
Benzoic Acid and Its Hydroxylated Derivatives Suppress Early Blight of Tomato (Alternaria solani) via the Induction of Salicylic Acid Biosynthesis and Enzymatic and Nonenzymatic Antioxidant Defense Machinery

 ,
,  ,
,  ,
,
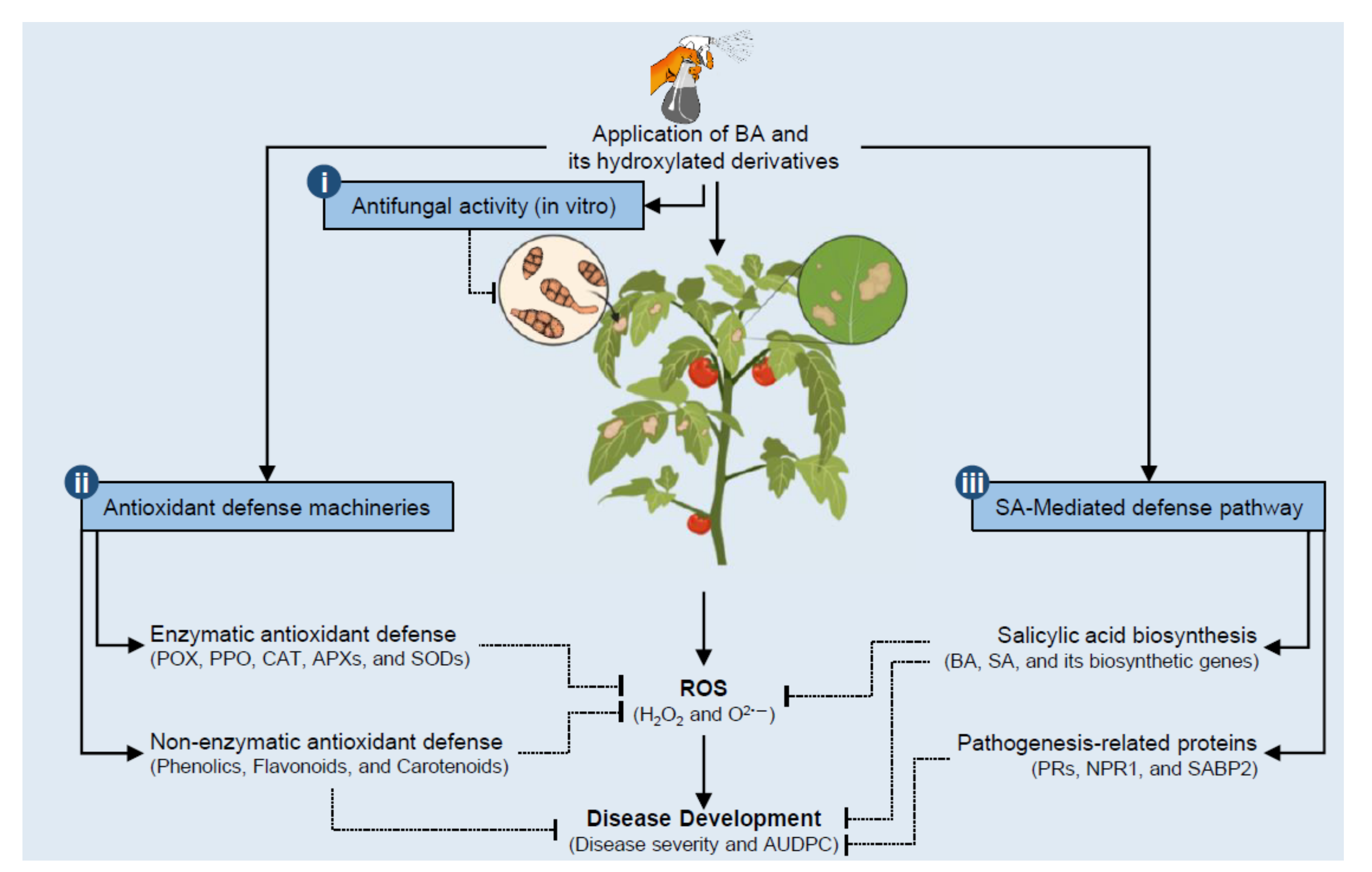
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Antifungal Activity of Benzoic Acid and Its Hydroxylated Derivatives
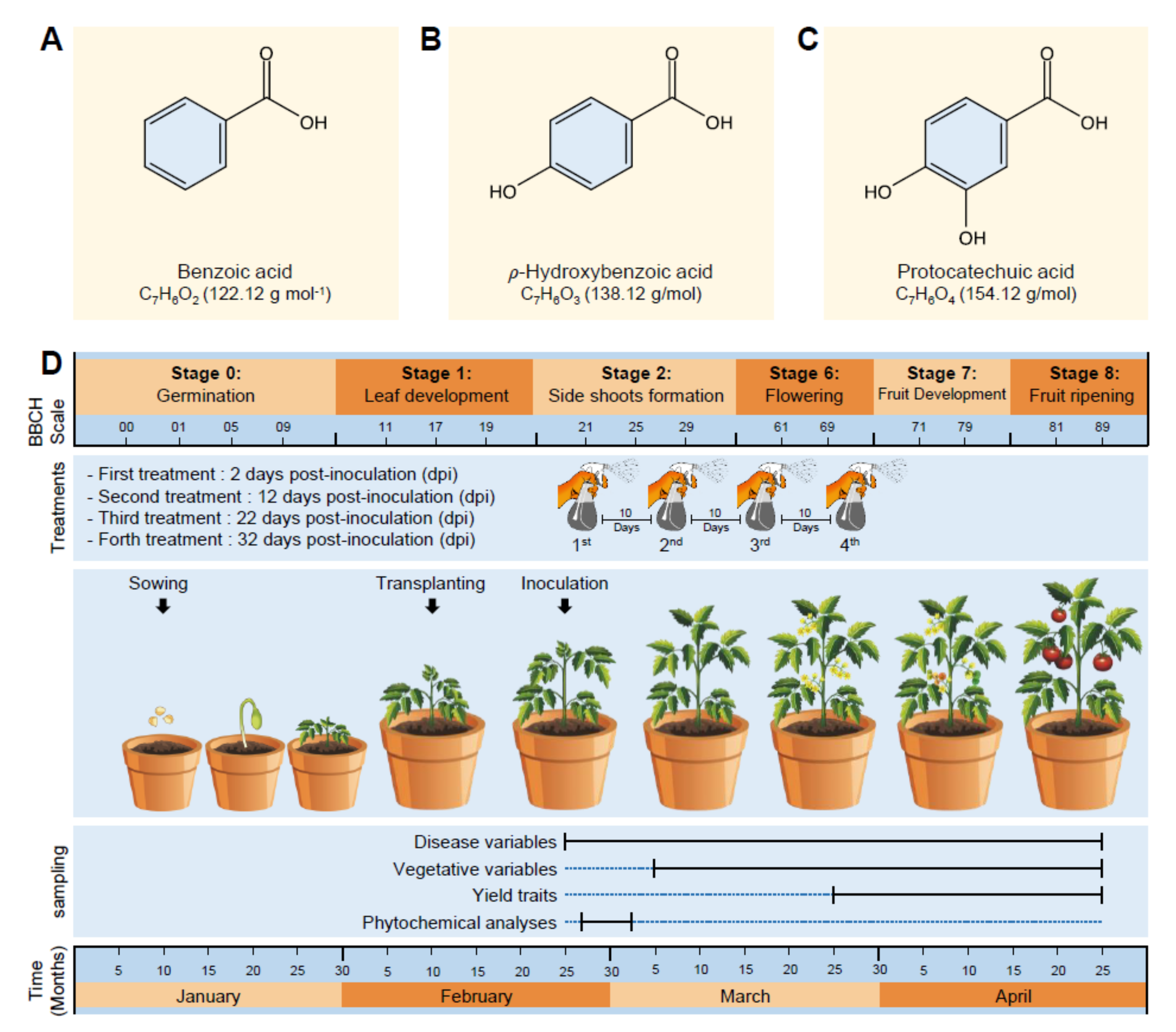
2.1.1. Tested Compounds
2.1.2. Antifungal Activity
2.1.3. Half-Maximal Inhibitory Concentration (IC50)
2.2. In Vivo Experiment
2.2.1. Plant Materials and Growth Conditions
2.2.2. Infection with A. solani and Treatment with Benzoic Acid and Its Derivatives
2.2.3. Treatment with BA, HBA, or PCA
2.2.4. Disease Assessments
2.2.5. Vegetative Growth and Yield Assessment
2.3. Phytochemical Analyses
2.3.1. Photosynthetic Pigments
2.3.2. Total Soluble Phenolic Compounds
2.3.3. Total Soluble Flavonoids
2.3.4. In Situ Histochemical Localization of O2•− and H2O2
2.3.5. Antioxidant Enzymes Activity
2.4. Quantification of Salicylic Acid and Its Precursors Using GC-MS
2.5. Gene Expression Analysis
2.6. Statistical Analysis
3. Results
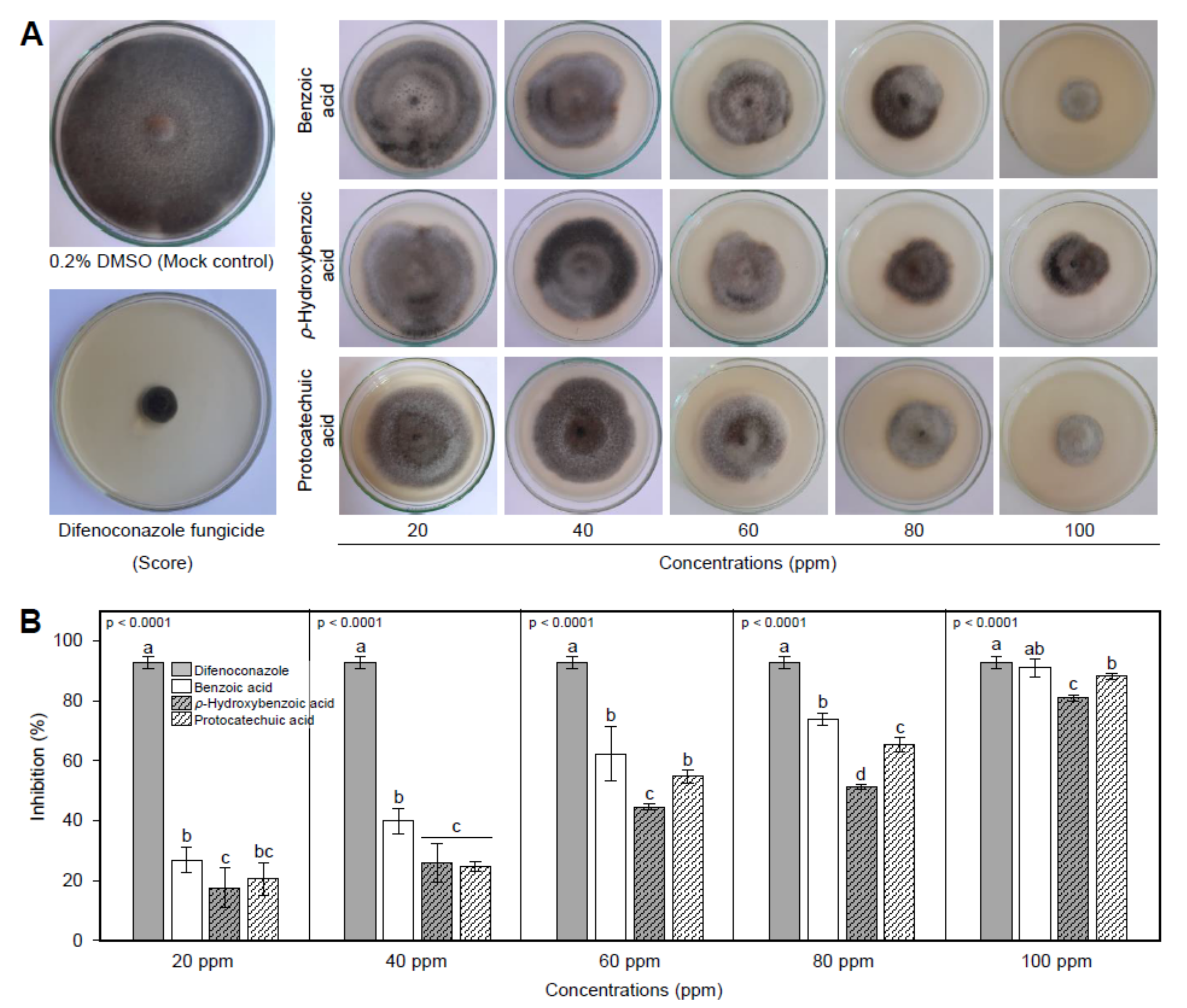
3.1. In Vitro Antifungal Activity of Benzoic Acid and Its Hydroxylated Derivatives
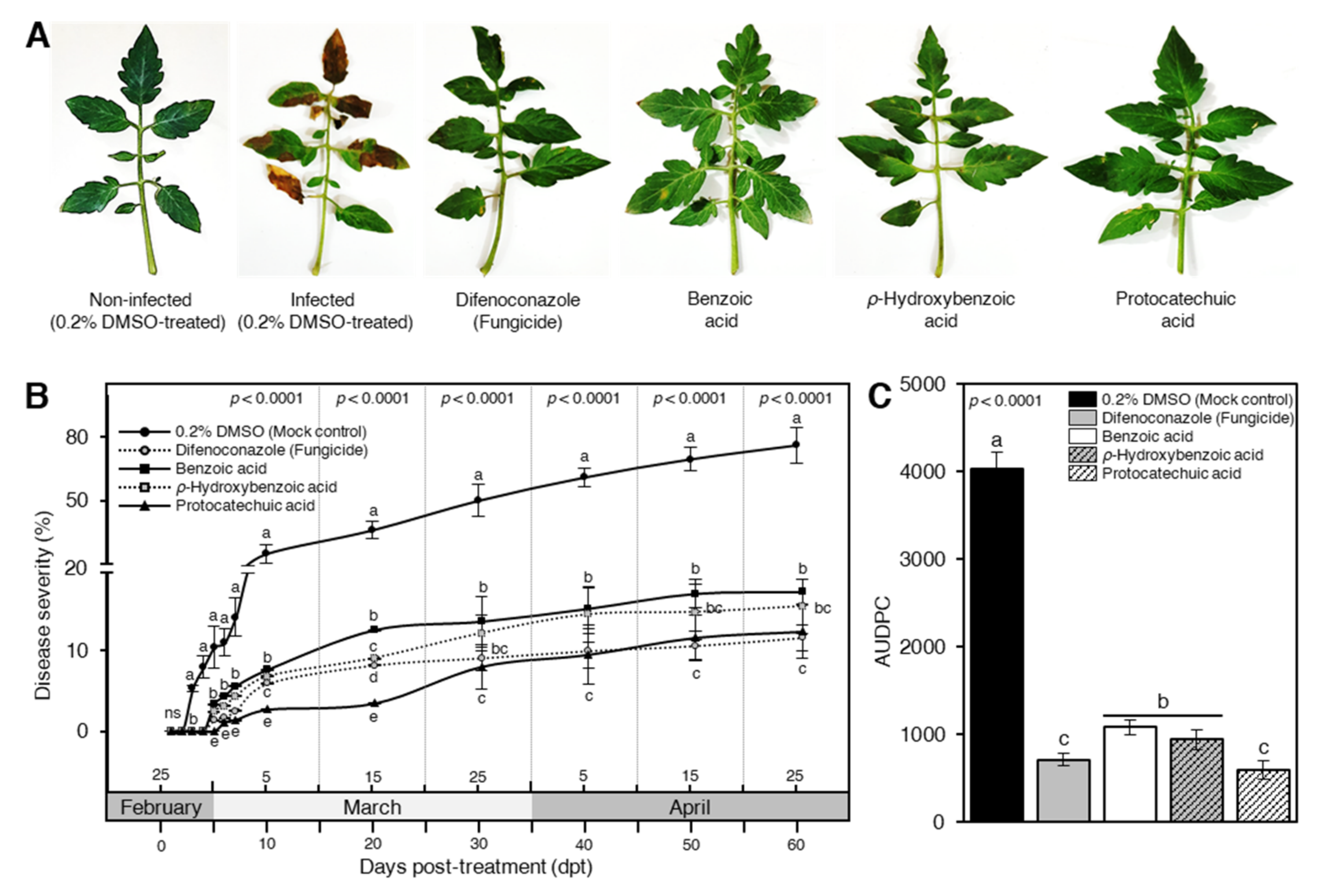
3.2. Benzoic Acid and Its Derivatives Reduced the Disease Evaluation of Early Blight
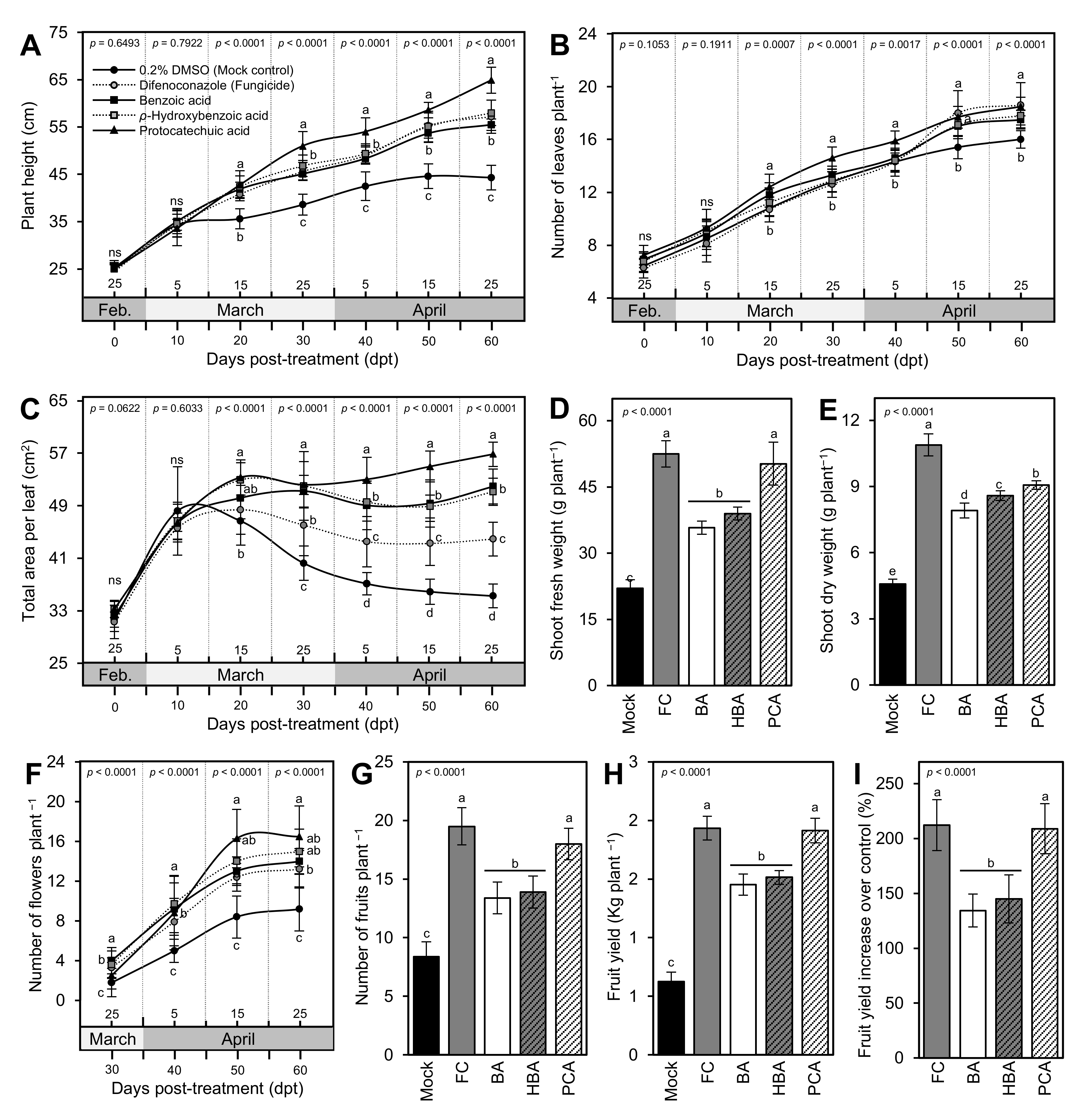
3.3. BA and Its Hydroxylated Derivatives Improve the Growth Variables of A. solani-Infected Tomato Plants
3.4. BA and Its Derivatives Enhance Fruit Yield and Its Components of Infected Tomato Plants
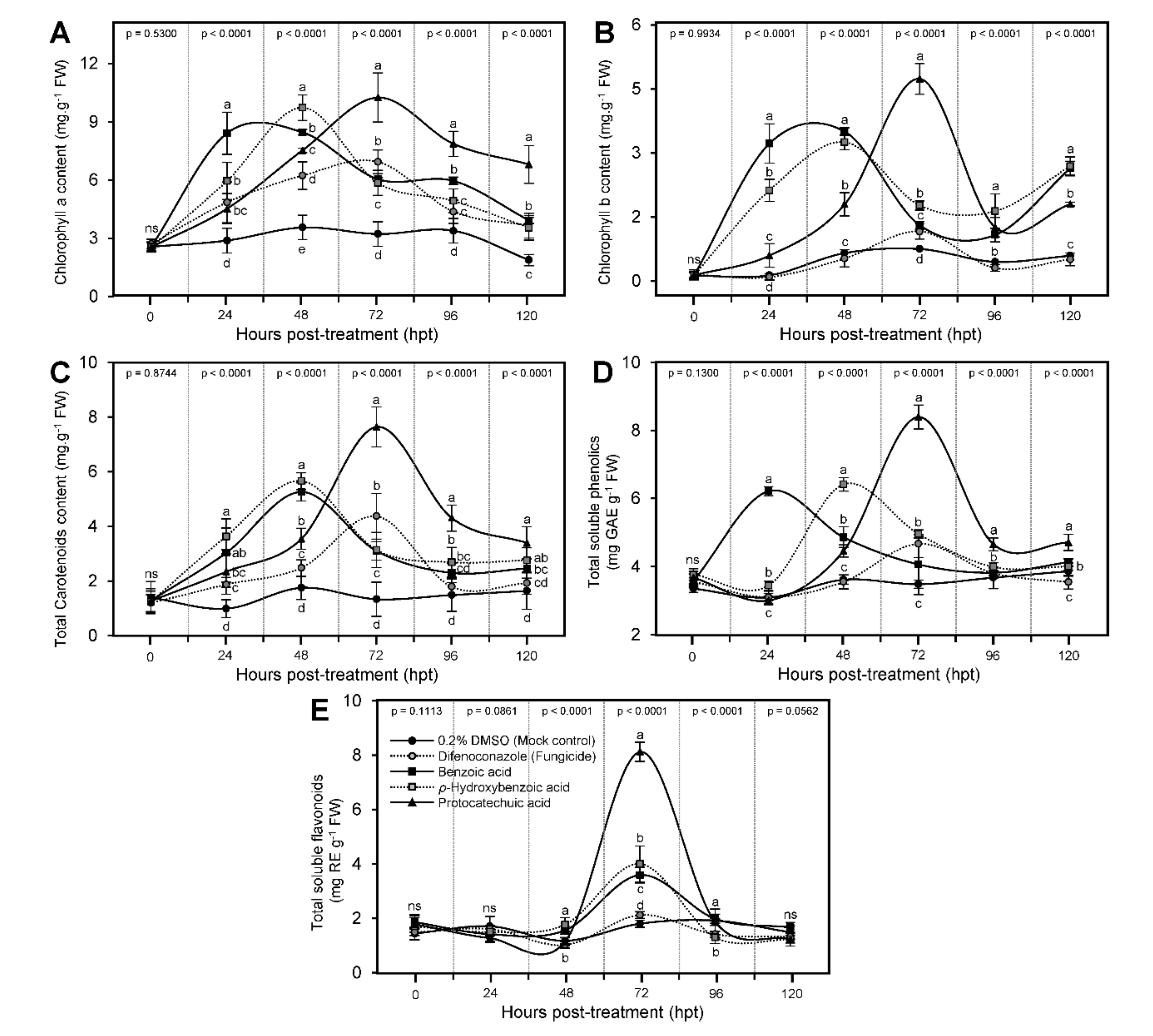
3.5. Effect of BA and Its Derivatives on Photosynthetic Pigments of Infected Leaves
3.6. Benzoic Acid and Its Derivatives Enhanced the Profile of Total Soluble Phenolics and Flavonoids of A. solani-Infected Tomato Plants
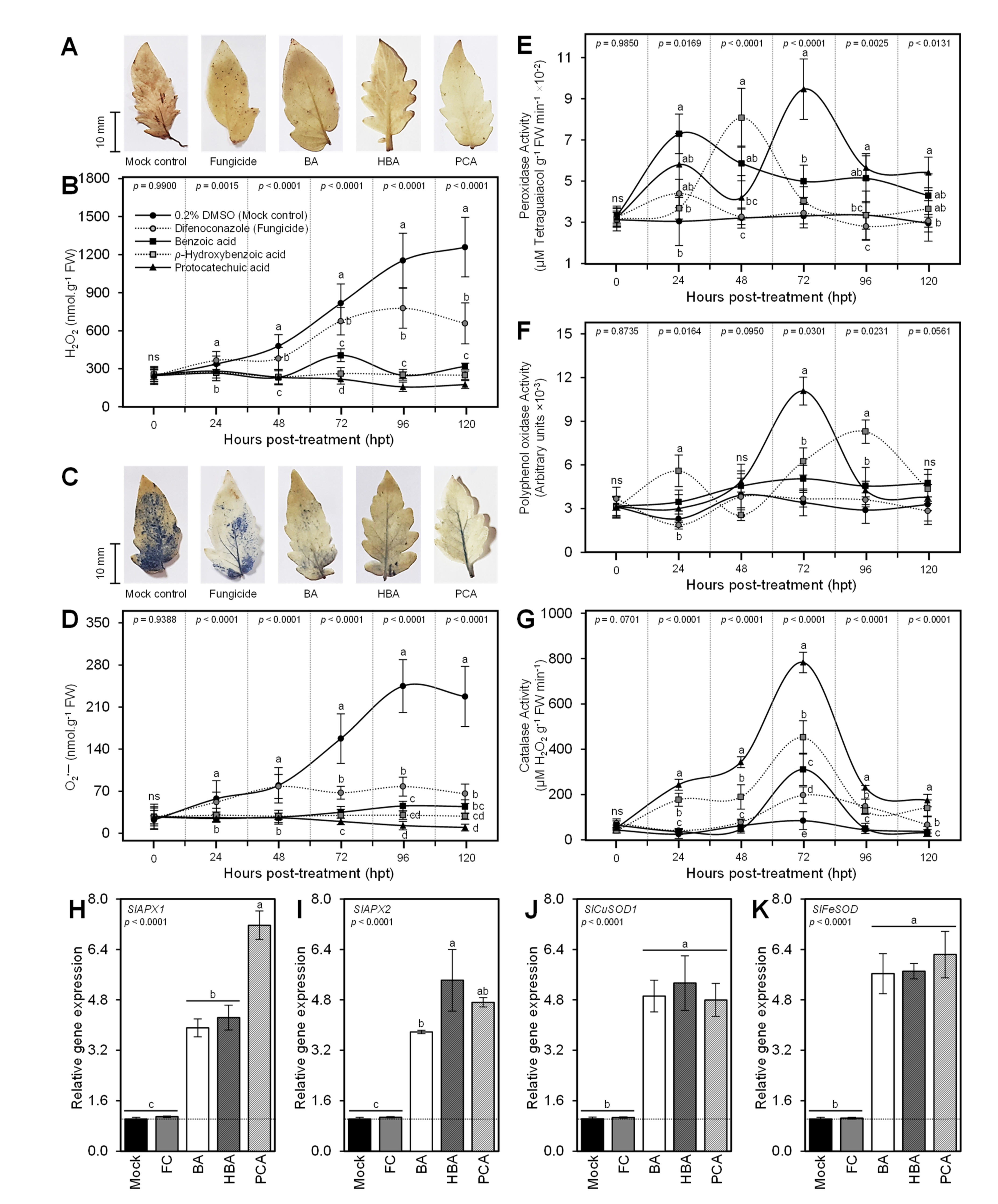
3.7. BA and Its Derivatives Alleviate the Oxidative Stress of A. solani-Infected Leaves
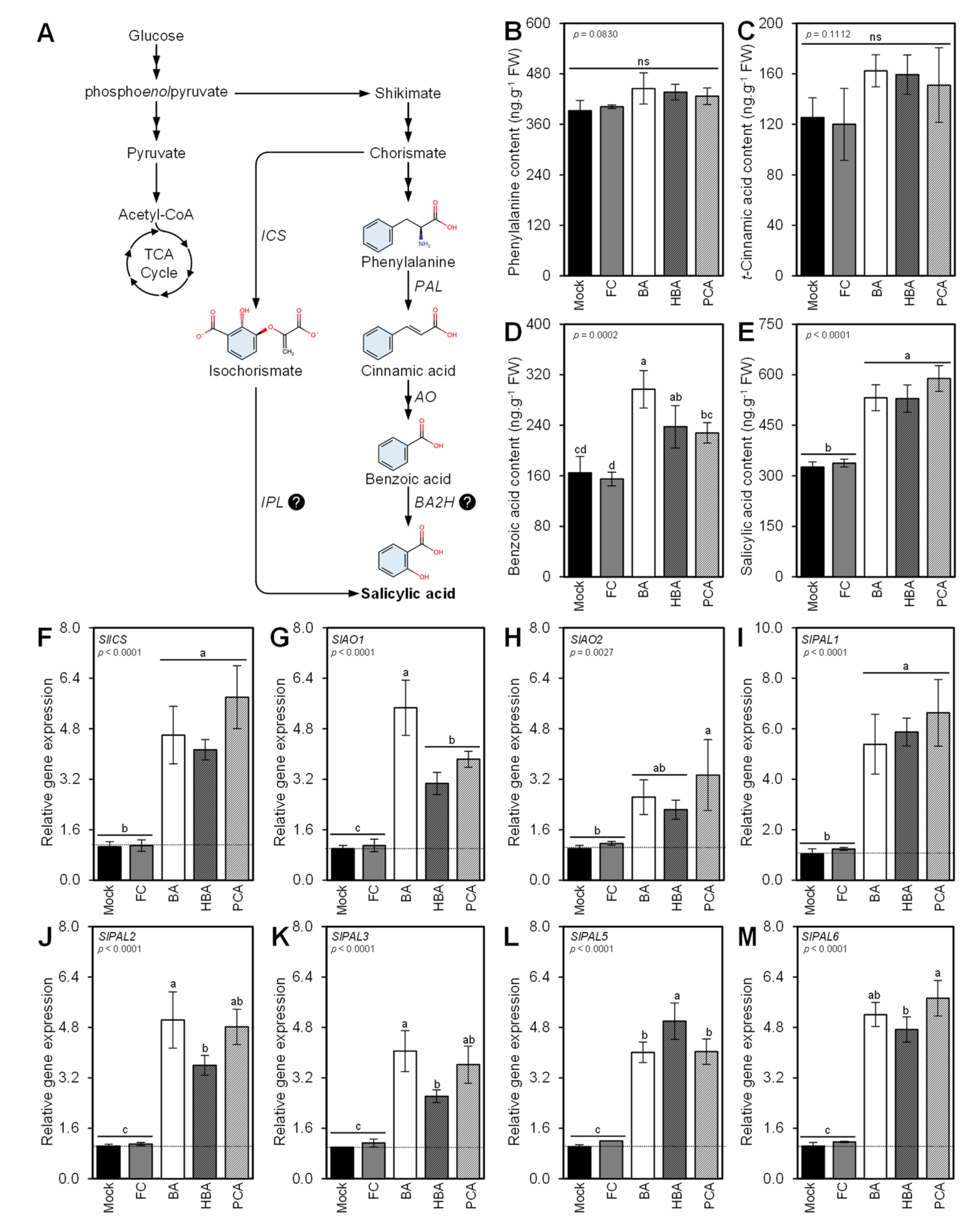
3.8. Benzoic Acid and Its Derivatives Induce the Salicylic Acid Biosynthesis
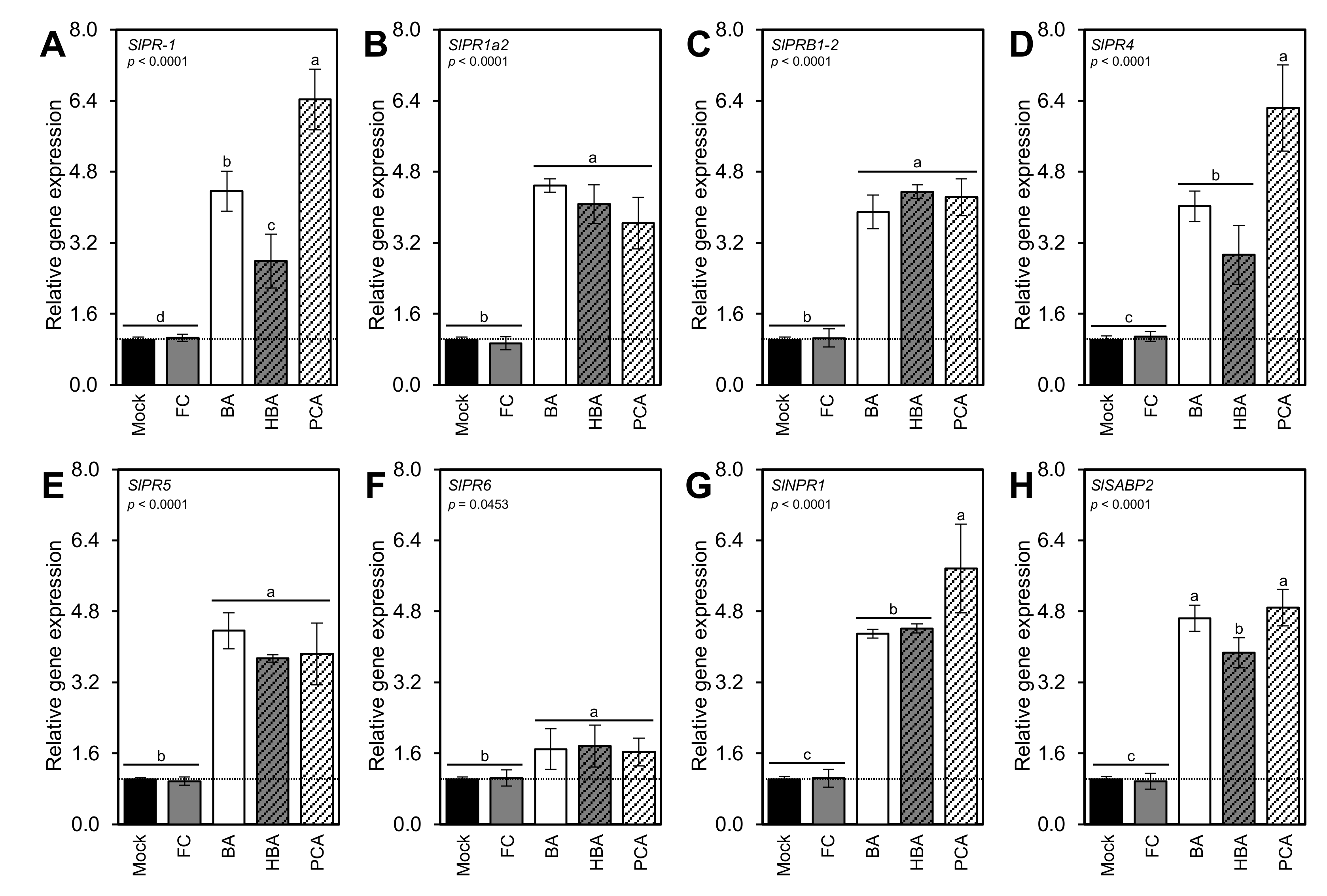
3.9. BA and Its Derivatives Stimulate the Expression of Pathogenesis-Related Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kimura, S.; Sinha, N. Tomato (Solanum lycopersicum): A model fruit-bearing crop. Cold Spring Harb. Protoc. 2008, 3. [Google Scholar] [CrossRef]
- FAOSTAT, Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 13 February 2020).
- Abada, K.A.; Mostafa, S.H.; Mervat, R.; Abada, K.A.; Mostafa, S.H.; Mervat, R. Effect of some chemical salts on suppressing the infection by early blight disease of tomato. Egypt. J. Appl. Sci. 2008, 23, 47–58. [Google Scholar]
- Chao, J.; Huo, T.I.; Cheng, H.Y.; Tsai, J.C.; Liao, J.W.; Lee, M.S.; Qin, X.M.; Hsieh, M.T.; Pao, L.H.; Peng, W.H. Gallic acid ameliorated impaired glucose and lipid homeostasis in high fat diet-induced NAFLD mice. PLoS ONE 2014, 9, e96969. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Kaur, T.; Yadav, A.N.; Sharma, S.; Singh, N. Diversity of fungal isolates associated with early blight disease of tomato from mid Himalayan region of India. Arch. Phytopathol. Plant Prot. 2020, 53, 612–624. [Google Scholar] [CrossRef]
- Pandey, S.; Giri, V.P.; Tripathi, A.; Kumari, M.; Narayan, S.; Bhattacharya, A.; Srivastava, S.; Mishra, A. Early blight disease management by herbal nanoemulsion in Solanum lycopersicum with bio-protective manner. Ind. Crops Prod. 2020, 150, 112421. [Google Scholar] [CrossRef]
- Meno, L.; Escuredo, O.; Rodríguez-Flores, M.S.; Seijo, M.C. Modification of the TOMCAST Model with Aerobiological Data for Management of Potato Early Blight. Agronomy 2020, 10, 1872. [Google Scholar] [CrossRef]
- Van der Waals, J.E.; Korsten, L.; Aveling, T.A.S. A review of early blight of potato. Afr. Plant Prot. 2001, 7, 91–102. [Google Scholar]
- Sahu, D.K.; Khare, C.P.; Singh, H.K.; Patel, R.; Thakur, M.P. Epidemiological Studies on Early Blight Disease of Tomato. Bioscan 2014, 9, 1345–1350. [Google Scholar]
- Attia, M.S.; El-Sayyad, G.S.; Abd Elkodous, M.; El-Batal, A.I. The effective antagonistic potential of plant growth-promoting rhizobacteria against Alternaria solani-causing early blight disease in tomato plant. Sci. Hortic. 2020, 266, 109289. [Google Scholar] [CrossRef]
- Agamy, R.; Alamri, S.; Moustafa, M.F.M.; Hashem, M. Management of tomato leaf spot caused by Alternaria tenuissima wiltshire using salicylic acid and agrileen. Int. J. Agric. Biol. 2013, 15, 266–272. [Google Scholar]
- De La Noval, B.; Pérez, E.; Martínez, B.; León, O.; Martínez-Gallardo, N.; Délano-Frier, J. Exogenous systemin has a contrasting effect on disease resistance in mycorrhizal tomato (Solanum lycopersicum) plants infected with necrotrophic or hemibiotrophic pathogens. Mycorrhiza 2007, 17, 449–460. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, W.; Wang, X.; Yang, L.; Han, S.; Chen, S.; Strasser, R.J.; Valverde, B.E.; Qiang, S. Comparative phytotoxicity of usnic acid, salicylic acid, cinnamic acid and benzoic acid on photosynthetic apparatus of Chlamydomonas reinhardtii. Plant Physiol. Biochem. 2018, 128, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dar, S.A.; Rather, B.A.; Wani, A.R.; Ganie, M.A. Chemical Science Review and Letters Resistance against Insect Pests by Plant Phenolics and their Derivative Compounds. Chem. Sci. Rev. Lett. 2017, 6, 1941–1949. [Google Scholar]
- De la Rosa, L.A.; Moreno-Escamilla, J.O.; Rodrigo-García, J.; Alvarez-Parrilla, E. Phenolic compounds. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E., Carrillo-Lopez, A., Eds.; Woodhead Publishing, Elsevier: Sawston, UK, 2018; pp. 253–271. ISBN 9780128132784. [Google Scholar]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef] [PubMed]
- Clé, C.; Hill, L.M.; Niggeweg, R.; Martin, C.R.; Guisez, Y.; Prinsen, E.; Jansen, M.A.K. Modulation of chlorogenic acid biosynthesis in Solanum lycopersicum; consequences for phenolic accumulation and UV-tolerance. Phytochemistry 2008, 69, 2149–2156. [Google Scholar] [CrossRef]
- Srinivasan, M.; Sudheer, A.R.; Menon, V.P. Ferulic acid: Therapeutic potential through its antioxidant property. J. Clin. Biochem. Nutr. 2007, 40, 92–100. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Lin, H.; Zhou, J.; Chen, L.; Pan, Z.; Fang, B. Synthesis and antimicrobial activity of the hybrid molecules between amoxicillin and derivatives of benzoic acid. Drug Dev. Res. 2020, 82, 198–206. [Google Scholar] [CrossRef]
- Senaratna, T.; Merritt, D.; Dixon, K.; Bunn, E.; Touchell, D.; Sivasithamparam, K. Benzoic acid may act as the functional group in salicylic acid and derivatives in the induction of multiple stress tolerance in plants. Plant Growth Regul. 2003, 39, 77–81. [Google Scholar] [CrossRef]
- Williams, M.; Senaratna, T.; Dixon, K.; Sivasithamparam, K. Benzoic acid induces tolerance to biotic stress caused by Phytophthora cinnamomi in Banksia attenuata. Plant Growth Regul. 2003, 41, 89–91. [Google Scholar] [CrossRef]
- Amborabé, B.E.; Fleurat-Lessard, P.; Chollet, J.F.; Roblin, G. Antifungal effects of salicylic acid and other benzoic acid derivatives towards Eutypa lata: Structure-activity relationship. Plant Physiol. Biochem. 2002, 40, 1051–1060. [Google Scholar] [CrossRef]
- Embaby, E.M.; Hazaa, M.; Hagag, L.F.; Ibrahim, T.E.; El-Azem, F.S.A. Decay of some citrus fruit quality caused by fungi and their control: II-control Alternaria rot or core rot decay by using some alternative fungicides. J. Appl. Sci. Res. 2013, 9, 5671–5678. [Google Scholar]
- Singh, H.; Singh, R.; Maurya, K. Management of Alternaria Blight of Indian Mustard through Resistance Inducing Chemicals. J. AgriSearch 2014, 1, 108–111. [Google Scholar]
- Liu, J.; Li, X.; Jia, Z.; Zhang, T.; Wang, X. Effect of benzoic acid on soil microbial communities associated with soilborne peanut diseases. Appl. Soil Ecol. 2017, 110, 34–42. [Google Scholar] [CrossRef]
- Heflish, A.; El samra, I.; Youssef, N. Occurrence and Control of Alternaria alternata, Penicilliun citrinum and Aspergillus flavus Mycotoxins in Broad Bean Seeds by Benzoic and Sorbic Acids. Egypt. Acad. J. Biol. Sci. G Microbiol. 2020, 12, 75–87. [Google Scholar] [CrossRef]
- Cheng, H.M.; Gao, X.W.; Zhang, K.; Wang, X.R.; Zhou, W.; Li, S.J.; Cao, X.L.; Yan, D.P. A novel antimicrobial composite: ZnAl-hydrotalcite with: P-hydroxybenzoic acid intercalation and its possible application as a food packaging material. New J. Chem. 2019, 43, 19408–19414. [Google Scholar] [CrossRef]
- Krzysztoforska, K.; Mirowska-Guzel, D.; Widy-Tyszkiewicz, E. Pharmacological effects of protocatechuic acid and its therapeutic potential in neurodegenerative diseases: Review on the basis of in vitro and in vivo studies in rodents and humans. Nutr. Neurosci. 2019, 22, 72–82. [Google Scholar] [CrossRef]
- Zhang, J.; Li, D.M.; Sun, W.J.; Wang, X.J.; Bai, J.G. Exogenous p-hydroxybenzoic acid regulates antioxidant enzyme activity and mitigates heat stress of cucumber leaves. Sci. Hortic. 2012, 148, 235–245. [Google Scholar] [CrossRef]
- Nguyen, X.H.; Naing, K.W.; Lee, Y.S.; Moon, J.H.; Lee, J.H.; Kim, K.Y. Isolation and characteristics of protocatechuic acid from Paenibacillus elgii HOA73 against Botrytis cinerea on strawberry fruits. J. Basic Microbiol. 2015, 55, 625–634. [Google Scholar] [CrossRef]
- Feller, C.; Bleiholder, H.; Buhr, L.; Hack, H.; He, M.; Klose, R.; Meier, U.; Stau/, R.; Van Den Boom, T.; Weber, E. Phanologische Entwicklungsstadien von Gemusepflanzen II. Fruchtgemuse und Hulsenfruchte. Nachrichtenbl. Deut. Pflanzenschutzd. 1995, 47, 217–232. [Google Scholar]
- Acosta-Quezada, P.G.; Riofrío-Cuenca, T.; Rojas, J.; Vilanova, S.; Plazas, M.; Prohens, J. Phenological growth stages of tree tomato (Solanum betaceum Cav.), an emerging fruit crop, according to the basic and extended BBCH scales. Sci. Hortic. 2016, 199, 216–223. [Google Scholar] [CrossRef]
- Grover, R.; Moore, J.D. Toximetric studies of fungicides against the brown root organisms, Sclerotinia fructicola and S. laxa. Phytopathology 1962, 52, 876–880. [Google Scholar]
- Prentice, R.L. A Generalization of the Probit and Logit Methods for Dose Response Curves. Biometrics 1976, 32, 761. [Google Scholar] [CrossRef]
- Pandey, K.K.; Pandey, P.K.; Kalloo, G.; Banerjee, M.K. Resistance to early blight of tomato with respect to various parameters of disease epidemics. J. Gen. Plant Pathol. 2003, 69, 364–371. [Google Scholar] [CrossRef]
- Shaner, G.; Finney, R.E. The Effect of Nitrogen Fertilization on the Expression of Slow-Mildewing Resistance in Knox Wheat. Phytopathology 1977, 67, 1051–1056. [Google Scholar] [CrossRef] [Green Version]
- Dere, S.; Güneş, T.; Sivaci, R. Spectrophotometric Determination of Chlorophyll-a, b and Total Carotenoid Contents of Some Algae Species Using Different Solvents. Turk. J. Botany 1998, 22, 13–17. [Google Scholar]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant activity of plant extracts containing phenolic compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some algerian medicinal plants extracts containing phenolic compounds. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Adám, A.; Farkas, T.; Somlyai, G.; Hevesi, M.; Király, Z. Consequence of O2·− generation during a bacterially induced hypersensitive reaction in tobacco: Deterioration of membrane lipids. Physiol. Mol. Plant Pathol. 1989, 34, 13–26. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzym. 1984, 105, 121–126. [Google Scholar]
- Harrach, B.D.; Fodor, J.; Pogány, M.; Preuss, J.; Barna, B. Antioxidant, ethylene and membrane leakage responses to powdery mildew infection of near-isogenic barley lines with various types of resistance. Eur. J. Plant Pathol. 2008, 121, 21–33. [Google Scholar] [CrossRef]
- Malik, C.P.; Singh, M.B. Plant Enzymology and Histo-Enzymology; Kalyani Publishers: New Delhi, India, 1980. [Google Scholar]
- Nehela, Y.; Hijaz, F.; Elzaawely, A.A.; El-Zahaby, H.M.; Killiny, N. Phytohormone profiling of the sweet orange (Citrus sinensis (L.) Osbeck) leaves and roots using GC-MS-based method. J. Plant Physiol. 2016, 199, 12–17. [Google Scholar] [CrossRef]
- Killiny, N.; Nehela, Y. Metabolomic Response to Huanglongbing: Role of Carboxylic Compounds in Citrus sinensis Response to ‘Candidatus Liberibacter asiaticus’ and Its Vector, Diaphorina citri. Mol. Plant-Microbe Interact. 2017, 30, 666–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nehela, Y.; Hijaz, F.; Elzaawely, A.A.; El-Zahaby, H.M.; Killiny, N. Citrus phytohormonal response to Candidatus Liberibacter asiaticus and its vector Diaphorina citri. Physiol. Mol. Plant Pathol. 2018, 102, 24–35. [Google Scholar] [CrossRef]
- Nehela, Y.; Killiny, N. Infection with phytopathogenic bacterium inhibits melatonin biosynthesis, decreases longevity of its vector, and suppresses the free radical-defense. J. Pineal Res. 2018, e12511. [Google Scholar] [CrossRef]
- Nehela, Y.; Killiny, N. Melatonin is involved in citrus response to the pathogen huanglongbing via modulation of phytohormonal biosynthesis. Plant Physiol. 2020, 184, 2216–2239. [Google Scholar] [CrossRef]
- Nehela, Y.; Killiny, N. ‘Candidatus Liberibacter asiaticus’ and Its Vector, Diaphorina citri, Augment the Tricarboxylic Acid Cycle of Their Host via the g-Aminobutyric Acid Shunt and Polyamines Pathway. Mol. Plant-Microbe Interact. 2019, 32, 413–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gan, R.Y.; Chan, C.L.; Yang, Q.Q.; Li, H.B.; Zhang, D.; Ge, Y.Y.; Gunaratne, A.; Ge, J.; Corke, H. Bioactive compounds and beneficial functions of sprouted grains. In Sprouted Grains: Nutritional Value, Production, and Applications; Feng, H., Nemzer, B., DeVries, J.W., Eds.; Woodhead Publishing and AACC International Press: Cambridge, UK, 2019; pp. 191–246. ISBN 9780128115251. [Google Scholar]
- Vondráková, Z.; Malbeck, J.; Trávníčková, A.; Černý, R.; Cvikrová, M. Phenolic acids in selected scab-resistant and mildew-tolerant apple cultivars. Acta Physiol. Plant. 2020, 42, 1–10. [Google Scholar] [CrossRef]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. In Phytochemistry: Advances in Research; Imperato, F., Ed.; Research Signpost: Cerala, India, 2006; ISBN 8130800349. [Google Scholar]
- Vagiri, M.; Johansson, E.; Rumpunen, K. Phenolic compounds in black currant leaves—An interaction between the plant and foliar diseases? J. Plant Interact. 2017, 12, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Berne, S.; Kovačič, L.; Sova, M.; Kraševec, N.; Gobec, S.; Križaj, I.; Komel, R. Benzoic acid derivatives with improved antifungal activity: Design, synthesis, structure-activity relationship (SAR) and CYP53 docking studies. Bioorg. Med. Chem. 2015, 23, 4264–4276. [Google Scholar] [CrossRef]
- Kim, J.H.; Campbell, B.C.; Mahoney, N.; Chan, K.L.; Molyneux, R.J.; Balajee, A. Augmenting the activity of antifungal agents against aspergilli using structural analogues of benzoic acid as chemosensitizing agents. Fungal Biol. 2010, 114, 817–824. [Google Scholar] [CrossRef]
- Mandal, D.; Pal, R.; Mohanty, A.K. Management of early blight of tomato. Indian J. Hortic. 2019, 76, 548–551. [Google Scholar] [CrossRef]
- Laborda, P.; Li, C.; Zhao, Y.; Tang, B.; Ling, J.; He, F.; Liu, F. Antifungal Metabolite p-Aminobenzoic Acid (pABA): Mechanism of Action and Efficacy for the Biocontrol of Pear Bitter Rot Disease. J. Agric. Food Chem. 2019, 67, 2157–2165. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.W.; McKoy, J.; Del Valle, R.; Armstrong, D.; Bernard, E.M.; Katz, N.; Gordon, R.E. Fungal cell wall septation and cytokinesis are inhibited by bleomycins. Antimicrob. Agents Chemother. 2003, 47, 3281–3289. [Google Scholar] [CrossRef] [Green Version]
- Freitag, J.; Lanver, D.; Böhmer, C.; Schink, K.O.; Bölker, M.; Sandrock, B. Septation of Infectious Hyphae Is Critical for Appressoria Formation and Virulence in the Smut Fungus Ustilago maydis. PLoS Pathog. 2011, 7, e1002044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Rocha Neto, A.C.; Maraschin, M.; Di Piero, R.M. Antifungal activity of salicylic acid against Penicillium expansum and its possible mechanisms of action. Int. J. Food Microbiol. 2015, 215, 64–70. [Google Scholar] [CrossRef]
- Warth, A.D. Mechanism of action of benzoic acid on Zygosaccharomyces bailii: Effects on glycolytic metabolite levels, energy production, and intracellular pH. Appl. Environ. Microbiol. 1991, 57, 3410–3414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EL-Nagar, A.; Elzaawely, A.A.; Taha, N.A.; Nehela, Y. The antifungal activity of gallic acid and its derivatives against Alternaria solani, the causal agent of tomato early blight. Agronomy 2020, 10, 1402. [Google Scholar] [CrossRef]
- Shabana, Y.M.; Abdel-Fattah, G.M.; Ismail, A.E.; Rashad, Y.M. Control of brown spot pathogen of rice (Bipolaris oryzae) using some phenolic antioxidants. Braz. J. Microbiol. 2008, 39, 438–444. [Google Scholar] [CrossRef] [Green Version]
- Zakariyya, F.; Wahyu Susilo, A.; Iman Santoso, T.; Susilo Addy, H.; Pancaningtyas, S. Role of Exogenous Salicylic Acid and Benzoic Acid Applications to Vascular Streak Dieback Disease Attack on Cocoa Seedlings. Pelita Perkeb. 2018, 34, 33. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Li, K.; Guo, X.; Ma, L.; Guo, Y.; Liu, Z. Developmental characteristics of grapevine seedlings root border cells and their response to ρ-hydroxybenzoic acid. Plant Soil 2019, 443, 199–218. [Google Scholar] [CrossRef]
- Sahu, R.; Sharaff, M.; Pradhan, M.; Sethi, A.; Bandyopadhyay, T.; Mishra, V.K.; Chand, R.; Chowdhury, A.K.; Joshi, A.K.; Pandey, S.P. Elucidation of defense-related signaling responses to spot blotch infection in bread wheat (Triticum aestivum L.). Plant J. 2016, 86, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sahu, R.; Navathe, S.; Mishra, V.K.; Chand, R.; Singh, P.K.; Joshi, A.K.; Pandey, S.P. Natural Variation in Elicitation of Defense-Signaling Associates to Field Resistance Against the Spot Blotch Disease in Bread Wheat (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 636. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Racchi, M.L. Antioxidant defenses in plants with attention to Prunus and Citrus spp. Antioxidants 2013, 2, 340–369. [Google Scholar] [CrossRef] [PubMed]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 2106. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Xu, B.T.; Xu, X.R.; Gan, R.Y.; Zhang, Y.; Xia, E.Q.; Li, H. Bin Antioxidant capacities and total phenolic contents of 62 fruits. Food Chem. 2011, 129, 345–350. [Google Scholar] [CrossRef]
- Rojas, C.M.; Senthil-Kumar, M.; Tzin, V.; Mysore, K.S. Regulation of primary plant metabolism during plant-pathogen interactions and its contribution to plant defense. Front. Plant Sci. 2014, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Hatcher, P.E.; Moore, J.; Taylor, J.E.; Tinney, G.W.; Paul, N.D. Phytohormones and plant-herbivore-pathogen interactions: Integrating the molecular with the ecological. Ecology 2004, 85, 59–69. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Lefevere, H.; Bauters, L.; Gheysen, G. Salicylic Acid Biosynthesis in Plants. Front. Plant Sci. 2020, 11, 338. [Google Scholar] [CrossRef] [PubMed]
- Catinot, J.; Buchala, A.; Abou-Mansour, E.; Métraux, J.-P. Salicylic acid production in response to biotic and abiotic stress depends on isochorismate in Nicotiana benthamiana. FEBS Lett. 2008, 582, 473–478. [Google Scholar] [CrossRef] [Green Version]
- Hao, Q.; Wang, W.; Han, X.; Wu, J.; Lyu, B.; Chen, F.; Caplan, A.; Li, C.; Wu, J.; Wang, W.; et al. Isochorismate-based salicylic acid biosynthesis confers basal resistance to Fusarium graminearum in barley. Mol. Plant Pathol. 2018, 19, 1995–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zheng, Z.; Huang, J.; Lai, Z.; Fan, B. Biosynthesis of salicylic acid in plants. Plant Signal. Behav. 2009, 4, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Mercado-Blanco, J.; Van Der Drift, K.M.G.M.; Olsson, P.E.; Thomas-Oates, J.E.; Van Loon, L.C.; Bakker, P.A.H.M. Analysis of the pmsCEAB gene cluster involved in biosynthesis of salicylic acid and the siderophore pseudomonine in the biocontrol strain Pseudomonas fluorescens WCS374. J. Bacteriol. 2001, 183, 1909–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coquoz, J.-L.; Buchala, A.; Métraux, J.-P. The Biosynthesis of Salicylic Acid in Potato Plants1. Plant Physiol. 1998, 117, 1095–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, C.-J.; Li, L.; Shang, Q.-M.; Liu, X.-Y.; Zhang, Z.-G. Endogenous salicylic acid accumulation is required for chilling tolerance in cucumber (Cucumis sativus L.) seedlings. Planta 2014, 240, 687–700. [Google Scholar] [CrossRef]
- León, J.; Shulaev, V.; Yalpani, N.; Lawton, M.A.; Raskin, I. Benzoic acid 2-hydroxylase, a soluble oxygenase from tobacco, catalyzes salicylic acid biosynthesis. Proc. Natl. Acad. Sci. USA 1995, 92, 10413–10417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spletzer, M.E.; Enyedi, A.J. Salicylic acid induces resistance to Alternaria solani in hydroponically grown tomato. Phytopathology 1999, 89, 722–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, C.K.; Wang, C.Y.; Gross, K.C.; Smith, D.L. Jasmonate and salicylate induce the expression of pathogenesis-related-protein genes and increase resistance to chilling injury in tomato fruit. Planta 2002, 214, 895–901. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 (ppm) | 95% Confidence Interval | IC99 (ppm) | 95% Confidence Interval | Overall Model Fit | ||||
|---|---|---|---|---|---|---|---|---|---|
| Lower | Upper | Lower | Upper | χ2 | p-Value | Cox and Snell R2 | |||
| Difenoconazole | 1.24 | 0.81 | 1.71 | 226.67 | 134.61 | 462.59 | 168.92 | <0.0001 | 0.1423 |
| Benzoic acid | 44.69 | 40.05 | 49.31 | 296.60 | 223.04 | 445.77 | 129.36 | <0.0001 | 0.2280 |
| ρ-Hydroxybenzoic acid | 58.80 | 45.96 | 75.24 | 1317.74 | 1015.76 | 1709.49 | 52.84 | <0.0001 | 0.1003 |
| Protocatechuic acid | 49.79 | 44.84 | 54.96 | 341.62 | 251.26 | 532.72 | 123.18 | <0.0001 | 0.2184 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nehela, Y.; Taha, N.A.; Elzaawely, A.A.; Xuan, T.D.; A. Amin, M.; Ahmed, M.E.; El-Nagar, A. Benzoic Acid and Its Hydroxylated Derivatives Suppress Early Blight of Tomato (Alternaria solani) via the Induction of Salicylic Acid Biosynthesis and Enzymatic and Nonenzymatic Antioxidant Defense Machinery. J. Fungi 2021, 7, 663. https://doi.org/10.3390/jof7080663
Nehela Y, Taha NA, Elzaawely AA, Xuan TD, A. Amin M, Ahmed ME, El-Nagar A. Benzoic Acid and Its Hydroxylated Derivatives Suppress Early Blight of Tomato (Alternaria solani) via the Induction of Salicylic Acid Biosynthesis and Enzymatic and Nonenzymatic Antioxidant Defense Machinery. Journal of Fungi. 2021; 7(8):663. https://doi.org/10.3390/jof7080663
Chicago/Turabian StyleNehela, Yasser, Naglaa A. Taha, Abdelnaser A. Elzaawely, Tran Dang Xuan, Mohammed A. Amin, Mohamed E. Ahmed, and Asmaa El-Nagar. 2021. "Benzoic Acid and Its Hydroxylated Derivatives Suppress Early Blight of Tomato (Alternaria solani) via the Induction of Salicylic Acid Biosynthesis and Enzymatic and Nonenzymatic Antioxidant Defense Machinery" Journal of Fungi 7, no. 8: 663. https://doi.org/10.3390/jof7080663