
Sponges as Emerging By-Product of Integrated Multitrophic Aquaculture (IMTA)
by
 , , , and
, , , and
Joseba Aguilo-Arce
1,
Pere Ferriol
2,
Roberta Trani
1,* ,
,
Patrizia Puthod
1,
Cataldo Pierri
1 and
Caterina Longo
1
1
Department of Bioscience, Biotechnology and Environment, University of Bari Aldo Moro, Via Orabona 4, 70125 Bari, Italy
2
Interdisciplinary Ecology Group, Department of Biology, University of the Balearic Islands, Carretera de Valldemossa km 7.5, 07122 Palma de Mallorca, Spain
*
Author to whom correspondence should be addressed.
J. Mar. Sci. Eng. 2023, 11(1), 80; https://doi.org/10.3390/jmse11010080
Submission received: 29 November 2022
/
Revised: 29 December 2022
/
Accepted: 30 December 2022
/
Published: 3 January 2023
(This article belongs to the Special Issue New Perspectives in Sustainable Aquaculture)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The use of marine sponges dates back thousands of years, and interest in these animals is increasing as new applications are discovered. Their potential is extensive, both in their ancient and still popular use as bath sponges for cosmetics and regarding the more recent discovery of bioactive secondary metabolites mainly of interest for the pharmaceutical industry and the less developed aquariology. Despite their proven biofiltration and ecosystem restoration ability and the biomass supply problem for the interested industries, few integrated multi-trophic aquaculture (IMTA) systems incorporate these invertebrates in their facilities. Therefore, in this brief review, the benefits that marine sponges could bring to rapidly growing IMTA systems are summarized, highlighting their suitability for a circular blue economy.
1. Introduction
Marine sponges have been targeted by fishermen for thousands of years, including in the Egyptian civilization, and their presence in our society has been continuous ever since [1]. However, the increasing interest in these animals during the last century has entailed the impossibility of providing the arising sponge market with an adequate amount of natural biomass. In this sense, when diving and collecting techniques improved in the Mediterranean area, different sponge species were collected without any specific legislation to supply a worldwide trade demanding dozens of tons of marine sponges, causing a dramatic reduction of natural stocks [2]. With the development of the pharmaceutical industry in the last decades of the 1900s, for example, and the strong demand for sponge biomass for the industrial production of avarol, e.g., (a powerful anti-inflammatory, anti-tumor, and anti-psoriatic) [3,4,5], the systematic collection of Dysidea avara caused a dramatic depletion of natural stocks, particularly along the eastern Mediterranean coasts, to the point of it being considered endangered [2]. In response to this intensive withdrawal, many demosponge species have been included in the lists of species with high conservation value for which specific protectionist policies are recommended (Annex I of the Berne Convention).
On the other hand, the world’s population has tripled in the last seven decades. Contemporaneously, several problems have arisen, one of the most important concerns being food supply [6]. The inability of natural populations to achieve this has accelerated the search for sustainable production alternatives. Fisheries have not been left behind, with aquaculture and its derivative mariculture quadrupling production over the last 3 decades, nearly equalling that of capture fisheries [7]. Still, as in most developing production techniques, there are problems that need to be solved to minimize ecological impact.
Coastal fish farms can have a significant environmental impact due to the release of catabolites by cultured organisms and feed waste, leading to an overall increase in sedimentation, organic matter, and a potentially pathogenic or antibiotic-resistant bacterial load in the water [8]. However, Integrated Multitrophic Aquaculture (IMTA), the practice that combines the cultivation of fed aquaculture species with other organisms able to extract organic and/or inorganic substances from seawater, represents an eco-friendly alternative to monoculture. IMTA allows better water quality and promises high economic return and social suitability, implying an improvement in human welfare and ecosystem services provided by the marine environment.
Among the extractive organisms, economically profitable edible filter-feeders are the most utilized, including the predominant bivalves [9,10]. Nevertheless, Porifera seems to be one of the most promising, although they are still underexploited. Up to now, few sponge species have been tested to assess their rearing suitability and performance, bioactive compound production, or bioremediation capability: on the Mediterranean scale, Geodia cydonium [11], Spongia officinalis and Hippospongia communis [12,13], Dysidea avara [14], Hymeniacidon perlevis [15], Chondrosia reniformis [14,16], Ircinia variabilis [17], and Sarcotragus spinosulus [18]. In these trials, G. cydonium almost doubled its initial weight in six months. D. avara and S. spinosulus doubled their size in four and twelve months, respectively, and C. reniformis achieved a growth rate of 170% in one year, while S. officinalis and H. communis sponges increased their volume by 100–200% over two years. Additionally, impressive growth exceeding 2000% per year was registered in the Southwestern Pacific Ocean with Mycale hentscheli cultured near a mussel farm [19]. Altogether, these impressive results prove the suitability of the rearing system and the species, opening new possibilities in the aquaculture sector.
In this sense, their environmental and economic potential and, thus, the interest in these organisms, is increasing as new sustainable applications arise. This review aims to highlight the suitability and benefits of marine sponges in IMTA systems in a circular blue economy framework by discussing the profitability of these organisms and emphasizing their eco-economic potential. From a literature search conducted in Scopus and Google Scholar databases, the available papers on sponges in IMTA systems were selected to gather current knowledge on this practice, along with relevant literature, providing an informative and comprehensive framework of the status and potential of marine sponges. The results of this bibliographic research are included in the references section of this work.
2. Past and Present Concerns
The trade of sea sponges dates back thousands of years to when they were collected mainly for cosmetic use and known as “bath sponges”. In the last century, sponges could be purchased for USD 30 per individual, and the annual value of imports is estimated to be millions of USD. However, in a span of 50 years, sponge production declined by more than 80% due to unregulated overexploitation, catastrophic diseases, and the introduction of synthetic sponges into the market [20]. Even so, today in the Mediterranean basin, countries such as Italy (e.g., Spugnificio Incorvaia, Gela; Bellini Let’s BE natural, Cogozzo di Viadana; Spugnificio Rosenfeld, Muggia), Greece (e.g., Cipreo J.-Kypreos Natural Sea Sponges, Kalymnos; Gerakios traders of natural sponges, Peristeri; Bioesti, Agios Nikolaos), Croatia (e.g., Spuga2, Kapranj), Cyprus (e.g., Old Port Sea Sponges, Limassol), and Tunisia (e.g., Éponge, Kraten) still maintain this economic sector based on traditional harvesting.
Although attempts to culture sponges have been documented since the late 18th century [21], the lack of knowledge and the failure of experiments made harvesting individuals from natural populations the most efficient and profitable methodology [22]. Still, the aforementioned negative events during the 20th century prompted the search for new business strategies. Thus, by the early 1970s, organized and successful mariculture trials with bathing sponges had already been conducted. The number of attempts has only increased over the years, becoming the second market interest for the culture of these animals [23,24]. In the search for sustainable and profitable production, as reviewed in [25], many studies have tried to define the best rearing system for bath sponges and have concluded it to be strongly dependent upon species and environmental conditions.
In recent years, marine natural products have gained popularity and the demand for sponges, though they are more expensive than those of synthetic origin, is high. However, despite the research effort and promising growth rates of cultured bath sponges (doubling or tripling in size annually) [26,27,28,29,30], few countries currently cultivate them for commercial purposes, where they are mostly sold in the local markets, e.g., Micronesia [31] and Zanzibar [32]. Additionally, few studies have been carried out concerning bath sponges within an IMTA system [12,13], but everything remains in the theoretical framework since, to date, there is no such production method at a commercial level. Incorporating bath sponges in these environments where bacterial load is high involves zero cost of consumables like food, while the material for their cultivation would barely reach a few hundred USD [30]. Since their maintenance is based on monthly growth monitoring, bath sponges rearing seems an ideal candidate for promising high economic profitability when looking for a blue circular economy in IMTA systems.
3. Bioremediation, a Way of Life
One of the main advantages of filter-feeding organisms such as Porifera is that their energy source is obtained by the highly efficient filtration of organic particles between 0.1–50 µm, such as dissolved and particulate organic particles, heterotrophic bacteria and eukaryotes, phytoplankton, and even viruses [33,34,35], so that their mere growth leads to bioremediation of the surrounding environment. Therefore, they are considered a possible solution to reduce eutrophication and bacterial load.
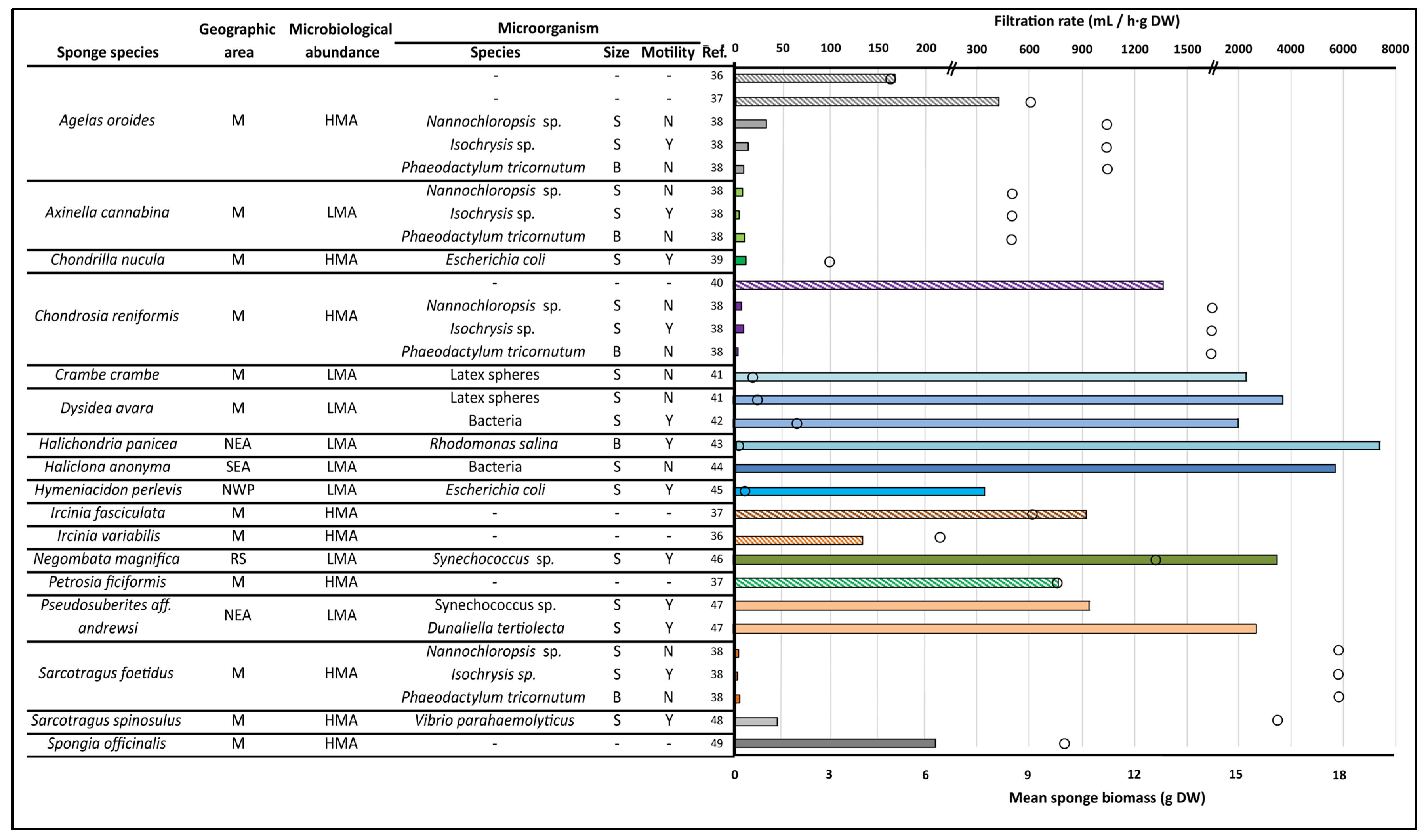
In this sense, numerous works have tried to calculate this potential by employing experiments on the filtration and retention capacity of sponges. When it comes to those performed ex situ, controlled conditions reduce the variability of factors, such as temperature, salinity, nutrients, or microorganisms used, which, together with visualization techniques and cell counting (such as spectrometry or cytometry), have served to prove that this capacity differs significantly depending on the species and size of the sponge explant or the size and even the motility of the microorganism used (Figure 1).
However, the logistics and maintenance of conditions involve an expense that does not exist in IMTA systems, being the natural environment of the sponges. In addition, variables such as season, currents, or depth are excluded in ex situ experiments, despite being proven to affect the growth, survival, morphology, composition, and pumping activity of some sponges [39,40,53,54,55,56]. Although the remediation and conversion of organic carbon have been studied (references already reviewed in [57]), little work has focused on in situ filtration capacity (e.g., [36,37,40]) and even less on IMTA systems, where knowledge is very limited [58,59]. Despite that, it has been shown that carbon uptake in these animals may exceed that of commercial species, such as bivalves, whose market is well established and generates large economic benefits while that of sponges is still under development [60].
Similarly, being filter-feeding organisms, they are constantly exposed to contamination of the water column. More specifically, since the last century, numerous experiments have been carried out to test the accumulation capacity and levels of pollutants, such as heavy metals (HM, related to diseases such as autism, Alzheimer, or diabetes mellitus) [61] from mercury or copper to heavier metals such as uranium, americium, or rare earth elements, which in some cases have been seen to be higher than in mussels [62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83]. As with filtration, it has been shown that this ability depends on the sponge species and the contaminant tested, including the morphology of the individual, and that, although spicules represent the main component of the biomass, they accumulate mainly in the organic tissue [84,85]. Additionally, they could potentially consume contaminated bacteria and incorporate their HMs [86] and are being proposed as a source of biomagnification of these compounds in higher trophic levels [87]. However, subjecting sponges to elevated HM concentrations has been found to negatively affect growth, survival, fecundity, and filtering ability and to cause DNA damage, ROS production increase, alterations in macromolecule composition, or even apoptosis, suggesting them as bioremediators of environments partially exposed to these contaminants [64,67,68,69,88,89,90,91,92].
The same is true for organic pollutants (OP), whose accumulation in sponges and biomagnification has also been demonstrated, including natural compounds that have been classified as such due to their possible toxic effects [93,94,95,96,97,98]. In this regard, there is a diversity of results according to contaminants and sponge species. While some results state that the accumulation of some polycyclic aromatic hydrocarbons (PAHs) and polychlorinated biphenyls (PCBs) is higher than in some commercial oysters and mussels, others have found similar levels [72,76,94,99]. However, as expected with pollutant compounds, also OPs induce stress, DNA damage, and cell apoptosis in sponges [93,100,101].
On the other hand, microplastics (MP) represent a threat that has been gaining importance for the past decade, despite having been found in sponge samples dated more than 20 years ago [102,103]. Therefore, work and knowledge on the accumulation of these contaminants and their derivatives in these animals are scarce [104,105,106,107,108]. As might be expected, a higher concentration of MPs has been seen in filter feeders (similar between sponges and mussels) than in other feeding strategies, and in the case of sponges, a possible preference for very small fibers and particles as well [102,104]. While no negative effect has also been reported [106,109], a more recent study demonstrates that the uptake of some of these contaminants reduces filtration and respiration rates [110]. However, being that the levels of MPs in the surrounding water are higher than in their tissues, these studies agree and suggest a possible resistance or ejection mechanism that allows them to thrive in contaminated waters [106,110].
When talking about remediation, and considering the concept of the sponge as a holobiont, the capacity of the associated microbiota to resist, accumulate, and degrade contaminants cannot be disregarded. In this sense, recent works have demonstrated the HM accumulation capacity of some symbionts and identified genes that grant resistance to them, which could have been transmitted via plasmids [111,112,113,114,115,116,117]. However, not all these microorganisms appear to possess them, and, although no differences have also been reported [118,119], the accumulation of HMs in the host has been shown to negatively affect symbiont communities [79,120]. The usefulness of these microorganisms does not stop there. They have also been proposed to biotransform and can produce biosurfactants for the bioremediation of these metals [121,122,123,124,125,126], for the degradation of MPs [127], and of many OPs, such as pyrenes, brominated phenols, steroids, crude oil, dyes used in the textile industry, or even alkylbenzene sulfonates present in detergents [96,128,129,130,131,132,133,134,135,136], which increases the interest in and confirms sponges as a hot-spot for the search of new green alternatives in this field.
All in all, the works cited above define marine sponges as good biofilters and bioindicators of pollution caused by increased bacterial load and organic and inorganic contaminants due to their ability to filter and accumulate them. In search of new remediation technologies, these animals have even inspired the creation of synthetic sponges to treat water [137,138,139]. Therefore, incorporating them into the production systems of species for human consumption, such as IMTAs, implies the possibility of monitoring and improving water quality and, thanks to the demonstrated ability to reduce the pathogen bacterial load, a possible reduction in antibiotic costs.
4. A Marine Bioactive Compound Factory
The first drugs of marine origin reaching clinical trials were extracted from a Caribbean sponge around seven decades ago [140]; since then, the number of these compounds has only increased, and, together with cnidarians, they are the most prominent organisms discovered [141]. By November 2022, nearly 12,000 compounds were reported from the phylum Porifera (exceeding a quarter of the total of marine origin), more than 200 of which were described this year (Figure 2). These are synthesized not only by sponges but also by their symbiont microorganisms. The value of the approved and artificially synthesized ones can reach from hundreds to thousands of USD [142,143], demonstrating the potential of profitable natural production carried out by these organisms.
The different sponge species and their micro inhabitants open the possibility to search for and characterize a great number of biomolecules, with different structures and activities that review works have already tried to collect and synthesize, e.g., [141,143,144,145,146]. In a very generalized way and without detailing, they have a wide range of applications: from anti-tumour, antiviral, and antimicrobials in human medicine to antimicrobial and antifouling activities in the aquaculture industry or biomaterial production in different sectors. However, obtaining the needed biomass involves a considerable challenge, as natural populations cannot supply this market sustainably.
Therefore, as with bath sponges, attempts to culture them for this purpose have increased in recent years to become the main commercial target of these activities [24]. However, few attempts have been carried out in IMTA systems, and some of these are cultures close to other fisheries facilities [11,14,16,18,19,147,148,149]. Considering the importance of symbionts in the synthesis of bioactive compounds, the conditions to ensure their production should be optimal not only for the sponge but also for their inhabitants, which ex situ can be costly and difficult to maintain. Therefore, co-culture strategies such as IMTA appear to be possible and economically profitable solutions, as from reared sponge biomass, as well as others for the pharmaceutical industry, antifouling and antibiotic compounds could be obtained that would reduce the annual losses of billions for fisheries [150], fostering a circular economy of the sector.
5. From the Sea to the Tank
With hundreds of aquariums worldwide and millions of recreational ones, another economic potential of marine sponges is their sale for this activity, as due to their diverse colors and shapes they are particularly attractive (Figure 3). There are companies dedicated to their trade (mainly in the US), and their prices vary from tens to hundreds of USD per individual, depending on species and size (e.g., Marine World Aquatics Ltd., Bradford, UK; RUSALTY, St Okeechobee, FL, USA; Aquarium Creations Online, Lauderdale Lakes, FL, USA).
In addition, there are fish families such as angelfish (Pomacanthidae), filefish (Monacanthidae), boxfish (Ostraciidae), or pufferfish (Tetraodontidae) that graze on different sponge species and can contribute to over 70% of their diet [151,152,153]. As it is used as an aquarium species, sponge-based food supplies have already been developed (e.g., Mega-Marine Angel, Hikari; Sponge Professional Softgran, EBO; Angel Formula, Ocean Nutrition).
Considering that some of the sponges consumed by these fish have already been the target of in situ culture attempts (albeit if for a different purpose, such as Agelas sp., Callyspongia spp., Chondrilla sp., Chondrosia sp., Geodia sp., Haliclona sp., Ircinia spp., Mycale spp., Spongia spp., or Xestospongia sp.) [11,13,14,16,26,27,30,36,53,147,148,149,154,155,156,157,158,159,160], incorporating them into IMTA systems would be a viable option for the production of aquarium feed supplements. On the other hand, physical traits such as color, shape, or size are vital when selecting ornamental aquarium sponges. Depending on the geographic position, these facilities could provide a suitable environment for the correct growth and sustainable production of local ornamental aquarium species.
6. The Challenge in Marine Sponges’ Restoration
As many sponge species are considered endangered, efforts to recover natural populations are increasing (Annex I of the Berne Convention). From artificial reefs to vertical collectors, ideas are emerging for different restoration structures to increase biodiversity and create a biotope for these species [161]. Transplanting sponges to areas where they had been almost eradicated has been seen as a viable option to recover the populations of these animals, with 100% survival rates and maintaining fertile individuals to favor the natural recruitment of juveniles and other organisms, such as corals, would improve ecological succession [162,163].
In this sense, wisely planned IMTA systems could be reasonably proposed for environmental restoration and conservation purposes. Such facilities combine bioremediation with biodiversity increase thanks to the fouling attached to artificial substrates and reared specimens (Figure 4). In the case of sponges, the biomass obtained can be valorized for transplantation or restocking of threatened or vulnerable species up to benthic habitat restoration. In addition to the environmental value, the creation of these ecosystems and the indirect recruitment of other species could represent a benchmark of underwater tourism that, at zero cost, would bring income to the local community [161].
7. Deciding the Methodological Approach
Planning any sponge farm (including those in an IMTA system) requires in-depth preliminary studies to first evaluate the consistency of the natural stocks which will act as “donors”. Once the distribution and abundance of the sponge donor beds have been characterized, it is necessary to study thoroughly the biological and ecological characteristics of the potentially selected species. Among the features that must be considered in the species selection, expertise in reproduction, growth performance, resistance to survival in critical conditions, and resistance to manipulation are fundamental.
Going into more detail on the methodology and practical aspects of sponge culture, the design of the systems and the survival and growth rates of the explants used vary according to the sponge species, their skeletal consistency, and the aim of production. Thus, for secondary metabolite production, the shape and appearance of the individual are taken less into consideration, whereas maturity or symbiotic composition are. At the same time, the opposite would be true if the cultured individuals were to be destined for an aquarium.
In this sense, sponge farming systems could be performed vertically in the water column or horizontally on the seabed using different materials and tools. Mobile structures suspended in the water column allow the rearing of many sponge explants around the fish farming cages. In such systems, different tools have been tested, such as ropes, tubular nets, mesh panels, lantern nets, and SEAPA cages where the sponge explants have been inserted (Figure 5). [24] reviews the methodology of studies that have cultured sponges in situ and concludes that artificial substrates are the predominant choice, followed by rope and mesh systems, all with similar percentages (between 25% and 30% of the studies reviewed for each). However, when the search is narrowed down to IMTA systems (present study), those with thread lines running through explants and mesh systems become the most studied (encompassing almost all the studies), while artificial substrates are hardly used [16]. For Dysidea avara, for example, Ref. [164] concludes that in pristine natural waters a cage system promotes a higher growth rate when compared with rope and mesh systems, while [14] and the Porifarma company described in [25], being fish-farming associated sponge rearing systems, utilize nylon rope systems with promising results. This change in the choice of methodology could be mainly because, in IMTA systems, the organic load in the water column coming from fish production is high and increases sedimentation [8], which has been shown to negatively affect sponge culture structures with large exposed or semi-enclosed surfaces, such as artificial substrates or cages [14]. Therefore, designs in which more of the sponge surface area is in contact with the water column and less is available for sedimentation and fouling colonization seem to be the most suitable for explant rearing, such as the rope and mesh system variants.
However, when choosing the type of culture system, other factors must also be considered, which in some cases determine the biomass production yield or its usefulness for the established objective. Depth, for example, is a determining factor for collagen production in Chondrosia reniformis (higher content in shallow waters) [40]. Consequently, culture structures should be located a few meters from the surface. Heat stress, on the other hand, induces the loss of symbionts [165], which can be crucial to produce secondary metabolites; thus, in areas where the water surface temperature rises above 30 °C, the culture should be placed at greater depths. All in all, the selection of the cultivation method for marine sponge explants is the result of different factors, such as environmental factors, species, or production objectives, that will determine the type of system, material, and placement of the structures.
8. Fitting Strategic and Sustainable Development Goals
The European Union, in the amended 2022 “New strategic guidelines for aquaculture”, improved the “Strategic Guidelines for a more sustainable and competitive European aquaculture for the period from 2021 to 2030” with a shared vision for further development of aquaculture in the EU. The goal is to build a sustainable, resilient, and competitive aquaculture (marine and freshwater). Particular attention is paid to the need to put science into practice through applied research and innovation, nationally and internationally. Among the strategic points that member states must consider to achieve the proposed goal by 2030 are supporting the development of new aquaculture methods, in particular those with low environmental impact (e.g., IMTA), and promoting diversified production to expand the supply of aquaculture products, using promising new species in the EU, including marine invertebrates.
From a global point of view and with a focus on future feasible socio-economic development, the use of marine sponges in IMTA systems fits perfectly with the Sustainable Development Goals for 2030 described by the United Nations [166], which detail objectives, such as reducing biodiversity loss and poverty, making sustainable use of marine resources, and promoting environmentally sustainable economic development, among others. Although the inclusion of sponges in IMTA is still underutilized, keeping avoiding the over-exploitation of natural sponge populations in mind, the spreading of such practices can combine environmental needs and sustainable development [24,25,57].
As mentioned above, sponge mariculture has been known for a long time but with only a few large-scale applications, mainly related to “bath sponges” [24,25,57]. Currently, a positive impact has been reported in regions such as Zanzibar and Micronesia, where in situ marine sponge culture systems enhance the recruitment of other species and provide a source of income for the local population [31,32]. Thus, despite being underexploited species in IMTA systems, the environmental and economic potentials of marine sponges are important in the pursuit of the fixed goals of the aquaculture sector and sustainable societal development.
9. Conclusions
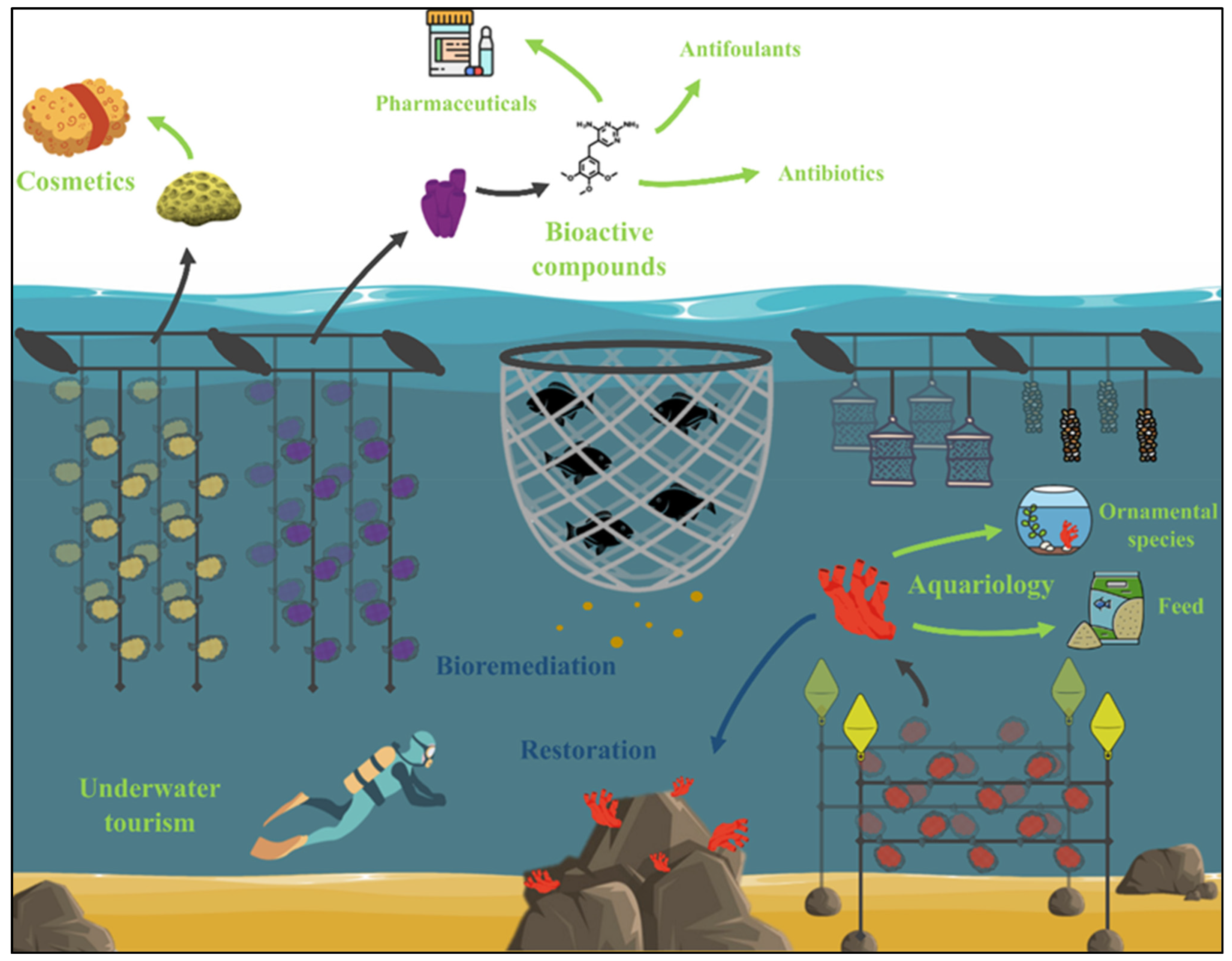
The production of biomass from non-edible species in properly designed IMTAs, such as marine sponges with important profitable applications, implies using new resources that promote a blue circular economy (Figure 6). In this sense, these systems represent an eco-friendly sponge biomass supply not only for economic interests, such as the bath sponge market, green extraction of bioactive molecules, and aquariology (ornamental species or food for fishes) but also for environmental welfare as bioremediation or restoration purposes that increase ecosystem biodiversity. The IMTA systems not only fit perfectly with the general Sustainable Development Goals proposed by the United Nations but, at the same time, lowering the use of antibiotics would reduce costs and economic losses in the pursuit of the aquaculture sector’s environmentally friendly progress.
Author Contributions
Conceptualization, C.L. and J.A.-A.; formal analysis, J.A.-A.; data curation, J.A.-A., R.T., P.P., C.P. and P.F.; writing—original draft preparation, C.L. and J.A.-A.; writing—review and editing, J.A.-A., R.T., P.P., C.P. and P.F.; supervision, C.L.; project administration, C.L.; funding acquisition, C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the European Community, Life Environment funding program: Remedia-Life project (LIFE16 ENV/IT/000343): Remediation of Marine Environment and Development of Innovative Aquaculture: exploitation of Edible/not Edible biomass.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Authors would like to acknowledge the time and effort devoted by reviewers to improve the quality of the manuscript. We sincerely appreciate all valuable comments and suggestions, which helped us to improve the quality of the published manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chaviarà, D. Le spugne e i loro pescatori dai tempi antichi ad ora. Mem. Reg. Com. Talassogr. Ital. 1920, 74, 1–49. [Google Scholar]
- Pronzato, R.; Manconi, R. Mediterranean commercial sponges: Over 5000 years of natural history and cultural heritage. Mar. Ecol. 2008, 29, 146–166. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Zahn, R.K.; Gasić, M.J.; Dogović, N.; Maidhof, A.; Becker, C.; Diehl-Seifert, B.; Eich, E. Avarol, a cytostatically active compound from the marine sponge Dysidea avara. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1985, 80, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Ferrándiz, M.L.; Sanz, M.J.; Bustos, G.; Payá, M.; Alcaraz, M.J.; De Rosa, S. Avarol and avarone, two new anti-inflammatory agents of marine origin. Eur. J. Pharmacol. 1994, 253, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Amigó, M.; Payá, M.; Braza-Boïls, A.; De Rosa, S.; Terencio, M.C. Avarol inhibits TNF-α generation and NF-κB activation in human cells and in animal models. Life Sci. 2008, 82, 256–264. [Google Scholar] [CrossRef]
- United Nations Department of Economic and Social Affairs, Population Division. World Population Prospects 2022: Summary of Results; UN DESA/POP/2022/TR/NO. 3; United Nations Publications: New York, NY, USA, 2022. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2022. In Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Gowen, R.J.; Bradbury, N.B. The ecological impact of salmonid farming in coastal waters: A review. Oceanogr. Mar. Biol. 1987, 25, 563–575. [Google Scholar] [CrossRef]
- Holdt, S.L.; Edwards, M.D. Cost-effective IMTA: A comparison of the production efficiencies of mussels and seaweed. J. App. Phycol. 2014, 26, 933–945. [Google Scholar] [CrossRef] [Green Version]
- Irisarri, J.; Fernández-Reiriz, M.J.; Labarta, U.; Cranford, P.J.; Robinson, S.M. Availability and utilization of waste fish feed by mussels Mytilus edulis in a commercial integrated multi-trophic aquaculture (IMTA) system: A multi-indicator assessment approach. Ecol. Indic. 2015, 48, 673–686. [Google Scholar] [CrossRef]
- Müller, W.E.; Wimmer, W.; Schatton, W.; Böhm, M.; Batel, R.; Filic, Z. Initiation of an aquaculture of sponges for the sustainable production of bioactive metabolites in open systems: Example, Geodia cydonium. Mar. Biotechnol. 1999, 1, 569–579. [Google Scholar] [CrossRef]
- Pronzato, R.; Cerrano, C.; Cubeddu, T.; Lanza, S.; Magnino, G.; Manconi, R.; Pantelis, J.; Sarà, A.; Sidri, M. Sustainable development in coastal areas: Role of sponge farming in integrated aquaculture. In Proceedings of the Aquaculture Europe, Bordeaux, France, 7–10 October 1998. [Google Scholar] [CrossRef]
- Pronzato, R. Sponge Farming in the Mediterranean Sea: New Perspectives. Mem. Qld. Mus. 1999, 44, 485–491. [Google Scholar]
- Osinga, R.; Sidri, M.; Cerig, E.; Gokalp, S.Z.; Gokalp, M. Sponge Aquaculture Trials in the East-Mediterranean Sea: New Approaches to Earlier Ideas. Open. Mar. Biol. J. 2010, 4, 74–81. [Google Scholar] [CrossRef]
- Longo, C.; Cardone, F.; Corriero, G.; Licciano, M.; Pierri, C.; Stabili, L. The co-occurrence of the demosponge Hymeniacidon perlevis and the edible mussel Mytilus galloprovincialis as a new tool for bacterial load mitigation in aquaculture. Environ. Sci. Pollut. Res. 2016, 23, 3736–3746. [Google Scholar] [CrossRef] [PubMed]
- Gökalp, M.; Wijgerde, T.; Sarà, A.; De Goeij, J.M.; Osinga, R. Development of an integrated mariculture for the collagen-rich sponge Chondrosia reniformis. Mar. Drugs 2019, 17, 29. [Google Scholar] [CrossRef] [Green Version]
- Longo, C.; Scrascia, M.; Trani, R.; Pierri, C.; Cariglia, A.; Cariglia, F.; Cariglia, M. Assesment of sponge mariculture potential in polycolture system in Manfredonia Gulf toward the IMTA implementation. In Proceedings of the Aquafarm Novelfarm, Pordenone, Italy, 19–20 February 2020. [Google Scholar]
- Giangrande, A.; Pierri, C.; Arduini, D.; Borghese, J.; Licciano, M.; Trani, R.; Corriero, G.; Basile, G.; Cecere, E.; Petrocelli, A. An Innovative IMTA System: Polychaetes, Sponges and Macroalgae Co-Cultured in a Southern Italian in-Shore Mariculture Plant (Ionian Sea). J. Mar. Sci. Eng. 2020, 8, 733. [Google Scholar] [CrossRef]
- Page, M.J.; Northcote, P.T.; Webb, V.L.; Mackey, S.; Handley, S.J. Aquaculture trials for the production of biologically active metabolites in the New Zealand sponge Mycale hentscheli (Demospongiae: Poecilosclerida). Aquaculture 2005, 250, 256–269. [Google Scholar] [CrossRef]
- FAO. Available online: https://www.fao.org/3/ac286e/AC286E00.htm (accessed on 26 November 2022).
- Cavolini, F. Memorie Per Servire Alla Storia de’ Polipi Marini; Wentworth Press: Napoli, Italy, 1785; pp. 262–265. [Google Scholar]
- Crawshay, L.R. Studies in the market sponges I. Growth from the planted cutting. J. Mar. Biol. Assoc. UK 1939, 23, 553–574. [Google Scholar] [CrossRef] [Green Version]
- Gökalp, N. Türkiye’de Ilk Sünger Yetistirme Tecrübeleri. Balık ve Balıkçılık 1974, 12, 1–10. [Google Scholar]
- Bierwirth, J.; Mantas, T.P.; Villechanoux, J.; Cerrano, C. Restoration of marine sponges—What can we learn from over a century of experimental cultivation? Water 2022, 14, 1055. [Google Scholar] [CrossRef]
- Duckworth, A. Farming Sponges to Supply Bioactive Metabolites and Bath Sponges: A Review. Mar. Biotechnol. 2009, 11, 669–679. [Google Scholar] [CrossRef]
- Verdenal, B.; Vacelet, J. Sponge culture on vertical ropes in the Northwestern Mediterranean Sea. In New Perspectives in Sponge Biology; Rützler, K., Ed.; Smithsonian Institution Press: Washington, DC, USA, 1990; pp. 416–424. [Google Scholar]
- Corriero, G.; Longo, C.; Mercurio, M.; Marzano, C.N.; Lembo, G.; Spedicato, M.T. Rearing performance of Spongia officinalis on suspended ropes off the Southern Italian coast (Central Mediterranean Sea). Aquaculture 2004, 238, 195–205. [Google Scholar] [CrossRef]
- Duckworth, A.R.; Wolff, C. Sath sponge aquaculture in Torres Strait, Australia: Effect of explant size, farming method and the environment on culture success. Aquaculture 2007, 271, 188–195. [Google Scholar] [CrossRef]
- Duckworth, A.R.; Wolff, C.; Evans-Illidge, E. Developing methods for commercially farming bath sponges in tropical Australia. Porifera Res.-Biodivers. Innov. Sustain. Rio de Jan. Mus. Nac. Ser. Livros 2007, 28, 297–302. [Google Scholar]
- Oronti, A.; Danylchuk, A.J.; Elmore, C.E.; Auriemma, R.; Pesle, G. Assessing the Feasibility of Sponge Aquaculture as a Sustainable Industry in The Bahamas. Aquac. Int. 2012, 20, 295–303. [Google Scholar] [CrossRef]
- Tobey, J.A.; Haws, M.C.; Ellis, S.S. Aquaculture Profile for Pohnpei Federated States of Micronesia; Technical Report; Pohnpei State Division of Marine Development, Office of Economic Affairs and the Conservation Society of Pohnpei. 2004. Available online: https://repository.library.noaa.gov/view/noaa/39936 (accessed on 26 November 2022).
- Marinecultures. Available online: https://www.marinecultures.org/en/projects/spongefarming/spongefarming/ (accessed on 26 November 2022).
- Simpson, T.L. The Cell Biology of Sponges, 1st ed.; Springer: New York, NY, USA, 1984. [Google Scholar]
- Riisgård, H.U.; Larsen, P.S. Filter-feeding in marine macro-invertebrates: Pump characteristics, modelling and energy cost. Biol. Rev. 1995, 70, 67–106. [Google Scholar] [CrossRef]
- Hadas, E.; Marie, D.; Shpigel, M.; Ilan, M. Virus predation by sponges is a new nutrient-flow pathway in coral reef food webs. Limnol. Oceanogr. 2006, 51, 1548–1550. [Google Scholar] [CrossRef] [Green Version]
- Ledda, F.D.; Pronzato, R.; Manconi, R. Mariculture for bacterial and organic waste removal: A field study of sponge filtering activity in experimental farming. Aquac. Res. 2014, 45, 1389–1401. [Google Scholar] [CrossRef]
- Gili, J.M.; Bibiloni, M.A.; Montserrat, A. Tasas de filtración y retención de bacterias “in situ” de tres especies de esponjas litorales. Estudio preliminar. Misc. Zool. 1984, 8, 13–21. [Google Scholar]
- Varamogianni-Mamatsi, D.; Anastasiou, T.I.; Vernadou, E.; Papandroulakis, N.; Kalogerakis, N.; Dailianis, T.; Mandalakis, M.A. Multi-species investigation of sponges’ filtering activity towards marine microalgae. Mar. Drugs 2021, 20, 24. [Google Scholar] [CrossRef]
- Milanese, M.; Chelossi, E.; Manconi, R.; Sarà, A.; Sidri, M.; Pronzato, R. The marine sponge Chondrilla nucula Schmidt, 1862 as an elective candidate for bioremediation in integrated aquaculture. Biomol. Eng. 2003, 20, 363–368. [Google Scholar] [CrossRef] [Green Version]
- Gökalp, M.; Kooistra, T.; Rocha, M.S.; Silva, T.H.; Osinga, R.; Murk, A.J.; Wijgerde, T. The effect of depth on the morphology, bacterial clearance, and respiration of the mediterranean sponge Chondrosia reniformis (Nardo, 1847). Mar. Drugs 2020, 18, 358. [Google Scholar] [CrossRef]
- Turon, X.; Galera, J.; Uriz, M.J. Clearance rates and aquiferous systems in two sponges with contrasting life-history strategies. J. Exp. Zool. 1997, 278, 22–36. [Google Scholar] [CrossRef]
- Ribes, M.; Coma, R.; Gili, J.-M. Natural diet and grazing rate of the temperate sponge Dysidea avara (Demospongiae, Dendroceratida) throughout an annual cycle. Mar. Ecol. Prog. Ser. 1999, 176, 179–190. [Google Scholar] [CrossRef]
- Kumala, L.; Riisgård, H.; Canfield, D. Osculum dynamics and filtration activity in small single-osculum explants of the Demosponge Halichondria panicea. Mar. Ecol. Prog. Ser. 2017, 572, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Stuart, V.; Klumpp, D. Evidence for food-resource partitioning by kelp-bed filter feeders. Mar. Ecol. Prog. Ser. 1984, 16, 27–37. [Google Scholar] [CrossRef]
- Fu, W.; Sun, L.; Zhang, X.; Zhang, W. Potential of the marine sponge Hymeniacidon perleve as a bioremediator of pathogenic bacteria in integrated aquaculture ecosystems. Biotechnol. Bioeng. 2006, 93, 1112–1122. [Google Scholar] [CrossRef]
- Hadas, E.; Shpigel, M.; Ilan, M. Particulate organic matter as a food source for a coral reef sponge. J. Exp. Biol. 2009, 212, 3643–3650. [Google Scholar] [CrossRef] [Green Version]
- Osinga, R.; Kleijn, R.; Groenendijk, E.; Niesink, P.; Tramper, J.; Wijffels, R.H. Development of in vivo sponge cultures: Particle feeding by the tropical sponge Pseudosuberites aff. andrewsi. Mar. Biotechnol. 2001, 3, 544–554. [Google Scholar] [CrossRef]
- Trani, R.; Corriero, G.; de Pinto, M.C.; Mercurio, M.; Pazzani, C.; Pierri, C.; Scrascia, M.; Longo, C. Filtering activity and nutrient release by the keratose sponge Sarcotragus spinosulus Schmidt, 1862 (Porifera, Demospongiae) at the laboratory scale. J. Mar. Sci. Eng. 2021, 9, 178. [Google Scholar] [CrossRef]
- Stabili, L.; Licciano, M.; Giangrande, A.; Longo, C.; Mercurio, M.; Marzano, C.N.; Corriero, G. Filtering activity of Spongia officinalis var. adriatica (Schmidt) (Porifera, Demospongiae) on bacterioplankton: Implications for bioremediation of polluted seawater. Water Res. 2006, 40, 3083–3090. [Google Scholar] [CrossRef]
- Riisgård, H.; Thomassen, S.; Jakobsen, H.; Weeks, J.; Larsen, P. Suspension feeding in marine sponges Halichondria panicea and Haliclona urceolus: Effects of temperature on filtration rate and energy cost of pumping. Mar. Ecol. Prog. Ser. 1993, 96, 177–188. [Google Scholar] [CrossRef]
- Ricciardi, A.; Bourget, E. Weight-to-Weight conversion factors for marine benthic macroinvertebrates. Mar. Ecol. Prog. Ser. 1998, 163, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Moitinho-Silva, L.; Steinert, G.; Nielsen, S.; Hardoim, C.C.; Wu, Y.-C.; McCormack, G.P.; López-Legentil, S.; Marchant, R.; Webster, N.; Thomas, T. Predicting the HMA-LMA status in marine sponges by machine learning. Front. Microbiol. 2017, 8, 752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, C.R.; Vacelet, J. Transplantation of marine sponges to different conditions of light and current. J. Exp. Mar. Biol. Ecol. 1979, 37, 91–104. [Google Scholar] [CrossRef]
- Duckworth, A.R.; Battershill, C.N.; Bergquist, P.R. Influence of explant procedures and environmental factors on culture success of three sponges. Aquaculture 1997, 156, 251–267. [Google Scholar] [CrossRef]
- Duckworth, A.R.; Battershill, C.N.; Schiel, D.R. Effects of depth and water flow on growth, survival and bioactivity of two temperate sponges cultured in different seasons. Aquaculture 2004, 242, 237–250. [Google Scholar] [CrossRef]
- Mendola, D.; De Caralt, S.; Uriz, M.; Van den End, F.; Van Leeuwen, J.L.; Wijffels, R.H. Environmental Flow Regimes for Dysidea avara Sponges. Mar. Biotechnol. 2008, 10, 622–630. [Google Scholar] [CrossRef]
- Gökalp, M.; Mes, D.; Nederlof, M.; Zhao, H.; Merijn de Goeij, J.; Osinga, R. The potential roles of sponges in integrated mariculture. Rev. Aquac. 2021, 13, 1159–1171. [Google Scholar] [CrossRef]
- Fu, W.; Wu, Y.; Sun, L.; Zhang, W. Efficient bioremediation of total organic carbon (TOC) in integrated aquaculture system by marine sponge Hymeniacidon perleve. Biotechnol. Bioeng. 2007, 97, 1387–1397. [Google Scholar] [CrossRef]
- Longo, C.; Corriero, G.; Licciano, M.; Stabili, L. Bacterial accumulation by the Demospongiae Hymeniacidon perlevis: A tool for the bioremediation of polluted seawater. Mar. Pollut. Bull. 2010, 60, 1182–1187. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Larsen, P.S. Comparative ecophysiology of active zoobenthic filter feeding, essence of current knowledge. J. Sea Res. 2000, 44, 169–193. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leatherland, T.M.; Burton, J.D. The occurrence of some trace metals in coastal organisms with particular reference to the Solent Region. J. Mar. Biol. Assoc. UK 1974, 54, 457–468. [Google Scholar] [CrossRef]
- Patel, B.; Balani, M.C.; Patel, S. Sponge ‘sentinel’ of heavy metals. Sci. Total Environ. 1985, 41, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Olesen, T.M.E.; Weeks, J.M. Accumulation of Cd by the marine sponge Halichondria panicea Pallas: Effects upon filtration rate and its relevance for biomonitoring. Bull. Environ. Contam. Toxicol. 1994, 52, 722–728. [Google Scholar] [CrossRef]
- Hansen, I.V.; Weeks, J.M.; Depledge, M.H. Accumulation of Copper, Zinc, Cadmium and Chromium by the marine sponge Halichondria panicea Pallas and the implications for biomonitoring. Mar. Pollut. Bull. 1995, 31, 133–138. [Google Scholar] [CrossRef]
- Philp, R.B. Cadmium content of the marine sponge Microciona prolifera, other sponges, water and sediment from the eastern Florida panhandle: Possible effects on Microciona cell aggregation and potential roles of low pH and low salinity. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1999, 124, 41–49. [Google Scholar] [CrossRef]
- Cebrian, E.; Martı, R.; Uriz, J.M.; Turon, X. Sublethal effects of contamination on the mediterranean sponge Crambe crambe: Metal accumulation and biological responses. Mar. Pollut. Bull. 2003, 46, 1273–1284. [Google Scholar] [CrossRef]
- Rao, J.V.; Kavitha, P.; Reddy, N.C.; Rao, T.G. Petrosia testudinaria as a biomarker for metal contamination at Gulf of Mannar, Southeast Coast of India. Chemosphere 2006, 65, 634–638. [Google Scholar] [CrossRef]
- Rao, J.V.; Kavitha, P.; Srikanth, K.; Usman, P.K.; Rao, T.G. Environmental contamination using accumulation of metals in marine sponge, Sigmadocia fibulata inhabiting the coastal waters of Gulf of Mannar, India. Toxicol. Environ. Chem. 2007, 89, 487–498. [Google Scholar] [CrossRef]
- Venkateswara Rao, J.; Srikanth, K.; Pallela, R.; Gnaneshwar Rao, T. The use of marine sponge, Haliclona tenuiramosa as bioindicator to monitor heavy metal pollution in the coasts of Gulf of Mannar, India. Environ. Monit. Assess. 2009, 156, 451. [Google Scholar] [CrossRef]
- Annibaldi, A.; Truzzi, C.; Illuminati, S.; Bassotti, E.; Finale, C.; Scarponi, G. First systematic voltammetric measurements of Cd, Pb, and Cu in hydrofluoric acid-dissolved siliceous spicules of marine sponges: Application to Antarctic specimens. Anal. Lett. 2011, 44, 2792–2807. [Google Scholar] [CrossRef]
- Mahaut, M.-L.; Basuyaux, O.; Baudinière, E.; Chataignier, C.; Pain, J.; Caplat, C. The Porifera Hymeniacidon perlevis (Montagu, 1818) as a bioindicator for water quality monitoring. Environ. Sci. Pollut. Res. 2013, 20, 2984–2992. [Google Scholar] [CrossRef] [PubMed]
- Batista, D.; Muricy, G.; Rocha, R.C.; Miekeley, N.F. Marine sponges with contrasting life histories can be complementary biomonitors of heavy metal pollution in coastal ecosystems. Environ. Sci. Pollut. Res. 2014, 21, 5785–5794. [Google Scholar] [CrossRef] [PubMed]
- Maloubier, M.; Michel, H.; Solari, P.L.; Moisy, P.; Tribalat, M.-A.; Oberhaensli, F.R.; Bottein, M.Y.D.; Thomas, O.P.; Monfort, M.; Moulin, C. Speciation of Americium in seawater and accumulation in the marine sponge Aplysina cavernicola. Dalton Trans. 2015, 44, 20584–20596. [Google Scholar] [CrossRef] [PubMed]
- Celik, F.; Camas, M.; Camas, A.S.; Ozalp, H.B. Uranium (VI) biosorption on marine sponge, Sarcotragus foetidus (Schmidt, 1862) and its statistical investigation using central composite design. Turkish J. Fish. Aquat. Sci. 2016, 16, 899–911. [Google Scholar] [CrossRef]
- Gentric, C.; Rehel, K.; Dufour, A.; Sauleau, P. Bioaccumulation of metallic trace elements and organic pollutants in marine sponges from the South Brittany Coast, France. J. Environ. Sci. Health A 2016, 51, 213–219. [Google Scholar] [CrossRef]
- Ferrante, M.; Vassallo, M.; Mazzola, A.; Brundo, M.V.; Pecoraro, R.; Grasso, A.; Copat, C. In Vivo Exposure of the marine sponge Chondrilla nucula Schmidt, 1862 to Cadmium (Cd), Copper (Cu) and Lead (Pb) and its potential use for bioremediation purposes. Chemosphere 2018, 193, 1049–1057. [Google Scholar] [CrossRef]
- Orani, A.M.; Barats, A.; Vassileva, E.; Thomas, O.P. Marine sponges as a powerful tool for trace elements biomonitoring studies in coastal environment. Mar. Pollut. Bull. 2018, 131, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, G.R.; Morales, E.O. Assessment of heavy metal contamination at Tallaboa Bay (Puerto Rico) by marine sponges’ bioaccumulation and fungal community composition. Mar. Pollut. Bull. 2020, 161, 111803. [Google Scholar] [CrossRef]
- Srikanth, K.; Rao, J.V.; Rao, A.R. Trace elements in Endectyon fruticosa collected from a sewage outfall site, Therespuram, Tuticorin coast, India. Int. J. Environ. Sci. Technol. 2020, 17, 267–272. [Google Scholar] [CrossRef]
- Gravina, M.F.; Longo, C.; Puthod, P.; Rosati, M.; Colozza, N.; Scarselli, M. Heavy metal accumulation capacity of Axinella damicornis (Esper, 1794) (Porifera, Demospongiae): A tool for bioremediation of polluted seawaters. Mediterr. Mar. Sci. 2022, 23, 125–133. [Google Scholar] [CrossRef]
- Krikech, I.; Jafarabadi, A.R.; Leermakers, M.; Le Pennec, G.; Cappello, T.; Ezziyyani, M. Insights into bioaccumulation and bioconcentration of potentially toxic elements in marine sponges from the Northwestern Mediterranean coast of Morocco. Mar. Pollut. Bull. 2022, 180, 113770. [Google Scholar] [CrossRef] [PubMed]
- Orani, A.M.; Vassileva, E.; Thomas, O.P. Marine sponges as coastal bioindicators of rare earth elements bioaccumulation in the French Mediterranean Sea. Environ. Pollut. 2022, 304, 119172. [Google Scholar] [CrossRef] [PubMed]
- Genta-Jouve, G.; Cachet, N.; Oberhänsli, F.; Noyer, C.; Teyssié, J.-L.; Thomas, O.P.; Lacoue-Labarthe, T. Comparative bioaccumulation kinetics of trace elements in Mediterranean marine sponges. Chemosphere 2012, 89, 340–349. [Google Scholar] [CrossRef]
- Illuminati, S.; Annibaldi, A.; Truzzi, C.; Scarponi, G. Heavy metal distribution in organic and siliceous marine sponge tissues measured by square wave anodic stripping voltammetry. Mar. Pollut. Bull. 2016, 111, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Lacoue-Labarthe, T.; Warnau, M.; Beaugeard, L.; Pascal, P.-Y. Trophic transfer of radioisotopes in Mediterranean sponges through bacteria consumption. Chemosphere 2016, 144, 1885–1892. [Google Scholar] [CrossRef]
- Agusa, T.; Takagi, K.; Kubota, R.; Anan, Y.; Iwata, H.; Tanabe, S. Specific accumulation of arsenic compounds in green turtles (Chelonia mydas) and hawksbill turtles (Eretmochelys imbricata) from Ishigaki Island, Japan. Environ. Pollut. 2008, 153, 127–136. [Google Scholar] [CrossRef]
- Batel, R.; Bihari, N.; Rinkevich, B.; Dapper, J.; Schäcke, H.; Schröder, H.C.; Müller, W.E. Modulation of organotin-induced apoptosis by the water pollutant methyl mercury in a human lymphoblastoid tumour cell line and a marine sponge. Mar. Ecol. Prog. Ser. 1993, 93, 245–251. [Google Scholar] [CrossRef]
- Wagner, C.; Steffen, R.; Koziol, C.; Batel, R.; Lacorn, M.; Steinhart, H.; Simat, T.; Müller, W.E.G. Apoptosis in marine sponges: A biomarker for environmental stress (cadmium and bacteria). Mar. Biol. 1998, 131, 411–421. [Google Scholar] [CrossRef]
- Schröder, H.C.; Hassanein, H.M.A.; Lauenroth, S.; Koziol, C.; Mohamed, T.-A.; Lacorn, M.; Steinhart, H.; Batel, R.; Müller, W.E.G. Induction of DNA strand breaks and expression of HSP70 and GRP78 homolog by cadmium in the marine sponge Suberites domuncula. Arch. Environ. Contam. Toxicol. 1999, 36, 47–55. [Google Scholar] [CrossRef]
- Wanick, R.C.; de Sousa Barbosa, H.; Frazão, L.R.; Santelli, R.E.; Arruda, M.A.Z.; Coutinho, C.C. Evaluation of differential protein expression in Haliclona aquarius and sponge-associated microorganisms under cadmium stress. Anal. Bioanal. Chem. 2013, 405, 7661–7670. [Google Scholar] [CrossRef]
- Akpiri, R.U.; Konya, R.S.; Hodges, N.J. Development of cultures of the marine sponge Hymeniacidon perleve for genotoxicity assessment using the alkaline comet assay. Environ. Toxicol. Chem. 2017, 36, 3314–3323. [Google Scholar] [CrossRef] [PubMed]
- Zahn, R.K.; Kurelec, B.; Zahn-Daimler, G.; Müller, W.E.G.; Rijavec, M.; Batel, R.; Given, R.; Pondeljak, V.; Beyer, Ŕ. The effect of benzo[a]pyrene on sponges as model organisms in marine pollution. Chem. Biol. Interact. 1982, 39, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Perez, T.; Wafo, E.; Fourt, M.; Vacelet, J. Marine sponges as biomonitor of polychlorobiphenyl contamination: Concentration and fate of 24 congeners. Environ. Sci. Technol. 2003, 37, 2152–2158. [Google Scholar] [CrossRef] [PubMed]
- Haraguchi, K.; Kato, Y.; Ohta, C.; Koga, N.; Endo, T. Marine sponge: A potential source for methoxylated polybrominated diphenyl ethers in the Asia-Pacific food web. J. Agric. Food Chem. 2011, 59, 13102–13109. [Google Scholar] [CrossRef]
- Aresta, A.; Marzano, C.N.; Lopane, C.; Corriero, G.; Longo, C.; Zambonin, C.; Stabili, L. Analytical investigations on the lindane bioremediation capability of the Demosponge Hymeniacidon perlevis. Mar. Pollut. Bull. 2015, 90, 143–149. [Google Scholar] [CrossRef]
- Webster, L.; Russell, M.; Shepherd, N.; Packer, G.; Dalgarno, E.J.; Neat, F. Monitoring of Polycyclic Aromatic Hydrocarbons (PAHs) in Scottish Deepwater environments. Mar. Pollut. Bull. 2018, 128, 456–459. [Google Scholar] [CrossRef]
- Wu, Q.; Eisenhardt, N.; Holbert, S.S.; Pawlik, J.R.; Kucklick, J.R.; Vetter, W. Naturally occurring organobromine compounds (OBCs) including polybrominated dibenzo-p-dioxins in the marine sponge Hyrtios proteus from The Bahamas. Mar. Pollut. Bull. 2021, 172, 112872. [Google Scholar] [CrossRef]
- Batista, D.; Tellini, K.; Nudi, A.H.; Massone, T.P.; Scofield, A.D.L.; de LR Wagener, A. Marine sponges as bioindicators of oil and combustion derived PAH in coastal waters. Mar. Environ. Res. 2013, 92, 234–243. [Google Scholar] [CrossRef]
- Schröder, H.C.; Badria, F.A.; Ayyad, S.N.; Batel, R.; Wiens, M.; Hassanein, H.M.; Kurelec, B.; Müller, W.E. Inhibitory effects of extracts from the marine alga Caulerpa taxifolia and of toxin from Caulerpa racemosa on multixenobiotic resistance in the marine sponge Geodia cydonium. Environ. Toxicol. Pharmacol. 1998, 5, 119–126. [Google Scholar] [CrossRef]
- Châtel, A.; Talarmin, H.; Hamer, B.; Schröder, H.C.; Müller, W.E.G.; Dorange, G. MAP Kinase cell signaling pathway as biomarker of environmental pollution in the sponge Suberites domuncula. Ecotoxicology 2011, 20, 1727–1740. [Google Scholar] [CrossRef] [PubMed]
- Modica, L.; Lanuza, P.; García-Castrillo, G. Surrounded by microplastic, since when? Testing the feasibility of exploring past levels of plastic microfibre pollution using natural history museum collections. Mar. Pollut. Bull. 2020, 151, 110846. [Google Scholar] [CrossRef] [PubMed]
- Soares, G.M.; Barros, F.; Lanna, E.; da Silva, M.V.S.; Cavalcanti, F.F. Sponges as libraries: Increase in microplastics in Cinachyrella alloclada after 36 years. Mar. Pollut. Bull. 2022, 185, 114339. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, T.M.; Vethaak, A.D.; Almroth, B.C.; Ariese, F.; van Velzen, M.; Hassellöv, M.; Leslie, H.A. Screening for microplastics in sediment, water, marine invertebrates and fish: Method development and microplastic accumulation. Mar. Pollut. Bull. 2017, 122, 403–408. [Google Scholar] [CrossRef]
- Celis-Hernández, O.; Ávila, E.; Ward, R.D.; Rodríguez-Santiago, M.A.; Aguirre-Téllez, J.A. Microplastic distribution in urban vs pristine mangroves: Using marine sponges as bioindicators of environmental pollution. Environ. Pollut. 2021, 284, 117391. [Google Scholar] [CrossRef] [PubMed]
- Fallon, B.R.; Freeman, C.J. Plastics in Porifera: The occurrence of potential microplastics in marine sponges and seawater from Bocas Del Toro, Panamá. PeerJ 2021, 9, e11638. [Google Scholar] [CrossRef] [PubMed]
- Giametti, S.D.; Finelli, C.M. Detection of plastic-associated compounds in marine sponges. Mar. Pollut. Bull. 2022, 175, 113141. [Google Scholar] [CrossRef]
- Saliu, F.; Biale, G.; Raguso, C.; La Nasa, J.; Degano, I.; Seveso, D.; Galli, P.; Lasagni, M.; Modugno, F. Detection of plastic particles in marine sponges by a combined infrared micro-spectroscopy and pyrolysis-gas chromatography-mass spectrometry approach. Sci. Total Environ. 2022, 819, 152965. [Google Scholar] [CrossRef]
- Baird, C.A. Measuring the Effects of Microplastics on Sponges. Ph.D. Thesis, Te Herenga Waka-Victoria University of Wellington, Wellington, New Zealand, 2016. [Google Scholar]
- De Marchi, L.; Renzi, M.; Anselmi, S.; Pretti, C.; Guazzelli, E.; Martinelli, E.; Cuccaro, A.; Oliva, M.; Magri, M.; Bulleri, F. Polyethylene microplastics reduce filtration and respiration rates in the Mediterranean sponge Petrosia ficiformis. Environ. Res. 2022, 211, 113094. [Google Scholar] [CrossRef]
- Selvin, J.; Priya, S.S.; Kiran, G.S.; Thangavelu, T.; Bai, N.S. Sponge-associated marine bacteria as indicators of heavy metal pollution. Microbiol. Res. 2009, 164, 352–363. [Google Scholar] [CrossRef]
- Mori, T.; Iwamoto, K.; Wakaoji, S.; Araie, H.; Kohara, Y.; Okamura, Y.; Shiraiwa, Y.; Takeyama, H. Characterization of a novel gene involved in cadmium accumulation screened from sponge-associated bacterial metagenome. Gene 2016, 576, 618–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, E.; Gonçalves, J.M.; Reis, M.; Costa, R. Draft genome Sequence of Microbacterium Sp. strain Alg239_V18, an Actinobacterium retrieved from the marine sponge Spongia Sp. Genome Announc. 2017, 5, e01457-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Liu, C.; Zhang, F.; Zhao, M.; Li, Z. Comparative genomics provides insights into the marine adaptation in sponge-derived Kocuria flava S43. Front. Microbiol. 2018, 9, 1257. [Google Scholar] [CrossRef] [Green Version]
- Freitas-Silva, J.; de Oliveira, B.F.R.; Vigoder, F.D.M.; Muricy, G.; Dobson, A.D.; Laport, M.S. Peeling the layers away: The genomic characterization of Bacillus pumilus 64-1, an isolate with antimicrobial activity from the marine sponge Plakina cyanorosea (Porifera, Homoscleromorpha). Front. Microbiol. 2021, 11, 592735. [Google Scholar] [CrossRef]
- Marzuki, I.; Kamaruddin, M.; Ahmad, R.; Asaf, R.; Armus, R.; Siswanty, I. Performance of cultured marine sponges-symbiotic bacteria as a heavy metal bio-adsorption. Biodiversitas 2021, 22, 5536–5543. [Google Scholar] [CrossRef]
- Chen, Y.; Pan, T.; Chai, G.; Li, Z. Complete Genome of Mycetocola spongiae MSC19T isolated from deep-sea sponge Cacospongia mycofijiensis indicates the adaptation to deep-sea environment and sponge-microbe symbioses. Mar. Genom. 2022, 63, 100955. [Google Scholar] [CrossRef] [PubMed]
- Bauvais, C.; Zirah, S.; Piette, L.; Chaspoul, F.; Domart-Coulon, I.; Chapon, V.; Gallice, P.; Rebuffat, S.; Pérez, T.; Bourguet-Kondracki, M.-L. Sponging up metals: Bacteria associated with the marine sponge Spongia officinalis. Mar. Environ. Res. 2015, 104, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Gantt, S.E.; Lopez-Legentil, S.; Erwin, P.M. Stable microbial communities in the sponge Crambe crambe from inside and outside a polluted Mediterranean harbor. FEMS Microbiol. Lett. 2017, 364, 11. [Google Scholar] [CrossRef] [PubMed]
- Webster, N.S.; Webb, R.I.; Ridd, M.J.; Hill, R.T.; Negri, A.P. The effects of copper on the microbial community of a coral reef sponge. Environ. Microbiol. 2001, 3, 19–31. [Google Scholar] [CrossRef]
- Santos-Gandelman, J.F.; Cruz, K.; Crane, S.; Muricy, G.; Giambiagi-deMarval, M.; Barkay, T.; Laport, M.S. Potential application in mercury bioremediation of a marine sponge-isolated Bacillus cereus strain Pj1. Curr. Microbiol. 2014, 69, 374–380. [Google Scholar] [CrossRef]
- Sajayan, A.; Kiran, G.S.; Priyadharshini, S.; Poulose, N.; Selvin, J. Revealing the ability of a novel polysaccharide bioflocculant in bioremediation of heavy metals sensed in a Vibrio bioluminescence reporter assay. Environ. Pollut. 2017, 228, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Orani, A.M.; Barats, A.; Zitte, W.; Morrow, C.; Thomas, O.P. Comparative study on the bioaccumulation and biotransformation of arsenic by some northeastern Atlantic and northwestern Mediterranean sponges. Chemosphere 2018, 201, 826–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orani, A.M.; Vassileva, E.; Azemard, S.; Thomas, O.P. Comparative study on Hg bioaccumulation and biotransformation in Mediterranean and Atlantic sponge species. Chemosphere 2020, 260, 127515. [Google Scholar] [CrossRef]
- Ravindran, A.; Sajayan, A.; Priyadharshini, G.B.; Selvin, J.; Kiran, G.S. Revealing the efficacy of thermostable biosurfactant in heavy metal bioremediation and surface treatment in vegetables. Front. Microbiol. 2020, 11, 222. [Google Scholar] [CrossRef]
- Freitas-Silva, J.; de Oliveira, B.F.R.; Dias, G.R.; de Carvalho, M.M.; Laport, M.S. Unravelling the sponge microbiome as a promising source of biosurfactants. Crit. Rev. Microbiol. 2022, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Almeida, E.L.; Rincón, A.F.C.; Jackson, S.A.; Dobson, A.D. In silico screening and heterologous expression of a polyethylene terephthalate hydrolase (PETase)-like enzyme (SM14est) with polycaprolactone (PCL)-degrading activity, from the marine sponge-derived strain Streptomyces sp. SM14. Front. Microbiol. 2019, 10, 2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, T.; Sarrazin, L.; Rebouillon, P.; Vacelet, J. First evidences of surfactant biodegradation by marine sponges (porifera): An experimental study with a linear alkylbenzenesulfonate. Hydrobiologia 2002, 489, 225–233. [Google Scholar] [CrossRef]
- Bonugli-Santos, R.C.; Durrant, L.R.; Sette, L.D. The production of ligninolytic enzymes by marine-derived Basidiomycetes and their biotechnological potential in the biodegradation of recalcitrant pollutants and the treatment of textile effluents. Water Air Soil Pollut. 2012, 223, 2333–2345. [Google Scholar] [CrossRef]
- Dhasayan, A.; Kiran, G.S.; Selvin, J. Production and characterisation of glycolipid biosurfactant by Halomonas Sp. MB-30 for potential application in enhanced oil recovery. Appl. Biochem. Biotechnol. 2014, 174, 2571–2584. [Google Scholar] [CrossRef]
- Loredana, S.; Graziano, P.; Antonio, M.; Carlotta, N.M.; Caterina, L.; Maria, A.A.; Carlo, Z.; Giuseppe, C.; Pietro, A. Lindane bioremediation capability of bacteria associated with the Demosponge Hymeniacidon perlevis. Mar. Drugs 2017, 15, 108. [Google Scholar] [CrossRef]
- Holert, J.; Cardenas, E.; Bergstrand, L.H.; Zaikova, E.; Hahn, A.S.; Hallam, S.J.; Mohn, W.W. Metagenomes reveal global distribution of bacterial steroid catabolism in natural, engineered, and host environments. mBio 2018, 9, e02345-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bojko, B.; Onat, B.; Boyaci, E.; Psillakis, E.; Dailianis, T.; Pawliszyn, J. Application of in situ solid-phase microextraction on Mediterranean sponges for untargeted exometabolome screening and environmental monitoring. Front. Mar. Sci. 2019, 6, 632. [Google Scholar] [CrossRef]
- Vasconcelos, M.R.; Vieira, G.A.; Otero, I.V.; Bonugli-Santos, R.C.; Rodrigues, M.V.; Rehder, V.L.; Ferro, M.; Boaventura, S.; Bacci, M.; Sette, L.D. Pyrene degradation by marine-derived ascomycete: Process optimization, toxicity, and metabolic analyses. Environ. Sci. Pollut. Res. 2019, 26, 12412–12424. [Google Scholar] [CrossRef] [PubMed]
- Horna-Gray, I.; Lopez, N.A.; Nijenhuis, I.; Ahn, Y.; Richnow, H.H.; Häggblom, M.M. Reductive debromination by sponge-associated anaerobic bacteria coupled to carbon isotope fractionation. Int. Biodeterior. Biodegrad. 2020, 155, 105093. [Google Scholar] [CrossRef]
- Marzuki, I.; Nisaa, K.; Asaf, R.; Athirah, A.; Paena, M.; Susianingsih, E.; Nurhidayah, N.; Kadriah, I.A.K.; Kamaruddin, K.; Sahabuddin, S. Comparison of pyrene biodegradation using two types of marine bacterial isolates. Sustainability 2022, 14, 9890. [Google Scholar] [CrossRef]
- Schleuter, D.; Günther, A.; Paasch, S.; Ehrlich, H.; Kljajić, Z.; Hanke, T.; Bernhard, G.; Brunner, E. Chitin-based renewable materials from marine sponges for uranium adsorption. Carbohydr. Polym. 2013, 92, 712–718. [Google Scholar] [CrossRef]
- Wang, D.; Song, J.; Lin, S.; Wen, J.; Ma, C.; Yuan, Y.; Lei, M.; Wang, X.; Wang, N.; Wu, H. A marine-inspired hybrid sponge for highly efficient uranium extraction from seawater. Adv. Funct. Mater. 2019, 29, 1901009. [Google Scholar] [CrossRef]
- Machałowski, T.; Jankowska, K.; Bachosz, K.; Smulek, W.; Ehrlich, H.; Kaczorek, E.; Zdarta, J.; Jesionowski, T. Biocatalytic system made of 3D chitin, silica nanopowder and horseradish peroxidase for the removal of 17α-ethinylestradiol: Determination of process efficiency and degradation mechanism. Molecules 2022, 27, 1354. [Google Scholar] [CrossRef]
- Bergmann, W.; Feeney, R.J. Contributions to the Study of Marine Products. XXXII. The Nucleosides of Sponges. I. J. Org. Chem. 1951, 16, 981–987. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [Green Version]
- Proksch, P.; Edrada, R.; Ebel, R. Drugs from the seas–current status and microbiological implications. Appl. Microbiol. Biotechnol. 2002, 59, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Maslin, M.; Gaertner-Mazouni, N.; Debitus, C.; Joy, N.; Ho, R. Marine sponge aquaculture towards drug development: An ongoing history of technical, ecological, chemical considerations and challenges. Aquac. Rep. 2021, 21, 100813. [Google Scholar] [CrossRef]
- Sipkema, D.; Franssen, M.C.; Osinga, R.; Tramper, J.; Wijffels, R.H. Marine sponges as pharmacy. Mar. Biotechnol. 2005, 7, 142–162. [Google Scholar] [CrossRef] [PubMed]
- Rotter, A.; Barbier, M.; Bertoni, F.; Bones, A.M.; Cancela, M.L.; Carlsson, J.; Carvalho, M.F.; Cegłowska, M.; Chirivella-Martorell, J.; Conk Dalay, M.; et al. The essentials of marine biotechnology. Front. Mar. Sci. 2021, 8, 629629. [Google Scholar] [CrossRef]
- Varijakzhan, D.; Loh, J.-Y.; Yap, W.-S.; Yusoff, K.; Seboussi, R.; Lim, S.-H.E.; Lai, K.-S.; Chong, C.M. Bioactive compounds from marine sponges: Fundamentals and applications. Mar. Drugs 2021, 19, 246. [Google Scholar] [CrossRef]
- Lipton, A.P.; Sunith, S. Mariculture of marine sponges for drug development: Bioactivity potentials of cultured sponges, Callyspongia subarmigera (Ridley) and Echinodictyum gorgonoides (Dendy). Mar. Fish. Infor. Serv. T&E Ser. 2009, 202, 7–10. Available online: http://eprints.cmfri.org.in/id/eprint/6469 (accessed on 26 November 2022).
- Page, M.J.; Handley, S.J.; Northcote, P.T.; Cairney, D.; Willan, R.C. Successes and pitfalls of the aquaculture of the sponge Mycale hentscheli. Aquaculture 2011, 312, 52–61. [Google Scholar] [CrossRef]
- Santiago, V.S.; Manzano, G.G.; Clairecynth, C.Y.; Aliño, P.M.; Salvador-Reyes, L.A. Mariculture potential of renieramycin-producing philippine blue sponge Xestospongia sp. (Porifera: Haplosclerida). Aquaculture 2019, 502, 356–364. [Google Scholar] [CrossRef]
- Mishra, S.; Das, R.; Swain, P. Status of fish diseases in aquaculture and assessment of economic loss due to disease. In Contemporary Trends in Fisheries and Aquaculture; Rao, P., Pandey, B., Pandey, P., Joshi, B.D., Eds.; Today & Tomorrow’s Printers and Publishers: New Delhi, India, 2018. [Google Scholar]
- Randall, J.E.; Hartman, W.D. Sponge-feeding fishes of the West Indies. Mar. Biol. 1968, 1, 216–225. [Google Scholar] [CrossRef]
- Wulff, J.L. Sponge Feeding by Caribbean Angelfishes, Trunkfishes, and Filefishes. In Sponges in Time and Space; van Soests, R.W.M., van Kempen, T.M.G., Braekman, J.C., Eds.; Balkema: Rotterdam, The Netherlands, 1994; pp. 265–271. [Google Scholar]
- Ruzicka, R.; Gleason, D.F. Latitudinal variation in spongivorous fishes and the effectiveness of sponge chemical defenses. Oecologia 2008, 154, 785–794. [Google Scholar] [CrossRef]
- Kaandorp, J.A.; De Kluijver, M.J. Verification of fractal growth models of the sponge Haliclona oculata (Porifera) with transplantation experiments. Mar. Biol. 1992, 113, 133–143. [Google Scholar] [CrossRef]
- van Treeck, P.; Eisinger, M.; Müller, J.; Paster, M.; Schuhmacher, H. Mariculture trials with Mediterranean sponge species: The exploitation of an old natural resource with sustainable and novel methods. Aquaculture 2003, 218, 439–455. [Google Scholar] [CrossRef]
- Kelly, M.; Handley, S.; Page, M.; Butterfield, P.; Hartill, B.; Kelly, S. Aquaculture trials of the New Zealand bath-sponge Spongia (Heterofibria) manipulatus using lanterns. N. Z. J. Mar. Freshw. Res. 2004, 38, 231–241. [Google Scholar] [CrossRef] [Green Version]
- De Voogd, N.J. An assessment of sponge mariculture potential in the Spermonde Archipelago, Indonesia. J. Mar. Biolog. Assoc. UK 2007, 87, 1777–1784. [Google Scholar] [CrossRef]
- de Voogd, N.J. The mariculture potential of the Indonesian reef-dwelling sponge Callyspongia (Euplacella) biru: Growth, survival and bioactive compounds. Aquaculture 2007, 262, 54–64. [Google Scholar] [CrossRef]
- Carballo, J.L.; Yañez, B.; Zubía, E.; Ortega, M.J.; Vega, C. Culture of explants from the sponge Mycale cecilia to obtain bioactive mycalazal-type metabolites. Mar. Biotechnol. 2010, 12, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Gökalp, M.; Wijgerde, T.; Murk, A.; Osinga, R. Design for large-scale maricultures of the mediterranean demosponge Chondrosia reniformis Nardo, 1847 for collagen production. Aquaculture 2022, 548, 737702. [Google Scholar] [CrossRef]
- Giangrande, A.; Gravina, M.F.; Rossi, S.; Longo, C.; Pierri, C. Aquaculture and restoration: Perspectives from mediterranean sea experiences. Water 2021, 13, 991. [Google Scholar] [CrossRef]
- Baldacconi, R.; Cardone, F.; Longo, C.; Mercurio, M.; Marzano, C.N.; Gaino, E.; Corriero, G. Transplantation of Spongia officinalis L. (Porifera, Demospongiae): A technical approach for restocking this endangered species. Mar. Ecol. 2010, 31, 309–317. [Google Scholar] [CrossRef]
- Biggs, B.C. Harnessing natural recovery processes to improve restoration outcomes: An experimental assessment of sponge-mediated coral reef restoration. PLoS ONE 2013, 8, e64945. [Google Scholar] [CrossRef] [Green Version]
- De Caralt, S.; Sánchez-Fontenla, J.; Uriz, M.J.; Wijffels, R.H. In situ aquaculture methods for Dysidea avara (Demospongiae, Porifera) in the Northwestern Mediterranean. Mar. Drugs 2010, 8, 1731–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, N.S.; Cobb, R.E.; Negri, A.P. Temperature thresholds for bacterial symbiosis with a sponge. ISME J. 2008, 2, 830–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United Nations Department of Economic and Social Affairs. The Sustainable Development Goals Report; United Nations: New York, NY, USA, 2022; p. 68. [Google Scholar] [CrossRef]
Figure 1.
Comparison of sponge filtration rates (bars) according to species, explant biomass (circles), and microorganism characteristics. The cited bibliographic references are shown in the ref column [36,37,38,39,40,41,42,43,44,45,46,47,48,49]. Rayed bars represent in situ studies where no specific microorganism was used. When standardizing explant size and filtration rates, volume-to-weight and weight-to-weight conversions were made based on [50,51], respectively. Microbiological abundance classification was that described in [52]. M (Mediterranean), NEA (North-eastern Atlantic), SEA (South-eastern Atlantic), NWP (North-western Pacific), and RS (Red Sea) for the geographic area; HMA (High Microbiological Abundance) and LMA (Low Microbiological Abundance) for microbiological abundance and S (Small, <15 µm), B (Big, >15 µm), Y (motile), and N (non-motile) for microorganism description are the abbreviations used.
Figure 1.
Comparison of sponge filtration rates (bars) according to species, explant biomass (circles), and microorganism characteristics. The cited bibliographic references are shown in the ref column [36,37,38,39,40,41,42,43,44,45,46,47,48,49]. Rayed bars represent in situ studies where no specific microorganism was used. When standardizing explant size and filtration rates, volume-to-weight and weight-to-weight conversions were made based on [50,51], respectively. Microbiological abundance classification was that described in [52]. M (Mediterranean), NEA (North-eastern Atlantic), SEA (South-eastern Atlantic), NWP (North-western Pacific), and RS (Red Sea) for the geographic area; HMA (High Microbiological Abundance) and LMA (Low Microbiological Abundance) for microbiological abundance and S (Small, <15 µm), B (Big, >15 µm), Y (motile), and N (non-motile) for microorganism description are the abbreviations used.

Figure 2.
Cumulative number of compound discoveries (light) and publications (dark) on marine sponges over the last seven decades. The proportion of marine natural products of poriferan origin regarding the total number of described molecules is shown in the pie chart. Row data from https://marinlit.rsc.org (accessed on 14 November 2022).
Figure 2.
Cumulative number of compound discoveries (light) and publications (dark) on marine sponges over the last seven decades. The proportion of marine natural products of poriferan origin regarding the total number of described molecules is shown in the pie chart. Row data from https://marinlit.rsc.org (accessed on 14 November 2022).

Figure 3.
The bright orange and stylized sponge Axinella cannabina as an ornamental aquarium species.
Figure 3.
The bright orange and stylized sponge Axinella cannabina as an ornamental aquarium species.

Figure 4.
Algae, ascidians, polychaetes, and sea anemones growing on Sarcotragus spinosulus (a) and Aplysina aerophoba (b) reared in the Remedia Life IMTA system.
Figure 4.
Algae, ascidians, polychaetes, and sea anemones growing on Sarcotragus spinosulus (a) and Aplysina aerophoba (b) reared in the Remedia Life IMTA system.

Figure 5.
Main rearing methods in Remedia Life (upper) and ASMAR (lower) IMTA systems. Sarcotragus spinosulus reared in tubular nets (a), Aplysina aerophoba (b), and Hymeniacydon perlevis (c) reared in rope systems with net bags in the Maricoltura del Mar Grande fish farm (Mar Grande of Taranto, northern Ionian Sea); S. spinosulus in SEAPA nets (d), mesh panels (e) and lantern nets (f) close to Gargano fish farm (Gulf of Manfredonia, south-eastern Adriatic Sea). ASMAR project: Assessment of Sponge Mariculture Potential in Polyculture System in the Manfredonia Gulf–Interreg Adrion Blue Boost 2019–2020.
Figure 5.
Main rearing methods in Remedia Life (upper) and ASMAR (lower) IMTA systems. Sarcotragus spinosulus reared in tubular nets (a), Aplysina aerophoba (b), and Hymeniacydon perlevis (c) reared in rope systems with net bags in the Maricoltura del Mar Grande fish farm (Mar Grande of Taranto, northern Ionian Sea); S. spinosulus in SEAPA nets (d), mesh panels (e) and lantern nets (f) close to Gargano fish farm (Gulf of Manfredonia, south-eastern Adriatic Sea). ASMAR project: Assessment of Sponge Mariculture Potential in Polyculture System in the Manfredonia Gulf–Interreg Adrion Blue Boost 2019–2020.

Figure 6.
Sponges as a by-product of an IMTA system. Environmental and economic benefits are represented in blue and green, respectively.
Figure 6.
Sponges as a by-product of an IMTA system. Environmental and economic benefits are represented in blue and green, respectively.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Aguilo-Arce, J.; Ferriol, P.; Trani, R.; Puthod, P.; Pierri, C.; Longo, C. Sponges as Emerging By-Product of Integrated Multitrophic Aquaculture (IMTA). J. Mar. Sci. Eng. 2023, 11, 80. https://doi.org/10.3390/jmse11010080
AMA Style
Aguilo-Arce J, Ferriol P, Trani R, Puthod P, Pierri C, Longo C. Sponges as Emerging By-Product of Integrated Multitrophic Aquaculture (IMTA). Journal of Marine Science and Engineering. 2023; 11(1):80. https://doi.org/10.3390/jmse11010080
Chicago/Turabian StyleAguilo-Arce, Joseba, Pere Ferriol, Roberta Trani, Patrizia Puthod, Cataldo Pierri, and Caterina Longo. 2023. "Sponges as Emerging By-Product of Integrated Multitrophic Aquaculture (IMTA)" Journal of Marine Science and Engineering 11, no. 1: 80. https://doi.org/10.3390/jmse11010080
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.