of the act, ITS, LSU, rpb1, rpb2, tef1, and tub2 regions. Differences between the new species and their close relatives were compared and discussed.
1. Introduction
The family Nectriaceae was established in 1865 to accommodate those fungi producing uniloculate perithecia that are yellow, orange-red to purple, or brown; often change color in 3% potassium hydroxide (KOH) and 100% lactic acid (LA); and have a tropical and subtropical distribution [1]. Approximately 69 genera are currently accepted [1,2,3], including genera such as Penicillifer Emden, Pseudocosmospora C.S. Herrera & P. Chaverri, and Thelonectria P. Chaverri & C. Salgado. The generic concepts and phylogenetic relationships of the family were comprehensively stated by Lombard et al. [2].
The genus Penicillifer, typified by Penicillifer pulcher Emden., was introduced by Emden [4]. It was treated as the asexual stage of Viridispora Samuels & Rossman [1] and recommended as the correct name for this group of fungi [2]. Seven species are currently known in the genus [1,2,5]. The genus Pseudocosmospora, typified by P. eutypellae C.S. Herrera & P. Chaverri, was established by Herrera et al. [6] to accommodate Cosmospora vilior (Starbäck) Rossman & Samuels and related species that are usually fungicolous. Sixteen species are recognized [6,7,8,9]. The genus Thelonectria, typified by T. discophora (Mont.) P. Chaverri & C. Salgado, was established by Chaverri et al. [10] to include the species formerly placed in the Nectria mammoidea and N. veuillotiana groups with a cosmopolitan distribution [11]. Forty-seven species are accepted in the genus [10,11,12,13,14,15,16,17,18].
In our study of the hypocrealean specimens from different regions of China, four unusual fungi were encountered. Judging by their perithecial gross morphology, anatomy, and culture characteristics, they represent four undescribed species of Penicillifer, Pseudocosmospora, and Thelonectria. Their taxonomic placements were further confirmed by multigene phylogenetic analyses of α-actin (act), nuclear ribosomal DNA ITS1-5.8S-ITS2 (ITS), large subunit of nuclear ribosomal DNA (LSU), the largest subunit of RNA polymerase II (rpb1), the second largest subunit of RNA polymerase II (rpb2), translation elongation factor 1-α (tef1), and β-tubulin (tub2). The differences between the novel taxa and their close relatives were compared.
2. Materials and Methods
2.1. Sampling and Morphological Studies
Specimens were collected from Beijing and the Guangxi Zhuang Autonomous Region, and they are preserved in Herbarium Mycologicum Academiae Sinicae (HMAS). Cultures were obtained by single ascospore isolation from the fresh perithecium and deposited in the China General Microbiological Culture Collection Center (CGMCC). The method of Lombard et al. [2] was followed for morphological observations. The ascomatal wall reactions to 3% KOH and 100% LA were tested. Sections were prepared with a freezing microtome (YD-1508-III, Jinhua, China) at a thickness of 6–8 μm for anatomic examination. Lactophenol cotton blue solution was used as a mounting medium for the measurements of the perithecia, asci, ascospores, conidiophores, and conidia. Photographs were taken with a Leica DFC450 digital camera (Wetzlar, Germany) attached to a Leica M125 stereomicroscope (Milton Keynes, UK) for gross morphology and a Zeiss AxioCam MRc 5 digital camera (Jena, Germany) attached to a Zeiss Axio Imager A2 microscope (Göttingen, Germany) for microscopic features. For colony morphology and growth rates, strains were grown on potato dextrose agar (PDA, 20% w/v potato + 2% w/v dextrose + 2% w/v agar) and synthetic nutrient-poor agar (SNA) [19] in 90 mm plastic Petri dishes at 25 °C for 14 d with alternating periods of light and darkness (12 h/12 h).
2.2. DNA Extraction, PCR Amplification, Sequencing, and Phylogenetic Analyses
Genomic DNA was extracted from fresh mycelium following the method of Lombard et al. [2]. Seven primer pairs, act1/act2 [20], ITS5/ITS4 [21], LR0R/LR5 [22,23], rpb1a/rpb1c [24], RPB2-5f/RPB2-7cR [25], 728F/EF2 [26,27], and T1/T22 [28], were used to amplify the sequences of the act, ITS, LSU, rpb1, rpb2, tef1, and tub2 regions, respectively. PCR reactions were performed using an ABI 2720 Thermal Cycler (Applied Biosciences, Foster City, CA, USA), and DNA sequencing was carried out in both directions on an ABI 3730XL DNA Sequencer (Applied Biosciences, Foster City, CA, USA).
Newly obtained sequences and those retrieved from GenBank are listed in Table 1, Table 2 and Table 3. The sequences were assembled and aligned, and the primer sequences were trimmed using BioEdit 7.0.5 [29] and converted to nexus files by ClustalX 1.83 [30]. A partition homogeneity test (PHT) was performed with 1000 replicates in PAUP*4.0b10 [31] to evaluate statistical congruence amongst these loci. The aligned sequences were combined in BioEdit and analyzed with Bayesian inference (BI), maximum likelihood (ML), and maximum parsimony (MP) methods to determine the phylogenetic positions of the new species. The BI analysis was conducted by MrBayes 3.1.2 [32] using a Markov chain Monte Carlo algorithm. Nucleotide substitution models were determined by MrModeltest 2.3 [33]. Four Markov chains were run simultaneously for 1,000,000 generations, with the trees sampled every 100 generations. A 50% majority rule consensus tree was computed after excluding the first 2500 trees as ‘burn-in’. The Bayesian inference posterior probability (BIPP) was determined from the remaining trees. Branch support measures were calculated with 1000 bootstrap replicates. The ML analysis was performed via IQ-Tree 1.6.12 [34] using the best model for each locus chosen by ModelFinder [35]. The MP analysis was performed with PAUP 4.0b10 [31] using heuristic searches with 1000 replicates of random addition of sequences and subsequent TBR (tree bisection and reconnection) branch swapping. The topological confidence of the resulting trees and the statistical supports of the branches were tested by maximum parsimony bootstrap proportion (MPBP) with 1000 replications, each with 10 replicates of random addition of taxa. Trees were examined by TreeView 1.6.6 [36]. Maximum likelihood bootstrap proportion (MLBP) and MPBP greater than 70% and BIPP greater than 90% were shown at the nodes.
3. Results
3.1. Phylogeny
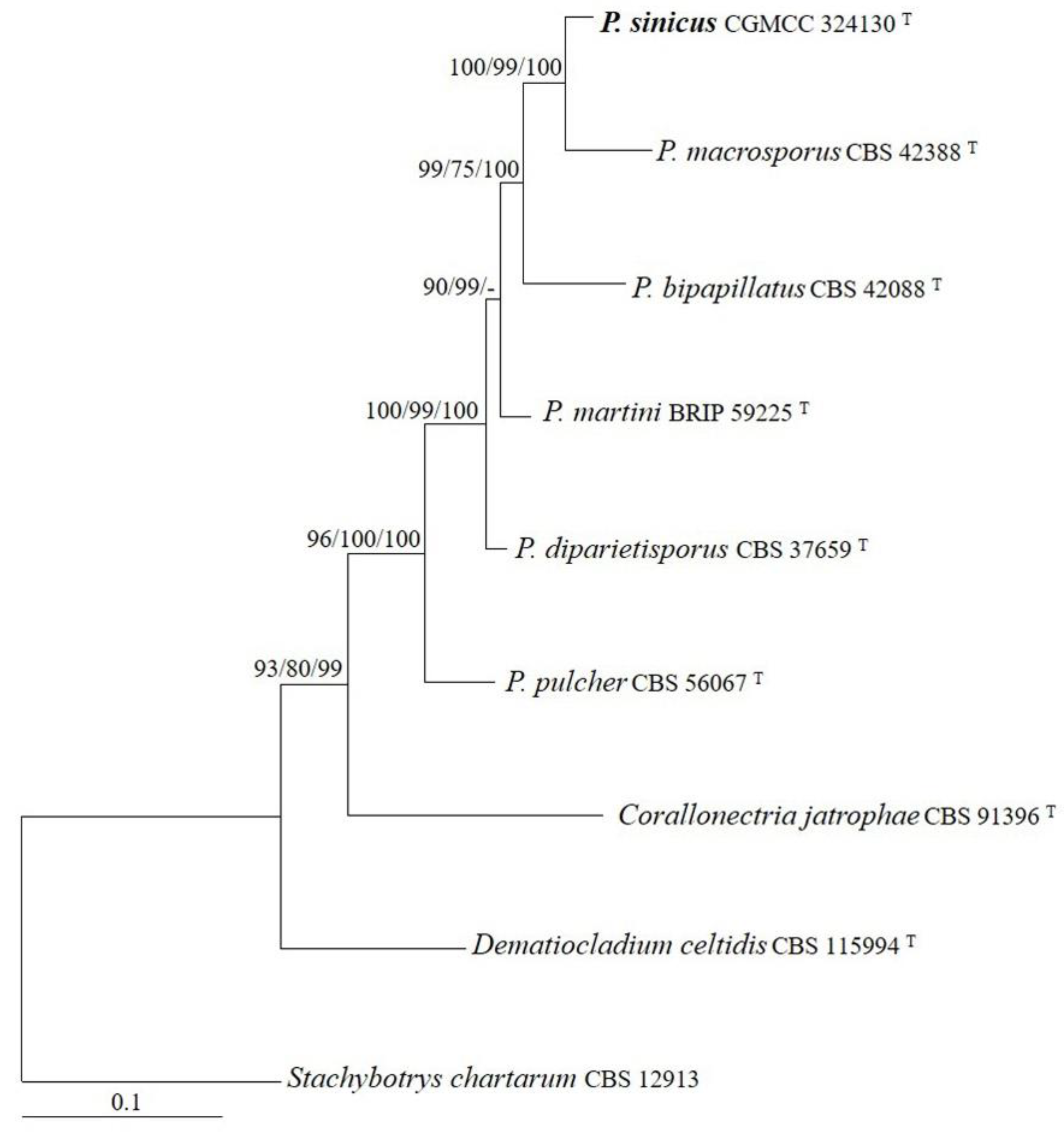
The sequences of the ITS, LSU, rpb2, and tef1 regions from six Penicillifer species were analyzed. Stachybotrys chartarum (Ehrenb.) S. Hughes was used as the outgroup taxon. The partition homogeneity test (p = 0.01) indicated that the individual partitions were not highly incongruent [37]; thus, these four loci were combined for the phylogenetic analyses. The ML tree is shown in Figure 1. The topologies of the BI and MP trees were similar to that of the ML tree. The isolate CGMCC 3.24130 grouped with other members of Penicillifer and received high statistical support (MLBP/MLBP/BIPP = 96%/100%/100%).
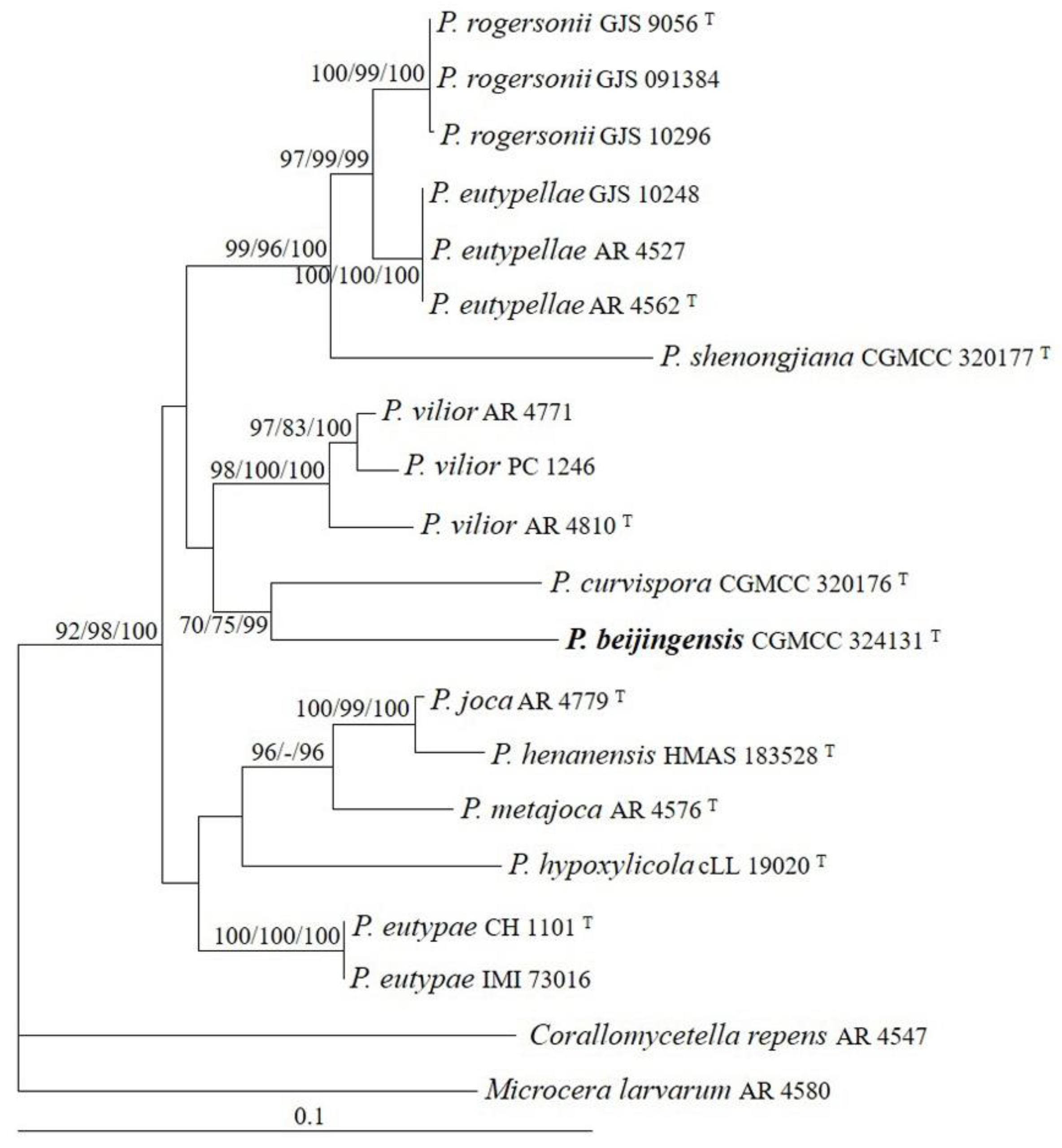
The sequences of ITS, LSU, and tub2 regions from 11 Pseudocosmospora species were analyzed. Corallomycetella repens (Berk. & Broome) Rossman & Samuels and Microcera larvarum (Fuckel) Gräfenhan, Seifert & Schroers were used as outgroup taxa. The partition homogeneity test (p = 0.01) indicated that the individual partitions were not highly incongruent [37]; thus, these three loci were combined for the phylogenetic analyses. The ML tree is shown in Figure 2. The topologies of the BI and MP trees were similar to that of the ML tree. The isolate CGMCC 3.24131 grouped with other species of Pseudocosmospora and received high statistical support (MLBP/MLBP/BIPP = 92%/98%/100%).
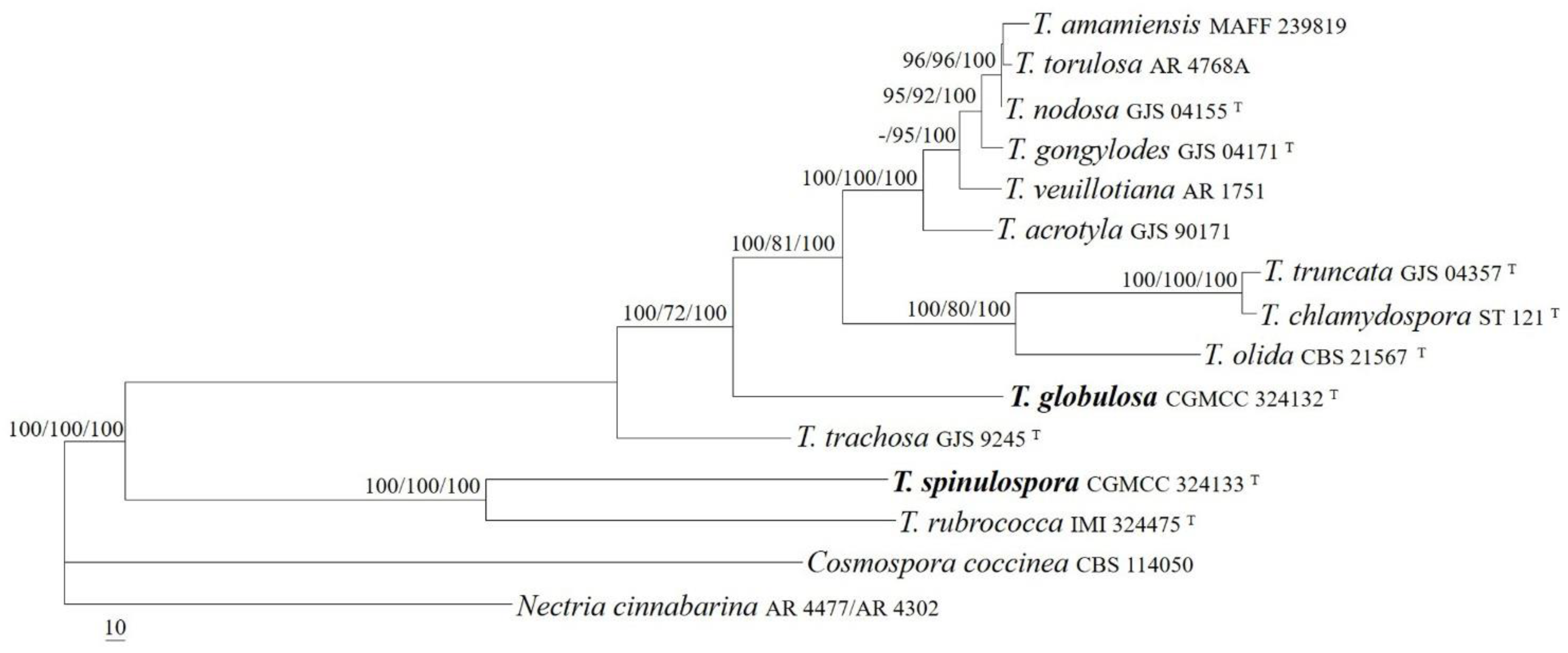
The sequences of the act, ITS, LSU, rpb1, and tub2 regions from 13 Thelonectria species were analyzed. Cosmospora coccinea Rabenh. and Nectria cinnabarina (Tode) Fr. were used as outgroup taxa. The partition homogeneity test (p = 0.01) indicated that the individual partitions were not highly incongruent [37]; thus, these five loci were combined for the phylogenetic analyses. The ML tree is shown in Figure 3. The topologies of the BI and MP trees were similar to that of ML tree. The isolates CGMCC 3.24132 and CGMCC 3.24133 were well-located among other Thelonectria species and received high supporting values (MLBP/MLBP/BIPP = 100%/100%/100%). The isolate CGMCC 3.24133 was related to T. rubrococca (Brayford & Samuels) Salgado & P. Chaverri, receiving high statistic values (MLBP/MLBP/BIPP = 100%/100%/100%), and the strain CGMCC 3.24132 formed an independent lineage and was related to the T. veuillotiana complex (MLBP/MLBP/BIPP = 100%/72%/100%).
3.2. Taxonomy
Fungal Names: FN571297.
Etymology: The epithet refers to the country where the fungus was collected.
Typification: CHINA, Guangxi Zhuang Autonomous Region, Guilin City, Mao’er Mountain, on rotten twigs, 7 December 2019, Z.Q. Zeng & H.D. Zheng 12496 (holotype HMAS 247865, ex-type strain CGMCC 3.24130).
GenBank accession numbers: ITS OP223439, LSU OP223435, rpb2 OP272863, tef1 OP272864, rpb1 OP586759, tub2 OP586763.
Mycelium was not visible on the natural substratum. Perithecia were superficial, solitary, non-stromatic or with a basal stroma, and subglobose to globose with an acute to blunt papilla; the surface was warted; they did not collapse upon drying; they were yellowish brown to brown and did not change color in 3% KOH or 100% LA, and the size was 235–314 × 176–274 μm. The perithecial surface had warts 15–55 μm high. Perithecial walls were two-layered, 23–35 μm thick; the outer layer was of the textura angularis, 16–25 μm thick, with cell 5–9 × 4–8 μm, and cell walls 1–1.2 μm thick; the inner layer was of the textura prismatica, 7–10 μm thick, with cell 5–13 × 2–3 μm, and cell walls 0.8–1 μm thick. Asci were unitunicate, cylindrical, and eight-spored with an apical ring, and 60–85 × 4.5–8 μm. Ascospores were ellipsoidal to fusiform, (0–)1-septate, constricted or not at septum, hyaline to light brown, smooth-walled, uniseriate, overlapping obliquely, and 10–15 × 4.5–5.3 μm.
Colony characteristics: On PDA, the colony was 20 mm in diam. after 1 week at 25 °C, the surface was cottony with dense, whitish aerial mycelium producing yellowish-brown pigments. On SNA, the colony was 26 mm in diam. after 1 week at 25 °C, the surface was velvet with sparse, whitish aerial mycelium. Conidiophores were verticillium-like, septate, and hyaline, with 1–2 whorls and a terminal whorl of 2–8 phialides, 30–120 μm long, and 2–3.5 μm wide at the base. Phialides were subulate, tapering toward the apex, 15–45 μm long, 1.5–2.5 μm wide at the base, and 0.2–0.3 μm wide at the apex. Macroconidia were ellipsoidal to fusiform or cylindrical, slightly curved, (0–)1(–3)-septate, smooth-walled, hyaline, and 10–28 × 3–5.5 μm.
Notes: Amongst the known species of the genus, P. sinicus is morphologically most similar to P. macrosporus Samuels in having superficial, solitary, non-stromatic, globose perithecia with warted surfaces; smooth-walled, 1-septate ascospores; and cylindrical, bicellular conidia [1]. However, P. macrosporus has a thicker perithecial wall (ca. 65 μm thick), clavate asci without apical rings, wider ascospores (5–7 μm wide), and longer conidia (33–47 μm long) [1]. In addition, there were 34 bp, 19 bp, 23 bp, 31 bp, and 23 bp divergences in the ITS, LSU, rpb1, rpb2, and tef1 regions between the ex-type cultures of the two species (CGMCC 3.24130 and CBS 423.88). Thus, both the morphological and the molecular evidence support their separation at the species level.
Fungal Names: FN571298.
Etymology: The epithet refers to the type locality of the fungus.
Typification: CHINA, Beijing, Beidagou forest, on rotten bark associated with other fungi, 10 August 2017, H.D. Zheng, X.C. Wang, Y.B. Zhang, C. Wang & P. Li 11339 (holotype HMAS 290896, ex-type strain CGMCC 3.24131).
GenBank accession numbers: ITS OP223438, LSU OP223434, tub2 OP272862.
Mycelium was not visible on the natural substratum. Perithecia were superficial and gregarious with a well-developed stroma, subglobose to globose, slightly roughened surface, laterally collapsed upon drying, orange-red to bright red, turning dark red in 3% KOH and light yellow in 100% LA, and 147–196 × 118–176 μm. Perithecial walls were two-layered, and 20–42 μm thick; the outer layer was of the textura globulosa to textura angularis, 15–25 μm thick, with cell 4–13 × 2.5–4.5 μm, and cell walls 1–1.2 μm thick; the inner layer was of the textura prismatica, 5–8 μm thick, with cell 6–10 × 2.5–3.5 μm, and cell walls 0.8–1 μm thick. Asci were unitunicate, cylindrical, with a simple apex, eight-spored, and 38–58 × 2.5–5 μm. Ascospores were ellipsoidal, 1-septate, not constricted at the septum, light yellow-brown, smooth-walled, uniseriate, and 8–10 × 2.5–4 μm.
Colony characteristics: On PDA, the colony was 25 mm in diam. after 1 week at 25 °C, surface crustose, producing yellowish-white pigments. On SNA, the colony was 15 mm in diam. after 1 week at 25 °C, surface velvet, with sparse, whitish aerial mycelium. Conidiophores were acremonium- to verticillium-like, septate, of indefinite length, and hyaline, with 1–2 whorls and a terminal whorl of 2–6 phialides. Phialides were subulate, tapering toward the apex, 10–55 μm long, 0.9–1.2 μm wide at the base, and 0.2–0.3 μm wide at the tip. Conidia were allantoid, curved, unicellular, smooth-walled, hyaline, and 2.6–4.5 × 0.9–1.8 μm.
Notes: Among the known species of Pseudocosmospora, P. beijingensis most resembles P. curvispora Z.Q. Zeng & W.Y. Zhuang in having subglobose to globose perithecia that are laterally collapsed upon drying; having asci without an apical ring; having ellipsoidal, 1-septate, smooth-walled, and light yellow-brown ascospores; and producing acremonium- to verticillium-like conidiophores, and allantoid, unicellular, curved conidia [8]. However, P. curvispora differs in clavate and somewhat longer asci (53–68 μm long) and narrower conidia (0.8–1.2 μm wide) [8]. Sequence comparisons revealed that there were 30 bp, 22 bp, and 90 bp divergences detected for the ITS, LSU, and tub2 regions. Obviously, they are not conspecific.
Fungal Names: FN571299.
Etymology: The epithet refers to the globose microconidia.
Typification: CHINA, Guangxi Zhuang Autonomous Region, Guilin City, Mao’er Mountain, on rotten roots, 5 December 2019, Z.Q. Zeng & H.D. Zheng 12434 (holotype HMAS 255835, ex-type strain CGMCC 3.24132).
GenBank accession numbers: act OP272865, ITS OP223436, LSU OP223432, rpb1 OP272867, rpb2 OP586760, tub2 OP586762.
Mycelium was not visible on the natural substratum. Perithecia were superficial, solitary to gregarious, with a basal or well-developed stroma, subglobose to globose, slightly roughened surface, with blunt papilla of 32–65 μm high and 52–75 μm wide at the base, did not collapse upon drying, orange-red to red, turning dark red in 3% KOH and light yellow in 100% LA, and 235–323 × 148–245 μm. Perithecial walls were two-layered, 20–40 μm thick; the outer layer was of the textura globulosa to textura angularis, 15–30 μm thick, with cell 5–15 × 4–12 μm, and cell walls 0.8–1 μm thick; the inner layer was of the textura prismatica, 5–10 μm thick, with cell 6–15 × 3–10 μm, and cell walls 1–1.2 μm thick. Asci were unitunicate, cylindrical to clavate, eight-spored, with a simple apex, and 53–75 × 8–13 μm. Ascospores were ellipsoidal to fusiform, 1-septate, constricted at the septum, hyaline, smooth to spinulose, uniseriate or irregular biseriate in asci, and 13–20 × 5.5–8 μm.
Colony characteristics: The colony on PDA was 22 mm in diam. after 1 week at 25 °C and had a cottony surface with dense, whitish aerial mycelium producing yellowish brown pigments. The colony on SNA was 35 mm in diam. after 1 week at 25 °C and had a cottony surface with sparse, whitish aerial mycelium. Conidiophores were mostly unbranched; rarely had simple branches; and were septate, hyaline, 25–89 μm long, and 1.5–2.5 μm wide at the base. Macroconidia were cylindrical to rod-shaped, slightly curved, 1–3(–4)-septate, smooth-walled, hyaline, and 20–58 × 3.2–5.8 μm. Microconidia were globose, smooth-walled, hyaline, and 3–4.5 μm in diam. Chlamydospores were globose to subglobose, and 4–10 × 3–8 μm.
Notes: Among the known species of Thelonectria, T. globulosa is distinct because of its globose microconidia. Morphologically, T. globulosa resembles T. nodosa C.G. Salgado & P. Chaverri in having solitary to gregarious, globose perithecia that do not collapse upon drying; cylindrical to clavate asci; ellipsoidal to fusiform ascospores; and cylindrical macroconidia. However, T. nodosa differs because of its larger asci (68–115 × 10–17 μm) with an apical ring, macroconidia possessing more septa (up to six septa), and lack of microconidia formation [12]. Moreover, there are 38 bp, 96 bp, 37 bp, and 65 bp divergences in the act, ITS, LSU, and rpb1 regions between the ex-type cultures of the two taxa (CGMCC 3.24132 and GJS 04155). Both the morphology and DNA sequence data distinguish them as different species.
Fungal Names: FN571300.
Etymology: The specific epithet refers to the spinulose ascospores.
Typification: CHINA, Guangxi Zhuang Autonomous Region, Guilin City, Mao’er Mountain, on rotten twigs, 7 December 2019, Z.Q. Zeng & H.D. Zheng 12499 (holotype HMAS 290897, ex-type strain CGMCC 3.24133).
GenBank accession numbers: act OP272866, ITS OP223437, LSU OP223433, rpb1 OP272868, rpb2 OP586761, tub2 OP586764.
Mycelium was not visible on the natural substratum. Perithecia were superficial, solitary, with a basal stroma, subglobose to broad-pyriform, surface slightly roughened, sometimes collapsed laterally upon drying, orange-red to red, turning dark red in 3% KOH and light yellow in 100% LA, and 123–195 × 143–212 μm. Perithecial walls were two-layered, 8–18 μm thick; the outer layer was of the textura globulosa to textura angularis, 5–13 μm thick, with cell 6–10 × 4–9 μm, and cell walls 1–1.2 μm thick; the inner layer was of the textura prismatica, 3–5 μm thick, with cell 2–8 × 3–10 μm, and cell walls 0.8–1 μm thick. Asci were not observed. Ascospores were ellipsoidal, 1-septate, not constricted or slightly constricted at the septum, hyaline, spinulose, and 12–18 × 5.6–8 μm.
Colony characteristics: On PDA, the colony was 40 mm in diam. after 2 weeks at 25 °C, the surface was cottony with dense, whitish aerial mycelium producing light yellow pigments. On SNA, the colony was 28 mm in diam. after 2 weeks at 25 °C, surface velvet, with sparse, whitish aerial mycelium. Conidiophores were acremonium-like, rarely with simple branches, septate, hyaline, and 45–102 × 2.2–4 μm. Macroconidia were cylindrical, slightly curved, (1–2–)3-septate, smooth-walled, hyaline, and 28–62 × 2.8–4.5 μm. Chlamydospores were globose to subglobose, smooth-walled, hyaline, and 6–8 μm in diam.
Notes: The morphological features, such as the superficial, globose to subglobose, broad-pyriform perithecia that do not collapse when dry; ellipsoidal, two-celled, and hyaline ascospores; and curved macroconidia with rounded ends, indicate the placement of T. spinulospora in Thelonectria, which was confirmed by sequence analyses of the act, ITS, LSU, rpb1, and tub2 regions (Figure 3). Amongst the known species of the genus, the new species is morphologically similar and phylogenetically related to T. rubrococca (Brayford & Samuels) C.G. Salgado & P. Chaverri in having solitary to gregarious, globose perithecia that do not collapse upon drying, ellipsoidal ascospores, and cylindrical macroconidia. However, the latter differs in its larger perithecia (200–450 μm in diam.), smaller ascospores (8–14.5 × 3.6–6.6 μm), and macroconidia with more septa (up to five septa) [38]. Sequence comparisons between the ex-type cultures of the two species revealed that 24 bp, 8 bp, 0 bp, 22 bp, and 28 divergences were detected for the act, ITS, LSU, rpb1, and tub2 regions. Both the morphology and DNA sequence data support their distinction at the species level.
4. Discussion
The genus Penicillifer is proposed as the preferable name over Viridispora [2], following the International Code of Nomenclature for algae, fungi, and plants [39]. Our analyses, inferred from sequences of ITS, LSU, rpb2, and tef1 and including the new taxon, revealed a tree topology (Figure 1) similar to that given by Lombard et al. [2]. The phylogenetic tree shows that Penicillifer species forms a well-supported monophyletic clade (MLBS/MPBP/BIPP/ = 96%/100%/100%) (Figure 1). Penicillifer sinicus is closely related to P. macrosporus (MLBS/MPBP/BIPP/ = 100%/99%/100%). The sequence comparisons revealed that there were 34 bp, 19 bp, 23 bp, 31 bp, and 23 bp differences detected for the ITS, LSU, rpb1, rpb2, and tef1 regions. Therefore, both the molecular and the morphological evidence supports the separation of the two fungi at a specific level. Among the known species of Penicillifer, P. martinii P. Wong, Y.P. Tan & R.G. Shivas is known solely by its sexual stage [5], and only asexual stages of P. japonicus Matsush. and P. pulcher have been discovered [4,40]; the rest species of the genus are holomorphic, including the newly added one.
Historically, nectriaceous species producing small, reddish, smooth, thin-walled perithecia were categorized as Cosmospora Rabenh. sensu lato [1]. The accumulated morphological and phylogenetic information indicated that the genus was not monophyletic [41,42]. Herrera et al. [6] established Pseudocosmospora to accommodate ten cosmospora-like fungi on Eutypa and Eutypella and with acremonium- to verticillium-like asexual stages. Since then, six additional taxa have joined the group [7,8,9]. The genus has become distributed worldwide and displays high species diversity in warm temperate and tropical regions [6]. Species of the genus have the following features in common: they are superficial perithecia, gregarious, KOH+, LA+, laterally collapsed upon drying, and usually less than 250 μm in height; and they have asci containing eight 1-septate ascospores, acremonium- to verticillium-like conidiophores, and non-septate conidia. Pseudocosmospora beijingensis fits well the generic concept. The multigene analyses indicated its distinctions from any other species of the genus (Figure 2).
Members of Thelonectria are often found on twigs and branches, trunks of recently killed or dying trees, and rotting roots; they occasionally cause small cankers and are mainly distributed in tropical, subtropical, and temperate regions [10,11,17]. Among the species of the genus, T. coronata (Penz. & Sacc.) P. Chaverri & C. Salgado, T. discophora, T. lucida (Höhn.) P. Chaverri & C. Salgado, and T. veuillotiana (Roum. & Sacc.) P. Chaverri & Salgado are cosmopolitan and are treated as species complexes [38,43,44]. Salgado-Salazar et al. [11,12,13] carried out a revisionary work on the above species complexes and described 30 cryptic species on the basis of genealogical concordance phylogenetic species recognition. Our phylogenetic results indicated that T. globulosa was associated with but clearly separated from members of the T. veuillotiana complex. Thelonectria aurea, known only by the asexual stage, can be easily distinguished in the absence of microconidia and chlamydospores in culture [17]. Moreover, there were 93 bp and 64 bp divergences in the ITS and tub2 regions between the ex-type culture of the two species.
There are 47 species currently known in this genus, of which 20 species have been reported in China [11,13,15]. Large-scale surveys of fungal resources in various regions with different climates, vegetation, geographic structures, and multiple niches will improve our understanding of the species diversity of nectriaceous fungi in the country.
5. Conclusions
The species diversity of the family Nectriaceae was investigated, and four novel taxa were discovered. With the joining of the new species, the phylogenetic relationships among species of these three genera were updated.
Author Contributions
Conceptualization, W.-Y.Z.; methodology, Z.-Q.Z.; software, Z.-Q.Z.; validation, Z.-Q.Z.; formal analysis, Z.-Q.Z.; investigation, Z.-Q.Z.; resources, W.-Y.Z. and Z.-Q.Z.; data curation, Z.-Q.Z.; writing—original draft preparation, Z.-Q.Z.; writing—review and editing, W.-Y.Z. and Z.-Q.Z.; visualization, Z.-Q.Z.; supervision, Z.-Q.Z. and W.-Y.Z.; project administration, W.-Y.Z.; funding acquisition, W.-Y.Z. and Z.-Q.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (31750001, 31870012) and the Frontier Key Program of the Chinese Academy of Sciences (QYZDY-SSW-SMC029).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Names of the new species were formally registered in the database Fungal Names (https://nmdc.cn/fungalnames (accessed on 22 August 2022)). Specimens were deposited in the Herbarium Mycologicum Academiae Sinicae (HMAS). The newly generated sequences were deposited in GenBank (https://www.ncbi.nlm.nih.gov/genbank (accessed on 5 October 2022)).
Acknowledgments
The authors would like to thank Huan-Di Zheng, Xin-Cun Wang, Yu-Bo Zhang, Chao Wang, and Peng Li for collecting specimens jointly and Ting Huang for technique help with this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rossman, A.Y.; Samuels, G.J.; Rogerson, C.T.; Lowen, R. Genera of Bionectriaceae, Hypocreaceae, and Nectriaceae (Hypocreales, Ascomycetes). Stud. Mycol. 1999, 42, 1–260. [Google Scholar]
- Lombard, L.; van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- van Emden, J.H. Penicillifer, a new genus of Hyphomycetes from soil. Acta Bot. Neerl. 1968, 17, 54–58. [Google Scholar] [CrossRef]
- Crous, P.W.; Shivas, R.G.; Quaedvlieg, W.; van der Bank, M.; Zhang, Y.; Summerell, B.A.; Guarro, J.; Wingfield, M.J.; Wood, A.R.; Alfenas, A.C.; et al. Fungal Planet description sheets: 214–280. Persoonia 2014, 32, 184–306. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.S.; Rossman, A.Y.; Samuels, G.J.; Chaverri, P. Pseudocosmospora, a new genus to accommodate Cosmospora vilior and related species. Mycologia 2013, 105, 1287–1305. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.Q.; Zhuang, W.Y. Eight new combinations of Bionectriaceae and Nectriaceae. Mycosystema 2017, 36, 278–281. [Google Scholar]
- Zeng, Z.Q.; Zhuang, W.Y. Our current understanding of the genus Pseudocosmospora (Hypocreales, Nectriaceae) in China. Mycol. Prog. 2021, 20, 419–429. [Google Scholar] [CrossRef]
- Lechat, C.; Fournier, J. Pseudocosmospora hypoxylicola (Nectriaceae), a new species from the French Alps. Ascomycete.org 2020, 12, 6–10. [Google Scholar]
- Chaverri, P.; Salgado, C.; Hirooka, Y.; Rossman, A.Y.; Samuels, G.J. Delimitation of Neonectria and Cylindrocarpon (Nectriaceae, Hypocreales, Ascomycota) and related genera with Cylindrocarpon-like anamorphs. Stud. Mycol. 2011, 68, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Salazar, C.; Rossman, A.Y.; Chaverri, P. The genus Thelonectria (Nectriaceae, Hypocreales, Ascomycota) and closely related species with Cylindrocarpon-like asexual states. Fungal Divers. 2016, 80, 411–455. [Google Scholar] [CrossRef]
- Salgado-Salazar, C.; Rossman, A.; Samuels, G.J.; Capdet, M.; Chaverri, P. Multigene phylogenetic analyses of the Thelonectria coronata and T. veuillotiana species complexes. Mycologia 2012, 104, 1325–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgado-Salazar, C.; Rossman, A.Y.; Samuels, G.J.; Hirooka, Y.; Sanchez, R.M.; Chaverri, P. Phylogeny and taxonomic revision of Thelonectria discophora (Ascomycota, Hypocreales, Nectriaceae) species complex. Fungal Divers. 2015, 70, 1–29. [Google Scholar] [CrossRef]
- Zeng, Z.Q.; Zhuang, W.Y. Four new taxa of Ilyonectria and Thelonectria (Nectriaceae) revealed by morphology and combined ITS and β-tubulin sequence data. Phytotaxa 2013, 85, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.Q.; Zhuang, W.Y. The genera Rugonectria and Thelonectria (Hypocreales, Nectriaceae) in China. MycoKeys 2019, 55, 101–120. [Google Scholar] [CrossRef]
- Crous, P.W.; Luangsa-ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal Planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Nouri, M.T.; Trouillas, F.P. Taxonomy and multi-locus phylogeny of cylindrocarpon-like species associated with diseased roots of grapevine and other fruit and nut crops in California. Fungal Syst. Evol. 2019, 4, 59–75. [Google Scholar] [CrossRef]
- Braun, U.; Bensch, K. Annotated list of taxonomic novelties published in “Fungi Rhenani Exsiccati” Supplementi Fasc. 1 to 5, issued by K. W. G. L. Fuckel between 1865 and 1867. Schlechtendalia 2020, 37, 80–133. [Google Scholar]
- Nirenberg, H.I. Studies on the morphologic and biologic differentiation in Fusarium section Liseola. Mitt. Biol. Bundesanst. Land-Forstw. 1976, 169, 1–117. [Google Scholar]
- Samuels, G.J.; Dodd, S.; Lu, B.S.; Petrini, O.; Schroers, H.J.; Druzhinina, I.S. The Trichoderma koningii aggregate species. Stud. Mycol. 2006, 56, 67–133. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfland, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analyzed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Matheny, P.B.; Liu, Y.J.; Ammirati, J.F.; Hall, B.D. Using RPB1 sequences to improve phylogenetic inference among mushrooms (Inocybe, Agaricales). Am. J. Bot. 2002, 89, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA Polymerase II Subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgin, D.G. The ClustalX windows interface: Flexible strategies for multiple sequences alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4883. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP 4.0b10: Phylogenetic Analysis Using Parsimony (* and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Mol. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, R.D. TreeView: An application to display phylogenetic trees on personal computers. Comp. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
- Cunningham, C.W. Can three incongruence tests predict when data should be combined? Mol. Biol. Evol. 1997, 14, 733–740. [Google Scholar] [CrossRef]
- Brayford, D.; Samuels, G.J. Some didymosporous species of Nectria with nonmicroconidial Cylindrocarpon anamorphs. Mycologia 1993, 85, 612–637. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.H.; Li, D.Z.; Marhold, K.; et al. International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code); Koeltz Scientific Books: Berlin/Heidelberg, Germany, 2018; pp. 1–254. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs No 4; Published by the author; Kobe, Japan, 1985; pp. 1–68. [Google Scholar]
- Zhang, X.M.; Zhuang, W.Y. Phylogeny of some genera in the Nectriaceae (Hypocreales, Ascomycetes) inferred from 28S nrDNA partial sequences. Mycosystema 2006, 25, 15–22. [Google Scholar]
- Luo, J.; Zhuang, W.Y. Chaetopsinectria (Nectriaceae, Hypocreales), a new genus with Chaetopsina anamorphs. Mycologia 2010, 102, 976–984. [Google Scholar] [CrossRef]
- Samuels, G.J.; Doi, Y.; Rogerson, C.T. Contributions toward a mycobiota of Indonesia: Hypocreales. Mem. N. Y. Bot. Gard. 1990, 59, 6–108. [Google Scholar]
- Samuels, G.J.; Brayford, D. Species of Nectria (sensu lato) with red perithecia and striate ascospores. Sydowia 1994, 46, 75–161. [Google Scholar]
Figure 1.
The maximum likelihood tree inferred from combined ITS, LSU, rpb2, and tef1 sequences of representative species of Penicillifer. MLBP (left) and MPBP (middle) values greater than 70% and BIPP (right) values greater than 90% are shown at the nodes.
Figure 1.
The maximum likelihood tree inferred from combined ITS, LSU, rpb2, and tef1 sequences of representative species of Penicillifer. MLBP (left) and MPBP (middle) values greater than 70% and BIPP (right) values greater than 90% are shown at the nodes.

Figure 2.
The maximum likelihood tree inferred from combined ITS, LSU, and tub2 sequences of representative species of Pseudocosmospora. MLBP (left) and MPBP (middle) values greater than 70% and BIPP (right) values greater than 90% were shown at the nodes.
Figure 2.
The maximum likelihood tree inferred from combined ITS, LSU, and tub2 sequences of representative species of Pseudocosmospora. MLBP (left) and MPBP (middle) values greater than 70% and BIPP (right) values greater than 90% were shown at the nodes.

Figure 3.
The maximum likelihood tree inferred from combined act, ITS, LSU, rpb1, and tub2 sequences of representative species of Thelonectria. MLBP (left) and MPBP (middle) values greater than 70% and BIPP (right) values greater than 90% were shown at the nodes.
Figure 3.
The maximum likelihood tree inferred from combined act, ITS, LSU, rpb1, and tub2 sequences of representative species of Thelonectria. MLBP (left) and MPBP (middle) values greater than 70% and BIPP (right) values greater than 90% were shown at the nodes.

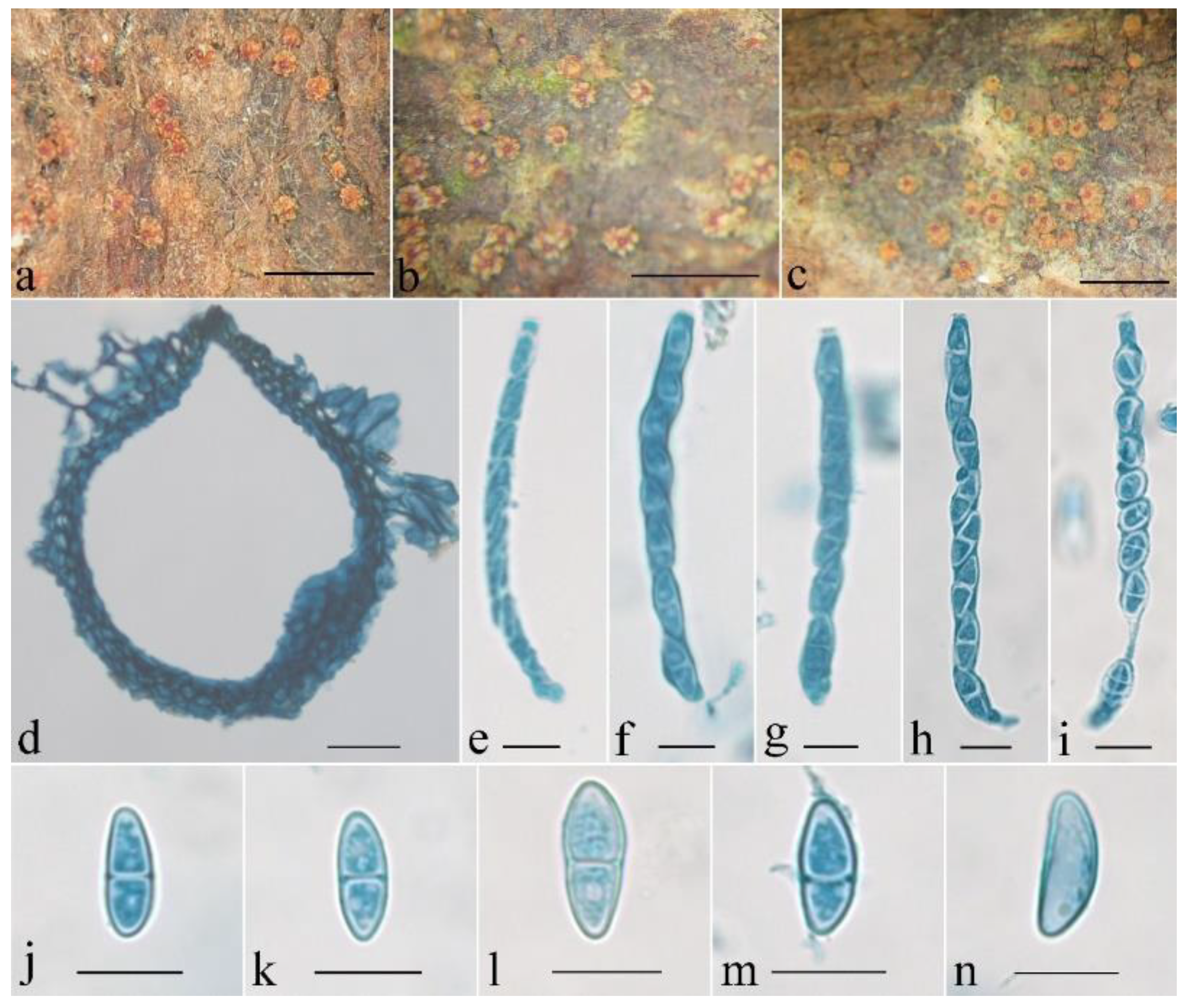
Figure 4.
Macroscopic and microscopic morphology of Penicillifer sinicus (HMAS 247865). (a–c) Ascomata on natural substratum; (d) median section of perithecium in lactophenol cotton blue; (e–i) asci with ascospores in lactophenol cotton blue; (j–n) ascospore in lactophenol cotton blue. Bars: (a–c) = 1 mm; (d) = 50 μm; (e–n) = 10 μm.
Figure 4.
Macroscopic and microscopic morphology of Penicillifer sinicus (HMAS 247865). (a–c) Ascomata on natural substratum; (d) median section of perithecium in lactophenol cotton blue; (e–i) asci with ascospores in lactophenol cotton blue; (j–n) ascospore in lactophenol cotton blue. Bars: (a–c) = 1 mm; (d) = 50 μm; (e–n) = 10 μm.

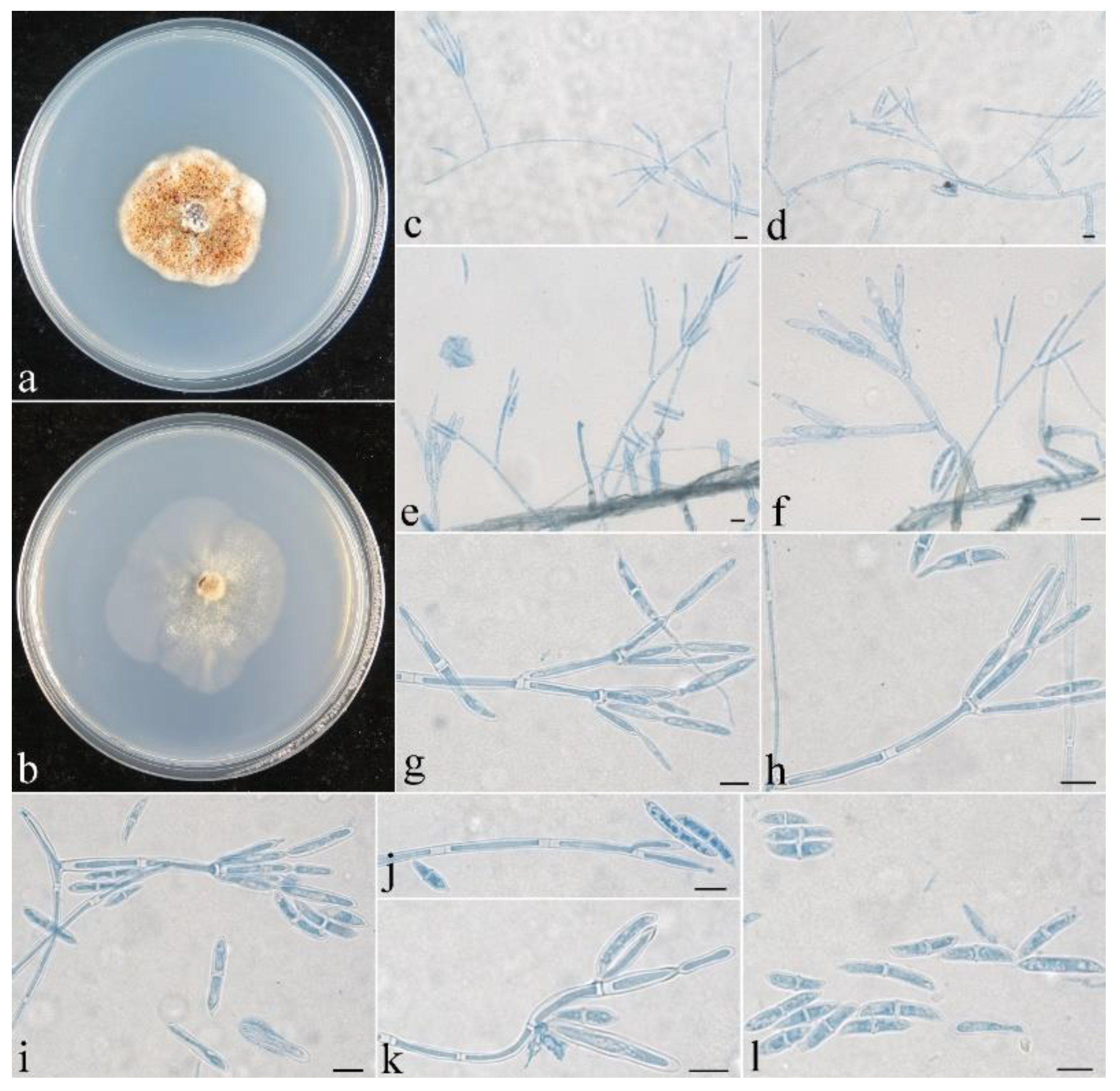
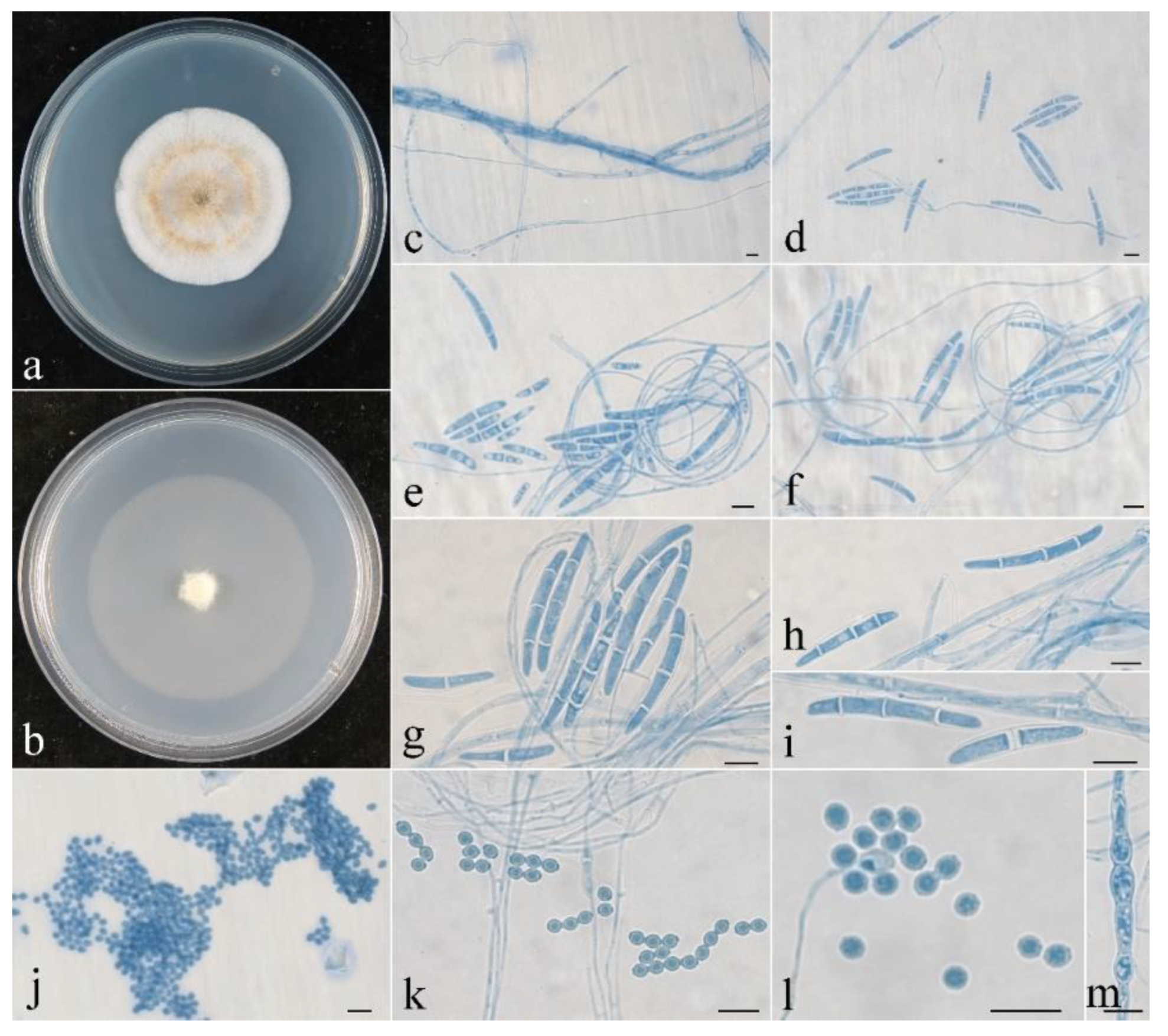
Figure 5.
Colonial and microscopic morphology of Penicillifer sinicus (CGMCC 3.24130). (a) Colony after 1 week at 25 °C on PDA; (b) colony after 1 week at 25 °C on SNA; (c–k) conidiophores, phialides, and macroconidia in lactophenol cotton blue; (l) macroconidia in lactophenol cotton blue. Bars: (c–l) = 10 μm.
Figure 5.
Colonial and microscopic morphology of Penicillifer sinicus (CGMCC 3.24130). (a) Colony after 1 week at 25 °C on PDA; (b) colony after 1 week at 25 °C on SNA; (c–k) conidiophores, phialides, and macroconidia in lactophenol cotton blue; (l) macroconidia in lactophenol cotton blue. Bars: (c–l) = 10 μm.

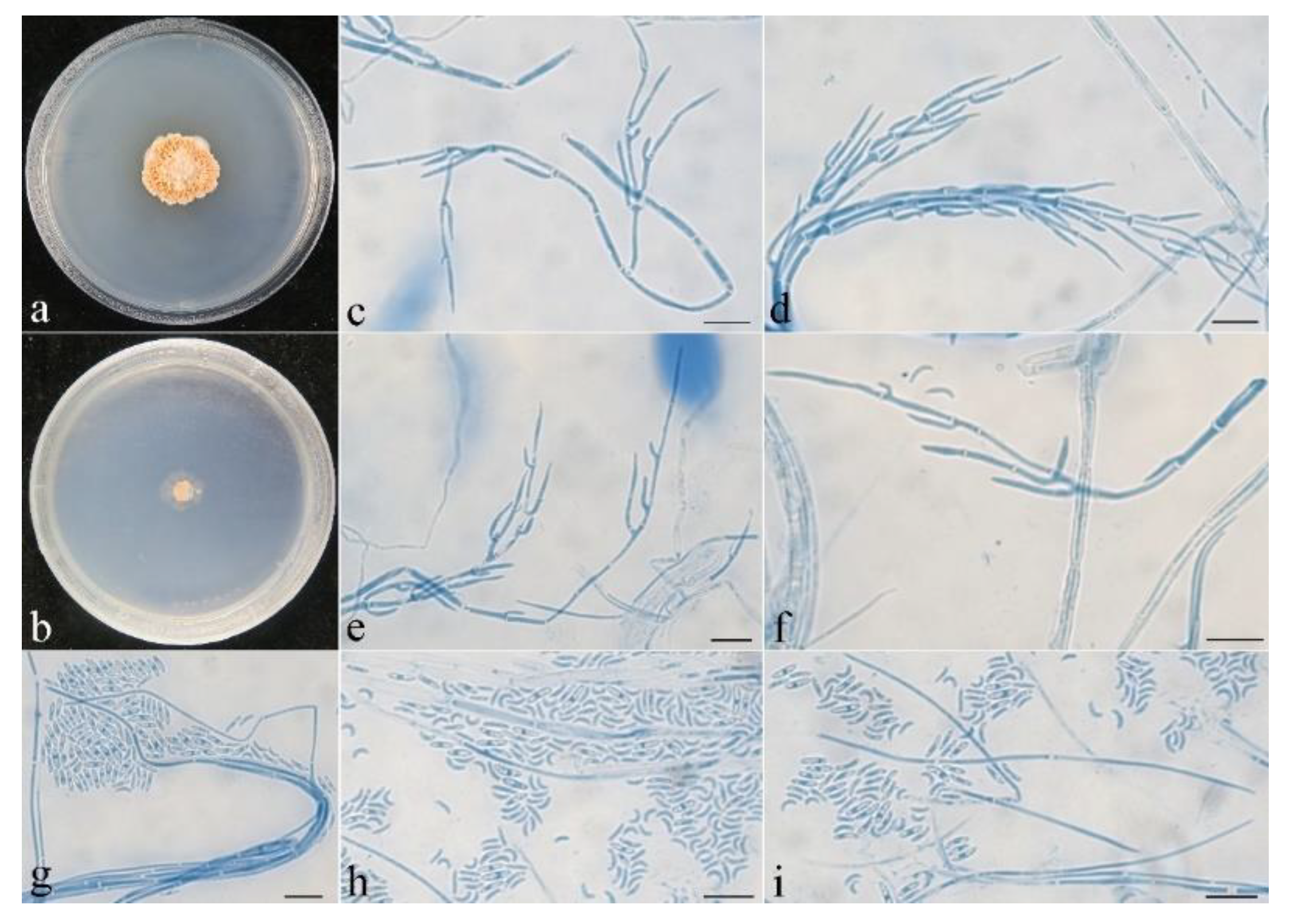
Figure 6.
Macroscopic and microscopic morphology of Pseudocosmospora beijingensis (HMAS 290896). (a–d) Ascomata on natural substratum; (e,f) median section of perithecia in lactic acid; (g,h) asci with ascospores in lactophenol cotton blue; (i–k) ascospore in lactophenol cotton blue. Bars: (a–d) = 1 mm; (e,f) = 50 μm; (g–k) = 10 μm.
Figure 6.
Macroscopic and microscopic morphology of Pseudocosmospora beijingensis (HMAS 290896). (a–d) Ascomata on natural substratum; (e,f) median section of perithecia in lactic acid; (g,h) asci with ascospores in lactophenol cotton blue; (i–k) ascospore in lactophenol cotton blue. Bars: (a–d) = 1 mm; (e,f) = 50 μm; (g–k) = 10 μm.

Figure 7.
Colonial and microscopic morphology of Pseudocosmospora beijingensis (CGMCC 3.24131). (a) Colony after 1 week at 25 °C on PDA; (b) colony after 1 week at 25 °C on SNA; (c–i) conidiophores, phialides, and microconidia in lactophenol cotton blue. Bars: (c–i) = 10 μm.
Figure 7.
Colonial and microscopic morphology of Pseudocosmospora beijingensis (CGMCC 3.24131). (a) Colony after 1 week at 25 °C on PDA; (b) colony after 1 week at 25 °C on SNA; (c–i) conidiophores, phialides, and microconidia in lactophenol cotton blue. Bars: (c–i) = 10 μm.

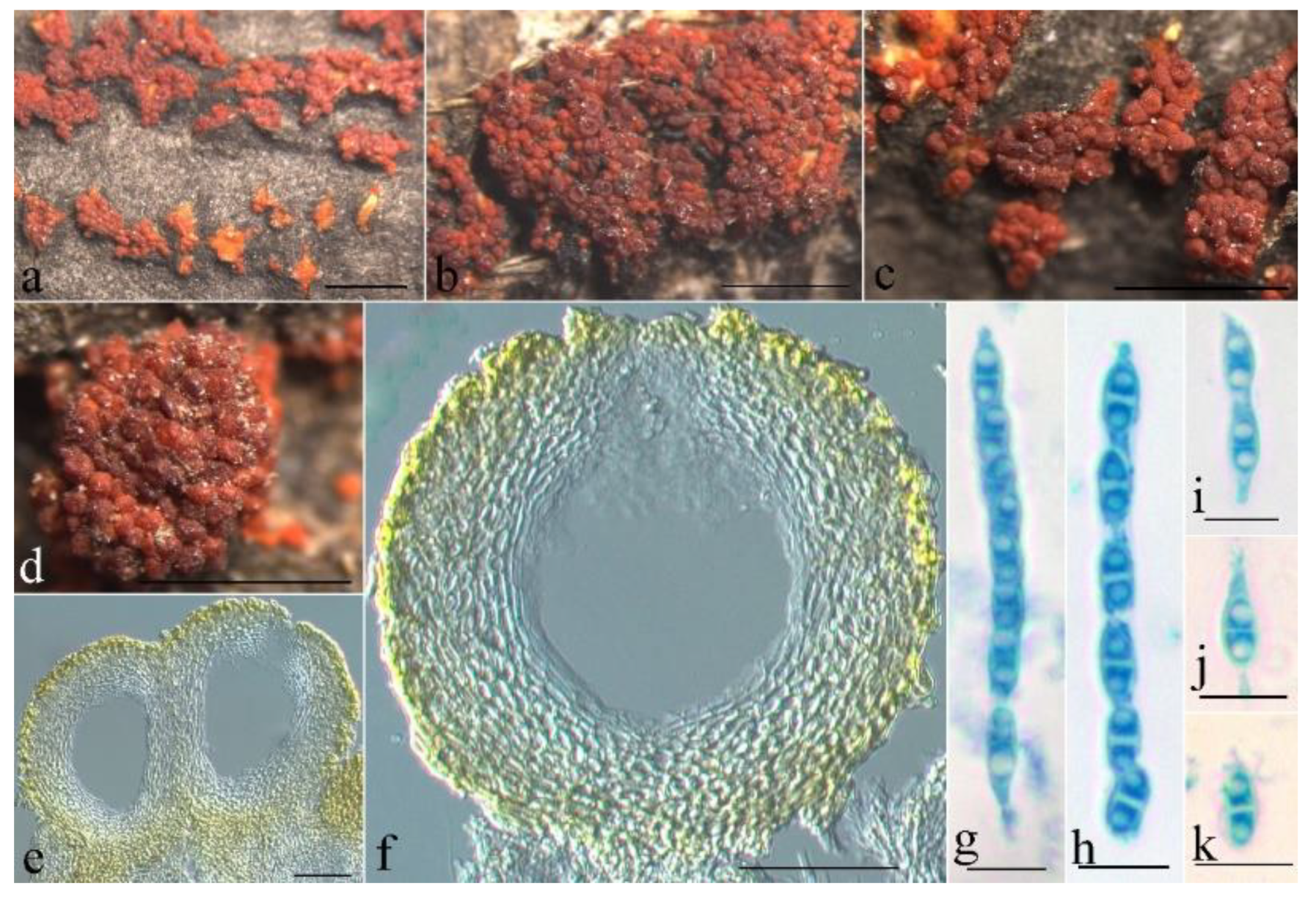
Figure 8.
Macroscopic and microscopic morphology of Thelonectria globulosa (HMAS 255835). (a–d) Ascomata on natural substratum; (e,f) median section of perithecium in lactophenol cotton blue; (g,h) asci with ascospores in lactophenol cotton blue; (i–k) ascospore in lactophenol cotton blue. Bars: (a–d) = 1 mm; (e,f) = 50 μm; (g–k) = 10 μm.
Figure 8.
Macroscopic and microscopic morphology of Thelonectria globulosa (HMAS 255835). (a–d) Ascomata on natural substratum; (e,f) median section of perithecium in lactophenol cotton blue; (g,h) asci with ascospores in lactophenol cotton blue; (i–k) ascospore in lactophenol cotton blue. Bars: (a–d) = 1 mm; (e,f) = 50 μm; (g–k) = 10 μm.

Figure 9.
Colonial and microscopic morphology of Thelonectria globulosa (CGMCC 3.24132). (a) Colony after 2 week at 25 °C on PDA; (b) colony after 2 week at 25 °C on SNA; (c) conidiophores in lactophenol cotton blue; (d–i) conidiophores and macroconidia in lactophenol cotton blue; (j–l) microconidia in lactophenol cotton blue; (m) chlamydospores in lactophenol cotton blue. Bars: (c–m) = 10 μm.
Figure 9.
Colonial and microscopic morphology of Thelonectria globulosa (CGMCC 3.24132). (a) Colony after 2 week at 25 °C on PDA; (b) colony after 2 week at 25 °C on SNA; (c) conidiophores in lactophenol cotton blue; (d–i) conidiophores and macroconidia in lactophenol cotton blue; (j–l) microconidia in lactophenol cotton blue; (m) chlamydospores in lactophenol cotton blue. Bars: (c–m) = 10 μm.

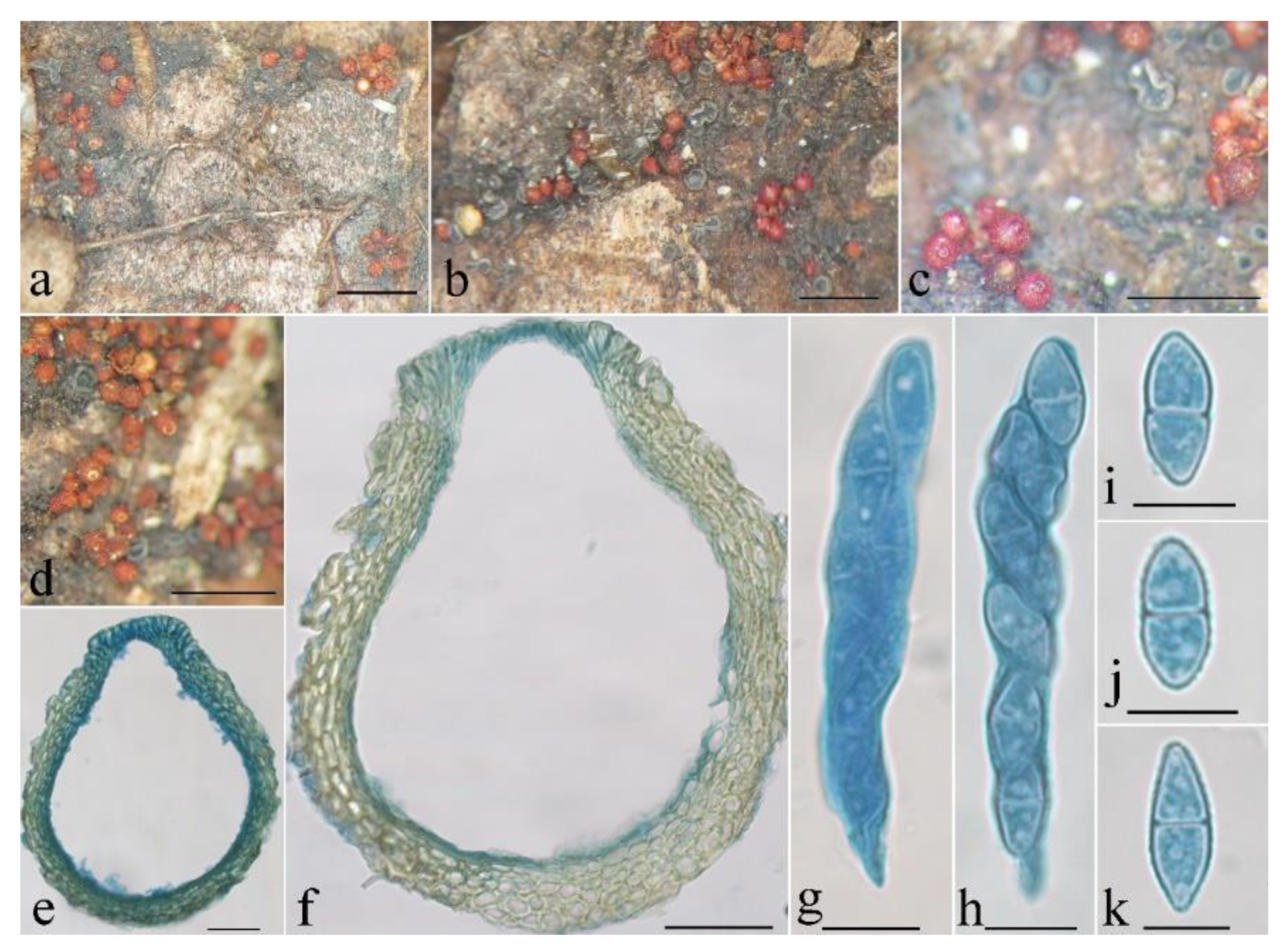
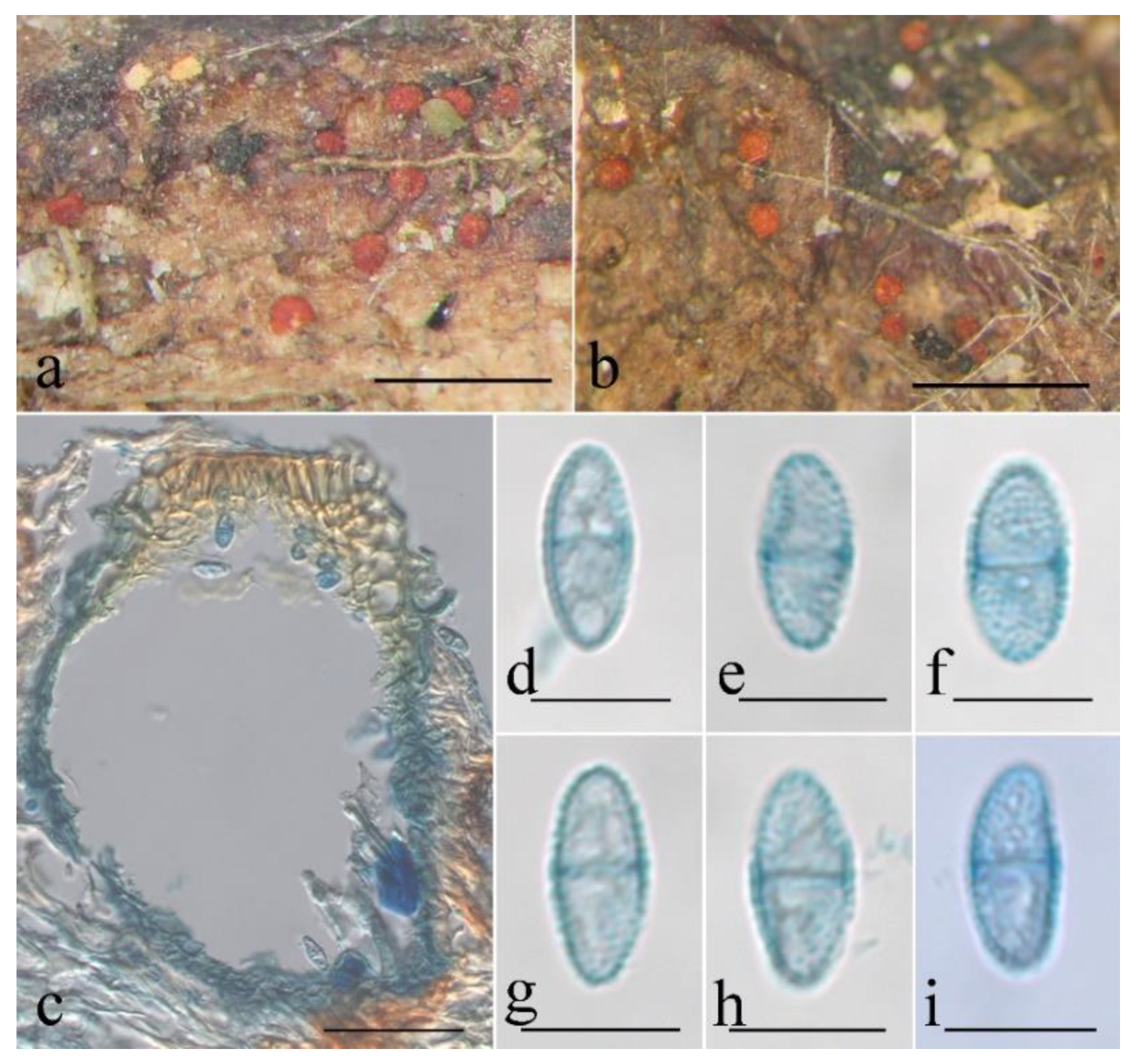
Figure 10.
Macroscopic and microscopic morphology of Thelonectria spinulospora (HMAS 290897). (a,b) Ascomata on natural substratum; (c) median section of perithecium in lactophenol cotton blue; (d–i) ascospore in lactophenol cotton blue. Bars: (a,b) = 1 mm; (c) = 50 μm; (d–i) = 10 μm.
Figure 10.
Macroscopic and microscopic morphology of Thelonectria spinulospora (HMAS 290897). (a,b) Ascomata on natural substratum; (c) median section of perithecium in lactophenol cotton blue; (d–i) ascospore in lactophenol cotton blue. Bars: (a,b) = 1 mm; (c) = 50 μm; (d–i) = 10 μm.

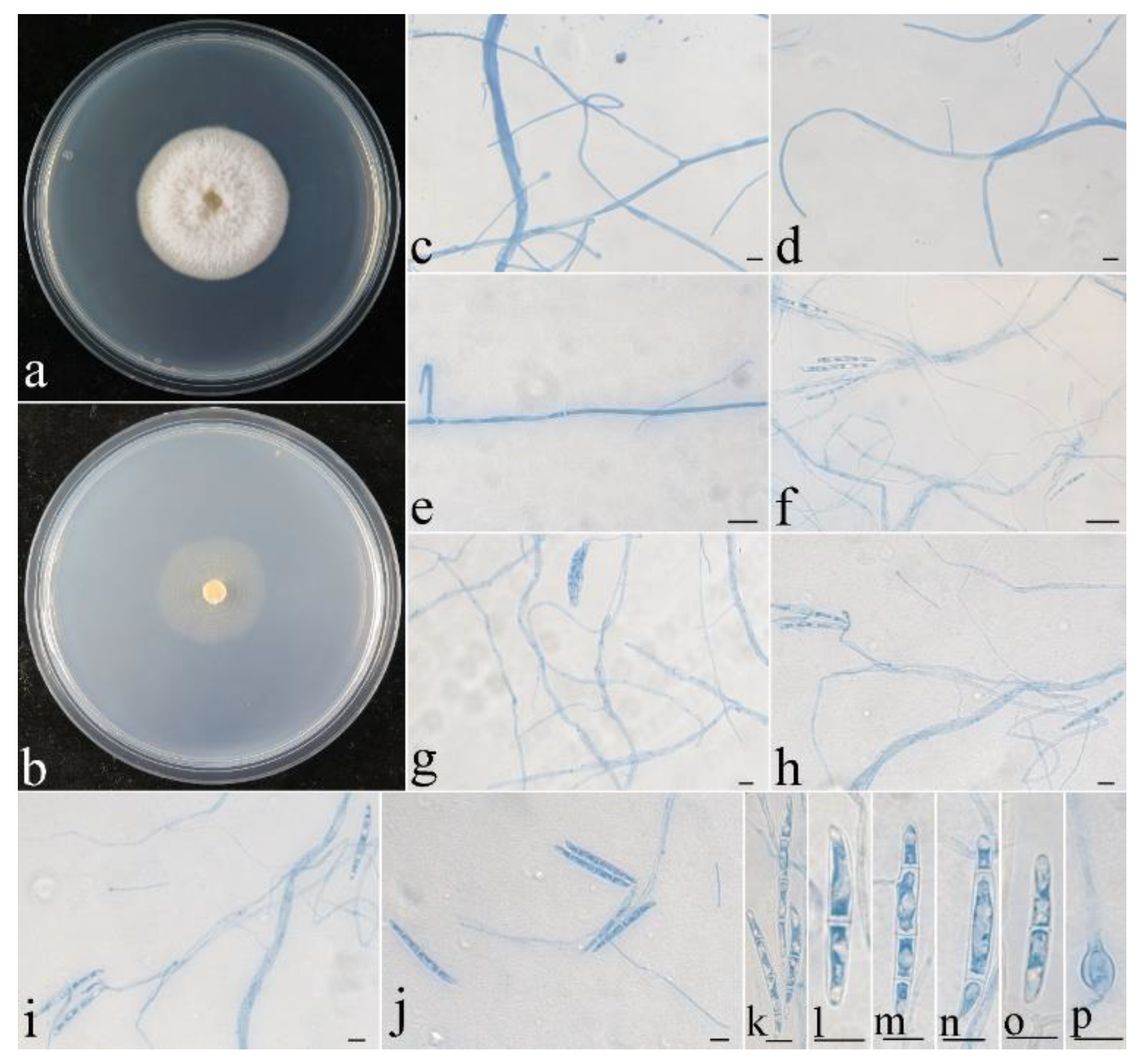
Figure 11.
Colonial and microscopic morphology of Thelonectria spinulospora (CGMCC 3.24133). (a) Colony after 2 week at 25 °C on PDA; (b) colony after 2 week at 25 °C on SNA; (c–e) conidiophores in lactophenol cotton blue; (f–j) conidiophores and macroconidia in lactophenol cotton blue; (k–o) macroconidia in lactophenol cotton blue; (p) chlamydospore in lactophenol cotton blue. Bars: (c–p) = 10 μm.
Figure 11.
Colonial and microscopic morphology of Thelonectria spinulospora (CGMCC 3.24133). (a) Colony after 2 week at 25 °C on PDA; (b) colony after 2 week at 25 °C on SNA; (c–e) conidiophores in lactophenol cotton blue; (f–j) conidiophores and macroconidia in lactophenol cotton blue; (k–o) macroconidia in lactophenol cotton blue; (p) chlamydospore in lactophenol cotton blue. Bars: (c–p) = 10 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of Penicillifer species, herbarium/strain numbers, and GenBank accession numbers of materials used in this study.
Table 1.
List of Penicillifer species, herbarium/strain numbers, and GenBank accession numbers of materials used in this study.
| Species | Herbarium/Strain Numbers | GenBank Accession Numbers | |||
|---|---|---|---|---|---|
| ITS | LSU | rpb2 | tef1 | ||
| Corallonectria jatrophae | CBS 91396 T | NR153873 | KM231611 | KM232298 | KM231863 |
| Dematiocladium celtidis | CBS 115994 T | AY793430 | AY793438 | - | KM231864 |
| P. bipapillatus | CBS 42088 T | KM231740 | KM231608 | KM232295 | KM231860 |
| P. diparietisporus | CBS 37659 T | NR154310 | MH869437 | KM232296 | KM231861 |
| P. macrosporus | CBS 42388 T | MH862133 | KM231607 | KM232294 | KM231859 |
| P. martinii | BRIP 59225 T | NR168155 | NG068753 | - | KJ869241 |
| P. pulcher | CBS 56067 T | NR154311 | NG058093 | KM232297 | KM231862 |
| P. sinicus | CGMCC 3.24130 T | OP223439 a | OP223435 | OP272863 | OP272864 |
| Stachybotrys chartarum | CBS 12913 | MH854622 | MH866145 | KM232434 | KM231994 |
T indicates the ex-type culture. a Numbers in bold indicate the newly provided sequences.
Table 2.
List of Pseudocosmospora species, herbarium/strain numbers, and GenBank accession numbers of materials used in this study.
Table 2.
List of Pseudocosmospora species, herbarium/strain numbers, and GenBank accession numbers of materials used in this study.
| Species | Herbarium/Strain Number | GenBank Accession Numbers | ||
|---|---|---|---|---|
| ITS | LSU | tub2 | ||
| Corallomycetella repens | AR 4547 | JF832594 | JF832679 | JF832838 |
| Microcera larvarum | AR 4580 | KC291751 | KC291759 | KC291935 |
| P. beijingensis | CGMCC 3.24131 T | OP223438 a | OP223434 | OP272862 |
| P. curvispora | CGMCC 3.20176 T | MT592897 | MT592879 | MT606156 |
| P. eutypae | CH 1101 T | KC291735 | KC291766 | KC291925 |
| IMI 73016 | KC291736 | KC291786 | - | |
| AR 4527 | KC291720 | KC291756 | KC291909 | |
| P. eutypellae | AR 4562 T | KC291721 | KC291757 | KC291912 |
| GJS 10248 | KC291722 | KC291772 | KC291911 | |
| P. henanensis | HMAS 183528 T | GU075856 | GU075863 | HM054103 |
| P. hypoxylicola | cLL 19020 T | MN886606 | MN886608 | - |
| P. joca | AR 4779 T | KC291746 | KC291762 | KC291924 |
| P. metajoca | AR 4576 T | KC291745 | KC291758 | KC291923 |
| P. rogersonii | GJS 9056 T | KC291729 | KC291780 | KC291915 |
| GJS 10296 | KC291727 | KC291774 | KC291917 | |
| GJS 091384 | KC291726 | KC291770 | KC291914 | |
| P. shennongjiana | CGMCC 3.20177 T | MT592898 | MT592880 | MT606157 |
| P. vilior | AR 4810 T | KC291737 | KC291763 | KC291928 |
| AR 4771 | KC291734 | KC291761 | KC291926 | |
| PC 1246 | KC291738 | KC291791 | KC291927 | |
T indicates the ex-type culture. a Numbers in bold indicate the newly provided sequences.
Table 3.
List of Thelonectria species, herbarium/strain numbers, and GenBank accession numbers of materials used in this study.
Table 3.
List of Thelonectria species, herbarium/strain numbers, and GenBank accession numbers of materials used in this study.
| Species | Herbarium/Strain Numbers | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| act | ITS | LSU | rpb1 | tub2 | ||
| Cosmospora coccinea | CBS 114050 | GQ505967 | FJ474072 | GQ505990 | GQ506020 | DQ522501 |
| Nectria cinnabarina | AR 4477/AR 4302 | HM484627 | HM484548 | HM484562 | M484577 | HM484820 |
| T. acrotyla | GJS 90171 | JQ365047 | JQ403329 | JQ403368 | JQ403407 | JQ394720 |
| T. amamiensis | MAFF 239819 | JQ365054 | JQ403337 | JQ403375 | KJ022408 | JQ394727 |
| T. chlamydospora | ST 121 T | LC519560 | LC509450 | LC509452 | - | - |
| T. globulosa | CGMCC 3.24132 T | OP272865 a | OP223436 | OP223432 | OP272867 | OP586762 |
| T. gongylodes | GJS 04171 T | JQ365038 | JQ4033 | JQ403358 | JQ403395 | JQ394710 |
| T. nodosa | GJS 04155 T | JQ365037 | JQ403317 | JQ403357 | JQ403394 | - |
| T. olida | CBS 21567 T | HM352884 | KJ021982 | KJ022058 | HM364334 | KM232024 |
| T. rubrococca | IMI 324475 T | KJ022275 | KJ022008 | KJ022061 | KJ022439 | KJ022329 |
| T. spinulospora | CGMCC 3.24133 T | OP272866 | OP223437 | OP223433 | OP272868 | OP586764 |
| T. torulosa | AR 4768A | JQ365031 | JQ403310 | JQ403350 | JQ403386 | JQ394702 |
| T. trachosa | GJS 9245 T | KF569832 | KF569842 | KF569851 | KF569879 | KF569869 |
| T. truncata | GJS 04357 T | JQ365039 | JQ403319 | JQ403359 | JQ403396 | KJ022324 |
| T. veuillotiana | AR 1751 | KJ022273 | JQ403305 | JQ403345 | JQ403382 | JQ394698 |
T indicates the ex-type culture. a Numbers in bold indicate the newly provided sequences.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zeng, Z.-Q.; Zhuang, W.-Y. New Species of Nectriaceae (Hypocreales) from China. J. Fungi 2022, 8, 1075. https://doi.org/10.3390/jof8101075
AMA Style
Zeng Z-Q, Zhuang W-Y. New Species of Nectriaceae (Hypocreales) from China. Journal of Fungi. 2022; 8(10):1075. https://doi.org/10.3390/jof8101075
Chicago/Turabian StyleZeng, Zhao-Qing, and Wen-Ying Zhuang. 2022. "New Species of Nectriaceae (Hypocreales) from China" Journal of Fungi 8, no. 10: 1075. https://doi.org/10.3390/jof8101075
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.