The Fission Yeast RNA-Binding Protein Meu5 Is Involved in Outer Forespore Membrane Breakdown during Spore Formation

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains, Media, and Culture Conditions
2.2. Fluorescence Microscopy
2.3. Quick-Freeze Deep-Etch Electron Microscopy
2.4. Thin-Section Electron Microscopy
2.5. Isolation of meu5+
2.6. Reverse Transcription PCR
2.7. Western Blotting
2.8. Construction of the Plasmid for meu5+ Overexpression
3. Results
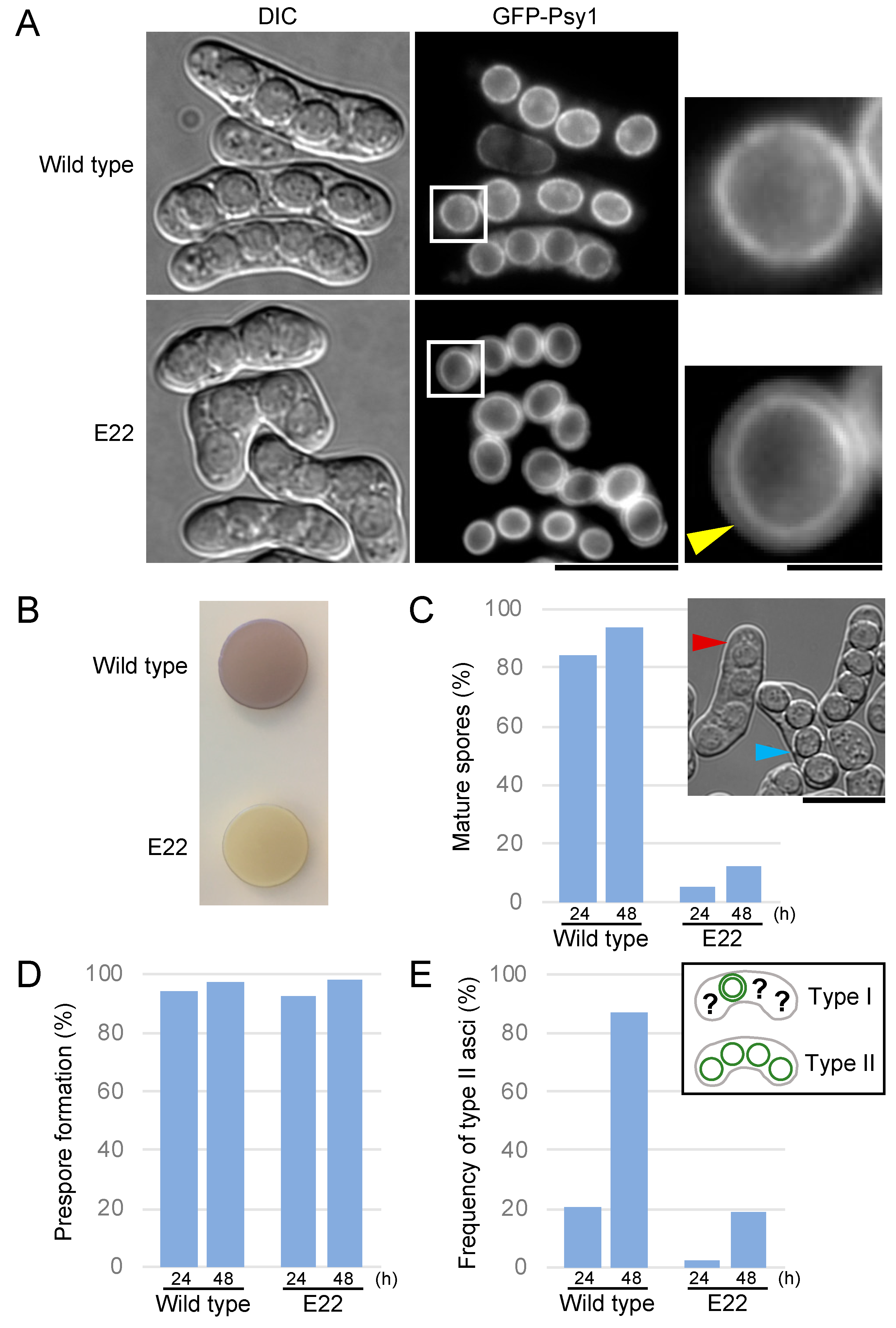
3.1. The E22 Mutant Shows a Defect in Spore Maturation
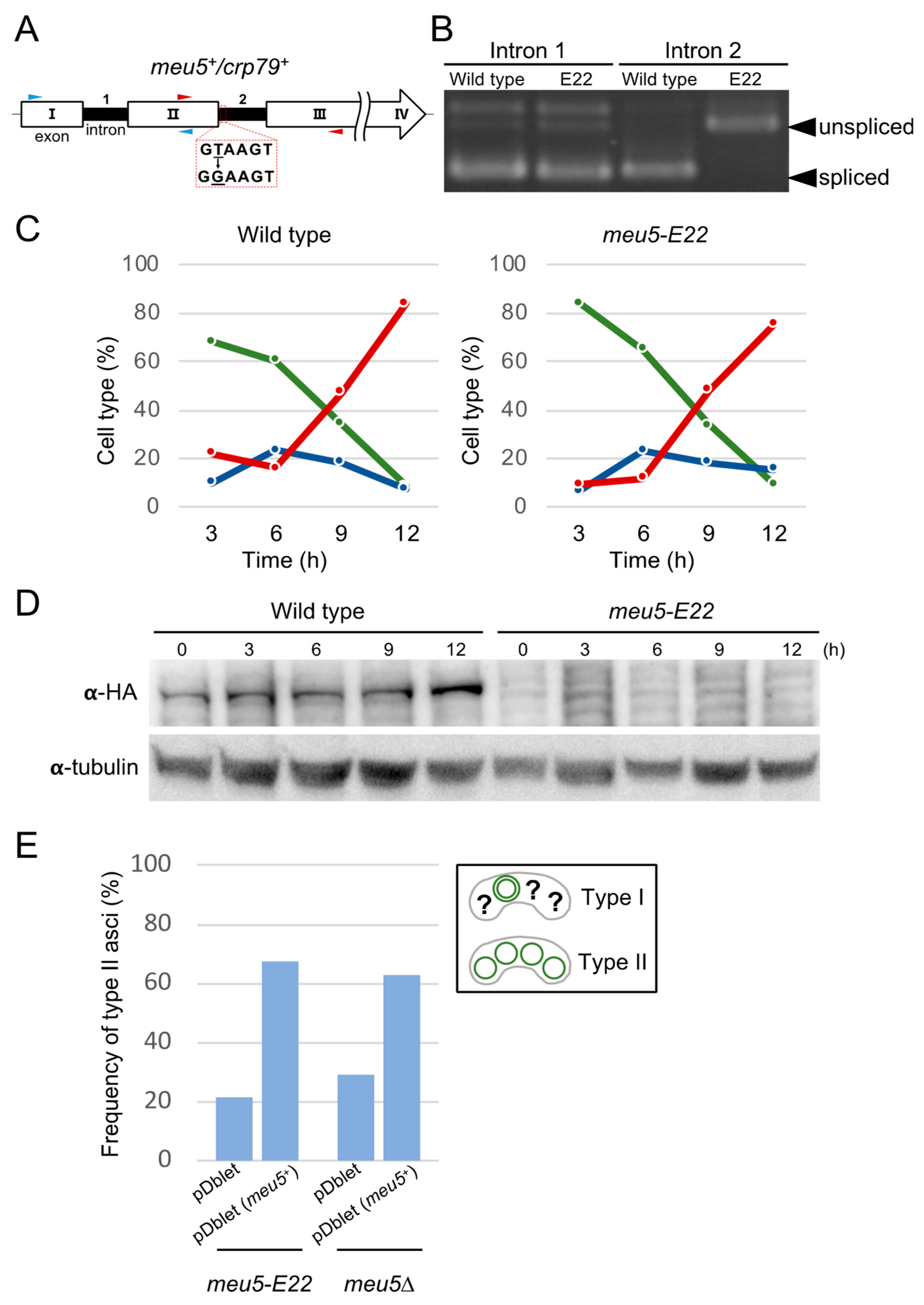
3.2. The E22 Mutation Is Located in the meu5+ Gene Encoding the RNA-Binding Protein Meu5
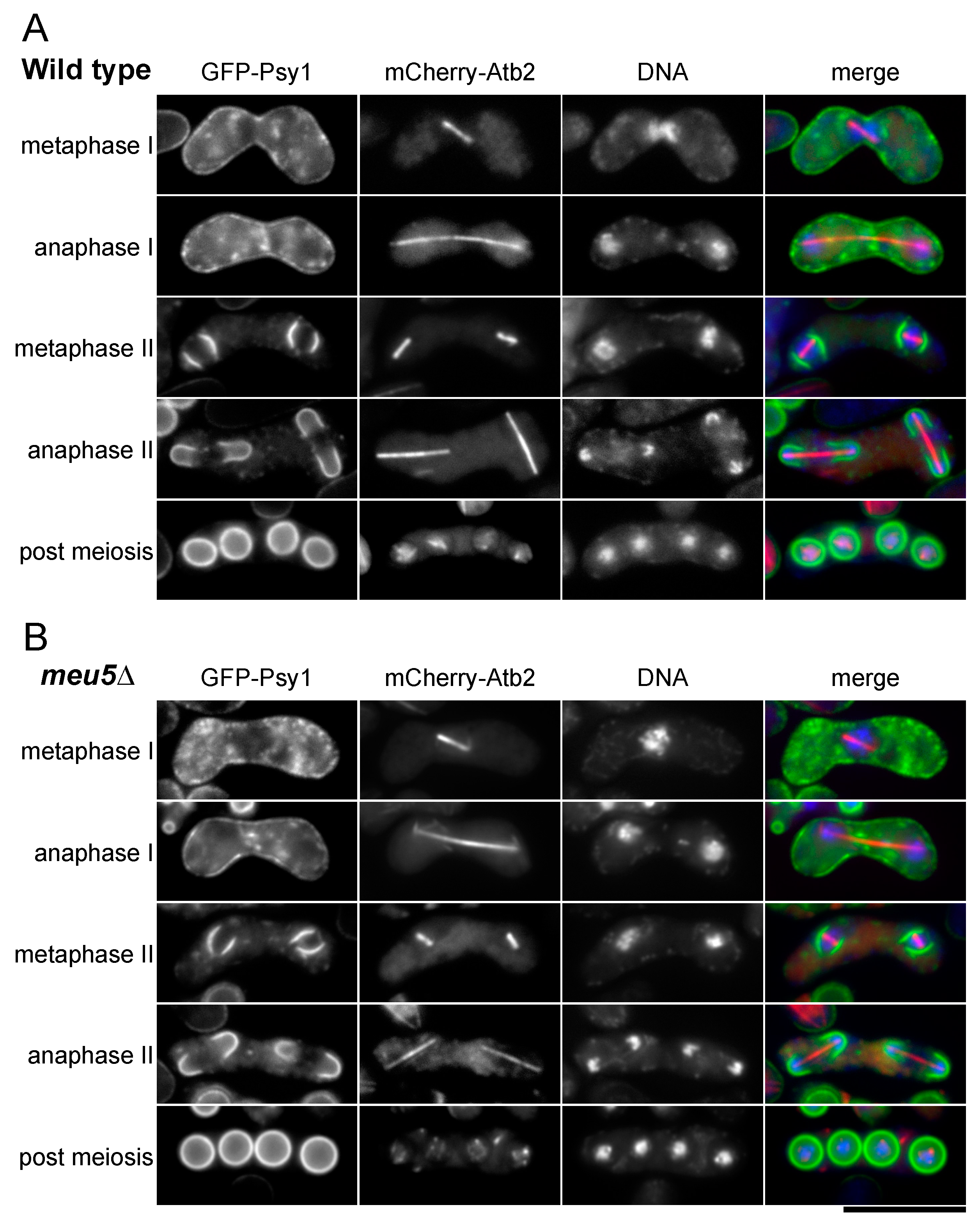
3.3. Meu5 Is Dispensable for Initiation of FSM Formation and Expansion of the FSM
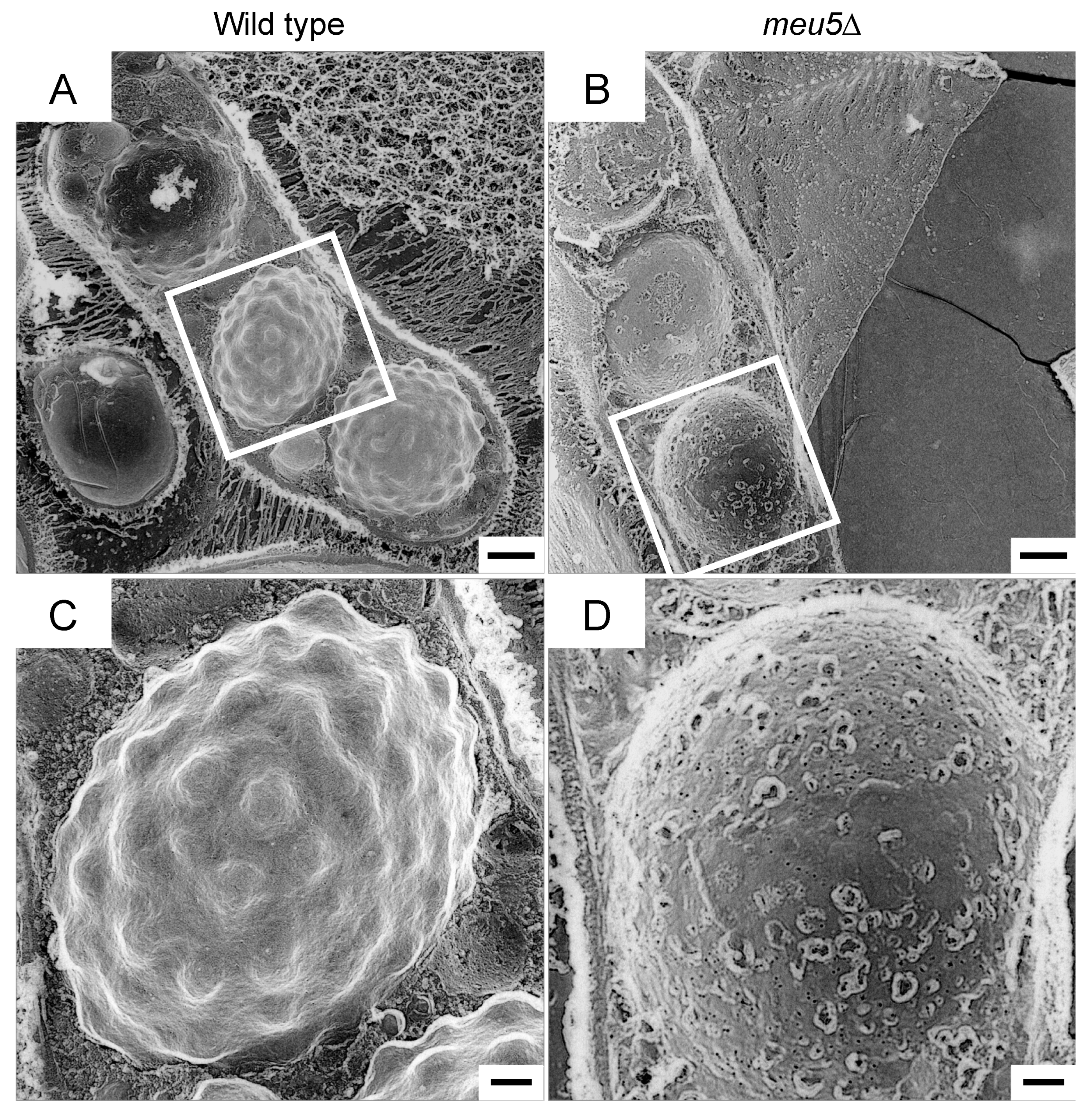
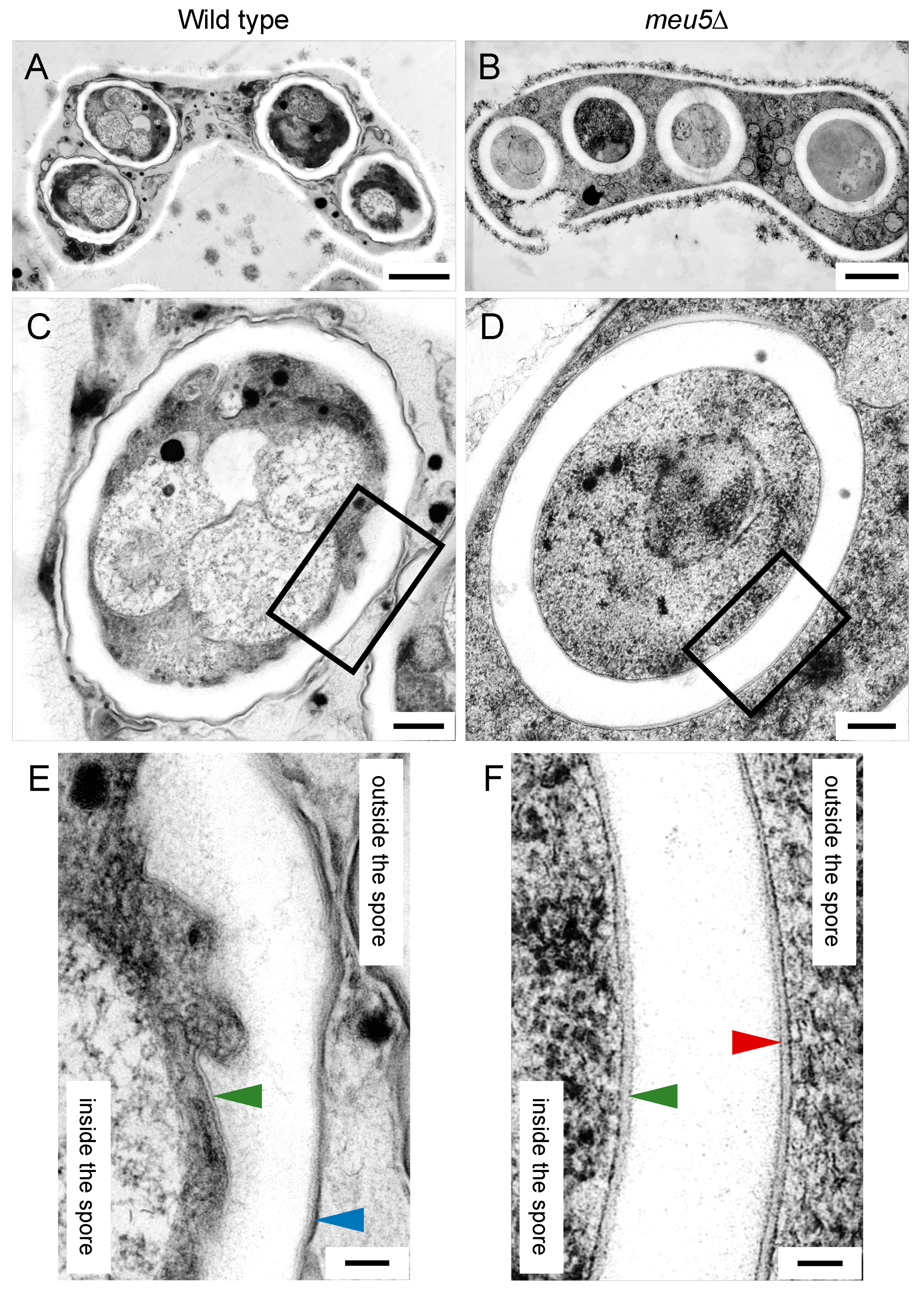
3.4. Meu5 Is Involved in Breakdown of the Outer FSM
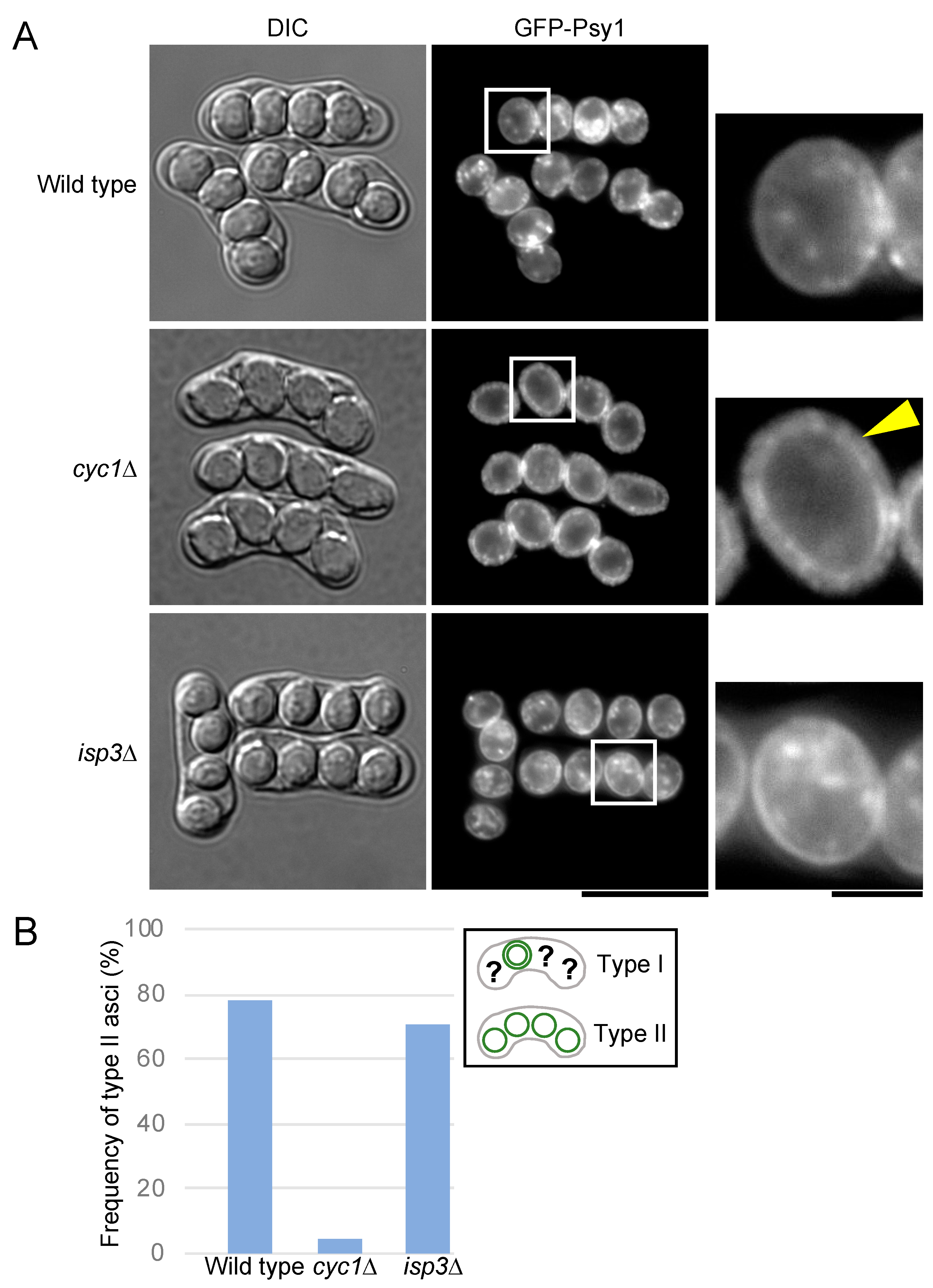
3.5. Cytochrome c Is Involved in Breakdown of the Outer FSM
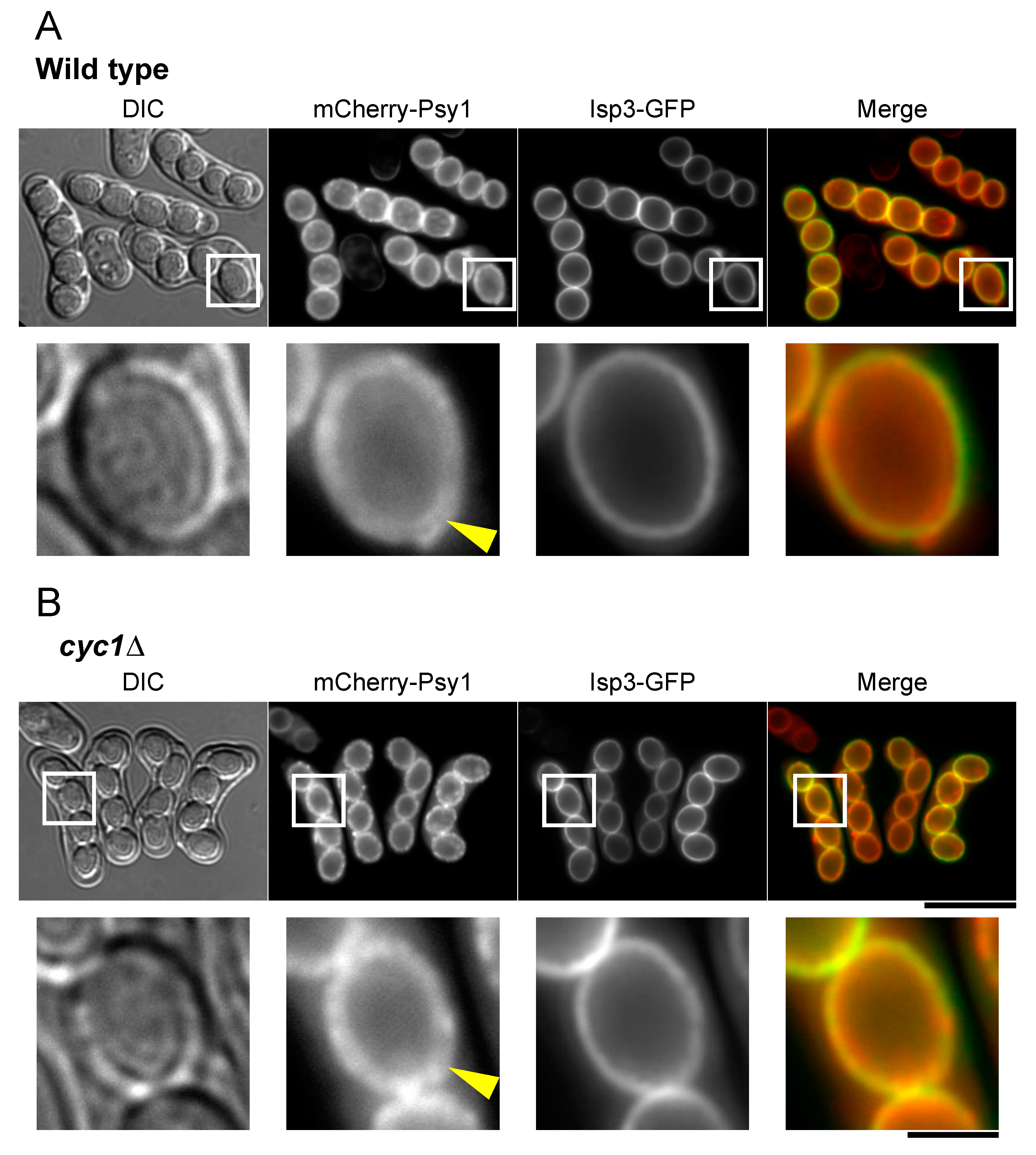
3.6. Localization of Isp3 to the Spore Periphery Precedes Outer Membrane Breakdown
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yoo, B.Y.; Calleja, G.B.; Johnson, B.F. Ultrastructural changes of the fission yeast (Schizosaccharomyces pombe) during ascospore formation. Arch. Mikrobiol. 1973, 91, 1–10. [Google Scholar] [CrossRef]
- Shimoda, C.; Nakamura, T. Control of late meiosis and ascospore formation. In Molecular Biology of Schizosaccharomyces pombe; Springer: Berlin/Heidelberg, Germany, 2004; pp. 311–327. [Google Scholar]
- Hirata, A.; Tanaka, K. Nuclear behavior during conjugation and meiosis in the fission yeast Schizosaccharomyces pombe. J. Gen. Appl. Microbiol. 1982, 28, 263–274. [Google Scholar] [CrossRef]
- Nakamura, T.; Nakamura-Kubo, M.; Hirata, A.; Shimoda, C. The Schizosaccharomyces pombe spo3+ gene is required for assembly of the forespore membrane and genetically interacts with psy1+ encoding syntaxin-like protein. Mol. Biol. Cell 2001, 12, 3955–3972. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Asakawa, H.; Nakase, Y.; Kashiwazaki, J.; Hiraoka, Y.; Shimoda, C. Live observation of forespore membrane formation in fission yeast. Mol. Biol. Cell 2008, 19, 3544–3553. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Hirata, A. Ascospore development in the fission yeasts Schizosaccharomyces pombe and S. japonicus. J. Cell. Sci. 1982, 56, 263–279. [Google Scholar] [PubMed]
- Egel, R. Selective spore survival during replica-plating of fission yeast. Arch. Microbiol. 1977, 112, 109–110. [Google Scholar] [CrossRef]
- Kupelic, M.; Byers, B.; Esposito, B. Meiosis and sporulation in Saccharomyces cerevisiae. In The Molecular and Cellular Biology of the Yeast Saccharomyces; Jones, E.W., Pringle, J.R., Broach, J.R., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997; pp. 889–1036. [Google Scholar]
- Neiman, A.M. Sporulation in the budding yeast Saccharomyces cerevisiae. Genetics 2011, 189, 737–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, G.J.; van den Ende, H.; Klis, F.M. Differential regulation of cell wall biogenesis during growth and development in yeast. Microbiology 2001, 147, 781–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreger-Van Rij, N.J.W. Electron microscopy of germinating ascospores of Saccharomyces cerevisiae. Arch. Microbiol. 1978, 117, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Briza, P.; Winkler, G.; Kalchhauser, H.; Breitenbach, M. Dityrosine is a prominent component of the yeast ascospore wall. A proof of its structure. J. Biol. Chem. 1986, 261, 4288–4294. [Google Scholar]
- Briza, P.; Ellinger, A.; Winkler, G.; Breitenbach, M. Chemical composition of the yeast ascospore wall. The second outer layer consists of chitosan. J. Biol. Chem. 1988, 263, 11569–11574. [Google Scholar] [PubMed]
- Bush, D.A.; Horisberger, M.; Horman, I.; Wursch, P. The wall structure of Schizosaccharomyces pombe. Microbiology 1974, 81, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arellano, M.; Cartagena-Lirola, H.; Nasser Hajibagheri, M.A.; Durán, A.; Henar Valdivieso, M. Proper ascospore maturation requires the chs1+ chitin synthase gene in Schizosaccharomyces pombe. Mol. Microbiol. 2000, 35, 79–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, Y.; Tanaka, K.; Nakagawa, T.; Matsuda, H.; Kawamukai, M. Genetic analysis of chs1+ and chs2+ encoding chitin synthases from Schizosaccharomyces pombe. Biosci. Biotechnol. Biochem. 2004, 68, 1489–1499. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Tanaka, K.; Matsuda, H.; Kawamukai, M. cda1+, encoding chitin deacetylase is required for proper spore formation in Schizosaccharomyces pombe. FEBS Lett. 2005, 579, 2737–2743. [Google Scholar] [CrossRef] [Green Version]
- Shimoda, C. Existence of a major spore wall protein (23K) in the fission yeast Schizosaccharomyces pombe: A possible sporulation-specific protein. Arch. Microbiol. 1983, 136, 26–27. [Google Scholar] [CrossRef]
- Fukunishi, K.; Miyakubi, K.; Hatanaka, M.; Otsuru, N.; Hirata, A.; Shimoda, C.; Nakamura, T. The fission yeast spore is coated by a proteinaceous surface layer comprising mainly Isp3. Mol. Biol. Cell 2014, 25, 1549–1559. [Google Scholar] [CrossRef]
- Coluccio, A.; Bogengruber, E.; Conrad, M.N.; Dresser, M.E.; Briza, P.; Neiman, A.M. Morphogenetic pathway of spore wall assembly in Saccharomyces cerevisiae. Eukaryotic Cell 2004, 3, 1464–1475. [Google Scholar] [CrossRef] [Green Version]
- Egel, R.; Egel-Mitani, M. Premeiotic DNA synthesis in fission yeast. Exp. Cell Res. 1974, 88, 127–134. [Google Scholar] [CrossRef]
- Moreno, S.; Klar, A.; Nurse, P. Molecular genetic analysis of fission yeast Schizosaccharomyces pombe. Methods Enzymol. 1991, 194, 793–823. [Google Scholar]
- Miyata, M.; Petersen, J.D. Spike structure at the interface between gliding Mycoplasma mobile cells and glass surfaces visualized by rapid-freeze-and-fracture electron microscopy. J. Bacteriol. 2004, 186, 4382–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tulum, I.; Tahara, Y.O.; Miyata, M. Peptidoglycan layer and disruption processes in Bacillus subtilis cells visualized using quick-freeze, deep-etch electron microscopy. Microscopy 2019, 68, 441–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Yonekawa, T.; Kawasaki, Y.; Kai, M.; Furuya, K.; Iwasaki, M.; Murakami, H.; Yanagida, M.; Okayama, H. Fission yeast Eso1p is required for establishing sister chromatid cohesion during S phase. Mol. Cell. Biol. 2000, 20, 3459–3469. [Google Scholar] [CrossRef]
- Gutz, H.; Heslot, H.; Leupold, U.; Loprieno, N. Schizosaccharomyces pombe. In Handbook of Genetics 1; Plenum Press: New York, NY, USA, 1974; pp. 395–446. [Google Scholar]
- Masai, H.; Miyake, T.; Arai, K. hsk1+, a Schizosaccharomyces pombe gene related to Saccharomyces cerevisiae CDC7, is required for chromosomal replication. EMBO J. 1995, 14, 3094–3104. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.; Sherwin, T.; Sasse, R.; MacRae, T.H.; Baines, A.J.; Gull, K. Definition of individual components within the cytoskeleton of Trypanosoma brucei by a library of monoclonal antibodies. J. Cell Sci. 1989, 93, 491–500. [Google Scholar] [PubMed]
- Brun, C.; Dubey, D.D.; Huberman, J. pDblet, a stable autonomously replicating shuttle vector for Schizosaccharomyces pombe. Gene 1995. [Google Scholar] [CrossRef]
- Imada, K.; Nakamura, T. The exocytic Rabs Ypt3 and Ypt2 regulate the early step of biogenesis of the spore plasma membrane in fission yeast. Mol. Biol. Cell 2016, 27, 3317–3328. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Miyashita, K.; Saito, T.T.; Yoneki, T.; Kakihara, Y.; Nabeshima, K.; Kishi, Y.A.; Shimoda, C.; Nojima, H. Comprehensive isolation of meiosis-specific genes identifies novel proteins and unusual non-coding transcripts in Schizosaccharomyces pombe. Nucleic Acids Res. 2001, 29, 2327–2337. [Google Scholar] [CrossRef] [Green Version]
- Thakurta, A.G.; Whalen, W.A.; Yoon, J.H.; Bharathi, A.; Kozak, L.; Whiteford, C.; Love, D.C.; Hanover, J.A.; Dhar, R. Crp79p, like Mex67p, is an auxiliary mRNA export factor in Schizosaccharomyces pombe. Mol. Biol. Cell 2002, 13, 2571–2584. [Google Scholar] [CrossRef] [Green Version]
- Amorim, M.J.; Cotobal, C.; Duncan, C.; Mata, J. Global coordination of transcriptional control and mRNA decay during cellular differentiation. Mol. Syst. Biol. 2010, 6, 380. [Google Scholar] [CrossRef]
- Heuser, J.; Reese, T.; Dennis, M.; Jan, Y.; Jan, L.; Evans, L. Synaptic vesicle exocytosis captured by quick freezing and correlated with quantal transmitter release. J. Cell Sci. 1979, 81, 275–300. [Google Scholar] [CrossRef] [Green Version]
- Heuser, J.E. The origins and evolution of freeze-etch electron microscopy. J. Electron Microsc. 2011, 60, S3–S29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilhofer, M.; Ladinsky, M.S.; McDowall, A.W.; Jensen, G.J. Bacterial TEM: New insights from cryo-microscopy. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2010; Volume 96, pp. 21–45. [Google Scholar]
- Zhao, X.; Norris, S.J.; Liu, J. Molecular architecture of the bacterial flagellar motor in cells. Biochemistry 2014, 53, 4323–4333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Abe, H.; Hirata, A.; Shimoda, C. ADAM family protein Mde10 is essential for development of spore envelopes in the fission yeast Schizosaccharomyces pombe. Eukaryot Cell 2004, 3, 27–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregan, J.; Rabitsch, P.K.; Sakem, B.; Csutak, O.; Latypov, V.; Lehmann, E.; Kohli, J.; Nasmyth, K. Novel genes required for meiotic chromosome segregation are identified by a high-throughput knockout screen in fission yeast. Curr. Biol. 2005, 15, 1663–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phadnis, N.; Cipak, L.; Polakova, S.; Hyppa, R.W.; Cipakova, I.; Anrather, D.; Karvaiova, L.; Mechtler, K.; Smith, G.R.; Gregan, J. Casein kinase 1 and phosphorylation of cohesin subunit Rec11 (SA3) promote meiotic recombination through linear element formation. PLoS Genet 2015, 11, e1005225. [Google Scholar] [CrossRef] [Green Version]
- Blyth, J.; Makrantoni, V.; Barton, R.E.; Spanos, C.; Rappsilber, J.; Marston, A.L. Genes important for Schizosaccharomyces pombe meiosis identified through a functional genomics screen. Genetics 2018, 208, 589–603. [Google Scholar] [CrossRef] [Green Version]
- Briza, P.; Ellinger, A.; Winkler, G.; Breitenbach, M. Characterization of a DL-dityrosine-containing macromolecule from yeast ascospore walls. J. Biol. Chem. 1990, 265, 15118–15123. [Google Scholar]
- Briza, P.; Kalchhauser, H.; Pittenauer, E.; Allmaier, G.; Breitenbach, M. N,N’-Bisformyl dityrosine is an in vivo precursor of the yeast ascospore wall. Eur. J. Biochem. 1996, 239, 124–131. [Google Scholar] [CrossRef]
- Russell, P.R.; Hall, B.D. Structure of the Schizosaccharomyces pombe cytochrome c gene. Mol. Cell. Biol. 1982, 2, 106–116. [Google Scholar] [CrossRef] [Green Version]
- Miki, R.; Saiki, R.; Ozoe, Y.; Kawamukai, M. Comparison of a coq7 deletion mutant with other respiration-defective mutants in fission yeast. FEBS J. 2008, 275, 5309–5324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suda, Y.; Nakanishi, H.; Mathieson, E.M.; Neiman, A.M. Alternative modes of organellar segregation during sporulation in Saccharomyces cerevisiae. Eukaryot Cell 2007, 6, 2009–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashiwazaki, J.; Nakamura, T.; Iwaki, T.; Takegawa, K.; Shimoda, C. A role for fission yeast Rab GTPase Ypt7p in sporulation. Cell Struct. Funct. 2005, 30, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Description (Pombase) | Frequency of Type II Asci (%) |
|---|---|---|
| cyc1 | cytochrome c | 4.3 |
| are2 | acyl-CoA-sterol acyltransferase Are2 | 26.8 |
| pmp3 | plasma membrane proteolipid Pmp3 (predicted) | 27.9 |
| SPBPB2B2.07c | S. pombe specific DUF999 protein family 7 | 29.8 |
| mug129 | Schizosaccharomyces specific protein Mug129 | 42.1 |
| agn2 | glucan endo-1,3-alpha-glucosidase Agn2 | 43.8 |
| mug63 | TLDc domain protein 1, implicated in response to oxidative stress | 43.8 |
| atg24 (snx4) | autophagy associated PX/BAR domain sorting nexin Atg24 | 44.1 |
| ctl1 | choline transporter-like, implicated in autophagy Ctl1 | 45.5 |
| hhp1 | serine/threonine protein kinase Hhp1 | 47.5 |
| pmc1 | vacuolar calcium transporting P-type ATPase P2 type, Pmc1 | 52.2 |
| fzr2 | meiotic fizzy-related APC coactivator Fzr2 (predicted) | 52.7 |
| ubp4 | ubiquitin C-terminal hydrolase Ubp4 | 52.8 |
| fis1 | mitochondrial fission protein Fis1 (predicted) | 53.0 |
| dsk1 | SR protein-specific kinase Dsk1 | 54.6 |
| SPAC750.06c | S. pombe specific DUF999 protein family 4 | 56.4 |
| uvi15 | tail anchored plasma membrane protein Uvi15 | 57.8 |
| pcy1 (pct1) | choline phosphate cytidylyltransferase Pcy1 (predicted) | 59.1 |
| meu6 | pleckstrin homology domain protein Meu6 | 59.5 |
| nep2 (mug120) | NEDD8 protease Nep2 | 62.6 |
| omh4 | alpha-1,2-mannosyltransferase Omh4 (predicted) | 62.8 |
| mug58 | GLYK family kinase of unknown specificity, implicated in nucleotide metabolism (predicted) | 63.6 |
| gsk31 | serine/threonine protein kinase Gsk31 (predicted) | 66.7 |
| gor1 | glyoxylate reductase (predicted) | 66.7 |
| SPBC1348.01 | S. pombe specific DUF999 protein family 5 | 67.0 |
| isp3 (meu4) | spore wall structural constituent Isp3 | 70.7 |
| sdu1 (mug67, hag1) | PPPDE peptidase family deubiquitinase/desumoylase Sdu1 (predicted) | 71.1 |
| psk1 | ribosomal protein S6 kinase Psk1 | 71.4 |
| elo1 | fatty acid elongase Elo1 | 72.0 |
| idn1 | gluconokinase | 73.5 |
| ryh1 (hos1, sat7) | GTPase Ryh1 | 73.7 |
| gma12 | alpha-1,2-galactosyltransferase Gma12 | 75.2 |
| git5 (gpb1) | heterotrimeric G protein beta (WD repeat) subunit Git5 | 77.0 |
| mug111 | major facilitator family transmembrane transporter Mug111 (predicted) | 77.3 |
| SPAC212.04c | S. pombe specific DUF999 family protein 1 | 77.3 |
| meu7 (aah4) | alpha-amylase homolog Aah4 | 77.5 |
| SPAC212.01c | S. pombe specific DUF999 family protein 2 | 78.3 |
| pmp31 (mug75) | plasma membrane proteolipid Pmp31 | 79.0 |
| mug110 | Schizosaccharomyces specific protein Mug110 | 79.2 |
| mde10 (mug139) | spore wall assembly ADAM family peptidase Mde10 | 79.2 |
| mug86 | plasma membrane acetate transmembrane transporter (predicted) | 79.6 |
| meu31 | Schizosaccharomyces specific protein Meu31 | 80.0 |
| pet2 | Golgi phosphoenolpyruvate transmembrane transporter Pet2 | 80.4 |
| pso2 (snm1) | DNA 5’ exonuclease (predicted) | 80.6 |
| fbp1 | fructose-1,6-bisphosphatase Fbp1 | 82.5 |
| meu34 (mug145) | ubiquitin-protein ligase E3 Meu34, human RNF13 family homolog, unknown biological role (predicted) | 83.3 |
| SPAC4F10.16c | plasma membrane phospholipid-translocating ATPase complex, ATPase subunit (predicted) | 84.5 |
| rit1 | initiator methionine tRNA 2’-O-ribosyl phosphate transferase (predicted) | 85.8 |
| gld1 | mitochondrial glycerol dehydrogenase Gld1 | 86.1 |
| SPCC1739.08c | short chain dehydrogenase (predicted) | 88.2 |
| erf2 (mug142) | palmitoyltransferase Erf2 | 89.3 |
| mug109 | Rab GTPase binding protein upregulated in meiosis II (predicted) | 91.6 |
| mde5 (meu30) | alpha-amylase homolog Mde5 | 92.4 |
| pdh1 | Golgi to ER retrograde transport protein (predicted) | 93.3 |
| eng2 | cell wall and ascospore endo-1,3-beta-glucanase Eng2 | 93.6 |
| mpf1 | meiotic pumilio family RNA-binding protein Mpf1 (predicted) | 94.1 |
| mok14 | alpha-1,4-glucan synthase Mok14 | 99.0 |
| mug113 | GIY-YIGT nuclease superfamily protein | 99.0 |
| ndk1 | nucleoside diphosphate kinase Ndk1 | 99.0 |
| SPAC977.06 | S. pombe specific DUF999 family protein 3 | unable to sporulate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Teraguchi, E.; Imada, K.; Tahara, Y.O.; Nakamura, S.; Miyata, M.; Kagiwada, S.; Nakamura, T. The Fission Yeast RNA-Binding Protein Meu5 Is Involved in Outer Forespore Membrane Breakdown during Spore Formation. J. Fungi 2020, 6, 284. https://doi.org/10.3390/jof6040284
Zhang B, Teraguchi E, Imada K, Tahara YO, Nakamura S, Miyata M, Kagiwada S, Nakamura T. The Fission Yeast RNA-Binding Protein Meu5 Is Involved in Outer Forespore Membrane Breakdown during Spore Formation. Journal of Fungi. 2020; 6(4):284. https://doi.org/10.3390/jof6040284
Chicago/Turabian StyleZhang, Bowen, Erika Teraguchi, Kazuki Imada, Yuhei O. Tahara, Shuko Nakamura, Makoto Miyata, Satoshi Kagiwada, and Taro Nakamura. 2020. "The Fission Yeast RNA-Binding Protein Meu5 Is Involved in Outer Forespore Membrane Breakdown during Spore Formation" Journal of Fungi 6, no. 4: 284. https://doi.org/10.3390/jof6040284