
Active Methanotrophs and Their Response to Temperature in Marine Environments: An Experimental Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sediment Source
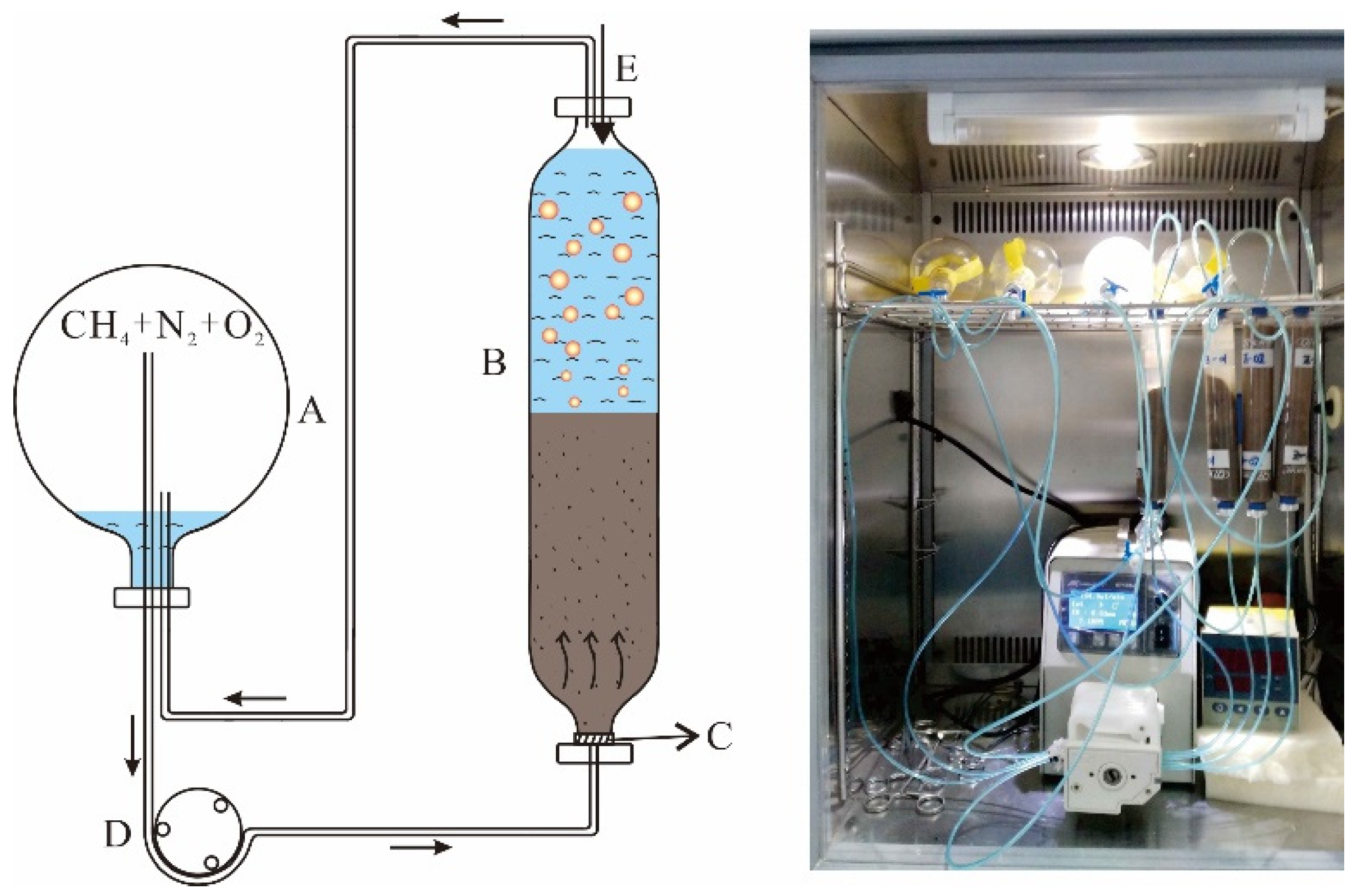
2.2. Sediment Incubations
2.3. Methane Oxidation Measurement
2.4. DNA Extraction
2.5. PCR Amplification, Illumina Miseq Sequencing, and Data Processing
2.6. Quantitative Real-Time PCR Assays
2.7. Statistical Analyses and Temperature Sensitivity Calculations
3. Results
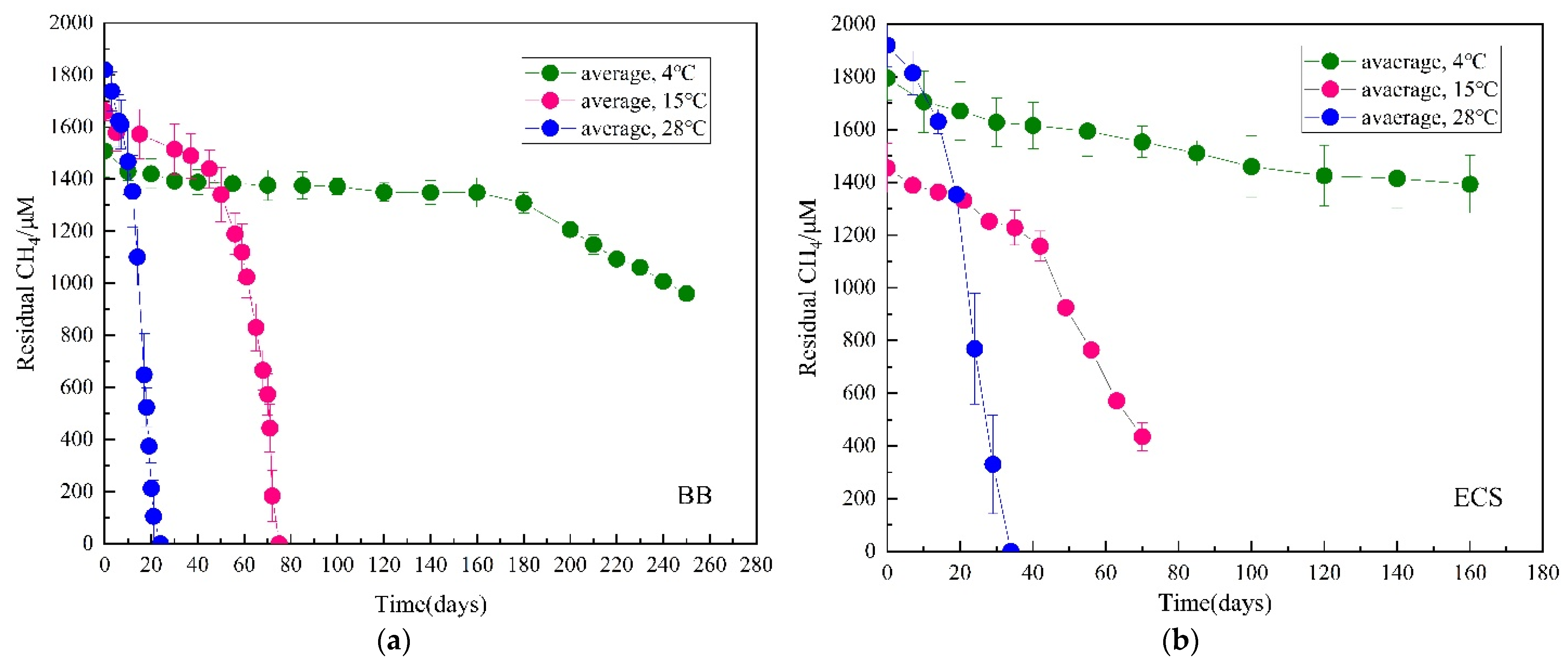
3.1. Methane Consumption
3.2. Methanotrophic Community Structure
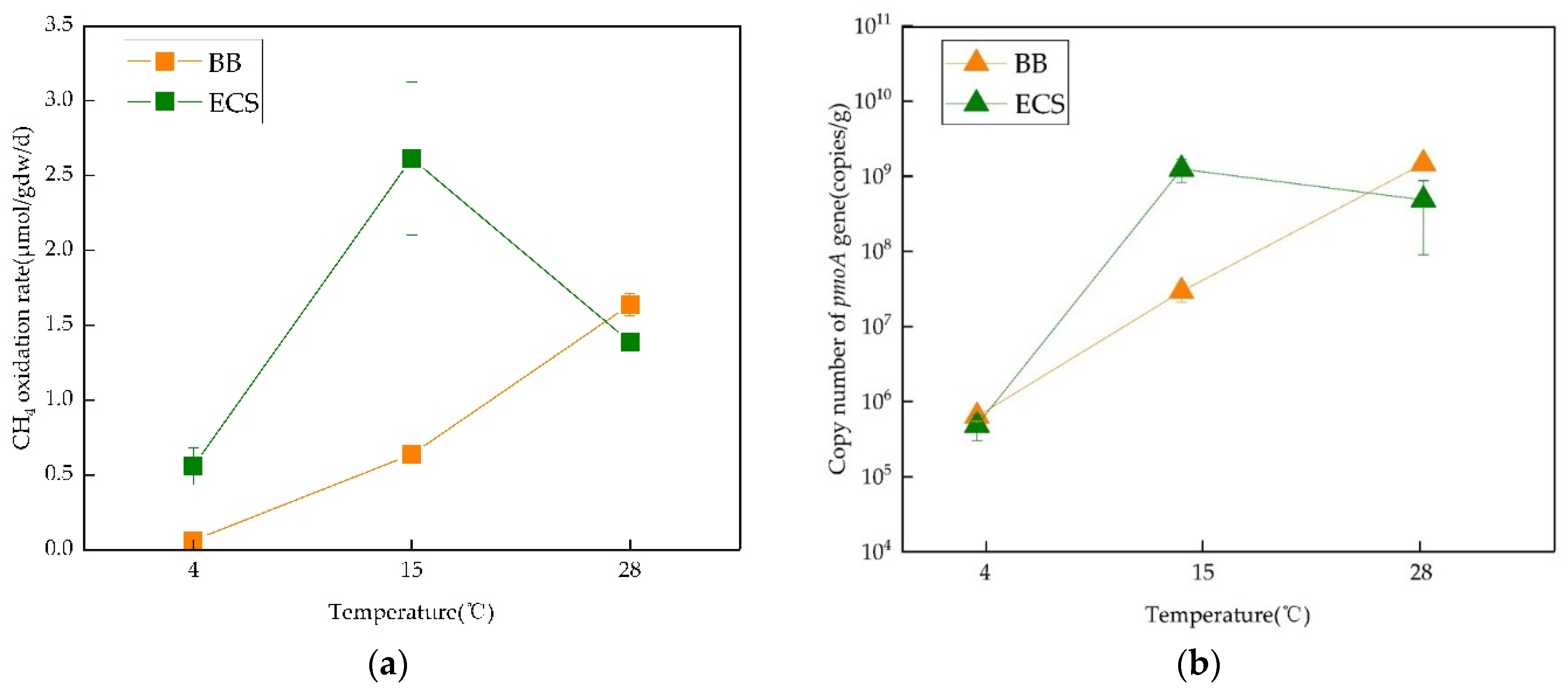
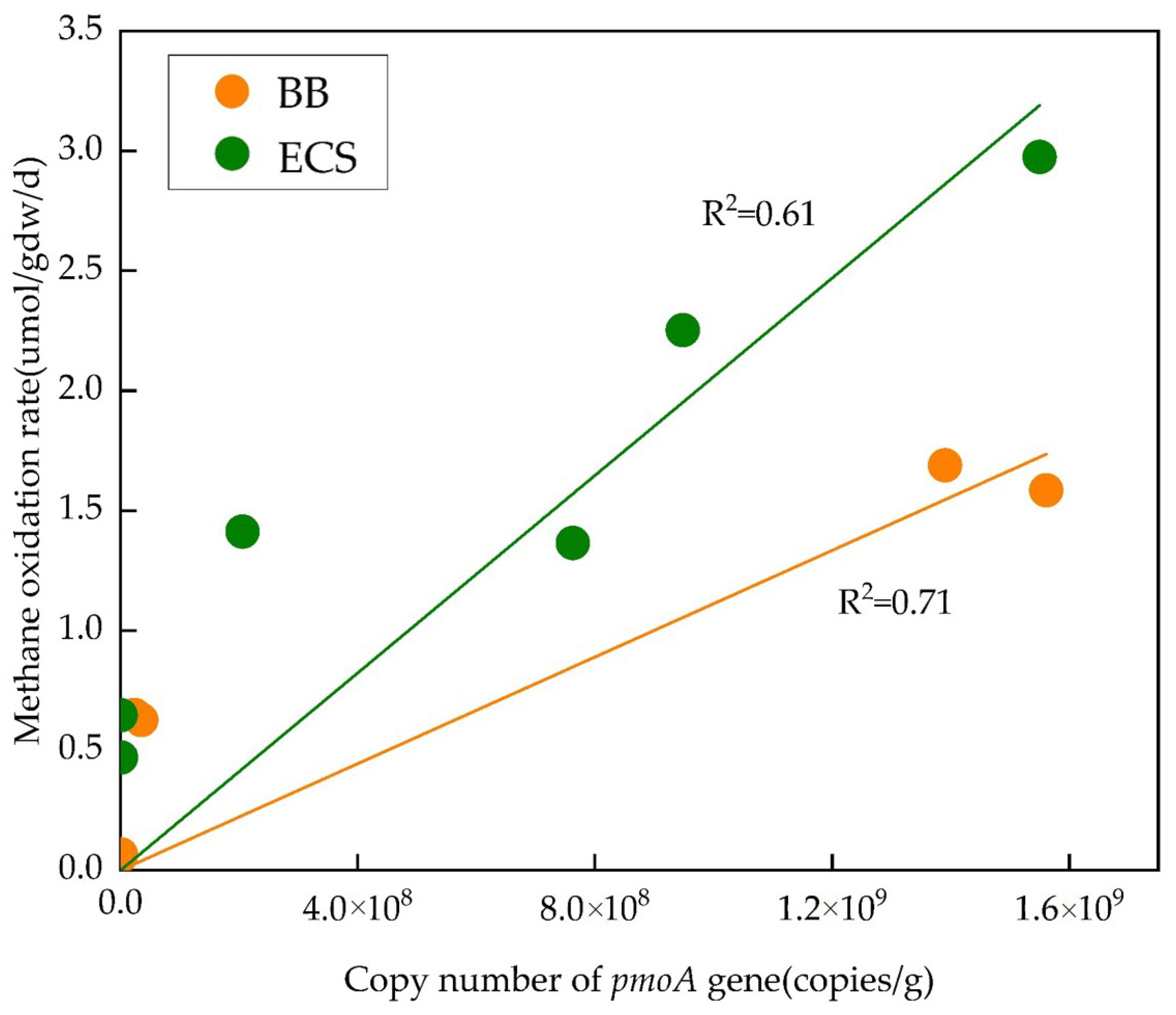
3.3. Abundance of Methanotrophic Population
4. Discussion
4.1. Temperature Effect on CH4 Oxidation Rate
4.2. Temperature Effect on Methanotrophic Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kushwaha, O.S.; Meshram, S.B.; Bhattacharjee, G.; Kumar, R. Molecular Insights about Gas Hydrate Formation. In Advances in Spectroscopy: Molecules to Materials; Springer: Berlin/Heidelberg, Germany, 2019; pp. 311–322. [Google Scholar]
- Lu, X.; Zhang, X.; Sun, F.; Wang, S.; Liu, L.; Liu, C. Experimental Study on the Shear Band of Methane Hydrate-Bearing Sediment. J. Mar. Sci. Eng. 2021, 9, 1158. [Google Scholar] [CrossRef]
- Bei, K.; Xu, T.; Shang, S.; Wei, Z.; Yuan, Y.; Tian, H. Numerical modeling of gas migration and hydrate formation in heterogeneous marine sediments. J. Mar. Sci. Eng. 2019, 7, 348. [Google Scholar] [CrossRef] [Green Version]
- Etiope, G. Natural Gas Seepage; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Yang, H.; Yu, S.; Lu, H. Iron-Coupled Anaerobic Oxidation of Methane in Marine Sediments: A Review. J. Mar. Sci. Eng. 2021, 9, 875. [Google Scholar] [CrossRef]
- Hanson, R.S.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [CrossRef]
- Shukla, P.N.; Pandey, K.; Mishra, V.K. Environmental determinants of soil methane oxidation and methanotrophs. Crit. Rev. Environ. Sci. Technol. 2013, 43, 1945–2011. [Google Scholar] [CrossRef]
- Haque, M.F.U.; Xu, H.-J.; Murrell, J.C.; Crombie, A. Facultative methanotrophs-diversity, genetics, molecular ecology and biotechnological potential: A mini-review. Microbiology 2020, 166, 894–908. [Google Scholar] [CrossRef] [PubMed]
- Kizilova, A.; Yurkov, A.; Kravchenko, I. Aerobic methanotrophs in natural and agricultural soils of European Russia. Diversity 2013, 5, 541–556. [Google Scholar] [CrossRef]
- Chowdhury, T.R.; Dick, R.P. Ecology of aerobic methanotrophs in controlling methane fluxes from wetlands. Appl. Soil Ecol. 2013, 65, 8–22. [Google Scholar] [CrossRef]
- Lehtoranta, K.; Koponen, P.; Vesala, H.; Kallinen, K.; Maunula, T. Performance and Regeneration of Methane Oxidation Catalyst for LNG Ships. J. Mar. Sci. Eng. 2021, 9, 111. [Google Scholar] [CrossRef]
- Wei, Z.; Xu, T.; Shang, S.; Tian, H.; Cao, Y.; Wang, J.; Shi, Z.; Liu, X. Laboratory Experimental Study on the Formation of Authigenic Carbonates Induced by Microbes in Marine Sediments. J. Mar. Sci. Eng. 2021, 9, 479. [Google Scholar] [CrossRef]
- Brindha, R.; Vasudevan, N. Methane oxidation capacity of methanotrophs isolated from different soil ecosystems. Int. J. Environ. Sci. Technol. 2018, 15, 1931–1940. [Google Scholar] [CrossRef]
- He, R.; Wooller, M.J.; Pohlman, J.W.; Quensen, J.; Tiedje, J.M.; Leigh, M.B. Shifts in identity and activity of methanotrophs in arctic lake sediments in response to temperature changes. Appl. Environ. Microbiol. 2012, 78, 4715–4723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Dumont, M.G.; Bodelier, P.L.; Adams, J.M.; He, D.; Chu, H. DNA stable-isotope probing highlights the effects of temperature on functionally active methanotrophs in natural wetlands. Soil Biol. Biochem. 2020, 149, 107954. [Google Scholar] [CrossRef]
- Praeg, N.; Wagner, A.O.; Illmer, P. Plant species, temperature, and bedrock affect net methane flux out of grassland and forest soils. Plant Soil 2017, 410, 193–206. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, S.R.; Bodelier, P.L.; Conrad, R. Effect of temperature on composition of the methanotrophic community in rice field and forest soil. FEMS Microbiol. Ecol. 2007, 62, 24–31. [Google Scholar] [CrossRef]
- Mau, S.; Heintz, M.B.; Valentine, D.L. Quantification of CH4 loss and transport in dissolved plumes of the Santa Barbara Channel, California. Cont. Shelf Res. 2012, 32, 110–120. [Google Scholar] [CrossRef]
- Li, J.; Liu, C.; He, X.; Santosh, M.; Hu, G.; Sun, Z.; Li, Y.; Meng, Q.; Ning, F. Aerobic microbial oxidation of hydrocarbon gases: Implications for oil and gas exploration. Mar. Pet. Geol. 2019, 103, 76–86. [Google Scholar] [CrossRef]
- Kawagucci, S.; Matsui, Y.; Makabe, A.; Fukuba, T.; Onishi, Y.; Nunoura, T.; Yokokawa, T. Hydrogen and carbon isotope fractionation factors of aerobic methane oxidation in deep-sea water. Biogeosciences 2021, 18, 5351–5362. [Google Scholar] [CrossRef]
- Chan, E.W.; Shiller, A.M.; Joung, D.J.; Arrington, E.C.; Valentine, D.L.; Redmond, M.C.; Breier, J.A.; Socolofsky, S.A.; Kessler, J.D. Investigations of aerobic methane oxidation in two marine seep environments: Part 1—Chemical kinetics. J. Geophys. Res. Oceans 2019, 124, 8852–8868. [Google Scholar] [CrossRef]
- Li, M.; Jain, S.; Baker, B.J.; Taylor, C.; Dick, G.J. Novel hydrocarbon monooxygenase genes in the metatranscriptome of a natural deep-sea hydrocarbon plume. Environ. Microbiol. 2014, 16, 60–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felden, J.; Lichtschlag, A.; Wenzhöfer, F.; Beer, D.d.; Feseker, T.; Pop Ristova, P.; Lange, G.d.; Boetius, A. Limitations of microbial hydrocarbon degradation at the Amon mud volcano (Nile deep-sea fan). Biogeosciences 2013, 10, 3269–3283. [Google Scholar] [CrossRef] [Green Version]
- Amos, C.L.; Al-Rashidi, T.B.; Rakha, K.; El-Gamily, H.; Nicholls, R. Sea surface temperature trends in the coastal ocean. Curr. Dev. Oceanogr. 2013, 6, 1–13. [Google Scholar]
- Hasse, L. The sea surface temperature deviation and the heat flow at the sea-air interface. Bound. -Layer Meteorol. 1971, 1, 368–379. [Google Scholar] [CrossRef]
- Doblin, M.A.; Van Sebille, E. Drift in ocean currents impacts intergenerational microbial exposure to temperature. Proc. Natl. Acad. Sci. USA 2016, 113, 5700–5705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Z.; Zhao, T.; Zhang, L.; Gao, Y.; Liu, S.; Yang, X. Effects of copper on expression of methane monooxygenases, trichloroethylene degradation, and community structure in methanotrophic consortia. Eng. Life Sci. 2018, 18, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Einola, J.-K.M.; Kettunen, R.H.; Rintala, J.A. Responses of methane oxidation to temperature and water content in cover soil of a boreal landfill. Soil Biol. Biochem. 2007, 39, 1156–1164. [Google Scholar] [CrossRef]
- Duc, N.T.; Crill, P.; Bastviken, D. Implications of temperature and sediment characteristics on methane formation and oxidation in lake sediments. Biogeochemistry 2010, 100, 185–196. [Google Scholar] [CrossRef]
- Houghton, K.M.; Stewart, L.C. Temperature-gradient incubation isolates multiple competitive species from a single environmental sample. Access Microbiol. 2020, 2, acmi000081. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Tian, J.; Chen, H.; Wu, N.; Yan, Z.; Du, L.; Shen, Y.; Wang, X. Changes in methane oxidation ability and methanotrophic community composition across different climatic zones. J. Soils Sediments 2019, 19, 533–543. [Google Scholar] [CrossRef]
- Xu, C.; Wu, N.; Sun, Z.; Zhang, X.; Geng, W.; Cao, H.; Wang, L.; Zhang, X.; Xu, G. Methane seepage inferred from pore water geochemistry in shallow sediments in the western slope of the Mid-Okinawa Trough. Mar. Pet. Geol. 2018, 98, 306–315. [Google Scholar] [CrossRef]
- Zang, K.; Zhang, G.; Wang, J. Methane emissions from oil and gas platforms in the Bohai Sea, China. Environ. Pollut. 2020, 263, 114486. [Google Scholar] [CrossRef]
- Zhang, N. Geochemical of Hydrocarbon Gas from the Surface Seabed Sediment in Bohai Bay Basin. Ph.D. Thesis, Jilin University, Changchun, China, 2016. [Google Scholar]
- Li, J.; Liu, C.; Wu, N.; He, X.; Hao, X.; Li, F.; Chen, Y.; Meng, Q. The Variation of Microbial (Methanotroph) Communities in Marine Sediments Due to Aerobic Oxidation of Hydrocarbons. J. Ocean Univ. China 2021, 20, 553–561. [Google Scholar] [CrossRef]
- Zang, K. Seasonal Variations and Regulatory Mechanisms of Dissolved Methane Concentration and Its Sea-to-Air Fluxes in the Seasonal Oxygen Deficient Zones in Bohai Sea. Ph.D. Thesis, Chinese Academy of Meteorological Sciences, Beijing, China, 2018. [Google Scholar]
- Chen, Y.-F.; Zheng, X.-L.; Jing, L.; He, X.-l.; Liu, C.-L.; Meng, Q.-G.; Qin, D.-D.; Zhang, P.-Y. Study on Oxidation Rate and Isotope Fractionation of Methane in Bohai Sea Sediments. Rock Miner. Anal. 2018, 37, 164–174. [Google Scholar]
- Gebert, J.; Groengroeft, A.; Miehlich, G. Kinetics of microbial landfill methane oxidation in biofilters. Waste Manag. 2003, 23, 609–619. [Google Scholar] [CrossRef]
- Pérez, R.; Cantera, S.; Bordel, S.; García-Encina, P.A.; Muñoz, R. The effect of temperature during culture enrichment on methanotrophic polyhydroxyalkanoate production. Int. Biodeterior. Biodegrad. 2019, 140, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.R.; Rai, R.K.; Green, S.J.; Chetri, J.K. Effect of temperature on methane oxidation and community composition in landfill cover soil. J. Ind. Microbiol. Biotechnol. 2019, 46, 1283–1295. [Google Scholar] [CrossRef]
- Dunfield, P.; Dumont, R.; Moore, T.R. Methane production and consumption in temperate and subarctic peat soils: Response to temperature and pH. Soil Biol. Biochem. 1993, 25, 321–326. [Google Scholar] [CrossRef]
- Segers, R. Methane production and methane consumption: A review of processes underlying wetland methane fluxes. Biogeochemistry 1998, 41, 23–51. [Google Scholar] [CrossRef]
- Scheutz, C.; Kjeldsen, P. Environmental factors influencing attenuation of methane and hydrochlorofluorocarbons in landfill cover soils. J. Environ. Qual. 2004, 33, 72–79. [Google Scholar] [CrossRef]
- Omelchenko, M.; Vasileva, L.L.; Khmelenina, V.; Trotsenko, Y.A. Pathways of primary and intermediate metabolism in a psychrophilic methanotroph. Microbiology 1993, 62, 509–512. [Google Scholar]
- Boetius, A.; Wenzhöfer, F. Seafloor oxygen consumption fuelled by methane from cold seeps. Nat. Geosci. 2013, 6, 725–734. [Google Scholar] [CrossRef]
- Ruff, S.E.; Felden, J.; Gruber-Vodicka, H.R.; Marcon, Y.; Knittel, K.; Ramette, A.; Boetius, A. In situ development of a methanotrophic microbiome in deep-sea sediments. ISME J. 2019, 13, 197–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshkin, I.Y.; Belova, S.E.; Danilova, O.V.; Miroshnikov, K.K.; Rijpstra, W.I.C.; Damsté, J.S.S.; Liesack, W.; Dedysh, S.N. Methylovulum psychrotolerans sp. nov., a cold-adapted methanotroph from low-temperature terrestrial environments, and emended description of the genus Methylovulum. Int. J. Syst. Evol. Microbiol. 2016, 66, 2417–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Water Depth (m) | Bottom Water Temperature 1 (°C) | Water Content | TOC | Median Grain Size (μm) | CH4 Concentration 2 (μL/L) | pH | ORP 3/mV |
|---|---|---|---|---|---|---|---|---|
| BB | 23 | 6.3–28.6 | 39.2% | 0.66 | 6.0 | 10–167 | 8.7 | −a28 |
| ECS | 979 | ~4.0 | 46.3% | 1.51 | 18.5 | 33 | 8.3 | - |
| Ecosystem | Temperature | CH4 Oxidation Rate 1 (R2 2) | Q10 Values | One-Way ANOVA 3 | Source | ||
|---|---|---|---|---|---|---|---|
| 4–15 °C | 15–28 °C | d.f4 | p | ||||
| BB | 4 °C | 0.06 (0.97–0.99) | 8.3 | 2.1 | 2 | 0.007 | This paper |
| 15 °C | 0.64 (0.89–0.91) | ||||||
| 28 °C | 1.64 (0.95–0.98) | ||||||
| ECS | 4 °C | 0.56 (0.92) | 4.1 | 0.6 | 2 | 0.009 | |
| 15 °C | 2.61 (0.97–0.99) | ||||||
| 28 °C | 1.39 (0.96–0.99) | ||||||
| Eight lakes in central Sweden | 4–30 °C | 0.01–0.63 (0.83–1.00) | 0.5–2.4 | - | - | [29] | |
| Two forest zones | 10–25 °C | <0.0002 (-) | 0.2–1.9 | - | - | [31] | |
| Zion landfill site | 6–40 °C | 0.85–4.84 (-) | 1.4–3.3 | - | <0.004 | [40] | |
| Ecosystem | Temperature | Type of Active Methanotrophs | Initial Abundance of pmoA Genes | Average Abundance of pmoA Genes | One-Way ANOVA 2 | Source | |
|---|---|---|---|---|---|---|---|
| d.f3 | p | ||||||
| BB | 4 °C | unclassified_Methylococcaceae | 4.18 × 104 | 6.35 × 105 | 2 | 0.135 | This paper |
| 15 °C | unclassified_Methylococcaceae Methylobacter | 2.94 × 107 | |||||
| 28 °C | 1.48 × 109 | ||||||
| ECS | 4 °C | unclassified_Methylococcaceae Methylobacter | 4.78 × 104 | 4.86 × 105 | 2 | 0.046 | |
| 15 °C | 1.25 × 109 | ||||||
| 28 °C | unclassified_Methylococcaceae | 4.84 × 108 | |||||
| Arctic lake | 4–21 °C | Methylobacter; Methylophilus; Methylomonas; Methylophilus | - | - | - | - | [14] |
| Two forest zones | 10–25 °C | Methylobacter; Methylocaldum; Methylomicrobium; Methylocystis | - | - | - | - | [31] |
| Zion landfill site | 6–40 °C | Methylobacter; Methylocystis; Methylocaldum; Methylotenera | - | - | - | - | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Xu, X.; Liu, C.; Wu, N.; Sun, Z.; He, X.; Chen, Y. Active Methanotrophs and Their Response to Temperature in Marine Environments: An Experimental Study. J. Mar. Sci. Eng. 2021, 9, 1261. https://doi.org/10.3390/jmse9111261
Li J, Xu X, Liu C, Wu N, Sun Z, He X, Chen Y. Active Methanotrophs and Their Response to Temperature in Marine Environments: An Experimental Study. Journal of Marine Science and Engineering. 2021; 9(11):1261. https://doi.org/10.3390/jmse9111261
Chicago/Turabian StyleLi, Jing, Xiaoqing Xu, Changling Liu, Nengyou Wu, Zhilei Sun, Xingliang He, and Ye Chen. 2021. "Active Methanotrophs and Their Response to Temperature in Marine Environments: An Experimental Study" Journal of Marine Science and Engineering 9, no. 11: 1261. https://doi.org/10.3390/jmse9111261