
Fourteen New Species of Foliar Colletotrichum Associated with the Invasive Plant Ageratinaadenophora and Surrounding Crops

1
Laboratory for Conservation and Utilization of Bio-Resources, Yunnan University, Kunming 650091, China
2
School of Life Sciences, Yunnan University, Kunming 650091, China
*
Authors to whom correspondence should be addressed.
J. Fungi 2022, 8(2), 185; https://doi.org/10.3390/jof8020185
Submission received: 20 January 2022
/
Revised: 11 February 2022
/
Accepted: 11 February 2022
/
Published: 13 February 2022
(This article belongs to the Topic Fungal Diversity)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Ageratina adenophora is one of the most invasive weeds in China. Following an outbreak in Yunnan in the 1960s, A. adenophora has been spreading in Southwest China at tremendous speed. Previous research indicated A. adenophora contained many Colletotrichum species as endophytes. In this study, we investigated the diversity of Colletotrichum in healthy and diseased leaves of the invasive plant A. adenophora and several surrounding crops in Yunnan, Guangxi, and Guizhou provinces in China, and obtained over 1000 Colletotrichum strains. After preliminary delimitation using the internal transcribed spacer region (ITS) sequences, 44 representative strains were selected for further study. Their phylogenetic positions were determined by phylogenetic analyses using combined sequences of ITS, actin (ACT), chitin synthase (CHS-1), glyceraldehyde-3-phosphate dehydrogenase (GAPDH), and beta-tubulin (TUB2). Combined with morphological characteristics, 14 new Colletotrichum species were named as C. adenophorae, C. analogum, C. cangyuanense, C. dimorphum, C. gracile, C. nanhuaense, C. nullisetosum, C. oblongisporum, C. parvisporum, C. robustum, C. simulanticitri, C. speciosum, C. subhenanense, and C. yunajiangense.
1. Introduction
Foliar fungal endophytes are defined as fungi that can colonize the internal plant leaf structure without causing apparent harm to the aerial parts of the host [1]. Foliar tissues of all plant species examined to date harbor at least one and, more commonly, a wide diversity of fungi lurking within their plant tissues [2]. These fungi phylogenetically span at least four phyla [3] and are categorized into two groups based on their manner of transmission: vertically transmitted endophytes (VTEs, e.g., Neotyphodium Glenn et al.) that inhabit temperate grasses (Lolium L.) and are transmitted from the mother plant [4] and horizontally transmitted endophytes (HTEs) transmitted mostly via spores from plant to plant via rain, wind, local leaf litter, or insect vectors [5,6]. HTEs are so common that the foliar tissues of all plant species examined thus far harbor at least one fungus [2]. Although the majority of these endophytic fungi are localized infections by either latent pathogens or dormant saprophytes [2], in many cases endophytes may promote host plant growth, improve nutrient supply and protect plants from both biotic and abiotic stresses [7,8]. Because a plant can be colonized by endophytic fungi that originate from a neighbor or by spores that persist in the local environment, it is not surprising that endophyte communities found in the same plant host species can show striking geographical differences [6].
Non-native (alien, exotic, introduced) species are species whose presence in a region is attributable to human actions that enabled them to overcome fundamentally biogeographic barriers. Only a fraction of non-native species become invasive; they are defined as species that become naturalized, that is, sustain self-replacing populations over several life cycles and have the potential to spread over long distances and dominate local communities [9]. When invasive plants are introduced into a new area, they will likely develop a novel relationship with the local microorganisms and generate impacts on ecosystems [10,11]. In many cases, these novel relationships rather than the plant itself determine the invasiveness of an exotic plant in an introduced range [12]. Many works have investigated the belowground microbes (soils) associated with invasive plants and their roles [13,14,15]. Invasive plants encounter foliar fungal endophytes; however, this community of endophytes is investigated less often. Until now, only a few reports have evaluated these fungi. Genetic analysis of endophytic fungi found in Centaurea stoebe L. in both its native and invaded ranges showed that 85% occurred in only one of the two ranges [16]. In Europe, the native range, the most common endophyte was a haplotype of Alternaria alternata (Fr.) Keissl. In the invaded range, no haplotype was dominant, and many were novel. Some fungal endophytes appear to have been introduced with C. stoebe, but the invader also acquired new endophytes after introduction [16]. The most common OTU corresponded most closely to Sarocladium strictum (W. Gams) Summerb. and comprised 25% of all fungal isolates. The functional role of the isolated endophytes is not yet known, but one genus isolated here (Stagonospora (Sacc.) Sacc.) has been reported to enhance Phragmites growth. Some of the fungal endophytes isolated here may correspond to latent pathogens (e.g., OTU 2) that either have not expressed their pathogenic phase [17] or are not pathogenic to non-native Phragmites [18,19]. Non-native P. australis (Cav.) Trin. ex Steud. decreases biodiversity and produces dense stands in North America. We surveyed the endophyte communities in the stems, leaves, and roots of collections of P. australis obtained from two sites with a low and high salt concentration to determine differences in endophyte composition and assess differences in the functional roles of microbes in plants from both sites. The high salt endophyte community showed higher resistance to zinc, mercury, and salt stress compared to fungal species from the low salt site. These endophytes also showed a greater propensity for growth promotion of rice seedlings (a model species) under salt stress. The results of this study were consistent with the ‘habitat-adapted symbiosis hypothesis’ that holds that endophytic microbes may help plants adapt to extreme habitats. The capacity of P. australis to establish symbiotic relationships with diverse endophytic microbes that enhance its tolerance to abiotic stresses could be a factor contributing to its invasiveness in saline environments [20].
Generally, endophytes are beneficial to the invasion of the host (there are also contrary examples, e.g., [21,22]). For example, the invasive forb species C. stoebe has allelopathic effects on native species that are significantly enhanced when it is infected by a fungal endophyte closely related to A. alternata [23]. The competitive effects of C. stoebe on grass species native to North America were enhanced by both Alternaria endophytes, but not on native European grasses [24]. Similarly, some fungal endophytes isolated from C. stoebe seeds caused significant declines in germination of Festuca idahoensis Elmer, a North American grass competitor [25]. Recently, fungi isolated from an invasive plant Vincetoxicum rossicum (Kleopow) Barbar. were proven to have differential growth effects between non-native invasive and native plants, which may be important for mediating plant community structure and the persistence of invasive plants [26].
Ageratina adenophora (Sprengel) King and Robinson (syn. Eupatorium adenophorum Spreng.) (Asteraceae), also known as Crofton weed or Mexican Devil weed, originates in Mexico and is an invasive weed in several countries worldwide, including Australia, China, Hawaii, India, New Zealand and South Africa [27,28]. It is one of the most invasive weeds in China. Following an outbreak in Yunnan in the 1960s, A. adenophora has been spreading north- and eastward through Guizhou, Sichuan, and Guangxi Provinces in Southwest China at approximately 20 km/year [29]. To determine if A. adenophora could benefit from the fungal endophytes accumulated in the introduced range, our team previously reported that A. adenophora actually contained diverse fungal endophytes; in particular, Colletotrichum Corda comprised 30% of total isolated strains [30]. However, the foliar Colletotrichum generally adversely affect leaf development in A. adenophora and may be a weak latent foliar pathogen [31]. Our further disease experiments verified that some of these Colletotrichum are pathogenic on local native plants [32]. Indeed, Colletotrichum species are among the most commonly occurring pathogens and foliar endophytes of terrestrial plants [33,34]. The Colletotrichum genus was recently included in the list of the 10 most important plant pathogenic fungi, based on perceived scientific and economic importance [35]. The genus Colletotrichum comprises ~600 species attacking over 3200 species of monocot and dicot plants [36]. In addition, many Colletotrichum species are latent plant pathogens, these species being essentially endophytes, epiphytes, or saprobes, switching to a pathogenic lifestyle when host plants are stressed or in postharvest storage [37,38]. It is common to find multiple Colletotrichum species infecting a single host [39]. Although it has also been shown that particular Colletotrichum endophytes confer protective benefits on cacao hosts by reducing disease incidence and damage caused by other plant pathogens [40,41,42], in this case, there is a possible ecological risk of disease transmission to local plants driven by A. adenophora invasion.
Our current understanding of Colletotrichum species is largely restricted to populations affecting crop or ornamental plants, with poor knowledge of non-agricultural crops [34]. Higgins et al. [43] studied Colletotrichum endophytes from grass and non-grass hosts in a tropical forest in Panama, recovering some genetically distinct taxa via direct sequence from surface-sterilized grass tissue that were not detected using cultural methods. They also observed that many taxa were detected from more than one grass host genus, corroborating observations by Lu et al. [44] and Arnold and Lutzoni [3] that the commonest tropical endophytes appear to be host generalists. All the studies of Colletotrichum associated with non-crop plants detailed above demonstrated considerable diversity of taxa. Despite preliminary evidence of less host specificity in native tropical forest ecosystems compared with managed environments, the sheer number of habitats (in the form of leaves, fruits, etc.) that remain unsampled indicates the likelihood that overall species-level diversity of the genus is still significantly under-represented. Studies of Colletotrichum from wild plants are likely to be particularly fruitful. It would be presumptuous even to speculate that the overall systematic framework for the genus cannot be improved [34]. Several endophyte taxa isolated from cacao in Panama by Rojas et al. [45] were thought to comprise part of the background endophytic community in the Panamanian forest ecosystem [45].
Previously, the identification of Colletotrichum species was based on morphological characteristics. Cai et al. [46] found that such morphological identification of Colletotrichum species depended on the experimental methods used, which caused the taxonomy and nomenclature to be inconsistent. Recently, Cai et al. recommended a polyphasic approach for accurate identification of Colletotrichum species using multi-locus phylogeny coupled with morphological data. Using this approach, many Colletotrichum strains have been successfully identified and epitypified. This increased understanding of Colletotrichum species can increase the effectiveness of plant disease control interventions [47].
In the present study, we investigated the diversity of Colletotrichum in healthy and diseased leaves of invasive plant A. adenophora and several surrounding crops in Yunnan, Guangxi, and Guizhou Provinces in China, and obtained over 1000 Colletotrichum strains. Of these, 44 representative strains were selected for further study, including five-locus phylogenetic analyses and morphological characteristics. Finally, these strains were assigned to 26 Colletotrichum species, including 12 known Colletotrichum species and 14 new species. The results of this study will contribute to the knowledge on the diversity and phylogeny of Colletotrichum associated with an invasive plant and surrounding crops. Because the Colletotrichum genus is one of the most important pathogenic fungi [35], our finding is also valuable for understanding the possible disease risk driven by invasive plants in the invaded ecosystem.
2. Materials and Methods
2.1. Sample Collection and Isolation of Fungi
The Colletotrichum strains used in this study were collected from healthy and diseased leaves of the invasive plant Ageratina adenophora, as well as diseased leaves of native plants, from 2012 to 2018, using leaf fragment incubation [32,48]. Briefly, for the isolation of leaf spot fungi, healthy leaf tissues and the margins of diseased tissues of each leaf spot were cut into 6 mm2 sections and surface sterilized as follows. These sections underwent initial immersion for 2 min in 0.5% sodium hypochlorite, followed by 1 min in sterile distilled water, 2 min in 75% ethanol, and, finally, 1 min in sterile distilled water. The disinfected fragments were then plated onto potato dextrose agar (PDA; 200 g potato, 20 g glucose, 18 g agar, 1 L distilled water) and incubated at ambient temperature for 6–8 days or until mycelia growing from the leaf fragments were observed. In total, over 1000 Colletotrichum strains were collected and we selected 44 representative strains for further study.
The pure cultures and dried cultures were deposited in the Herbarium of the Laboratory for Conservation and Utilization of Bio-resources, Yunnan University, Kunming, Yunnan, China (YMF) and the China General Microbiological Culture Collection Center (CGMCC).
2.2. Morphological Characterization
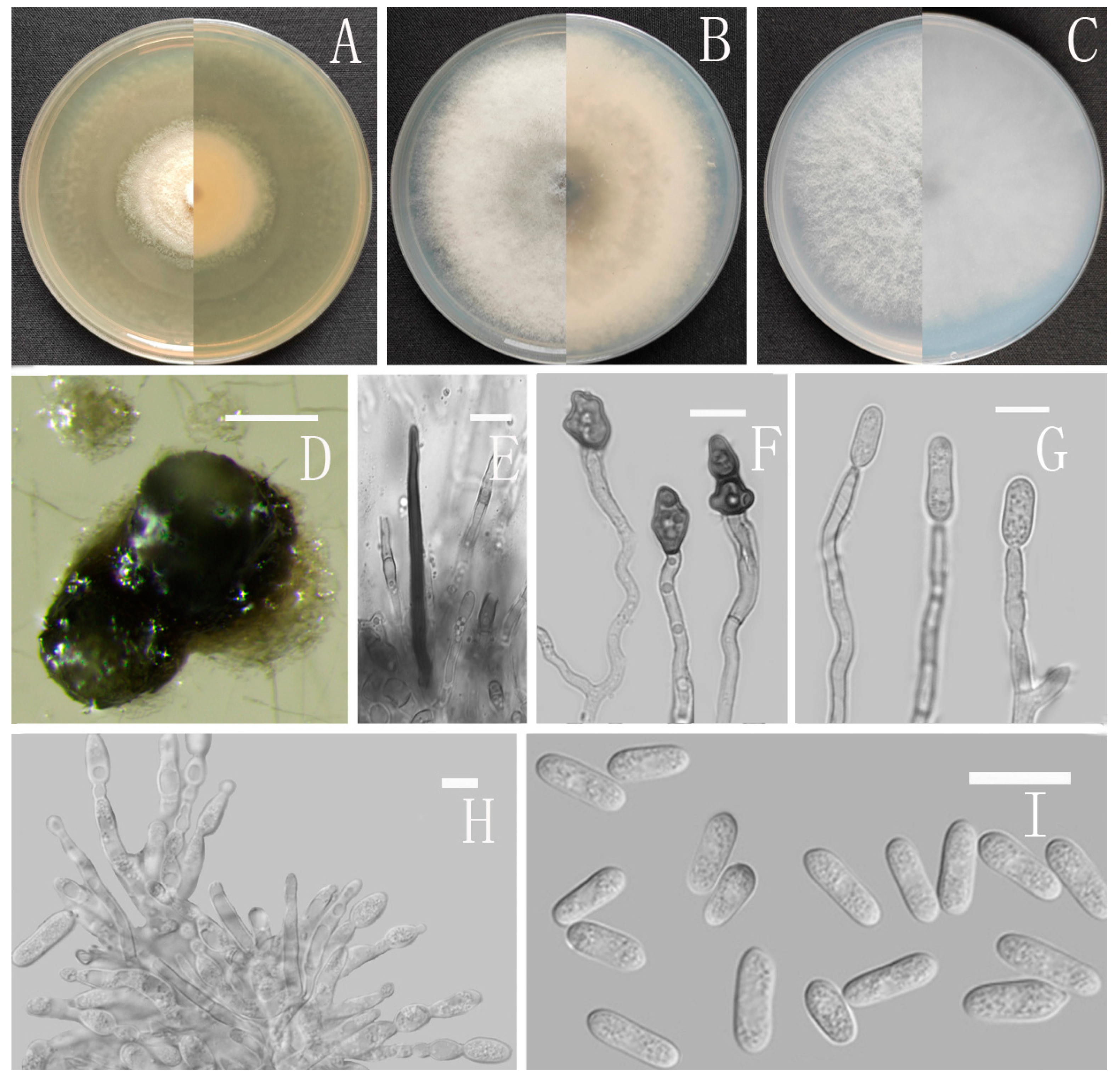
The growth rates of Colletotrichum isolates were determined on four common media: cornmeal dextrose agar (CMA; 20 g cornmeal, 18 g agar, 1 L distilled water), low nutrient agar (SNA; 1 g KH2PO4, 1 g KNO3, 0.5 g KCl, 0.5 g MgSO4, 0.2 g sucrose, 0.2 g glucose, 18 g agar, 1 L distilled water), malt extract agar (MEA; 30 g malt powder, 3 g peptone, 18 g agar, 1 L distilled water), and PDA, respectively. Six-millimeter plugs of actively growing mycelia were inoculated onto these media (20 mL per plate); these cultures were incubated at 25 °C for 7 days. During this time, the colony diameters and characteristics, such as colony appearance, color, and sporogony, on the four media were recorded. Moreover, microscopic characteristics were identified by photographing with a BX51 microscope connected to a DP Controller digital camera. Microscopic characteristics were created from cultures growing on CMA or SNA at 25 °C.
2.3. DNA Extraction, PCR Amplification, and Sequencing
Genomic DNA was extracted from the fresh mycelium harvested from PDA plates after 4 days, as described by Wang and Zhuang [49]. Five loci, the nuc rDNA internal transcribed spacers (ITS), actin (ACT), chitin synthase (CHS-1), glyceraldehyde-3-phosphate dehydrogenase (GAPDH), and beta-tubulin (TUB2), were amplified with the following primer pairs, ITS4 and ITS5 for ITS [50],GDF1 and GDR1 for GAPDH [51], ACT-512F and ACT-783R for ACT [52], CHS-79F and CHS-354R for CHS-1 [52], and Bt2a and Bt2b for TUB2 [53]. The ITS sequence fragment was used for original identification; then the ITS together with GAPDH, CHS-1, ACT and TUB2 were used to construct a phylogenetic tree of the genus Colletotrichum. A PCR reaction was conducted in a 25 μL reaction volume, and the components were as follows: 1 μL DNA template, 1 μL forward 10 μM primer, 1 μL reverse 10 μM primer, 12.5 μL T5 Super PCR Mix (containing Taq polymerase, dNTP and Mg2+, Beijing TisingKe Biotech Co., Ltd., Beijing, China), 5 μL enhance buffer and 4.5 μL sterile water. PCR reactions were run in an Eppendorf Mastercycler ep with related PCR thermal cycle programs described by Zhuang and Chen [54].
PCR products were firstly checked by performing 1.5% agarose gel electrophoresis stained with ethidium bromide, then purified using the PCR product purification kit (Biocolor BioScience and Technology Co., Shanghai, China), and finally sent for both forward and reverse sequencing on an ABI 3730 XL DNA sequencer (Applied Biosystems, Foster City, CA, USA) with the same primers at Tsingke Biotechnology Co., Ltd. (Kunming, China). Then, the forward and reverse sequences were assembled to obtain a consensus sequence using DNAMAN v. 9.0. These newly generated sequences were deposited in the GenBank database, and the accession numbers from this study in the National Center for Biotechnology Information (NCBI) are listed in Supplementary Table S1.
2.4. Multigene Phylogenetic Analyses
The phylogenetic tree was constructed with five loci ACT, CHS-1, GAPDH, TUB2, and ITS to identify these Colletotrichum isolates to the species level. Additional reference species sequences were selected based on recent studies on Colletotrichum species [55,56], referring to 14 species complexes and 13 singleton species. Both the reference sequences and new generated sequences in this study are listed in Supplementary Table S1. Multiple sequence alignments for the five loci were constructed and carried out using Clustalx v. 1.83 [57] with the default settings. The phylogeny was developed based on maximum-likelihood for all individual loci. Then, five datasets of ACT, CHS-1, GAPDH, TUB2, and ITS of Colletotrichum spp. were manually adjusted and linked through BioEdit v. 7.0 [58]. Of these, gap adjustments were performed and ambiguously aligned regions were also excluded. The final concatenated sequence matrix (Fasta file) generated by BioEdit v. 7.0 contained 2119 characters with gaps, ACT with 310 sites, CHS with 299 sites, GAPDH with 363 sites, TUB2 with 532 sites, and ITS with 615 sites. Gaps in the concatenated dataset were treated as missing data in maximum-likelihood (ML) trees and Bayesian inference (BI) methods.
ML analysis was performed using RAxML v. 8.2.10 [59] with the PHY files, which were generated using CLUSTAL_X v. 1.83 [57]. Final ML searches were conducted using the GTR-GAMMA model. Maximum likelihood bootstrap proportions (MLBP) were computed with 1000 replicates and were given above each node of the phylogenetic tree when it was equal to or greater than 70. For BI analysis, a Markov Chain Monte Carlo (MCMC) algorithm was used to generate phylogenetic trees using MrBayes v. 3.1.2 [60]. The concatenated dataset was converted to a NEXUS file with MEGA6 [61], then the Akaike information criterion (AIC) was implemented by using jModelTest v. 2.0 [62] to select the best fit models of nucleotide substitution. GTR+I+G+F was selected as the best fit model for the concatenated dataset. Two independent analyses with four chains were run for 4,000,000 generations sampling every 1000th generation, halting the analyses when the average standard deviation of split frequencies was less than 0.01. The first 25% of the tree generations were discarded as the burn-in phase of the analyses, and the Bayesian inference posterior probability (BIPP) was obtained from the remaining trees. Trees generated in this study were viewed with TreeView v. 1.6.6 [63], and the final topology of phylogenetic trees was based on maximum likelihood analysis, with MLBP greater than 70 and BIPP greater than 0.9, as shown at the nodes.
3. Results
3.1. Multi-Locus Phylogeny
All Colletotrichum spp. isolates were first identified at genus level based on their ITS sequences. To further identify these isolates at species level, we conducted multi-locus phylogenetic analysis referring to 2119 characters and 220 sequences. For the five-locus (CHS-1, GAPDH, ITS, ACT, and TUB2) phylogenetic analysis of the genus Colletotrichum, 173 reference sequences obtained from GenBank and our selected 46 representative sequences of Colletotrichum isolates were used in this study. Monilochaetes infuscans Harter CBS 869.96 was selected as the outgroup according to recent publications [55,56]. When the five individual sequence datasets did not show any conflicts in the tree topologies based on preliminary ML analyses, the phylogeny was developed based on both ML and BI for the combined multi-locus analyses (Figure 1).
The results of this analysis revealed that 46 isolates were distributed in eight species complex clades, which were, respectively, the C. dracaenophilum species complex, the C. magnum species complex, the C. orchidearum species complex, the C. gigasporum species complex, the C. gloeosporioides species complex, the C. boninense species complex, the C. acutatum species complex, and the C. graminicola-caudatum species complex clades, and one isolate cluster with the singleton species C. sydowii CBS 135819.
For the isolates in the C. dracaenophilum species complex, one isolate clustered with C. excelsum-altitudinum and C. tongrenense (MLBP 81/ BIBP 0.98); we designated the isolate as C. robustum.
For the isolates in the C. magnum species complex, two isolates clustered with the C. magnum clade (MLBP 80) corresponding to the known species C. magnum. For the isolates in the C. orchidearum species complex, two isolates were grouped with the C. plurivorum clade (100/1) and identified as the known species C. plurivorum.
For the isolates in the C. gigasporum species complex, two isolates clustered with the C. gigasporum clade (100/1) corresponding to C. gigasporum.
For the isolates in the C. gloeosporioides species complex, 23 isolates clustered in 11 clades corresponding to C. fructicola (100/1), C. parvisporum (87/0.98), C. dimorphum (100/1), C. nanhuaense (100/1), C. yunajiangense (100/1), C. analogum (95/1), C. oblongisporum (95/1), C. nullisetosum (MLBP 84), C. gracile (100/1), C. cangyuanense (100/1), and C. subhenanense (100/1).
For the isolates in the C. boninense species complex, four isolates were distributed in two clades and identified as two known species C. boninense (100/1) and C. karstii (100/1). For the isolates in the C. acutatum species complex, ten isolates were distributed in eight clades corresponding to C. citri (99/0.96), C. nymphaeae (97/1), C. simulanticitri (98/0.99), C. cosmi (100/1), C. guajavae (95/0.98), C. speciosum (100/1), C. fioriniae (100/1), and C. godetiae (100/1).
In addition, one isolate clustered with the singleton species C. sydowii (100/1), designated as C. adenophorae.
3.2. Taxonomy
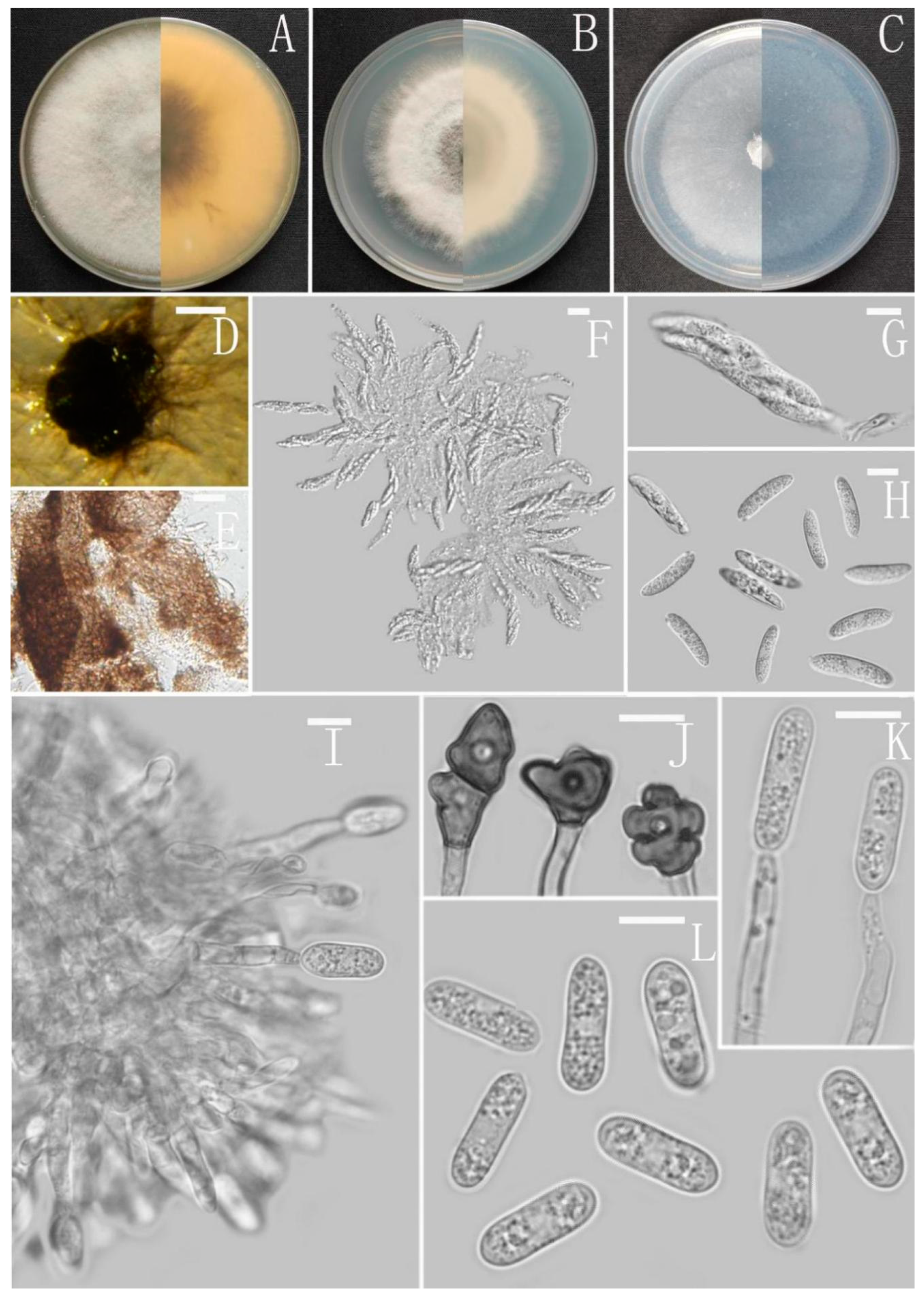
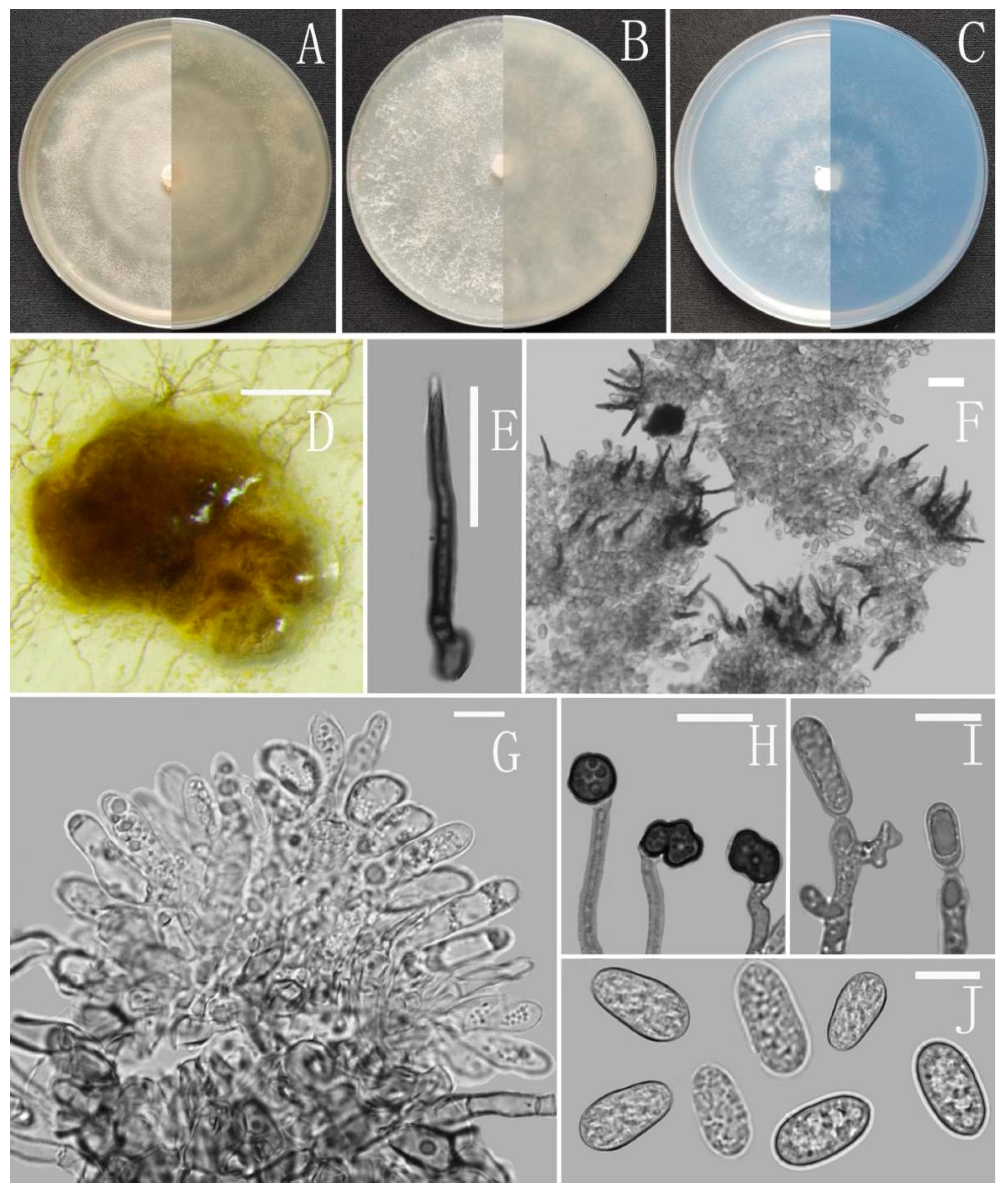
Colletotrichum adenophorae Z.F. Yu (Figure 2).
MycoBank Number: MB842546.
Etymology: The species epithet refers to the host plant Ageratina adenophora.
Type: China, Yunnan province, Lancang county, isolated as an endophyte from Ageratina adenophora, August 2016, Han-Bo Zhang. Holotype YMF 1.04952, Ex-type culture CGMCC 3.18952.
Sexual morph not observed. Asexual morph on CMA. Conidiomata rudimentary, dark olivaceous brown. Setae and conidiophores formed directly on hyphae. Setae abundant, cylindrical, straight or slightly flexuous, dark brown to black, 4–6-septate, base cylindrical to slightly inflated, tip acute to rounded, 54–105 μm long. Conidiophores hyaline to pale brown, smooth-walled, septate, unbranched. Conidiogenous cells hyaline, cylindrical, smooth-walled, collarette absent. Conidia straightly cylindrical, oblong to narrowly ovoid, obtuse at the base, rounded at the apex, contents granular, 18–22.5 × 5.5–7.5 μm, mean ± SD = 20.2 ± 1.1 × 6.2 ± 0.4 μm, l/w ratio = 3.3. Appressoria dark brown, irregular, margin sometimes lobed, 6.5–14 × 5.5–12 μm.
Culture characteristics: Colonies on MEA flat with entire margin, occupied the whole plate in 7 days at 25 °C, surface buff, partly covered with a short, felty, whitish aerial mycelium, reverse medium buff to pale honey. Colonies on PDA flat with entire margin, occupied the whole plate in 7 days at 25 °C, aerial mycelium fluffy, flocculent, white in edges, reverse white to light yellow. Colonies on SNA flat with entire margin, occupied the whole plate in 7 days at 25 °C, covered with thin, whitish aerial mycelium, producing pale gray conidiomata in 2 weeks, reverse same colors. Colonies on CMA reaching 59–60 mm diameter in 7 days at 25 °C, aerial mycelium fluffy, flocculent, white, reverse same colors.
Notes: Colletotrichum adenophorae is phylogenetically clustered with the singleton species C. sydowii Damm. However, C. adenophorae is easily distinguished from C. sydowii in morphology. For instance, C. sydowii has smaller conidia [(14–)15.5–18.5(–20.5) × 5–6 vs. 18.0–22.5 × 5.5–7.5 μm] and less lobate appressoria [64].
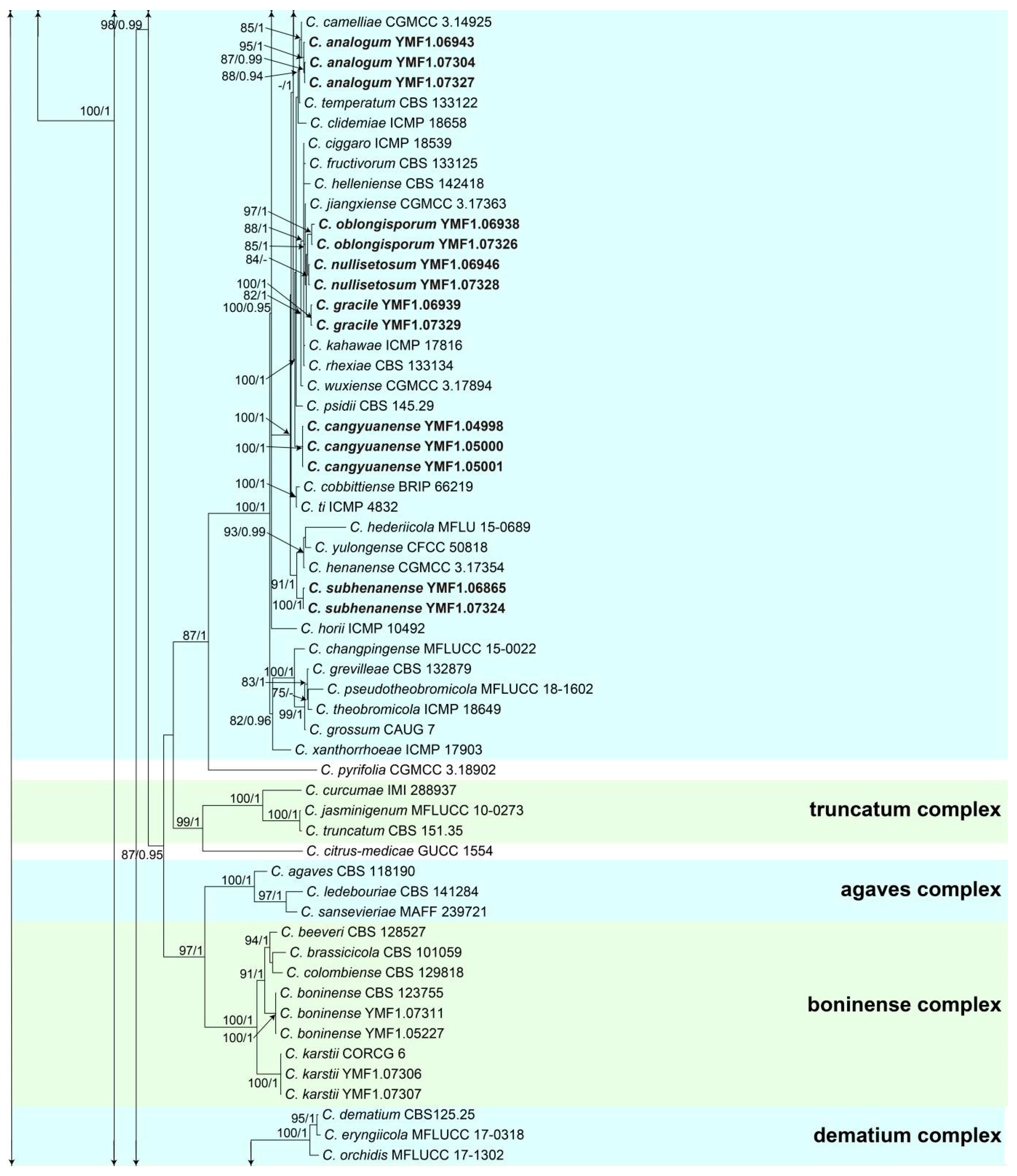
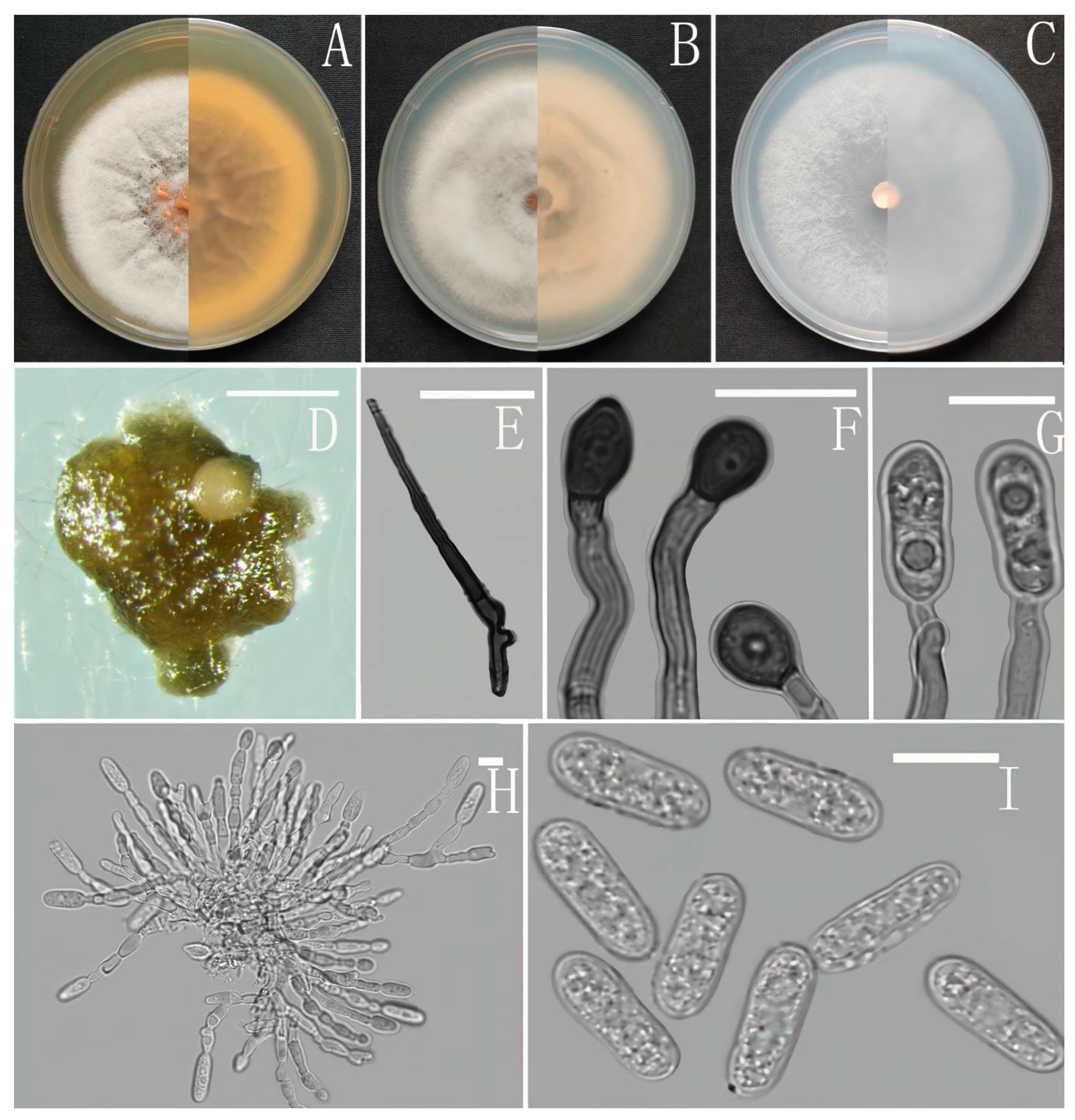
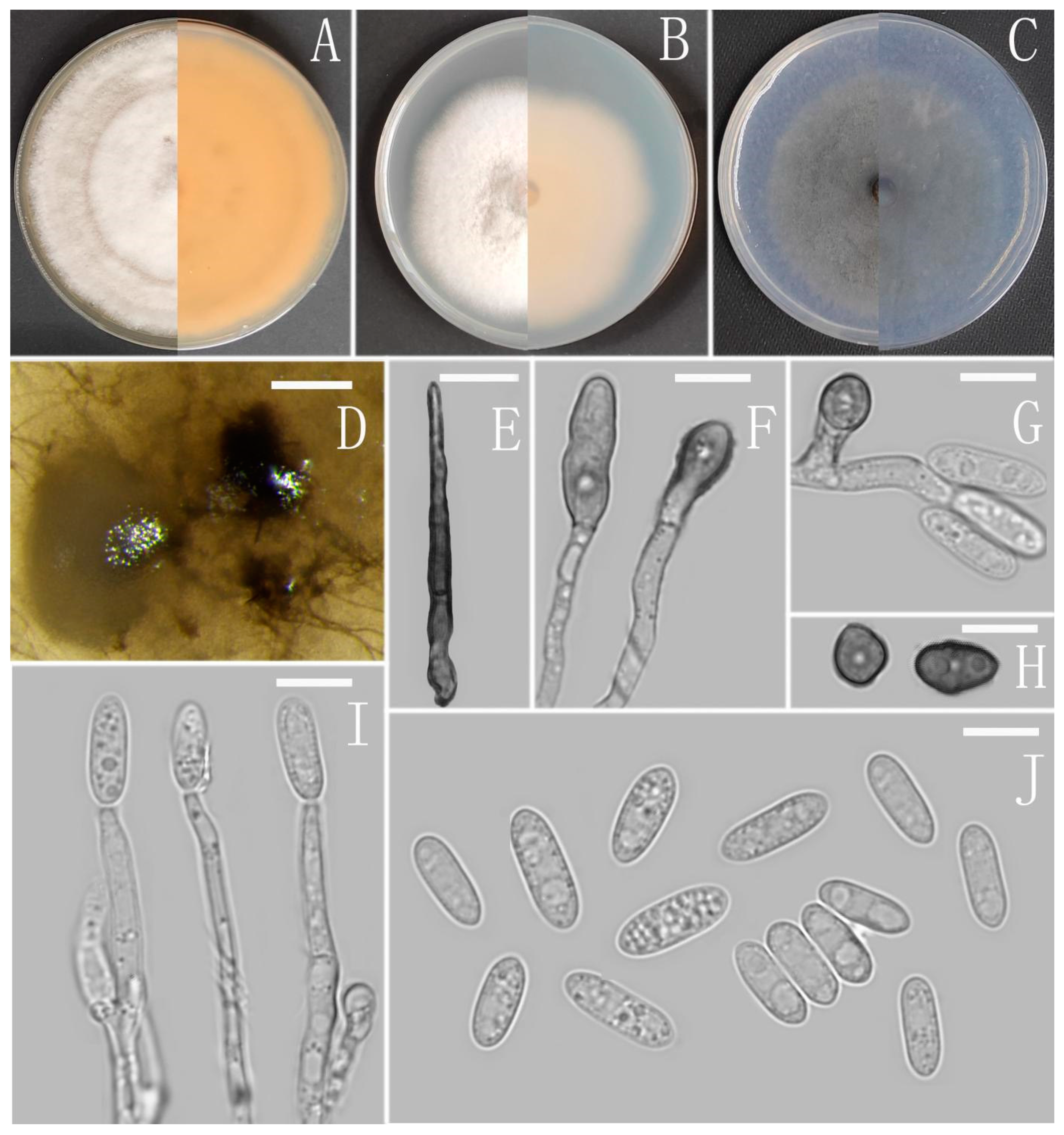
Colletotrichum analogum Z.F. Yu (Figure 3).
MycoBank Number: MB842547.
Etymology: The species epithet is based on the similarity or comparability with Colletotrichum clidemiae.
Type: China, Yunnan province, Ning’er county, from disease spot of Ageratina adenophora, August 2017, Han-Bo Zhang. Holotype YMF 1.06943, Ex-type culture CGMCC3.16079.
Sexual morph not observed. Asexual morph on CMA. Conidiomata rudimentary. Setae not observed. Chlamydospores pale yellow, 6.2–8.7 × 5.1–6.8 μm. Conidiophores directly formed on hyphae, hyaline, smooth-walled, 0–1-septate, unbranched. Conidiogenous cells hyaline, cylindrical, smooth-walled, collarette inconspicuous. Conidia cylindrical to clavate, unicellular, straight and slender, obtuse, subtruncate at the base, rounded at the apex, 15–20 × 5–6.5 μm, mean ± SD = 17.6 ± 1.4 × 5.5 ± 0.5 μm, l/w ratio = 3.2. Appressoria dark brown, septate, single or in small groups, irregular in outline, margin sometimes lobed, 9–18 × 6–10 μm.
Culture characteristics: Colonies on MEA flat with entire margin, reaching 48–50 mm diameter in 7 days at 25 °C, aerial mycelium fluffy, milky white, reverse same colors. Colonies on PDA reaching 84–88 mm diameter in 7 days at 25 °C, aerial mycelium dense, cottony, gray in the center and white in the margin, reverse grey to saffron with brown pigmentation ring. Colonies on SNA flat with entire margin, reaching 49–52 mm diameter in 7 days at 25 °C, medium buff to pale honey in the center, aerial mycelium fluffy, flocculent, pale yellow, white in edges, reverse buff to pale honey. Colonies on CMA reaching 39–40 mm diameter in 7 days at 25 °C, towards the margin honey to buff, partly covered with short felty whitish aerial mycelium, reverse olivaceous buff to olivaceous gray.
Additional specimens examined: China, Yunnan province, Ning’er county, from disease spot of Ageratina adenophora, August 2017, Han-Bo Zhang, living cultures YMF 1.07304, YMF1.07327.
Notes: Three strains of Colletotrichum analogum formed a sole clade in the gloeosporioides species complex, and the clade is close to C. camelliae Massee. Morphologically, C. analogum differs from C. camelliae by having narrowly cylindrical conidia, while those of C. camelliae are slightly constricted in the middle [65].
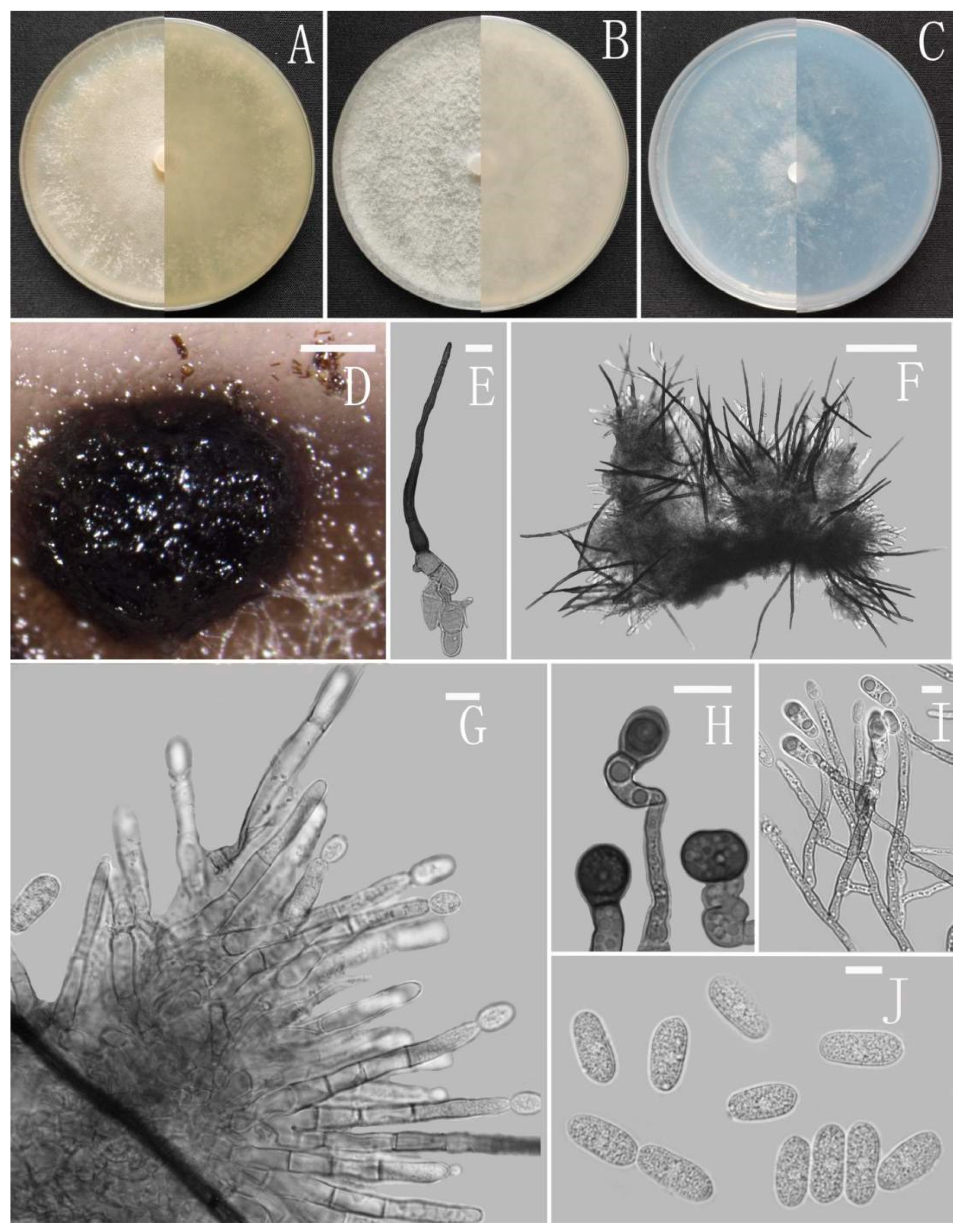
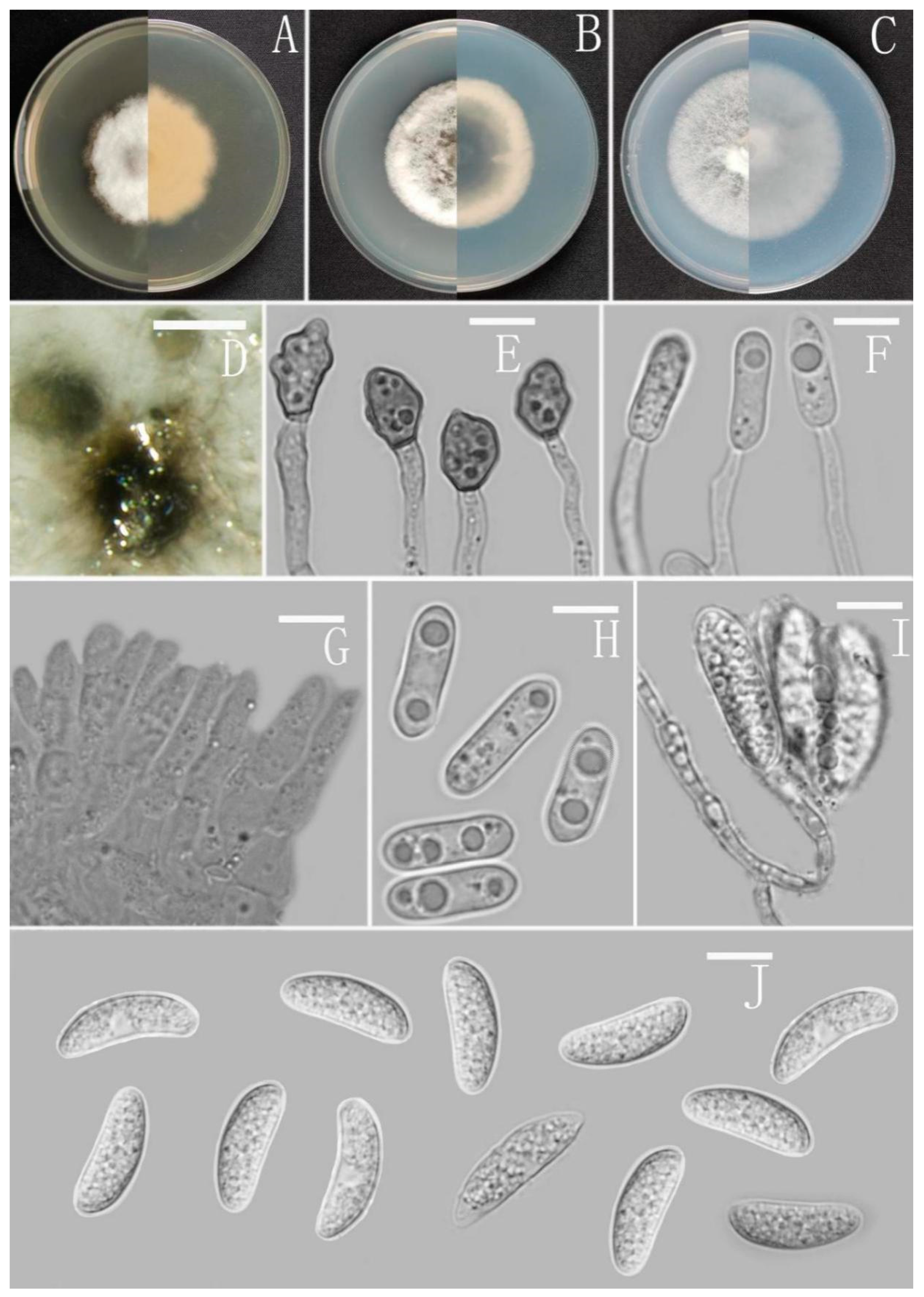
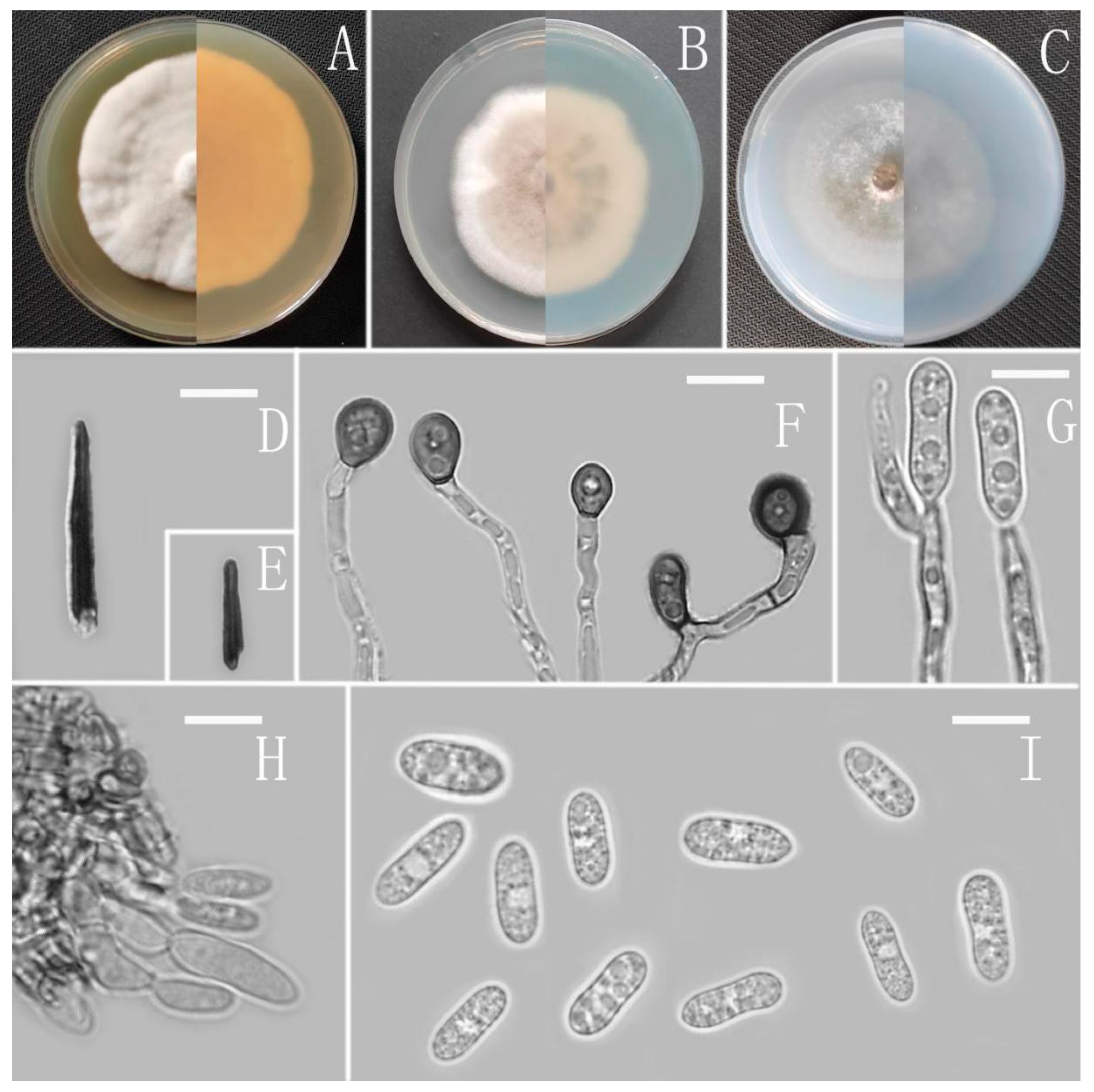
Colletotrichum cangyuanense Z.F. Yu (Figure 4).
MycoBank Number: MB842548.
Etymology: The species epithet refers to the county, where the type strain was collected, Cangyuan.
Type: China, Yunnan province, Cangyuan county, isolated as an endophyte from Ageratina adenophora, August 2016, Han-Bo Zhang. Holotype YMF 1.05001, Ex-type culture CGMCC3.18969.
Sexual morph not observed. Asexual morph on CMA. Conidiomata rudimentary, forming smaller aggregates of the mycelium with conidia droplets, dark olivaceous brown. Setae and conidiophores formed directly on hyphae. Setae scarce, cylindrical, straight or slightly flexuous, dark brown to black, 4–6-septate, base cylindrical to slightly inflated, tip acute to rounded, up to 102 μm. Conidiophores pale brown, aseptate, sometimes branched. Conidiogenous cells hyaline, smooth-walled, septate, subulate or cylindrical. Conidia hyaline, smooth-walled, cylindrical, slightly constricted at the middle, apex rounded, base obtuse, subtruncate, 15.5–20 × 4.5–5.5 μm, mean ± SD = 17.6 ± 1.3 × 5.1 ± 0.3 μm, l/w ratio = 3.5. Appressoria produced from mycelia, irregular, brown, branched, 12–16 × 6–11.5 μm.
Culture characteristics: Colonies on MEA flat forming a concentric ring with entire margin, occupied the whole plate in 7 days at 25 °C, surface grayish white, aerial mycelia medium hyaline to gray in the center, light gray to grayish white in the margin, partly covered with short felty whitish aerial mycelium, reverse same colors. Colonies on PDA flat forming a concentric ring with entire margin, occupied the whole plate in 7 days at 25 °C, aerial mycelia cotton-shaped, white to pale gray with circular margin, reverse same colors. Colonies on SNA flat with entire margin, occupied the whole plate in 7 days at 25 °C, surface covered with a radial, groove, thin, whitish aerial mycelium, reverse same colors. Colonies on CMA reaching 40–43 mm diameter in 7 days at 25 °C, aerial mycelium fluffy, flocculent, white, reverse same colors.
Additional specimens examined: China, Yunnan province, Cangyuan county, isolated as an endophyte from Ageratina adenophora, August 2016, Han-Bo Zhang, living cultures YMF 1.04998, YMF 1.05000.
Notes: Based on multi-locus phylogenetic analysis, three strains of Colletotrichum cangyuanense formed a sole clade in the gloeosporioides species complex, and the clade is close to C. psidii Curzi. Morphologically, C. cangyuanense shares similar morphological characteristics with other taxa in the gloeosporioides species complex, which have cylindrical conidia with rounded ends tapering slightly towards the base [66]. Nevertheless, C. cangyuanense is morphologically different from the phylogenetically nearest species C. psidii by having larger conidia, 15.5–20 × 4.5–5.5 vs. 12–15 × 3.5–4.5 μm [66].
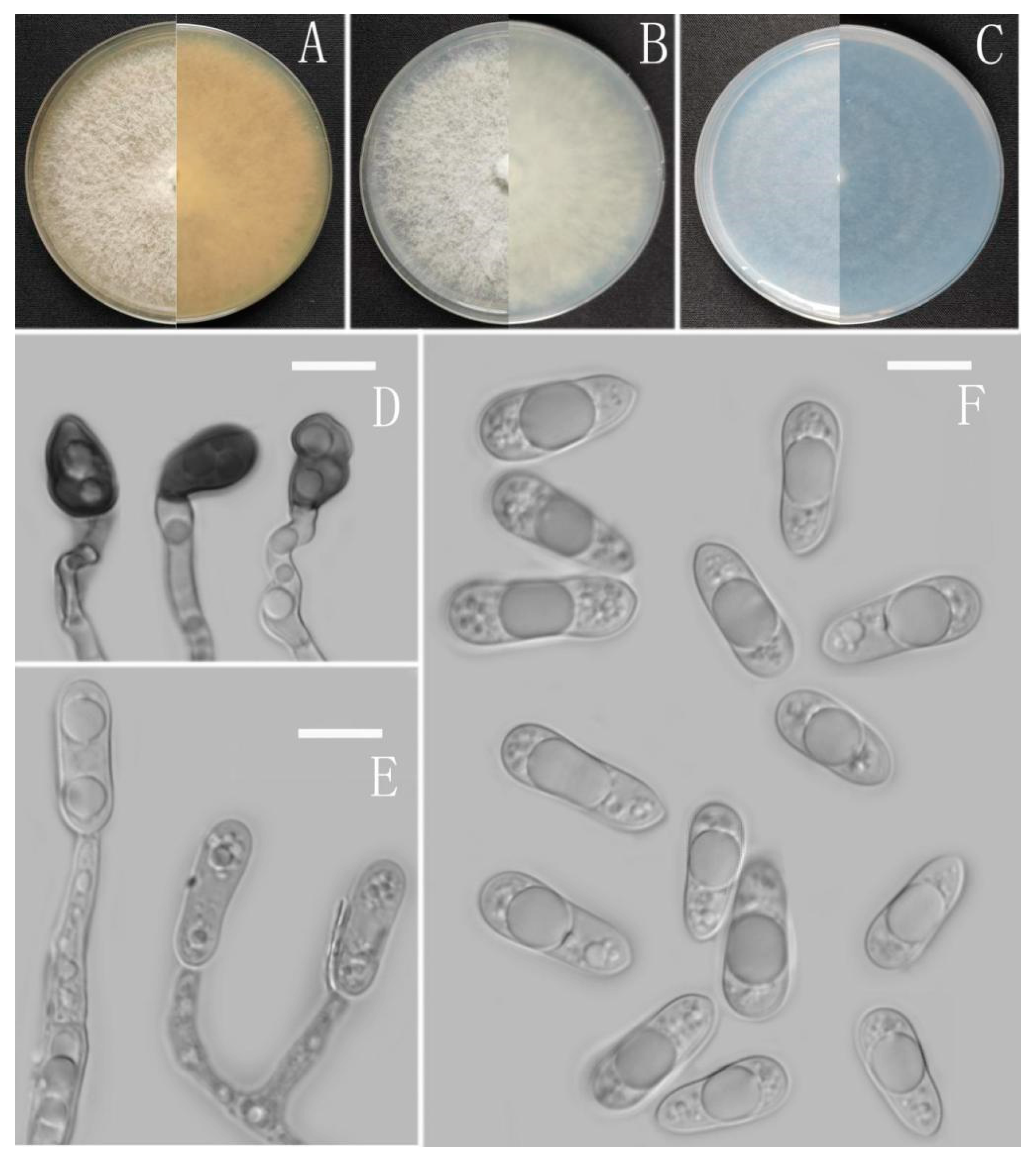
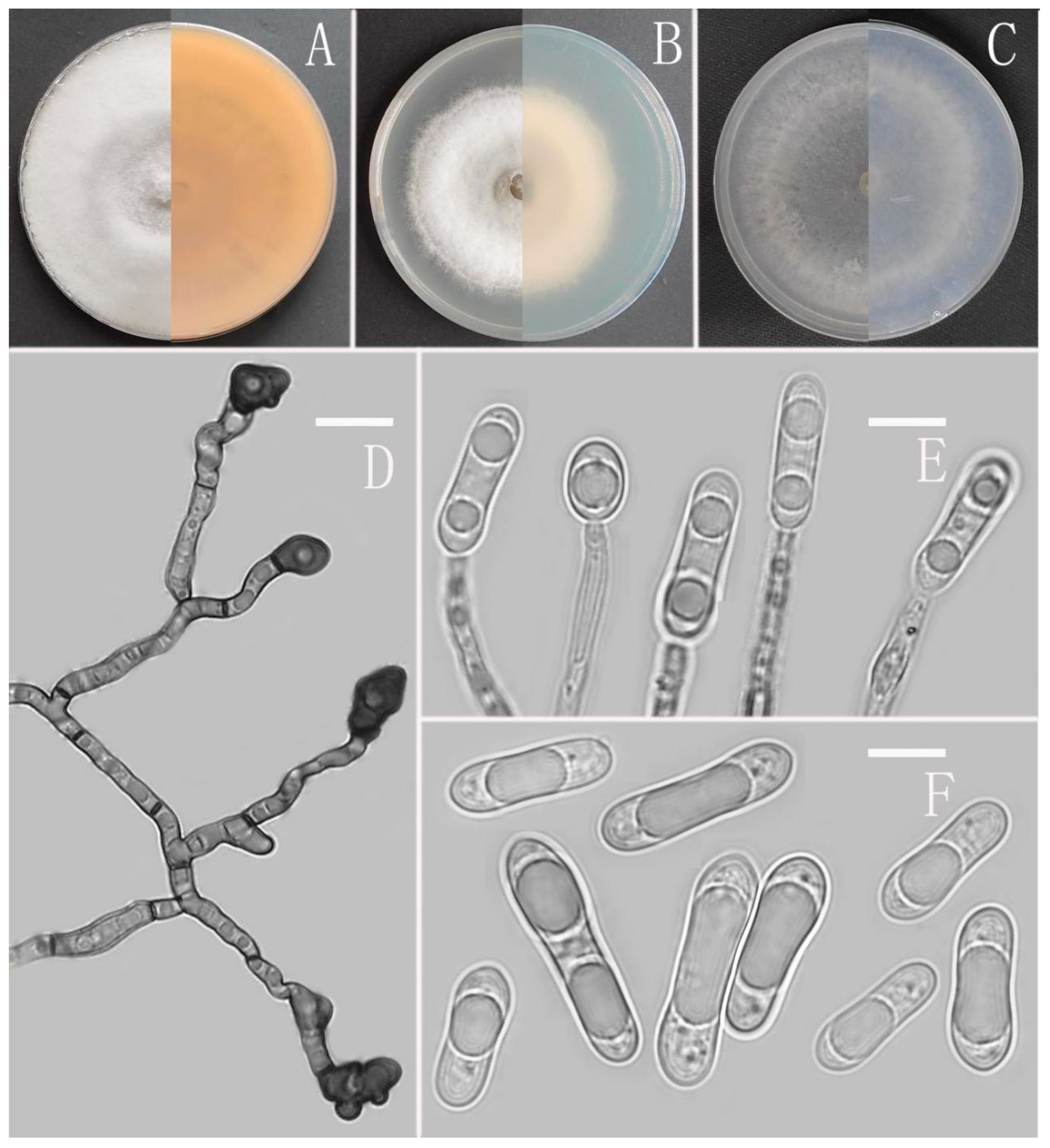
Colletotrichum dimorphum Z.F. Yu (Figure 5).
MycoBank Number: MB842549.
Etymology: The species epithet is based on the two forms of conidia.
Type: China, Guizhou province, Pingtang county, from disease spot of Ageratina adenophora, August 2018, Han-Bo Zhang. Holotype YMF 1.07309, Ex-type culture CGMCC3.16083.
Sexual morph not observed. Asexual morph on CMA. Conidiomata and setae not observed. Conidiophores directly formed on hyphae, hyaline, smooth-walled, unbranched. Conidiogenous cells hyaline, cylindrical, smooth-walled. Conidia hyaline, cylindrical to oblong, attenuate at the base, rounded at the apex, 10.5–19 × 4–6 μm, mean ± SD=14.6 ± 2 × 4.8 ± 0.7 μm, l/w ratio=3.1. Appressoria dark brown, aseptate, single, always elliptical in outline, the edge entire, 5.7–10.6 × 5–9 μm.
Culture characteristics: Colonies on MEA flat with entire margin, reaching 80 mm diameter in 7 days at 25 °C, surface grayish white, partly covered with fluffy, whitish aerial mycelium, light gray to grayish white in the margin, reverse medium grayish white to pale honey with brown pigmentation ring. Colonies on PDA flat with entire margin, occupied the whole plate in 7 days at 25 °C, aerial mycelia cotton-shaped, white to pale gray with circular margin, reverse iron grey to olivaceous grey. Colonies on SNA flat with entire margin, reaching 63–66 mm diameter in 7 days at 25 °C, aerial mycelia medium hyaline to grayish white in center, white to light gray in the margin, partly covered with short felty whitish aerial mycelium, reverse same colors. Colonies on CMA reaching 78 mm diameter in 7 days at 25 °C, aerial mycelium fluffy, flocculent, white, reverse same colors.
Additional specimen examined: China, Guizhou province, Pingtang county, from disease spot of Ageratina adenophora, August 2018, Han-Bo Zhang, living culture YMF1.07303.
Notes: Phylogenetically, Colletotrichum dimorphum fell into the gloeosporioides complex clade and near to the species C. gloeosporioides. Morphologically, C. dimorphum is distinguished from C. gloeosporioides by having slenderer conidia (l/w ratio: 3.1 vs. 2.6) and elliptical appressoria [67].
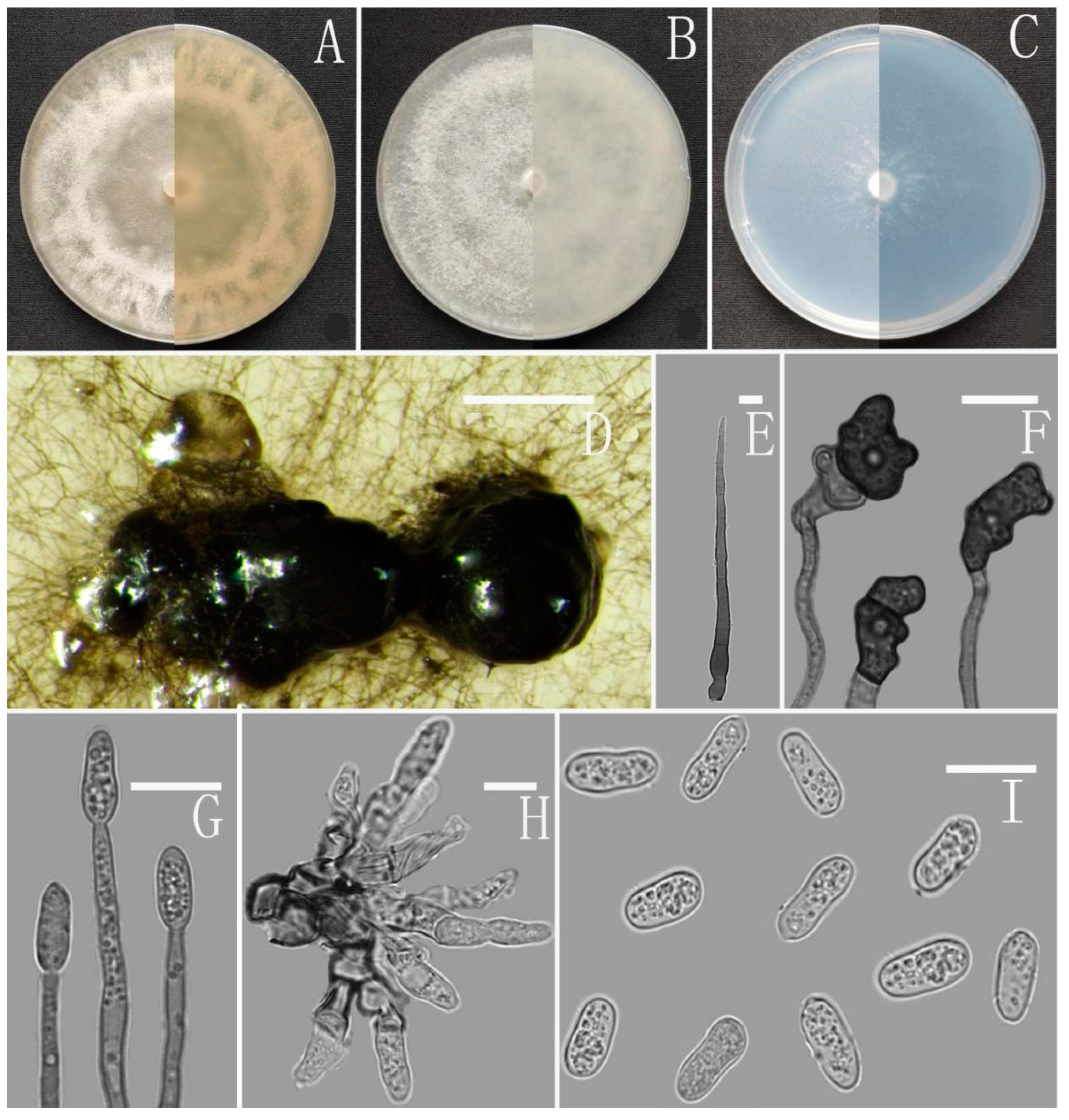
Colletotrichum gracile Z.F. Yu (Figure 6).
MycoBank Number: MB842550.
Etymology: The species epithet refers to the slender and thin conidia.
Type: China, Guizhou province, Nayong county, from disease spot of Ageratina adenophora, August 2018, Han-Bo Zhang. Holotype YMF 1.06939, Ex-type culture CGMCC3.16075.
Sexual morph on CMA: Clusters of ascoshell aggregates of various sizes were formed on the colony in the later stage of the culture. Ascomata perithecia, solitary, superficial or immersed, non-stromatic, medium to dark brown, subglobose to pyriform, 358–320 × 257–267 µm. Peridium composed of medium brown flattened textura angularis with cells 3.2–4.9 µm diameter. Asci unitunicate, 8-spored, cylindrical, tapering to apex and base, smooth-walled, the 8 ascospores are arranged in a hemp rope shape, 55–60 × 12–15 µm. Ascospores uni- or biseriately arranged, hyaline smooth-walled, allantoid, curved most in the middle, with rounded ends, 15–18 × 2–3 μm, mean ± SD = 17 ± 1.1 × 3 ± 0.4 μm, l/w ratio = 5.7.
Asexual morph on CMA: Conidiomata and setae not observed. Conidiophores directly formed on hyphae, hyaline, smooth-walled, aseptate, unbranched. Conidiogenous cells hyaline, cylindrical to clavate, smooth-walled, collarette absent. Conidia cylindrical to clavate, aseptate, straight or slightly curved, both ends obtusely rounded or one end slightly acute, 13–18 × 4–6 μm, mean ± SD = 15 ± 1.1 × 5.2 ± 0.4 μm, l/w ratio = 2.9. Appressoria dark brown, aseptate, unbranched, single and little, irregular but often subglobose to ellipsoidal, margin sometimes lobed, 7–17.5 × 5–9 μm.
Culture characteristics: Colonies on MEA flat with entire margin, occupied the whole plate in 7 days at 25 °C, aerial mycelium fluffy, milky white, reverse brown to buff. Colonies on PDA reaching 72–76 mm diameter in 7 days at 25 °C, aerial mycelium dense, cottony, gray in the center and white in the margin, reverse same colors. Colonies on SNA flat with entire margin, reaching 70–72 mm diameter in 7 days at 25 °C, aerial mycelia medium hyaline to grayish white in center, white in the margin, partly covered with short whitish aerial mycelium, reverse same colors. Colonies growing on CMA with entire margin, reaching 64–66 mm diameter in 7 days at 25 °C, aerial mycelia medium gray to pale buff in center, light gray to grayish white in the margin, entirely covered with floccose to dense, producing pale gray conidiomata in 2 weeks. Reverse dark white to gray with white margin.
Additional specimen examined: China, Guizhou province, Nayong county, from disease spot of Ageratina adenophora, August 2018, Han-Bo Zhang, living culture YMF1.07329.
Notes: Based on multi-locus phylogenetic analysis, two strains of Colletotrichum gracile formed a solitary clade in the gloeosporioides complex clade, and phylogenetically near to the two new species C. oblongisporum and C. nullisetosum. However, C. gracile is easily distinguished from them by observing sexual morph.
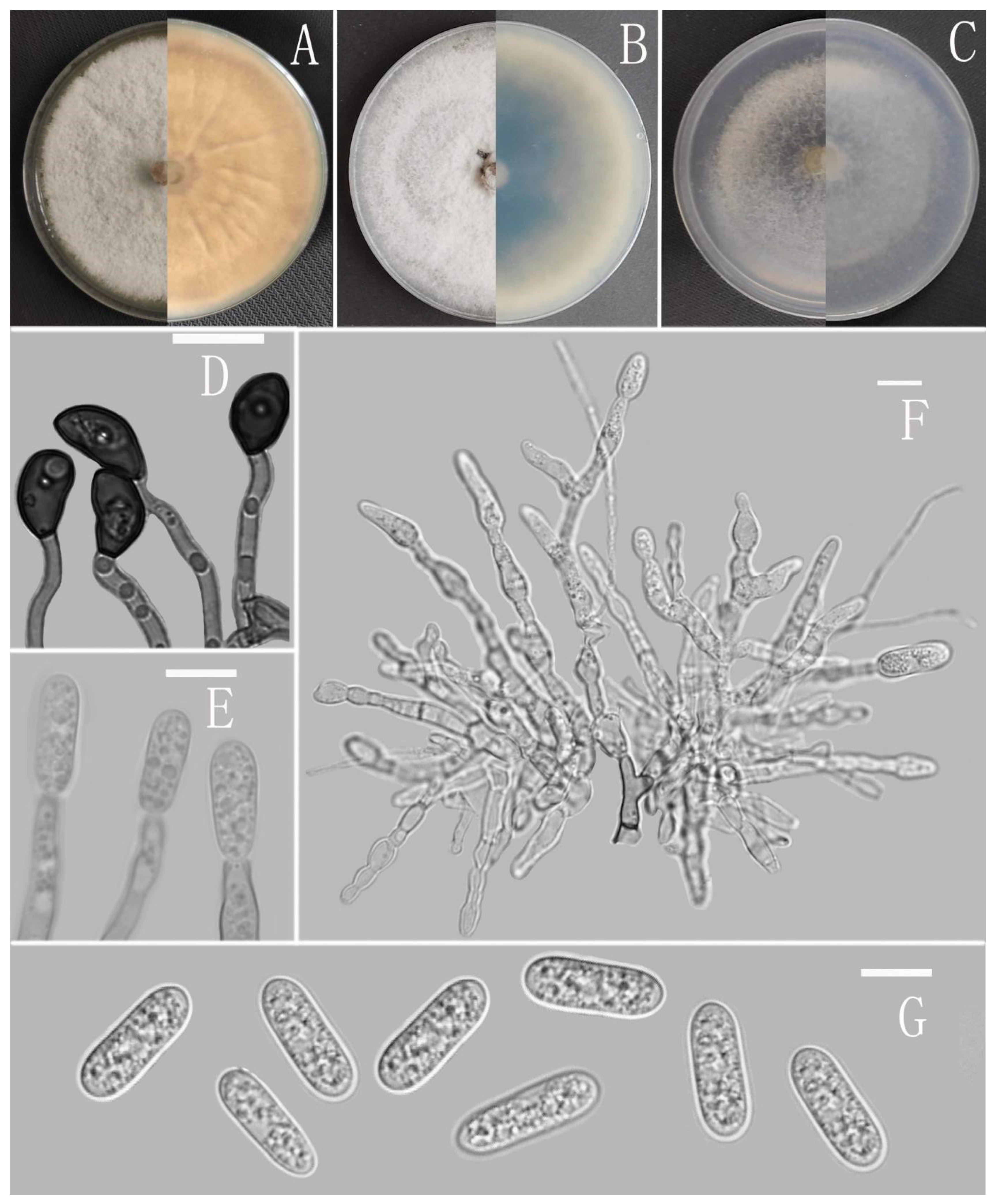
Colletotrichum nanhuaense Z. F. Yu (Figure 7).
MycoBank Number: MB842551.
Etymology: The species epithet refers to the county, where the type strain was collected, Nanhua.
Type: China, Yunnan province, Nanhua county, from disease spot of Ageratina adenophora, August 2017, Han-Bo Zhang. Holotype YMF1.04993, Ex-type culture CGMCC3.18962.
Sexual morph not observed. Asexual morph on CMA. Conidiomata rudimentary, dark olivaceous brown. Setae and conidiophores formed directly on hyphae. Setae scarce, cylindrical, straight, dark brown to black, 3–5-septate, base bent with bulge, tip acute to rounded, 25 μm. Conidiophores hyaline to pale brown, aseptate, unbranched. Conidiogenous cells hyaline, cylindrical, smooth-walled, collarette absent. Conidia straight cylindrical, oblong to narrowly ovoid, obtuse at the base, rounded at the apex, contents granular, 10.5–16 × 4.5–6 μm, mean ± SD=14 ± 1.1 × 5.4 ± 0.4 μm, l/w ratio = 2.6. Appressoria dark brown, irregular, margin sometimes lobed, 8–14 × 5–8 μm.
Culture characteristics: Colonies on MEA flat with entire margin, reaching 68–72 mm diameter in 7 days at 25 °C, surface covered with radial, groove, felty, whitish aerial mycelium, Conidiomata acervular, with orange conidial masses, reverse iron gray to buff. Colonies on PDA flat with entire margin, reaching 76–79 mm diameter in 7 days at 25 °C, aerial mycelium dense, cottony, gray in the center and white in the margin, reverse carneose to grayish white. Colonies on SNA flat with entire margin, reaching 70–73 mm diameter in 7 days at 25 °C, aerial mycelia medium hyaline to grayish white in the center, white in the margin, partly covered with short whitish aerial mycelium, producing conidiomata with yellow conidial masses in 2 weeks, reverse same colors. Colonies growing on CMA with entire margin, 73–74 mm diameter in 7 days at 25 °C, aerial mycelia medium gray to pale buff in center, light gray to grayish white in the margin, entirely covered with floccose to dense, producing pale gray conidiomata in 2 weeks. Reverse dark white to grey with white margin.
Additional specimen examined: China, Yunnan province, Nanhua county, from disease spot of Ageratina adenophora, August 2017, Han-Bo Zhang, living culture YMF 1.04990.
Notes: Phylogenetically, Colletotrichum nanhuaense fell into the gloeosporioides complex clade and near to C. gloeosporioides and the new species C. dimorphum. Nevertheless, C. nanhuaense is morphologically different from them by having setae [67].
Colletotrichum nullisetosum Z.F. Yu (Figure 8).
MycoBank Number: MB842556.
Etymology: The species epithet is based on the absence of setae.
Type: China, Yunnan province, Ning’er county, from disease spot of mango, August 2017, Han-Bo Zhang. Holotype YMF1.06946, Ex-type culture CGMCC3.16080.
Sexual morph not observed. Asexual morph on CMA. Conidiomata rudimentary, dark olivaceous brown. Setae not observed. Conidiophores directly formed on hyphae, hyaline, smooth-walled, unbranched. Conidiogenous cells hyaline, cylindrical to clavate, smooth-walled, collarette absent. Conidia unicellular, hyaline, dimorphic; (i) straight cylindrical, oblong to narrowly ovoid, obtuse at the base, rounded at the apex, contents 1–2-granular, 13.5–19.5 × 4–6.5 μm, mean ± SD = 15.5 ± 1.3 × 5 ± 0.5 μm, l/w ratio = 3.1. (ii) sickle, aseptate, blunt at both ends, curved to one side, contents 1-granular, 14.8–19.3 × 5–6.5, mean ± SD = 17.3 ± 1.1 × 5.8 ± 0.3 μm, l/w ratio = 3.0. Appressoria dark brown, sometimes branched, always little and oval in outline, contents granular, 9–15 × 5–8 μm.
Culture characteristics: Colonies on MEA reaching 44–45 mm diameter in 7 days at 25 °C, aerial mycelia cotton-shaped, grayish white to white, turning iron gray in the later stage, reverse same colors. Colonies on PDA reaching 49–51 mm diameter in 4 days at 25 °C, aerial mycelia cotton-shaped, white to pale gray with circular margin, reverse iron gray to olivaceous gray. Conidial mass whitish to pale yellow. Colonies on SNA reaching 58–60 mm diameter in 7 days at 25 °C, aerial mycelium fluffy, flocculent, white, producing black conidiomata in 2 weeks, reverse same colors. Colonies on CMA reaching 57–60 mm diameter in 7 days at 25 °C, aerial mycelium fluffy, flocculent, white, reverse same colors.
Additional specimen examined: China, Yunnan province, Ning’er county, from disease spot of the common fruit mango, August 2017, Han-Bo Zhang, living culture YMF 1.07328.
Notes: Colletotrichum nullisetosum is phylogenetically close to the new species C. oblongisporum, but it can be distinguished from C. oblongisporum by having dimorphic conidia.
Colletotrichum oblongisporum Z.F. Yu (Figure 9).
MycoBank Number: MB842557.
Etymology: The species epithet refers to oblong shape of conidia.
Type: China, Yunnan province, Kunming city, Xishan Park, isolated as an endophyte from Ageratina adenophora, September 2017, Han-Bo Zhang. Holotype YMF1.06938, Ex-type culture CGMCC3.16074.
Sexual morph not observed. Asexual morph on CMA. Conidiomata and setae not observed. Conidiophores directly formed on hyphae, hyaline, smooth-walled, septate, occasionally branched. Conidiogenous cells hyaline, cylindrical, collarette not visible. Conidia oblong, unicellular, subtruncate at the base, biguttulate near the ends, hyaline, smooth-walled, 13–18.5 × 4–5.5 μm, mean ± SD = 15.3 ± 1.4 × 4.7 ± 0.4 μm, l/w ratio = 3.3. Appressoria dark brown, subglobose, cuneiform to irregular, sometimes with lobate margin, 8–15 × 4.5–7.5 μm.
Culture characteristics: Colonies on MEA flat with entire margin, occupied the whole plate in 7 days at 25 °C, aerial mycelium fluffy, flocculent, white on the edges, reverse white to light yellow. Colonies on PDA flat with entire margin, occupied the whole plate in 7 days at 25 °C, aerial mycelium dense, cottony, in the later stage of culture, formed a four concentric ring pattern, from the center to the edge, pale gray to dark gray. Colonies on SNA flat with entire margin, occupied the whole plate in 7 days at 25 °C, aerial mycelium white, formed in the shape of four concentric ring pattern, reverse same colors. Colonies on CMA flat with entire margin, occupied the whole plate in 7 days at 25 °C, aerial mycelium fluffy, flocculent, white, reverse same colors.
Additional specimen examined: China, Yunnan province, Kunming city, Xishan Park, isolated as an endophyte from Ageratina adenophora, September 2017, Han-Bo Zhang, living culture YMF 1.07326.
Notes: Based on multi-locus phylogenetic analysis, two strains of Colletotrichum oblongisporum formed a solitary clade in the gloeosporioides complex clade, and phylogenetically near to the two new species C. oblongisporum and C. nullisetosum, but it can be easily distinguished from them in morphology. For instance, C. gracile differs from C. oblongisporum in the shape of conidia and appressoria. C. gracile has cylindrical to clavate conidia and subglobose to ellipsoidal appressoria, while C. oblongisporum has oblong conidia and subglobose and cuneiform to irregular appressoria.
Colletotrichum parvisporum Z.F. Yu (Figure 10).
MycoBank Number: MB842558.
Etymology: The species epithet is based on the relatively smaller conidia.
Type: China, Guangxi province, Debao county, from disease spot of Ageratina adenophora, August 2018, Han-Bo Zhang. Holotype YMF 1.06942, Ex-type culture CGMCC3.16078.
Sexual morph not observed. Asexual morph on CMA. Conidiomata and setae not observed. Conidiophores formed directly from mycelia, hyaline, sometimes branched. Conidiogenous cells hyaline, smooth-walled, cylindrical or oval, collarette not visible. Conidia oval or cylindrical, both with broadly rounded ends, straight or sometimes slightly bent, sometimes contracted in the middle, the contents are circular, 10–15 × 3.5–7.5 μm, mean ± SD = 12.5 ± 1.3 × 5.8 ± 0.9 μm, l/w ratio = 2.2. Appressoria produced from mycelia, clavate or irregular outline, aseptate, brown, 7.5–17 × 4.5–10 μm, mean ± SD = 11.7 ± 2.1 × 7.2 ± 1.6 μm.
Culture characteristics: Colonies on MEA reaching 55–58 mm diameter in 7 days at 25 °C, aerial mycelia cotton-shaped, grayish white to white, turning to iron gray in the later stage, reverse same colors. Colonies on PDA reaching 40–49 mm diameter in 5 days at 25 °C, felted aerial mycelia with grayish white appearance, reverse grayish white. Colonies on SNA flat with entire margin, reaching 65–69 mm diameter in 7 days at 25 °C, aerial mycelia medium hyaline to grayish white in center, white in the margin, partly covered with short whitish aerial mycelium, reverse same colors. Colonies on CMA flat with entire margin, reaching 50–54 mm diameter in 7 days at 25 °C, aerial mycelia hyaline to greyish white in center, white in the margin, partly covered with short whitish aerial mycelium, reverse same colors.
Notes: Phylogenetically, Colletotrichum parvisporum clustered with C. pandanicola Tibpromma and K.D. Hyde in the gloeosporioides complex clade. C. pandanicola was introduced by Tibpromma et al. in 2018; it was recorded as an endophyte isolated from the leaves of Pandanus sp. in Thailand and sexual morph was not reported [68]. Morphologically, C. pandanicola is easily distinguished from the new species C. parvisporum by having cylindrical conidia and lacking appressoria [68].
Colletotrichum robustum Z. F. Yu (Figure 11).
MycoBank Number: MB842559.
Etymology: The species epithet is based on the conidia, meaning robust, strong and vigorous.
Type: China, Guizhou province, Dushan county, on disease spot of Ageratina adenophora, August 2018, Han-Bo Zhang. Holotype YMF 1.06941, Ex-type culture CGMCC3.16077.
Sexual morph not observed. Asexual morph on CMA. Conidiomata rudimentary, pale brown. Setae and conidiophores formed directly on hyphae. Setae abundant, cylindrical, straight or slightly flexuous, dark brown to black, 3–5-septate, base cylindrical to slightly inflated, tip acute to rounded, 11.8–15.1 μm long. Conidiophores abundant, hyaline, branched. Conidiogenous cells hyaline, cylindrical to clavate, collarette not visible. Conidia smooth-walled, hyaline, broad cylindrical, rounded at the ends, sometimes slightly constricted in the middle, the circular or elliptical guttulate. 12.5–16.0 × 5.5–7.5 μm, mean ± SD = 14.0 ± 0.8 × 6.5 ± 0.5 μm, l/w ratio = 2.2. Appressoria circular or irregular outline, brown, branched, 6.0–8.0 × 5.0–7.0 μm.
Culture characteristics: Colonies on MEA flat forming a concentric ring with entire margin, occupied the whole plate in 7 days at 25 °C, surface grayish white, aerial mycelia medium hyaline to gray in center, light gray to grayish white in the margin, partly covered with short felty whitish aerial mycelium, reverse same colors. Colonies on PDA flat with entire margin, occupied the whole plate in 7 days at 25 °C, aerial mycelium fluffy, flocculent, white in edges, reverse white to light yellow, aerial mycelium sparse, white to pale gray, with copious yellow conidial masses in the later stage of culture, reverse pale luteous to sienna with circinate black colored pigmentation. Colonies on SNA flat is concentric ring with entire margin, occupied the whole plate in 7 days at 25 °C, aerial mycelia medium white to hyaline in center, partly covered with short felty whitish aerial mycelium, reverse same colors. Colonies on CMA flat with entire margin, reaching 68 mm diameter in 7 days at 25 °C, aerial mycelia medium hyaline to grayish white in center, white in the margin, partly covered with a short, whitish aerial mycelium, reverse same colors.
Notes: In this phylogenetic tree, Colletotrichum robustum fell into the dracaenophium complex clade, and clustered with C. excelsum-altitudinum G. Tao, Zuo Y. Liu and L. Cai and C. tongrenense S.X. Zhou, J.C. Kang and K.D. Hyde. However, C. robustum can be distinguished from them in morphology. C. excelsum-altitudinum was isolated as an endophyte from Bletilla ochracea in Guizhou [42]. It has larger conidia and appressoria than C. robustum but has a slower growth rate on PDA. C. tongrenense was also isolated as an endophyte from Nothapodytes pittosporoides in Guizhou [69]. C. tongrenense is obviously different in having shorter conidia, 11–14 × 5–7 μm, and lacking appressoria.
Colletotrichum simulanticitri Z. F. Yu (Figure 12).
MycoBank Number: MB842560.
Etymology: The species epithet is based on the similarity to Colletotrichum citri.
Type: China, Yunnan province, Dali county, from disease spot of Betula sp., August 2016, Han-Bo Zhang. Holotype YMF 1.07302, Ex-type culture CGMCC 3.16082.
Sexual morph not observed. Asexual morph on CMA. Conidiomata rudimentary, stromatic, subpulvinate, dark brown. Setae cylindrical to subulate, obtuse at the apex, 1–2-septate, smooth-walled, dark brown to black, 33–44 μm. Conidiophores abundant, hyaline, smooth-walled, septate, branched. Conidiogenous cells hyaline, cylindrical. Conidia hyaline, cylindrical to oblong, attenuated at the base, rounded at the apex, unicellular, guttulate, 10–13.5 × 4–5 μm, mean ± SD = 12 ± 1 × 4.3 ± 0.3 μm, l/w ratio = 2.8. Appressoria dark brown, septate, clavate or oval, smooth margin, 6–15.5 × 4–5.5 μm.
Culture characteristics: Colonies on MEA flat forming a concentric ring with entire margin, occupied the whole plate in 7 days at 25 °C, white to grayish white in the margin, partly covered with short felty whitish aerial mycelium, reverse white to buff. Colonies on PDA reaching 60–63 mm diameter in 7 d at 25 °C, felted aerial mycelia with grayish white appearance, reverse grayish white, aerial mycelia turning dark gray and later becoming cotton-like lately. Colonies on SNA flat with entire margin, reaching 59–61 mm diameter in 7 days at 25 °C, aerial mycelia medium iron gray in center, light gray in the margin, partly covered with short felty gray aerial mycelium, reverse same colors. Colonies on CMA flat with entire margin, reaching 57–61 mm diameter in 7 days at 25 °C, aerial mycelia dark brown, conidiomata scarced, dark, reverse same colors.
Additional specimens examined: China, Yunnan province, Dali county, from disease spot of Betula sp., August 2016, Han-Bo Zhang, living cultures YMF 1.07312, YMF 1.07308.
Notes: Based on multi-locus phylogenetic analysis, three strains of Colletotrichum simulanticitri formed a solitary clade in the gloeosporioides complex clade and near to C. nymphaeae (Pass.) Aa and C. citri F. Huang, L. Cai, K.D. Hyde and Hong Y. Li. Morphologically, C. simulanticitri can be distinguished from C. nymphaeae by having setae and and shorter conidia (10–13.5 μm vs. 14–18.5 μm) [70]. C. simulanticitri is distinguished from C. citri by the conidia shape and size. The shape conidia of C. citri is fusiform and acute to near-rounded at two ends, while C. simulanticitri is cylindrical to oblong and attenuate at the base and rounded at the apex [38]. Moreover, the conidia of C. simulanticitri are wider than C. citri, 4–5 μm vs. 3.3–4.4 μm.
Colletotrichum speciosum Z.F. Yu (Figure 13).
MycoBank Number: MB842562.
Etymology: The species epithet refers to the showy and splendid culture characteristics. Type: China, Yunnan province, Simao county, isolated as an endophyte from Ageratina adenophora, August 2016, Han-Bo Zhang. Holotype YMF 1.07301, Ex-type culture CGMCC3.16081.
Sexual morph not observed. Asexual morph on CMA. Conidiomata not observed. Setae scarce, cylindrical, straight, dark brown to black, 1–2-septate, obtuse or with a bulge at the apex, tip acute to rounded, 12–21 μm. Conidiophores formed directly on aerial mycelium, hyaline, 0–1-septate, sometimes branched. Conidiogenous cells hyaline, thin cylindrical to clavate, collarette inconspicuous. Conidia hyaline, unicellular, smooth-walled, cylindrical, slightly constricted at the middle, attenuated at the base, rounded at the apex, lumen granular, 10.5–14.5 × 4–5.5 μm, mean ± SD = 12.5 ± 1.0 × 4.8 ± 0.4 μm, l/w ratio = 2.6. Appressoria dark brown, broadly oboval to obovoid or subglobose, the margin smooth, 7–9.5 × 4.5–7.5 μm.
Culture characteristics: Colonies on MEA flat forming concentric ring with radial groove, reaching 65–67 mm diameter in 7 days at 25 °C, covered with short, felty, whitish aerial mycelium, reverse white to buff. Colonies on PDA reaching 59–61 mm diameter in 7 days at 25 °C, covered with felty aerial mycelium, whitish to pale gray with circular margin, reverse olivaceous gray to pale gray. Colonies on SNA flat with entire margin, reaching 58–60 mm diameter in 7 days at 25 °C, aerial mycelia medium grayish white in the center, light gray to hyaline in the margin, reverse pale gray to hyaline. Colonies on CMA flat with entire margin, reaching 59 mm diameter in 7 days at 25 °C, covered with thin whitish aerial mycelium, producing dark conidiomata in 2 weeks, reverse same colors.
Notes: The phylogenetic analyses based on multi-locus revealed that Colletotrichum speciosum belongs to the acutatum species complex and is closely related to C. chrysanthemi (Hori) Sawada and C. carthami (Fukui) S. Uematsu, Kageyama, Moriwaki & Toy. Sato. However, C. chrysanthemi is morphologically different from C. speciosum by its very short acute-ended conidia [70]. Furthermore, C. carthami also differs from C. speciosum by having fusiform to oblong and narrower (2.6–4.6 μm) conidia [71].
Colletotrichum subhenanense Z.F. Yu (Figure 14).
MycoBank Number: MB842563.
Etymology: The species epithet is based on the species Colletotrichum henanense, which is phylogenetically closely related.
Type: China, Yunnan province, Cangyuan county, from disease spot of Ageratina adenophora, August 2016, Han-Bo Zhang. Holotype YMF 1.06865, Ex-type culture CGMCC3.16073.
Sexual morph not observed. Asexual morph on CMA. Conidiomata and setae not observed. Conidiophores directly formed on hyphae, hyaline, smooth-walled, septate, sometimes branched. Conidiogenous cells hyaline, cylindrical, smooth-walled, collarette absent. Conidia plump cylindrical, oblong to narrowly ovoid, unicellular, hyaline, guttulate, straight or slightly curved, obtuse at the base, rounded at the apex, 10–20 × 4–6.5 μm, mean ± SD = 14.3 ± 1.9 × 5 ± 0.6 μm, l/w ratio = 2.9. Appressoria dark brown, septate, single, irregular but often subglobose in outline, margin smooth, 7–13 × 6–10 μm.
Culture characteristics: Colonies on MEA flat with entire margin, occupied the whole plate in 7 days at 25 °C, aerial mycelium fluffy, milky white, reverse pale gray to buff. Colonies on PDA reaching 59–62 mm diameter in 7 days at 25 °C, aerial mycelium dense, cottony, gray in the center and white in the margin, turning dark later, reverse olivaceous to grayish white. Colonies on SNA flat with entire margin, reaching 85–88 mm diameter in 7 days at 25 °C, aerial mycelia medium hyaline to grayish white in the center, white in the margin, partly covered with short, whitish aerial mycelium, reverse same colors. Colonies on CMA reaching 75–78 mm diameter in 7 days at 25 °C, aerial mycelium fluffy, flocculent, white, turning dark later, reverse same colors.
Additional specimen examined: China, Yunnan province, Cangyuan county, from disease spot of Ageratina adenophora, August 2016, Han-Bo Zhang, living culture YMF 1.07324.
Notes: Based on multi-locus phylogenetic analysis, two strains of Colletotrichum subhenanense formed a solitary clade in the gloeosporioides complex clade, and near to C. henanense F. Liu and L. Cai, C. yulongense C.L. Hou and X.T. Liu, and C. hederiicola Jayaward. Camporesi and K.D. Hyde. Morphologically, C. subhenanense is most similar to C. yulongense in culture characteristics on PAD and SNA, with cylindrical conidia with obtuse to slightly rounded ends, but C. yulongense is obviously distinguished by having septate setae and narrower conidia (4–5 μm) [72].
Colletotrichum yuanjiangense Z.F. Yu (Figure 15).
MycoBank Number: MB842564.
Etymology: The species epithet is derived from the county, where the type strain was collected, Yuanjiang.
Type: China, Yunnan province, Yuanjiang county, on disease spot of Ageratina adenophora, August 2017, Han-Bo Zhang. Holotype YMF 1.04996, Ex-type culture CGMCC3.18964.
Sexual morph not observed. Asexual morph on CMA. Conidiomata rudimentary, dark olivaceous brown. Setae and conidiophores formed directly on hyphae. Setae scarce, cylindrical, straight or slightly flexuous, dark brown to black, 4–6-septate, base cylindrical to slightly inflated, tip acute to rounded, 62 μm. Conidiophores directly formed on hyphae, hyaline, smooth-walled, aseptate, unbranched. Conidiogenous cells hyaline, cylindrical to clavate, smooth-walled, collarette absent. Conidia hyaline, smooth-walled, cylindrical, obtuse at both ends or slightly acute at one end, 10–14 × 4–6 μm, mean ± SD=12 ± 0.9 × 5.2 ± 0.5 μm, l/w ratio = 2.3. Appressoria dark brown, aseptate, single, subglobose or elliptical, the edge entire, sometimes slightly lobed, 7–13.5 × 6–10 μm.
Culture characteristics: Colonies on MEA flat with entire margin, reaching 38–42 mm diameter in 7 days at 25 °C, aerial mycelia medium buff to white, hyaline in the margin, partly covered with a short, whitish aerial mycelium, reverse buff to pale honey. Colonies on PDA reaching 80 mm diameter in 7 days at 25 °C, aerial mycelium cottony, pale gray in the center, grayish white at the edge, reverse olivaceous to grayish white. Colonies on SNA flat with entire margin, reaching 68–73 mm diameter in 7 days at 25 °C, aerial mycelia medium hyaline to white in center, white in the margin, partly covered with short whitish aerial mycelium, reverse same colors. Colonies growing on CMA with entire margin, 72–74 mm diameter in 7 days at 25 °C, aerial mycelia medium gray to pale buff in center, light gray to grayish white in the margin, entirely covered with floccose to dense, producing pale gray conidiomata in 2 weeks. Reverse same colors.
Additional specimen examined: China, Yunnan province, Yuanjiang county, from disease spot of Ageratina adenophora, August 2017, Han-Bo Zhang, living culture YMF 1.04997.
Notes: In this phylogenetic tree, Colletotrichum yuanjiangense fell into the gloeosporioides complex clade, and near to the two new species, C. nanhuaense and C. dimorphum, and C. gloeosporioides. Morphologically, C. yuanjiangense is most similar to C. nanhuaense in having septate setae and cylindrical conidia, but it can be distinguished by having subglobose or elliptical appressoria with an irregular edge.
4. Discussion
Although plant invasion is common and intensively studied [73,74], fungi associated with the host have received relatively less attention. A few cases have indicated that diverse endophytes were found in the leaves of invasive plants, so these fungi were co-introduced as well as local infection [16]. Regardless of the case, fungus identity is less known. Invasive plants invade a new range and commonly establish novel associations with local microbes [75,76]. Thus, invasive plants are good sources for novel microbial species. In this study, it is surprising that the diversity of Colletotrichum species was so abundant in Ageratina adenophora. We revealed 26 species associated with seven species complexes, including 14 new species C. adenophorae, C. analogum, C. cangyuanense, C. dimorphum, C. gracile, C. nanhuaense, C. nullisetosum, C. oblongisporum, C. parvisporum, C. robustum, C. simulanticitri, C. speciosum, C. subhenanense, C. yunajiangense and 12 known species C. boninense, C. citri, C. cosmi, C. fioriniae, C. fructicola, C. gigasporum, C. godetiae, C. guajavae, C. karstii, C. magnum, C. nymphaeae, C. plurivorum.
Although Colletotrichum is a common endophyte in most plants, its association with specific fungal taxa or its high relative abundance suggests it has a major effect on plant communities, given that Colletotrichum consistently occurs abundantly in the endophyte communities of A. adenophora. We hypothesized a pathogenic role in A. adenophora. The detection of many Colletotrichum spp., including the 14 novel species, in this study indicated the potential of Colletotrichum spread with A. adenophora invasion. The pathogenicity of these Colletotrichum strains (not shown here) indicated the necessity of investigating fungal endophytes in A. adenphora, as well as other invasive plants.
Recent evidence has indicated that the diversity of pathogens on highly abundant introduced hosts has been positively correlated over time with the geographical range of the introduced species and the diversity of invaded habitats [77]. Invasive species Microstegium vimineum (Trin.) A. Camus has been proven to be infected by Bipolaris spp. Shoemaker in the eastern USA [78]. Day et al. [26] isolated fungi from the roots of invasive Vincetoxicum rossicum (Kleopow) Barbar. (Apocynaceae) plants collected across Ontario, Canada, and indicated that some of them can reduce the total biomass of a native species Solidago Canadensis L., which may be important for mediating plant community structure and the persistence of invasive plants. Several studies have also found accompanying changes in populations of specific soil pathogens and their impacts on invasive and noninvasive species [79,80]. Recently, fungi isolated from the invasive plant Vincetoxicum rossicum have been proven to have differential growth effects between non-native invasive and native plants, which may be important for mediating plant community structure and the persistence of invasive plants [26]. Accumulations of pathogens are common in several detected invasive plants, including above-ground, for example, leaves [78], and belowground, e.g., soil oomycete pathogens in freshwater wetland soils invaded by non-native Phragmites australis (Cav.) Trin. ex Steud. (European common reed) [80]. We found that rhizosphere soils of Chromolaena odorata L., one of the world’s most destructive tropical invasive weeds, accumulate high concentrations of the generalist soil-borne fungi, Fusarium Link (tentatively identified as F. semitectum), thus creating a negative feedback for native plant species [79]. If Colletotrichum is pathogenic on A. adenophora, an abundance of this species may alleviate the spread of A. adenophora. If it is pathogenic on native plants, including fruits, the risk is great and should be emphasized in future.
Previously, association with the fungal endophyte Neotyphodium coenophialum (Morgan-Jones and W. Gams) Glenn et al. increased the ability of Lolium arundinaceum (Schreb.) S.J. Darbyshire to invade communities with greater species diversity. In the absence of the endophyte, the initial diversity of the community significantly reduced the establishment of L. arundinaceum. However, establishment was independent of initial diversity in the presence of the endophyte [81]. We tested the impact of the fungal endophyte Alternaria alternata (Fr.) Keissl. (phylotype CID 120) on the allelopathic effect of the invasive forb Centaurea stoebe when in competition with the North American native bunchgrass Koeleria macrantha (Ledeb.) Schultes in a greenhouse competition experiment. The allelopathic effect on K. macrantha of C. stoebe when infected with the fungal endophyte was more than twice that of endophyte-free C. stoebe. Fungal endophytes may increase the competitive ability of their hosts [23]. Two phylotypes of Alternaria endophytes influenced the growth, competitive effects, and competitive responses of the exotic invasive forb C. stoebe. The competitive effects of C. stoebe on grass species native to North America were enhanced by both fungal endophytes [24]. Arnold et al. [40] indicated that Colletotrichum-enhanced cacao trees resist pathogen attack. In this case, the role of the foliar fungus Colletotrichum sp. may be a weak latent foliar pathogen of A. adenophora and can enhance the pathogenicity of the fungus Diaporthe helianthi on the leaves of A. adenophora [31]. However, Colletotrichum may also as an endophyte of A. adenophora to resist herbivores, for example, Fang K [31] inoculated Collectotrichum into A. adenophora to enhance resistance herbivores. Therefore, the role of Colletotrichum in A. adenophora invasion or its interaction with fungal biocontrol agents is very complex and needs more investigation.
In conclusion, our study reported the diversity of Colletotrichum in the leaves of A. adenophora and several surrounding crops in the introduced range in China. There were several significant agronomic findings. Firstly, there is a potential application for controlling weeds in agriculture, although it remains to be clarified whether these endophytic Colletotrichum species interact with chemical herbicides. The fact that Colletotrichum sp. can enhance the pathogenicity of the fungus D. helianthi on the leaves of A. adenophora [31] provides a potential application for biocontrol of A. adenophora by combination of Colletotrichum with pathogenic fungi, such as D. helianthi. Secondly, because the Colletotrichum genus is one of the most important pathogenic fungi [33], our finding highlighted the high disease risk driven by invasive plants in the invaded ecosystem. Indeed, some of these Colletotrichum species were previously verified to be pathogenic on local native plants [32], as well as on tested fruits, including grape and mango, as the recent research by Zheng et al. [82] also revealed. This result suggested that invasive species can facilitate pathogen emergence and amplification, raising concerns about the movement of pathogens among agricultural, horticultural and wild grasses. For example, in all, 39 C. gloeosporioides and 3 C. acutatum isolates were recovered from diseased strawberry crowns, and 52 C. gloeosporioides and one C. acutatum isolate were recovered from non-cultivated hosts. The results were not inconsistent with the hypothesis that C. gloeosporioides isolates obtained from strawberry and noncultivated hosts adjacent to strawberry fields are from the same population and that noncultivated hosts can serve as potential inoculum sources for Colletotrichum crown rot of strawberry [83]. Finally, 14 novel Colletotrichum species were reported in invasive plants A. adenophora and several surrounding crops in south-western China. This result suggested that exotic plants are sources for novel fungal species, and the fungi associated with invasive plants are worth investigating in the future.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof8020185/s1. Table S1: Previously described Colletotrichum species and New species used in phylogenetic analyses.
Author Contributions
M.Q. and H.Z. (Hanbo Zhang) conceived and designed the study; Z.Y. wrote the manuscript; X.J. and H.Z. (Hua Zheng) conducted the experiments. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation Program of PR China, grant number 31760012 and 31970013.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequence data are available in NCBI GenBank following the accession numbers in the manuscript.
Acknowledgments
We are grateful to two reviewers for critically reviewing the manuscript and for providing helpful suggestions to improve this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hyde, K.D.; Soytong, K. The fungal endophyte dilemma. Fungal Divers. 2008, 33, 163–173. [Google Scholar]
- Saikkonen, K.; Wali, P.; Helander, M.; Faeth, S.H. Evolution of endophyte-plant symbioses. Trends Plant Sci. 2004, 9, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Muller, C.B.; Krauss, J. Symbiosis between grasses and asexual fungal endophytes. Curr. Opin. Plant Biol. 2005, 8, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.E. Understanding the diversity of foliar endophytic fungi: Progress, challenges, and frontiers. Fungal Biol. Rev. 2007, 21, 51–66. [Google Scholar] [CrossRef]
- Christian, N.S.; Sullivan, C.; Visser, N.D.; Clay, K. Plant Host and Geographic Location Drive Endophyte Community Composition in the Face of Perturbation. Microb. Ecol. 2016, 72, 621–632. [Google Scholar] [CrossRef]
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1114. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef] [Green Version]
- Seastedt, T.R.; Pysek, P. Mechanisms of Plant Invasions of North America and European Grasslands. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 133–153. [Google Scholar] [CrossRef] [Green Version]
- Callaway, R.M.; Maron, J.L. What have exotic plant invasions taught us over the past 20 years? Trends Ecol. Evol. 2006, 21, 369–374. [Google Scholar] [CrossRef]
- Vilá, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Putten, V.D.; Klironomos, J.N.; Wardle, D.A. Microbial ecology of biological invasions. Isme J. 2007, 1, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andonian, K.; Hierro, J.L. Species interactions contribute to the success of a global plant invader. Biol. Invasions 2011, 13, 2957–2965. [Google Scholar] [CrossRef] [Green Version]
- Jordan, N.R.; Larson, D.L.; Huerd, S.C. Soil modification by invasive plants: Effects on native and invasive species of mixed-grass prairies. Biol. Invasions 2008, 10, 177–190. [Google Scholar] [CrossRef]
- Wolfe, B.E.; Rodgers, V.L.; Stinson, K.A.; Pringle, A. The invasive plant Alliaria petiolata (garlic mustard) inhibits ectomycorrhizal fungi in its introduced range. J. Ecol. 2008, 96, 777–783. [Google Scholar] [CrossRef]
- Shipunov, A.; Newcombe, G.; Raghavendra, A.K.H.; Anderson, C.L. Hidden diversity of endophytic fungi in an invasive plant. Am. J. Bot. 2008, 95, 1096–1108. [Google Scholar] [CrossRef]
- Carroll, G. Fungal endophytes in stems and leaves—From latent pathogen to mutualistic symbiont. Ecology 1988, 69, 2–9. [Google Scholar] [CrossRef]
- Márquez, S.S.; Bills, G.F.; Herrero, N.; Zabalgogeazcoa, I. Non-Systemic fungal endophytes of grasses. Fungal Ecol. 2012, 5, 289–297. [Google Scholar] [CrossRef]
- Clay, K.; Shearin, Z.; Bourke, K.A.; Bickford, W.A.; Kowalski, K.P. Diversity of fungal endophytes in non-native Phragmites australis in the Great Lakes. Biol. Invasions 2016, 18, 2703–2716. [Google Scholar] [CrossRef]
- Soares, M.A.; Li, H.Y.; Kowalski, K.P.; Bergen, M.; Torres, M.S.; White, J.F. Evaluation of the functional roles of fungal endophytes of Phragmites australis from high saline and low saline habitats. Biol. Invasions 2016, 18, 2689–2702. [Google Scholar] [CrossRef]
- Faeth, S.H. Are endophytic fungi defensive plant mutualists? Oikos 2002, 98, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Molinari, N.; Knight, C. Correlated evolution of defensive and nutritional traits in native and non-native plants. Bot. J. Linn. Soc. 2010, 163, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Aschehoug, E.T.; Callaway, R.M.; Newcombe, G.; Tharayil, N.; Chen, S. Fungal endophyte increases the allelopathic effects of an invasive forb. Oecologia 2014, 175, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Aschehoug, E.T.; Metlen, K.L.; Callaway, R.M.; Newcombe, G. Fungal endophytes directly increase the competitive effects of an invasive forb. Ecology 2012, 93, 3–8. [Google Scholar] [CrossRef]
- Newcombe, G.; Shipunov, A.; Eigenbrode, S.D.; Raghavendra, A.K.H.; Ding, H.; Anderson, C.L.; Menjivar, R.; Crawford, M.; Schwarzländer, M. Endophytes influence protection and growth of an invasive plant. Commun. Integr. Biol. 2009, 2, 29–31. [Google Scholar] [CrossRef] [Green Version]
- Day, N.J.; Dunfield, K.E.; Antunes, P.M. Fungi from a non-native invasive plant increase its growth but have different growth effects on native plants. Biol. Invasions 2016, 18, 231–243. [Google Scholar] [CrossRef]
- Julien, M.H.; Griffiths, M.W. Biological Control of Weeds: A World Catalogue of Agents and Their Target Weeds, 4th ed.; CSIRO Entomology: Clayton, Australia; CABI Publishing: Wallingford, UK, 1998. [Google Scholar]
- Buccellato, L.; Byrne, M.J.; Witkowski, E.T.F. Interactions between a stem gall fly and a leaf-spot pathogen in the biological control of Ageratina adenophora. Biol. Control 2012, 61, 222–229. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Y.Z. Invasion dynamics and potential spread of the invasive alien plant species Ageratina adenophora (Asteraceae) in China. Divers. Distrib. 2006, 12, 397–408. [Google Scholar] [CrossRef]
- Mei, L.; Zhu, M.; Zhang, D.Z.; Wang, Y.Z.; Guo, J.; Zhang, H.B. Geographical and Temporal Changes of Foliar Fungal Endophytes Associated with the Invasive Plant Ageratina adenophora. Microbial Ecol. 2014, 67, 402–409. [Google Scholar] [CrossRef]
- Fang, K.; Chen, L.M.; Zhang, H.B. Evaluation of foliar fungus-mediated interactions with below and aboveground enemies of the invasive plant Ageratina adenophora. Ecol. Evol. 2021, 11, 526–535. [Google Scholar] [CrossRef]
- Chen, L.; Zhou, J.; Zeng, T.; Miao, Y.F.; Mei, L.; Yao, G.B.; Fang, K.; Dong, X.F.; Sha, T.; Yang, M.Z.; et al. Quantifying the sharing of foliar fungal pathogens by the invasive plant Ageratina adenophora and its neighbours. New Phytol. 2020, 227, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Cannon, P.F.; Simmons, C.M. Diversity and host preference of leaf endophytic fungi in the Iwokrama Forest Reserve, Guyana. Mycologia 2002, 94, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum—Current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, R.; Kan, J.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef]
- Hyde, K.D.; Cai, L.; McKenzie, E.H.C.; Yang, Y.L.; Zhang, J.Z.; Prihastuti, H. Colletotrichum: A catalogue of confusion. Fungal Divers. 2009, 39, 1–17. [Google Scholar]
- Huang, F.; Chen, G.Q.; Hou, X.; Fu, Y.S.; Cai, L.; Hyde, K.D.; Li, H.Y. Colletotrichum species associated with cultivated citrus in China. Fungal Divers. 2013, 61, 61–74. [Google Scholar] [CrossRef]
- Freeman, S.; Katan, T.; Shabi, T. Characterization of Colletotrichum species responsible for anthracnose diseases of various fruits. Plant Dis. 1998, 82, 596–605. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.E.; Mejía, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef] [Green Version]
- Herre, E.A.; Mejía, L.C.; Kyllo, D.A.; Rojas, E.; Maynard, Z.; Butler, A.; Van Bael, S.A. Ecological implications of anti-pathogen effects of tropical fungal endophytes and mycorrhizae. Ecology 2007, 88, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Tao, G.; Liu, Z.Y.; Liu, F.; Gao, Y.H.; Cai, L. Endophytic Colletotrichum species from Bletilla ochracea (Orchidaceae), with descriptions of seven new speices. Fungal Divers. 2013, 61, 139–164. [Google Scholar] [CrossRef]
- Higgins, K.L.; Coley, P.D.; Kursar, T.A.; Arnold, A.E. Culturing and direct PCR suggest prevalent host generalism among diverse fungal endophytes of tropical forest grasses. Mycologia 2011, 103, 247–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, G.; Cannon, P.F.; Reid, A.; Simmons, C.M. Diversity and molecular relationships of endophytic Colletotrichum isolates from the Iwokrama Forest Reserve, Gruyana. Mycol. Res. 2004, 108, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Rojas, E.I.; Rehner, S.A.; Samuels, G.J.; Van Bael, S.A.; Herre, E.A.; Cannon, P.; Chen, P.; Pang, J.F.; Wang, R.W.; Zhang, Y.P.; et al. Colletotrichum gloeosporioides s.l. associated with Theobroma cacao and other plants in Panama: Multilocus phylogenies distinguish host-associated pathogens from asymptomatic endophytes. Mycologia 2010, 102, 1318–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Hyde, K.D.; Taylor, P.W.J.; Weir, B.S.; Waller, J.M.; Abang, M.M.; Zhang, J.Z.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y.; et al. A polyphasic approach for studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Niu, X.P.; Gao, H.; Qi, J.M.; Chen, M.C.; Tao, A.F.; Xu, J.T.; Dai, Z.G.; Su, J.G. Colletotrichum species associated with jute (Corchorus capsularis L.) anthracnose in southeastern China. Sci. Rep. 2016, 6, 25179. [Google Scholar] [CrossRef] [PubMed]
- Fang, K.; Zhou, J.; Chen, L.; Li, Y.X.; Yang, A.L.; Dong, X.F.; Zhang, H.B. Virulence and community dynamics of fungal species with vertical and horizontal transmission on a plant with multiple infections. PLoS Pathog. 2021, 17, e1009769. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhuang, W.Y. Designing primer sets for amplification of partial calmodulin genes from Penicillia. Mygosystema 2004, 23, 466–473. [Google Scholar] [CrossRef]
- White, T.J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Templeton, M.D.; Rikkerink, E.H.A.; Solon, S.L.; Crowhurst, R.N. Cloning and molecular characterization of the glyceraldehyde-3-phosphate dehydrogenase-encoding gene and cDNA from the plant pathogenic fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Lousie, G.N.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from Filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Zhuang, W.Y. Discovery from a large-scaled survey of Trichoderma in soil of China. Sci. Rep. 2017, 7, 9090. [Google Scholar] [CrossRef] [Green Version]
- Jayawardena, R.S.; Hyde, K.D.; Chen, Y.J.; Papp, V.; Palla, B.; Papp, D.; Bhunjun, C.S.; Hurdeal, V.G.; Senwanna, C.; Manawasinghe, I.S.; et al. One stop shop IV: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 76-100 (2020). Fungal Divers. 2020, 103, 87–218. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Bhunjun, C.S.; Hyde, K.D.; Gentekaki, E.; Itthayakorn, P. Colletotrichum: Lifestyles, biology, morpho-species, species complexes and accepted species. Mycosphere 2021, 12, 519–669. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Page, R.D.M. TreeView: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin-Felix, Y.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Marincowitz, S.; Barnes, I.; Bensch, K.; Braun, U.; Camporesi, E.; Damm, U.; et al. Genera of phytopathogenic fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Weir, B.S.; Damm, U.; Crous, P.W.; Wang, Y.; Liu, B.; Wang, M.; Zhang, M.; Cai, L. Unravelling Colletotrichum species associated with Camellia: Employing ApMat and GS loci to resolve species in the C. gloeosporioides complex. Persoonia 2015, 35, 63–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [Green Version]
- Cannon, P.F.; Buddie, A.G.; Bridge, P.D. The typification of Colletotrichum gloeosporioides. Mycotaxon 2008, 104, 189–204. [Google Scholar]
- Tibpromma, S.; Hyde, K.D.; Bhat, J.D.; Mortimer, P.E.; Xu, J.C.; Promputtha, I.; Doilom, M.; Yang, J.B.; Tang, A.M.C.; Karunarathna, S.C. Identification of endophytic fungi from leaves of Pandanaceae based on their morphotypes and DNA sequence data from southern Thailand. Mycokeys 2018, 33, 25–67. [Google Scholar] [CrossRef]
- Zhou, S.X.; Qiao, L.J.; Jayawardena, R.S.; Hyde, K.D.; Ma, X.Y.; Wen, T.C.; Kang, J.C. Two new endophytic Colletotrichum species from nothapodytespittosporoides in China. MycoKeys 2019, 49, 1–14. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [Green Version]
- Uematsu, S.; Kageyama, K.; Moriwaki, J.; Sato, T. Colletotrichum carthami comb. nov., an anthracnose pathogen of safflower, garland chrysanthemum and pot marigold, revived by molecular phylogeny with authentic herbarium specimens. J. Gen. Plant Pathol. 2012, 78, 316–330. [Google Scholar] [CrossRef]
- Wang, Q.T.; Liu, X.T.; Ma, H.Y.; Shen, X.Y.; Hou, C.L. Colletotrichum yulongense sp. nov. and C. rhombiforme isolated as endophytes from vaccinium dunalianum var. urophyllum in China. Phytotaxa 2019, 394, 285. [Google Scholar] [CrossRef]
- Vila, M.; Basnou, C.; Pysek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W.; Olenin, S.; Roques, A.; Roy, D.; et al. How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front. Ecol. Environ. 2010, 8, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Turbelin, A.J.; Malamud, B.D.; Francis, R.A. Mapping the global state of invasive alien species: Patterns of invasion and policy responses. Glob. Ecol. Biogeogr. 2017, 26, 78–92. [Google Scholar] [CrossRef] [Green Version]
- Bufford, J.L.; Hulme, P.E.; Sikes, B.A.; Cooper, J.A.; Johnston, P.R.; Duncan, R.P. Taxonomic similarity, more than contact opportunity, explains novel plant-pathogen associations between native and alien taxa. New Phytol. 2016, 212, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Dickie, I.A.; Bufford, J.L.; Cobb, R.C.; Desprez-Loustau, M.L.; Grelet, G.; Hulme, P.E.; Klironomos, J.; Makiola, A.; Nunez, M.A.; Pringle, A.; et al. The emerging science of linked plant–fungal invasions. New Phytol. 2017, 215, 1314–1332. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, C.E.; Blumenthal, D.; Jarosik, V.; Puckett, E.E.; Pysek, P. Controls on pathogen species richness in plants’ introduced and native ranges: Roles of residence time, range size and host traits. Ecol. Lett. 2010, 13, 1525–1535. [Google Scholar] [CrossRef]
- Stricker, K.B.; Harmon, P.F.; Goss, E.M.; Clay, K.; Flory, S.L. Emergence and accumulation of novel pathogens suppress aninvasive species. Ecol. Lett. 2016, 19, 469–477. [Google Scholar] [CrossRef]
- Mangla, S.; Inderjit Callaway, R.M. Exotic invasive plant accumulates native soil pathogens which inhibit native plants. J. Ecol. 2008, 96, 58–67. [Google Scholar] [CrossRef]
- Crocker, E.V.; Karp, M.A.; Nelson, E.B. Virulence of oomycete pathogens from Phragmites australis—Invaded and noninvaded soils to seedlings of wetland plant species. Ecol. Evol. 2015, 5, 2127–2139. [Google Scholar] [CrossRef]
- Rudgers, J.A.; Mattingly, W.B.; Koslow, J.M. Mutualistic fungus promotes plant invasion into diverse communities. Oecologia 2005, 144, 463–471. [Google Scholar] [CrossRef]
- Zheng, H.; Yu, Z.; Jiang, X.; Fang, L.; Qiao, M. Endophytic Colletotrichum Species from Aquatic Plants in Southwest China. J. Fungi 2022, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.L.; Mackenzie, S.J.; Legard, D.E. Genetic and pathogenic analyses of Colletotrichum gloeosporioides isolates from strawberry and noncultivated hosts. Phytopathology 2004, 94, 446–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Phylogenic tree generated by the maximum likelihood analysis using combined sequences of ACT, CHS-1, GAPDH, ITS, and TUB2 loci of the genus Colletotrichum. Bootstrap values ≥ 70 (left) and Bayesian posterior probability values ≥ 0.9 (right) are indicated at nodes (MLBP/BIBP). Monilochaetes infuscans CBS 869.96 was used as the outgroup. Novel species generated in this study are indicated in black bold.
Figure 1.
Phylogenic tree generated by the maximum likelihood analysis using combined sequences of ACT, CHS-1, GAPDH, ITS, and TUB2 loci of the genus Colletotrichum. Bootstrap values ≥ 70 (left) and Bayesian posterior probability values ≥ 0.9 (right) are indicated at nodes (MLBP/BIBP). Monilochaetes infuscans CBS 869.96 was used as the outgroup. Novel species generated in this study are indicated in black bold.




Figure 2.
Colletotrichum adenophorae (YMF 1.04952). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E,F) Setae. (G) Conidiophores and Conidia. (H) Appressoria. (I) Conidiophores. (J) Conidia. Scale bars: (D) = 100 μm, (E) = 10 μm, (F) = 50 μm, (G–J) = 10 μm.
Figure 2.
Colletotrichum adenophorae (YMF 1.04952). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E,F) Setae. (G) Conidiophores and Conidia. (H) Appressoria. (I) Conidiophores. (J) Conidia. Scale bars: (D) = 100 μm, (E) = 10 μm, (F) = 50 μm, (G–J) = 10 μm.

Figure 3.
Colletotrichum analogum (YMF 1.06943). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Chlamydospores. (F,G) Appressoria. (H) Conidiophores. (I) Conidia. Scale bars: (D) = 100 μm, (E–I) = 10 μm.
Figure 3.
Colletotrichum analogum (YMF 1.06943). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Chlamydospores. (F,G) Appressoria. (H) Conidiophores. (I) Conidia. Scale bars: (D) = 100 μm, (E–I) = 10 μm.

Figure 4.
Colletotrichum cangyuanense (YMF 1.05001). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Setae. (F) Appressoria. (G) Conidiophores. (H) Conidiophores and Conidia. (I) Conidia. Scale bars: (D) = 100 μm, (E–I) = 10 μm.
Figure 4.
Colletotrichum cangyuanense (YMF 1.05001). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Setae. (F) Appressoria. (G) Conidiophores. (H) Conidiophores and Conidia. (I) Conidia. Scale bars: (D) = 100 μm, (E–I) = 10 μm.

Figure 5.
Colletotrichum dimorphum (YMF 1.07309). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Appressoria. (E) Conidiophores. (F) Conidiophores and Conidia. (G) Conidia. Scale bars: (D–G) = 10 μm.
Figure 5.
Colletotrichum dimorphum (YMF 1.07309). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Appressoria. (E) Conidiophores. (F) Conidiophores and Conidia. (G) Conidia. Scale bars: (D–G) = 10 μm.

Figure 6.
Colletotrichum gracile (YMF1.06939). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Ascomata. (E) Outer surface of peridium. (F,G) Asci. (H) Ascospores. (I) Conidiophores and Conidia. (J) Appressoria. (K) Conidiophores. (L) Conidia. Scale bars: (D) = 100 μm, (E–K) = 10 μm.
Figure 6.
Colletotrichum gracile (YMF1.06939). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Ascomata. (E) Outer surface of peridium. (F,G) Asci. (H) Ascospores. (I) Conidiophores and Conidia. (J) Appressoria. (K) Conidiophores. (L) Conidia. Scale bars: (D) = 100 μm, (E–K) = 10 μm.

Figure 7.
Colletotrichum nanhuaense (YMF1.04993). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Seta. (F) Appressoria. (G) Conidiophores. (H) Conidiophores and Conidia. (I) Conidia. Scale bars: (D) = 100 μm, (E–I) = 10 μm.
Figure 7.
Colletotrichum nanhuaense (YMF1.04993). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Seta. (F) Appressoria. (G) Conidiophores. (H) Conidiophores and Conidia. (I) Conidia. Scale bars: (D) = 100 μm, (E–I) = 10 μm.

Figure 8.
Colletotrichum nullisetosum (YMF1.06946). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Appressoria. (F,I) Conidiophores. (G) Conidiophores and Conidia. (H,J) Conidia. Scale bars: (D) = 100 μm, (E–J) = 10 μm.
Figure 8.
Colletotrichum nullisetosum (YMF1.06946). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Appressoria. (F,I) Conidiophores. (G) Conidiophores and Conidia. (H,J) Conidia. Scale bars: (D) = 100 μm, (E–J) = 10 μm.

Figure 9.
Colletotrichum oblongisporum (YMF 1.06938). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Appressoria. (E) Conidiophores. (F) Conidia. Scale bars: (D–F) = 10 μm.
Figure 9.
Colletotrichum oblongisporum (YMF 1.06938). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Appressoria. (E) Conidiophores. (F) Conidia. Scale bars: (D–F) = 10 μm.

Figure 10.
Colletotrichum parvisporum (YMF1.06942). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Appressoria. (E) Conidiophores. (F) Conidia. Scale bars: (D–F) = 10 μm.
Figure 10.
Colletotrichum parvisporum (YMF1.06942). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Appressoria. (E) Conidiophores. (F) Conidia. Scale bars: (D–F) = 10 μm.

Figure 11.
Colletotrichum robustum (YMF1.06941). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E,F) Setae. (G) Conidiophores and Conidia. (H) Appressoria. (I) Conidiophores. (J) Conidia. Scale bars: (D) = 100 μm, (F) = 20 μm, (E,G–J) = 10 μm.
Figure 11.
Colletotrichum robustum (YMF1.06941). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E,F) Setae. (G) Conidiophores and Conidia. (H) Appressoria. (I) Conidiophores. (J) Conidia. Scale bars: (D) = 100 μm, (F) = 20 μm, (E,G–J) = 10 μm.

Figure 12.
Colletotrichum simulanticitri (YMF1.07302). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Setae. (F,H) Appressoria. (G) Appressoria and Conidia. (I) Conidiophores. (J) Conidia. Scale bars: (D) = 100 μm, (E–J) = 10 μm.
Figure 12.
Colletotrichum simulanticitri (YMF1.07302). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Setae. (F,H) Appressoria. (G) Appressoria and Conidia. (I) Conidiophores. (J) Conidia. Scale bars: (D) = 100 μm, (E–J) = 10 μm.

Figure 13.
Colletotrichum speciosum (YMF1.07301). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D,E) Setae. (F) Appressoria. (G) Conidiophores. (H) Conidiophores and Conidia. (I) Conidia. Scale bars: (D–I) = 10 μm.
Figure 13.
Colletotrichum speciosum (YMF1.07301). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D,E) Setae. (F) Appressoria. (G) Conidiophores. (H) Conidiophores and Conidia. (I) Conidia. Scale bars: (D–I) = 10 μm.

Figure 14.
Colletotrichum subhenanense (YMF1.06865). (A–C) Colonies on media above and below after 7 d at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Appressoria. (E) Conidiophores. (F) Conidia. Scale bars: (D–F) = 10 μm.
Figure 14.
Colletotrichum subhenanense (YMF1.06865). (A–C) Colonies on media above and below after 7 d at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Appressoria. (E) Conidiophores. (F) Conidia. Scale bars: (D–F) = 10 μm.

Figure 15.
Colletotrichum yunajiangense (YMF1.04996). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Setae. (F) Appressoria. (G) Conidiophores. (H) Conidiophores and Conidia. (I) Conidia. Scale bars: (D) = 100 μm, (E–I) = 10 μm.
Figure 15.
Colletotrichum yunajiangense (YMF1.04996). (A–C) Colonies on media above and below after 7 days at 25 °C ((A) MEA; (B) PDA; (C) SNA). (D) Conidiomata. (E) Setae. (F) Appressoria. (G) Conidiophores. (H) Conidiophores and Conidia. (I) Conidia. Scale bars: (D) = 100 μm, (E–I) = 10 μm.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yu, Z.; Jiang, X.; Zheng, H.; Zhang, H.; Qiao, M. Fourteen New Species of Foliar Colletotrichum Associated with the Invasive Plant Ageratinaadenophora and Surrounding Crops. J. Fungi 2022, 8, 185. https://doi.org/10.3390/jof8020185
AMA Style
Yu Z, Jiang X, Zheng H, Zhang H, Qiao M. Fourteen New Species of Foliar Colletotrichum Associated with the Invasive Plant Ageratinaadenophora and Surrounding Crops. Journal of Fungi. 2022; 8(2):185. https://doi.org/10.3390/jof8020185
Chicago/Turabian StyleYu, Zefen, Xinwei Jiang, Hua Zheng, Hanbo Zhang, and Min Qiao. 2022. "Fourteen New Species of Foliar Colletotrichum Associated with the Invasive Plant Ageratinaadenophora and Surrounding Crops" Journal of Fungi 8, no. 2: 185. https://doi.org/10.3390/jof8020185
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.