
Ecomorphological Differentiation of Feeding Structures within the Antarctic Fish Species Flock Trematominae (Notothenioidei) from Terra Nova Bay (Ross Sea)
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
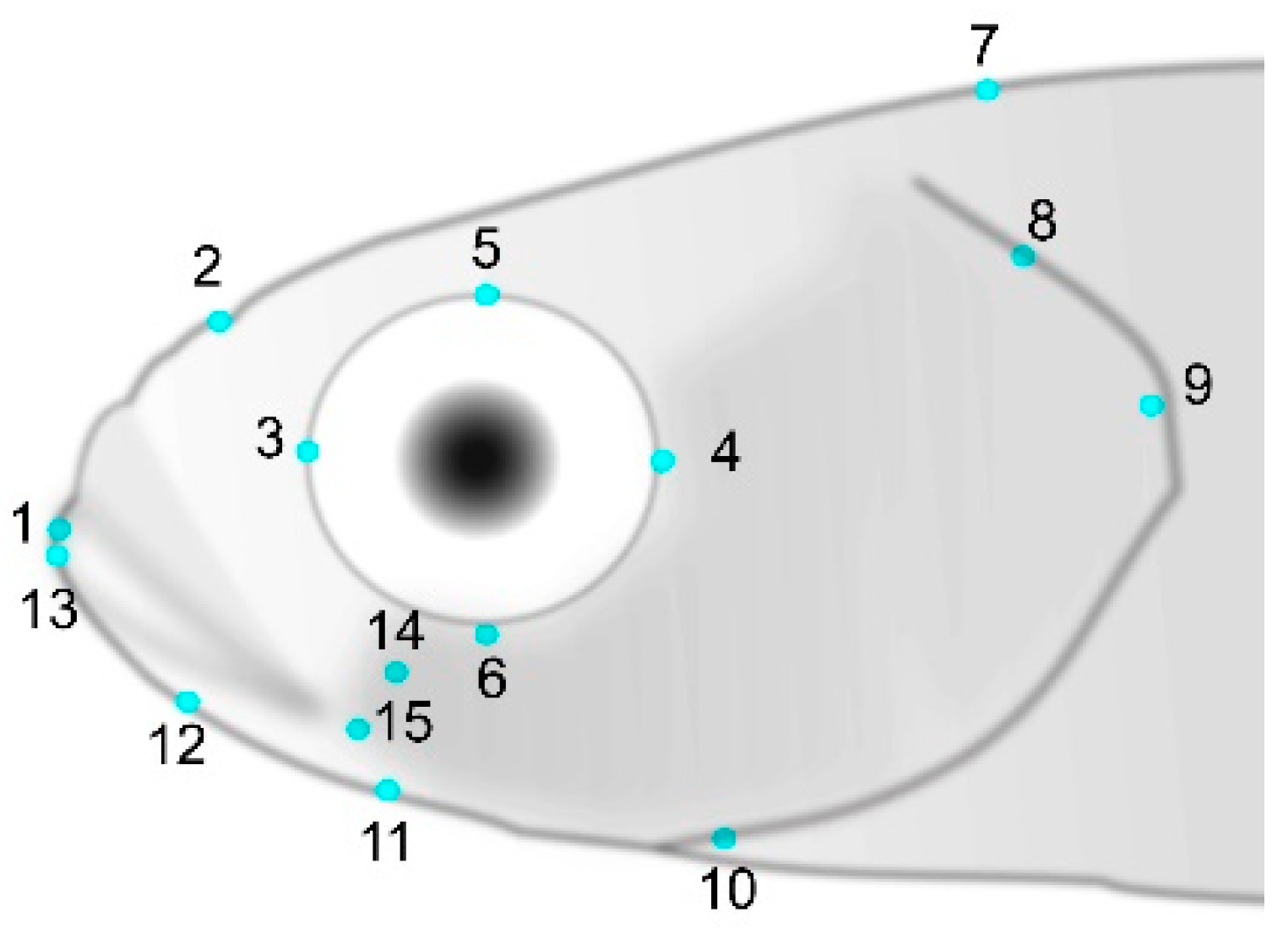
2.2. Morphological Traits
2.3. Geometric Morphometrics
2.4. Statistical Analyses
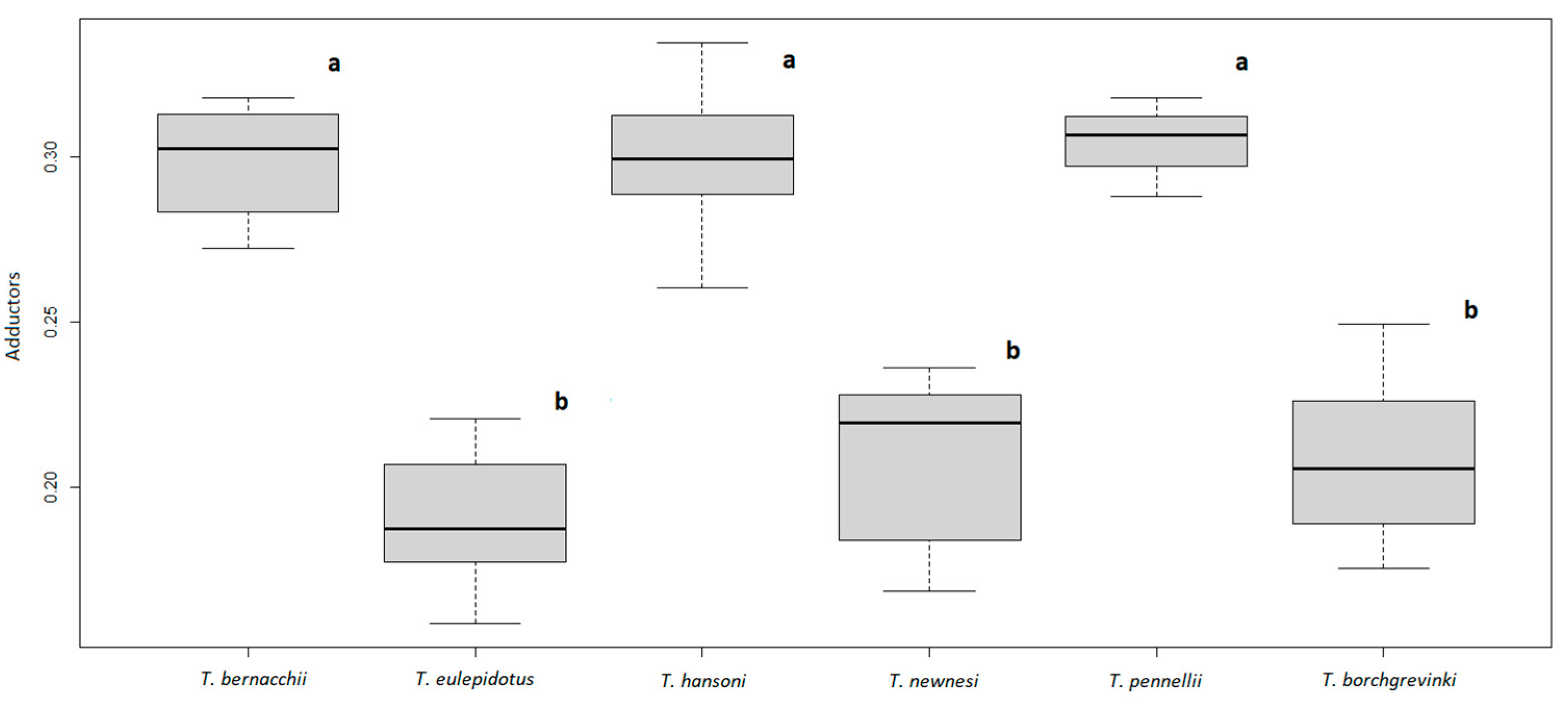
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schluter, D. The Ecology of Adaptive Radiation; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Colombo, M.; Damerau, M.; Hanel, R.; Salzburger, W.; Matschiner, M. Diversity and disparity through time in the adaptive radiation of Antarctic notothenioid fishes. J. Evol. Biol. 2015, 28, 376–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastman, J.T.; McCune, A.R. Fishes on the Antarctic continental shelf: Evolution of a marine species flock? J. Fish Biol. 2000, 57, 84–102. [Google Scholar] [CrossRef]
- Eastman, J.T. The nature of the diversity of Antarctic fishes. Polar Biol. 2005, 28, 93–107. [Google Scholar] [CrossRef]
- La Mesa, M.; Eastman, J.T.; Vacchi, M. The role of notothenioid fish in the food web of the Ross Sea shelf waters: A review. Polar Biol. 2004, 27, 321–338. [Google Scholar] [CrossRef]
- Janko, K.; Marshall, C.; Musilová, Z.; Van Houdt, J.; Couloux, A.; Cruaud, C.; Lecointre, G. Multilocus analyses of an Antarctic fish species flock (Teleostei, Notothenioidei, Trematominae): Phylogenetic approach and test of the early-radiation event. Mol. Phylogenetics Evol. 2011, 60, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T.; La Mesa, M. Neuromorphological disparity in deep-living sister species of the Antarctic fish genus Trematomus. Polar Biol. 2021, 44, 315–334. [Google Scholar] [CrossRef]
- Lannoo, M.J.; Eastman, J.T. Nervous and sensory system correlates of an epibenthic evolutionary radiation in Antarctic no-tothenioid fishes, genus Trematomus (Perciformes; Nototheniidae). J. Morphol. 2000, 245, 67–79. [Google Scholar] [CrossRef]
- Duhamel, G.; Hulley, P.A.; Causse, R.; Koubbi, P.; Vacchi, M.; Pruvost, P.; Vigetta, S.; Irisson, J.O.; Mormède, S.; Belchier, M.; et al. Biogeographic patterns of fish. In Biogeographic Atlas of the Southern Ocean; De Broyer, C., Koubbi, P., Griffiths, H.J., Raymond, B., d’Udekem d’Acoz, C., Van de Putte, A.P., Danis, B., David, B., Grant, S., Gutt, J., et al., Eds.; Scientific Committee on Antarctic Research: Cambridge, UK, 2014; pp. 328–362. [Google Scholar]
- Eastman, J.T.; Devries, A.L. Biology and phenotypic plasticity of the Antarctic nototheniid fish Trematomus newnesi in McMurdo Sound. Antarct. Sci. 1997, 9, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Vacchi, M.; La Mesa, M.; Greco, S. The Coastal Fish Fauna of Terra Nova Bay, Ross Sea, Antarctica. In Ross Sea Ecology; Faranda, F.M., Guglielmo, L., Ianora, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 457–468. [Google Scholar] [CrossRef]
- Parker, E.; Zapfe, K.L.; Yadav, J.; Frédérich, B.; Jones, C.D.; Economo, E.P.; Federman, S.; Near, T.J.; Dornburg, A. Periodic Environmental Disturbance Drives Repeated Ecomorphological Diversification in an Adaptive Radiation of Antarctic Fishes. bioRxiv 2000. [Google Scholar] [CrossRef]
- Eastman, J.T.; Eakin, R.R. Checklist of the species of notothenioid fishes. Antarct. Sci. 2021, 33, 273–280. [Google Scholar] [CrossRef]
- Gambi, M.C.; Lorenti, M.; Russo, G.F.; Scipione, M.B. Benthic associations of the shallow hard bottoms off Terra Nova Bay, Ross Sea: Zonation, biomass and population structure. Antarct. Sci. 1994, 6, 449–462. [Google Scholar] [CrossRef]
- La Mesa, M.; Vacchi, M.; Castelli, A.; Diviacco, G. Feeding ecology of two nototheniid fishes, Trematomus hansoni and Trematomus loennbergii, from Terra Nova Bay, Ross Sea. Polar Biol. 1997, 17, 62–68. [Google Scholar] [CrossRef]
- Chiantore, M.; Cattaneo-Vietti, R.; Berkman, P.A.; Nigro, M.; Vacchi, M.; Schiaparelli, S.; Albertelli, G. Antarctic scallop (Adamussium colbecki) spatial population variability along the Victoria Land Coast, Antarctica. Polar Biol. 2001, 24, 139–143. [Google Scholar] [CrossRef]
- Dell’Acqua, O.; Brey, T.; Vacchi, M.; Chiantore, M. Predation impact of the notothenioid fish Trematomus bernacchii on the size structure of the scallop Adamussium colbecki in Terra Nova Bay (Ross Sea, Antarctica). Polar Biol. 2017, 40, 1557–1568. [Google Scholar] [CrossRef] [Green Version]
- Cattaneo-Vietti, R.; Chiantore, M.; Schiaparelli, S.; Albertelli, G. Shallow- and deep-water mollusc distribution at Terra Nova Bay (Ross Sea, Antarctica). Polar Biol. 2000, 23, 173–182. [Google Scholar] [CrossRef]
- Schiaparelli, S.; Cattaneo-Vietti, R.; Chiantore, M. Adaptive morphology of Capulus subcompressus Pelseneer, 1903 (Gastropoda: Capulidae) from Terra Nova Bay, Ross Sea (Antarctica). Polar Biol. 2000, 23, 11–16. [Google Scholar] [CrossRef]
- La Mesa, M.; Canese, S.; Montagna, P.; Schiaparelli, S. Underwater Photographic Survey of Coastal Fish Community of Terra Nova Bay, Ross Sea. Diversity 2022, 14, 315. [Google Scholar] [CrossRef]
- Wainwright, P.C. Morphology and Ecology: Functional Basis of Feeding Constraints in Caribbean Labrid Fishes. Ecology 1988, 69, 635–645. [Google Scholar] [CrossRef]
- Sonnefeld, M.J.; Turingan, R.G.; Sloan, T.J. Functional Morphological Drivers of Feeding Mode in Marine Teleost Fishes. Adv. Zool. Bot. 2014, 2, 6–14. [Google Scholar] [CrossRef]
- Gidmark, N.J.; Pos, K.; Matheson, B.; Ponce, E.; Westneat, M.W. Functional Morphology and Biomechanics of Feeding in Fishes. In Feeding in Vertebrates; Bels, V., Whishaw, I., Eds.; Springer: Cham, Switzerland, 2019; pp. 297–332. [Google Scholar] [CrossRef]
- Delariva, R.L.; Neves, M.P. Morphological traits correlated with resource partitioning among small characin fish species coexisting in a Neotropical river. Ecol. Freshw. Fish 2020, 29, 640–653. [Google Scholar] [CrossRef]
- Wainwright, P.C.; Bellwood, D.R. Ecomorphology of feeding in coral reef fishes. In Coral Reef Fishes: Dynamics and Diversity in a Complex Ecosystem; Sale, P.F., Ed.; Academic Press: San Diego, CA, USA, 2002; pp. 33–55. [Google Scholar]
- Liem, K.F. Acquisition of Energy by Teleosts: Adaptive Mechanisms and Evolutionary Patterns. In Environmental Physiology of Fishes, NATO Advanced Study Institute Series; Ali, M.A., Ed.; Springer: Boston, MA, USA, 1980; Volume 35, pp. 299–334. [Google Scholar] [CrossRef]
- Norton, S.F.; Brainerd, E.L. Convergence in the feeding mechanics of ecomorphologically similar species in the Centrarchidae and Cichlidae. J. Exp. Biol. 1993, 176, 11–29. [Google Scholar] [CrossRef]
- Motta, P.J.; Norton, S.F.; Luczkovich, J.J. Perspectives on the ecomorphology of bony fishes. Env. Biol. Fish. 1995, 44, 11–20. [Google Scholar] [CrossRef]
- Carroll, A.M.; Wainwright, P.C.; Huskey, S.H.; Collar, D.C.; Turingan, R.G. Morphology predicts suction feeding performance in centrarchid fishes. J. Exp. Biol. 2004, 207, 3873–3881. [Google Scholar] [CrossRef] [Green Version]
- Westneat, M.W. Skull Biomechanics and Suction Feeding in Fishes. In Fish Phisiology. Fish Biomechanics; Shadwick, R.E., Lauder, G.V., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2005; Volume 23, pp. 29–75. [Google Scholar] [CrossRef]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Rohlf, F.J.; Marcus, L.F. A revolution morphometrics. Trends Ecol. Evol. 1993, 8, 129–132. [Google Scholar] [CrossRef]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. Geometric morphometrics: Ten years of progress following the ‘revolution’. Ital. J. Zool. 2004, 71, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.C.; Otárola-Castillo, E. geomorph: An R package for the collection and analysis of geometric morphometric shape data. Methods Ecol. Evol. 2013, 4, 393–399. [Google Scholar] [CrossRef]
- Frédérich, B.; Heindler, F.M.; Christiansen, H.; Dettai, A.; Van de Putte, A.P.; Volckaert, F.A.; Lepoint, G. Repeated morphological diversification in endemic Antarctic fishes of the genus Trematomus. Belg. J. Zool. 2022, 152, 55–73. [Google Scholar] [CrossRef]
- Mittelheiser, L.; Lepoint, G.; Gillet, A.; Frédérich, B. Ecomorphology of six goatfish species (Mullidae) from Toliara Reef, Madagascar. Env. Biol. Fish. 2022, 105, 1015–1032. [Google Scholar] [CrossRef]
- Hu, Y.; Ghigliotti, L.; Vacchi, M.; Pisano, E.; Detrich, H.W.; Albertson, R.C. Evolution in an extreme environment: Developmental biases and phenotypic integration in the adaptive radiation of antarctic notothenioids. BMC Evol. Biol. 2016, 16, 142. [Google Scholar] [CrossRef] [Green Version]
- Carlig, E.; Di Blasi, D.; Ghigliotti, L.; Pisano, E.; Faimali, M.; O’Driscoll, R.; Parker, S.; Vacchi, M. Diversification of feeding structures in three adult Antarctic nototheniid fish. Polar Biol. 2018, 41, 1707–1715. [Google Scholar] [CrossRef]
- Barnett, A.; Bellwood, D.R.; Hoey, A.S. Trophic ecomorphology of cardinalfish. Mar. Ecol. Prog. Ser. 2006, 322, 249–257. [Google Scholar] [CrossRef]
- Collar, D.C.; Wainwright, P.C. Discordance between Morphological and Mechanical Diversity in the Feeding Mechanism of Centrarchid Fishes. Evolution 2006, 60, 2575–2584. [Google Scholar] [CrossRef]
- Bansode, M.A.; Eastman, J.T.; Aronson, R.B. Feeding biomechanics of five demersal Antarctic fishes. Polar Biol. 2014, 37, 1835–1848. [Google Scholar] [CrossRef]
- Westneat, M.W. Evolution of Levers and Linkages in the Feeding Mechanisms of Fishes. Integr. Comp. Biol. 2004, 44, 378–389. [Google Scholar] [CrossRef]
- Carlig, E.; Christiansen, J.S.; Di Blasi, D.; Ferrando, S.; Pisano, E.; Vacchi, M.; O’Driscoll, R.L.; Ghigliotti, L. Midtrophic fish feeding modes at the poles: An ecomorphological comparison of polar cod (Boreogadus saida) and Antarctic silverfish (Pleuragramma antarctica). Polar Biol. 2021, 44, 1629–1642. [Google Scholar] [CrossRef]
- Pombo-Ayora, L.; Tavera, J. Are Feeding Modes Concealing Morphofunctional Diversity? The Case of the New World Parrotfishes. Front. Mar. Sci. 2021, 8, 634046. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Olsen, A.M.; Westneat, M.W. StereoMorph: An R package for the collection of 3D landmarks and curves using a stereo camera set-up. Methods Ecol. Evol. 2015, 6, 351–356. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Slice, D. Extensions of the Procrustes Method for the Optimal Superimposition of Landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: www.R-project.org (accessed on 15 July 2022).
- Near, T.J.; Dornburg, A.; Kuhn, K.L.; Eastman, J.T.; Pennington, J.N.; Patarnello, T.; Zane, L.; Fernández, D.A.; Jones, C.D. Ancient climate change, antifreeze, and the evolutionary diversification of Antarctic fishes. Proc. Natl. Acad. Sci. USA 2012, 109, 3434–3439. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.M.; Wainwright, P.C. Muscle function and power output during suction feeding in largemouth bass, Micropterus salmoides. Comp. Biochem. Phys. A 2006, 143, 389–399. [Google Scholar] [CrossRef]
- Daniels, R.A. Feeding ecology of some fishes of the Antarctic Peninsula. Fish Bull 1982, 80, 575–588. [Google Scholar]
- Pakhomov, E.A. Feeding plasticity of the Antarctic fish Trematomus hansoni Boulenger, 1902 (Pisces: Nototheniidae): The influence of fishery waste on the diet. Polar Biol. 1998, 19, 289–292. [Google Scholar] [CrossRef]
- Di Blasi, D.; Canese, S.; Carlig, E.; Parker, S.J.; Pisano, E.; Vacchi, M.; Ghigliotti, L. The Challenge to Observe Antarctic Toothfish (Dissostichus mawsoni) under Fast Ice. J. Mar. Sci. Eng. 2021, 9, 255. [Google Scholar] [CrossRef]
- Janssen, J. Use of the lateral line and tactile senses in feeding in four antarctic nototheniid fishes. Env. Biol. Fish. 1996, 47, 51–64. [Google Scholar] [CrossRef]
- La Mesa, M.; Caputo, V.; Eastman, J.T. The reproductive biology of two epibenthic species of Antarctic nototheniid fish of the genus Trematomus. Antarct. Sci. 2008, 20, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, J.C.; MacDonald, J.A. Sensory Tuning of Lateral Line Receptors in Antarctic Fish to the Movements of Planktonic Prey. Science 1987, 235, 195–196. [Google Scholar] [CrossRef]
- Macdonald, J.A.; Montgomery, J.C.; Prisco, G. The Sensory Biology of Notothenioid Fish. In Biology of Antarctic Fish; di Prisco, G., Maresca, B., Tota, B., Eds.; Springer: Berlin/Heidelberg, Germany, 1991; pp. 145–162. [Google Scholar] [CrossRef]
- Foster, B.A.; Montgomery, J.C. Planktivory in benthic nototheniid fish in McMurdo Sound, Antarctica. Environ. Biol. Fishes 1993, 36, 313–318. [Google Scholar] [CrossRef]
- Barrera-Oro, E.; Eastman, J.T.; Moreira, E. Phenotypic plasticity in the Antarctic nototheniid fish Trematomus newnesi: A guide to the identification of typical, large mouth and intermediate morphs. Polar Biol. 2012, 35, 1047–1056. [Google Scholar] [CrossRef]
- La Mesa, M.; Vacchi, M.; Zunini Sertorio, T.Z. Feeding plasticity of Trematomus newnesi (Pisces, Nototheniidae) in Terra Nova Bay, Ross Sea, in relation to environmental conditions. Polar Biol. 2000, 23, 38–45. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
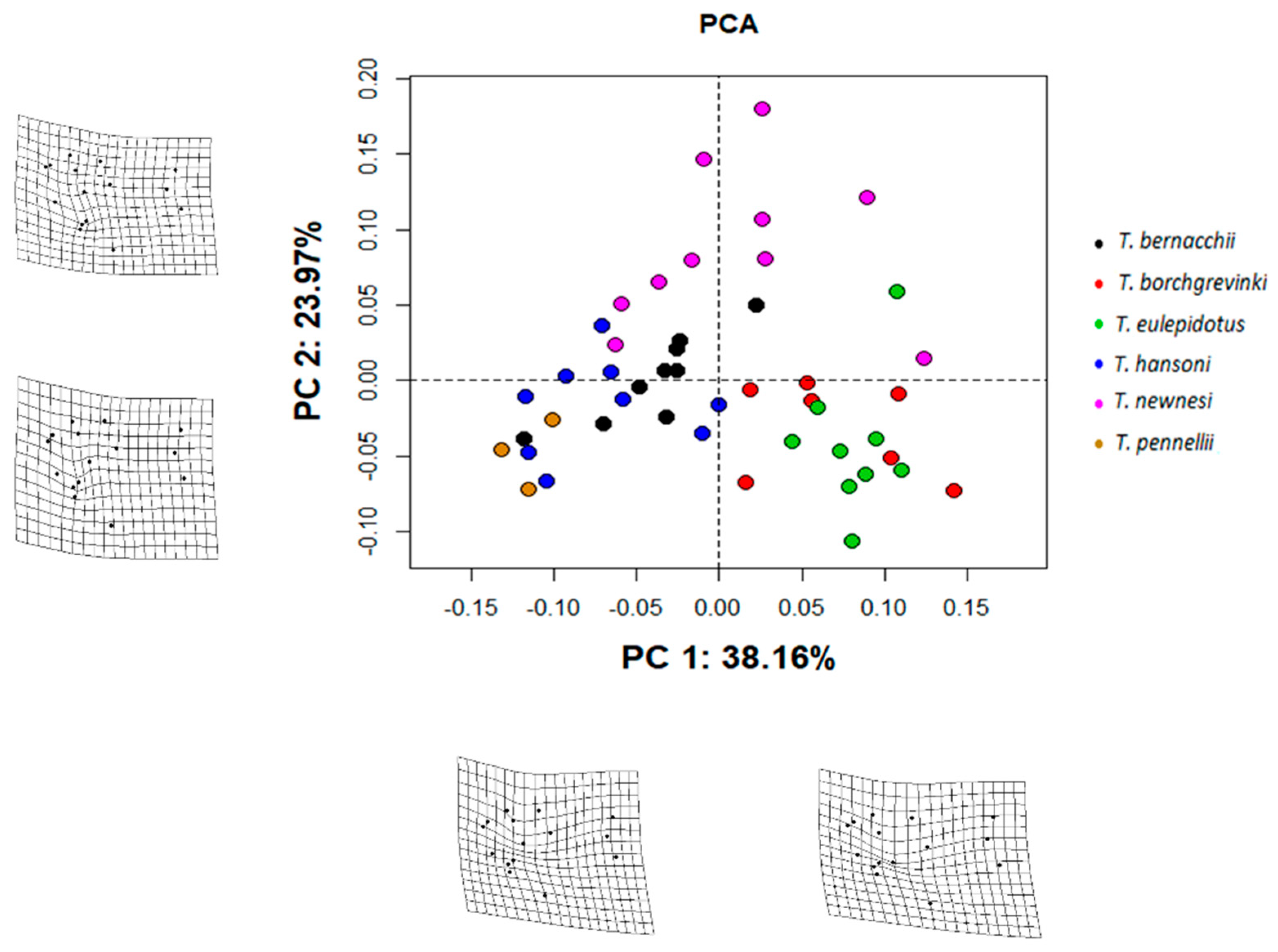{kind=link}
| Species | n | SL Min (mm) | SL Max (mm) | Habitat | Feeding Habits |
|---|---|---|---|---|---|
| Trematomus bernacchii | 9 | 173 | 203 | benthic | ominivorous |
| Trematomus borchgrevinki | 7 | 146 | 181 | (cryo)pelagic | zooplanktivorous |
| Trematomus eulepidotus | 9 | 135 | 192 | epibenthic | zooplanktivorous |
| Trematomus hansoni | 9 | 177 | 281 | benthic | ominivorous |
| Trematomus newnesi | 10 | 130 | 159 | pelagic | zooplanktivorous |
| Trematomus pennellii | 3 | 126 | 145 | benthic | ominivorous |
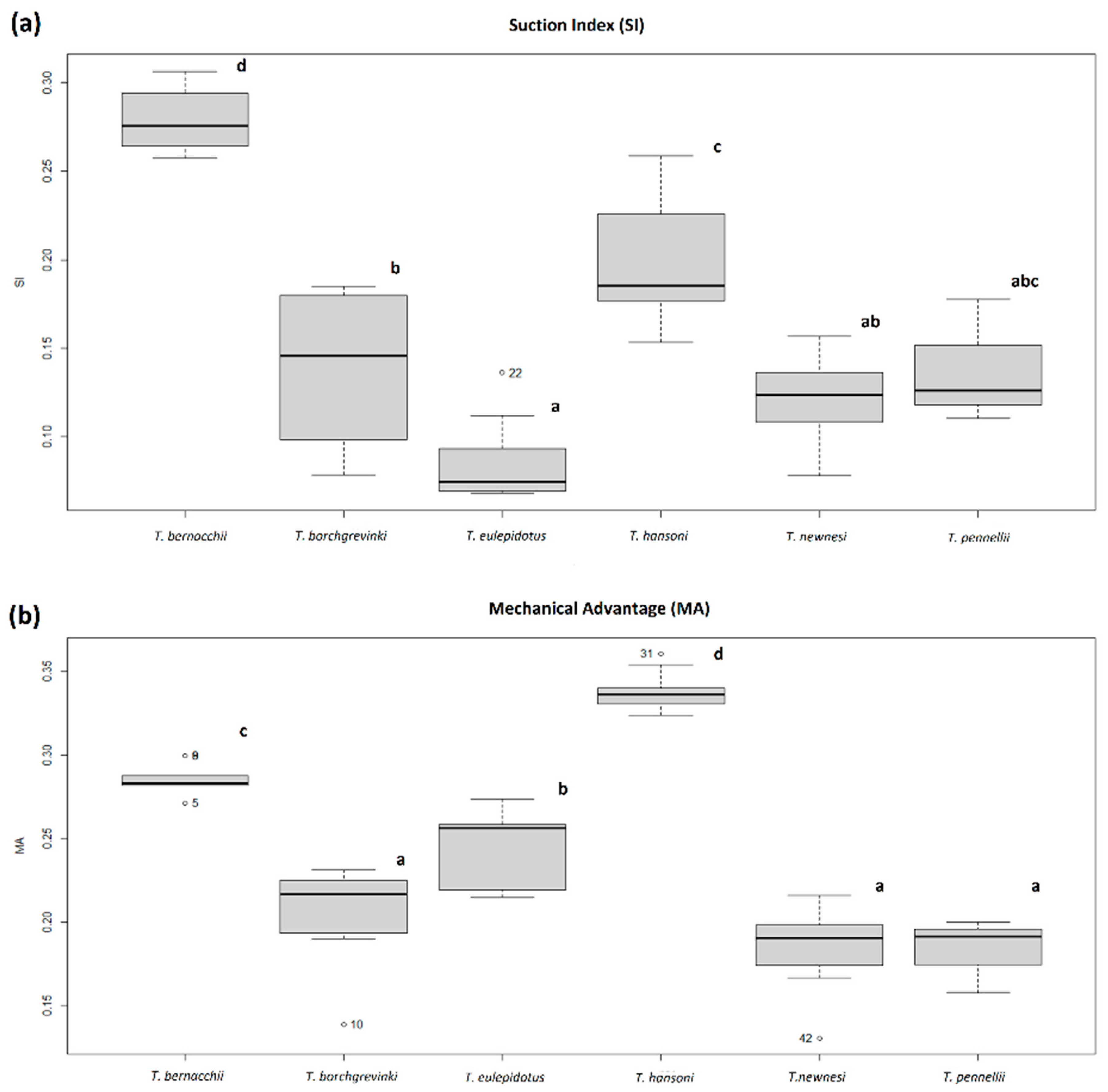
| Species | n | Suction Index (Mean ± SD) | Mechanical Advantage (Mean ± SD) |
|---|---|---|---|
| T. bernacchii | 9 | 0.279 ± 0.017 | 0.286 ± 0.009 |
| T. borchgrevinki | 7 | 0.138 ± 0.046 | 0.203 ± 0.032 |
| T. eulepidotus | 9 | 0.087 ± 0.024 | 0.243 ± 0.025 |
| T. hansoni | 9 | 0.199 ± 0.038 | 0.338 ± 0.012 |
| T. newnesi | 10 | 0.123 ± 0.024 | 0.183 ± 0.023 |
| T. pennellii | 3 | 0.138 ± 0.035 | 0.183 ± 0.022 |
| Traits | T. bernacchii Mean ± SD | T. borchgrevinki Mean ± SD | T. eulepidotus Mean ± SD | T. hansoni Mean ± SD | T. newnesi Mean ± SD | T. pennellii Mean ± SD |
|---|---|---|---|---|---|---|
| ED | 0.070 ± 0.002 | 0.081 ± 0.005 | 0.083 ± 0.013 | 0.067 ± 0.007 | 0.071 ± 0.006 | 0.085 ± 0.006 |
| HL | 0.304 ± 0.011 | 0.280 ± 0.010 | 0.288 ± 0.016 | 0.280 ± 0.012 | 0.228 ± 0.114 | 0.287 ± 0.004 |
| GW | 0.103 ± 0.010 | 0.114 ± 0.006 | 0.104 ± 0.007 | 0.114 ± 0.004 | 0.116 ± 0.013 | 0.100 ± 0.003 |
| BL | 0.090 ± 0.002 | 0.195 ± 0.008 | 0.195 ± 0.015 | 0.107 ± 0.007 | 0.204 ± 0.019 | 0.209 ± 0.008 |
| CSAepaxialis | 0.101 ± 0.002 | 0.092 ± 0.008 | 0.076 ± 0.010 | 0.096 ± 0.007 | 0.094 ± 0.003 | 0.096 ± 0.003 |
| LinSI | 0.039 ± 0.001 | 0.037 ± 0.004 | 0.031 ± 0.005 | 0.040 ± 0.002 | 0.038 ± 0.002 | 0.039 ± 0.003 |
| LoutSI | 0.155 ± 0.005 | 0.107 ± 0.005 | 0.105 ± 0.009 | 0.154 ± 0.008 | 0.119 ± 0.013 | 0.128 ± 0.008 |
| LinMA | 0.040 ± 0.001 | 0.023 ± 0.002 | 0.032 ± 0.003 | 0.041± 0.002 | 0.028 ± 0.004 | 0.021 ± 0.003 |
| LoutMA | 0.142 ± 0.004 | 0.118 ± 0.024 | 0.133 ± 0.007 | 0.120 ± 0.005 | 0.152 ± 0.008 | 0.116 ± 0.004 |
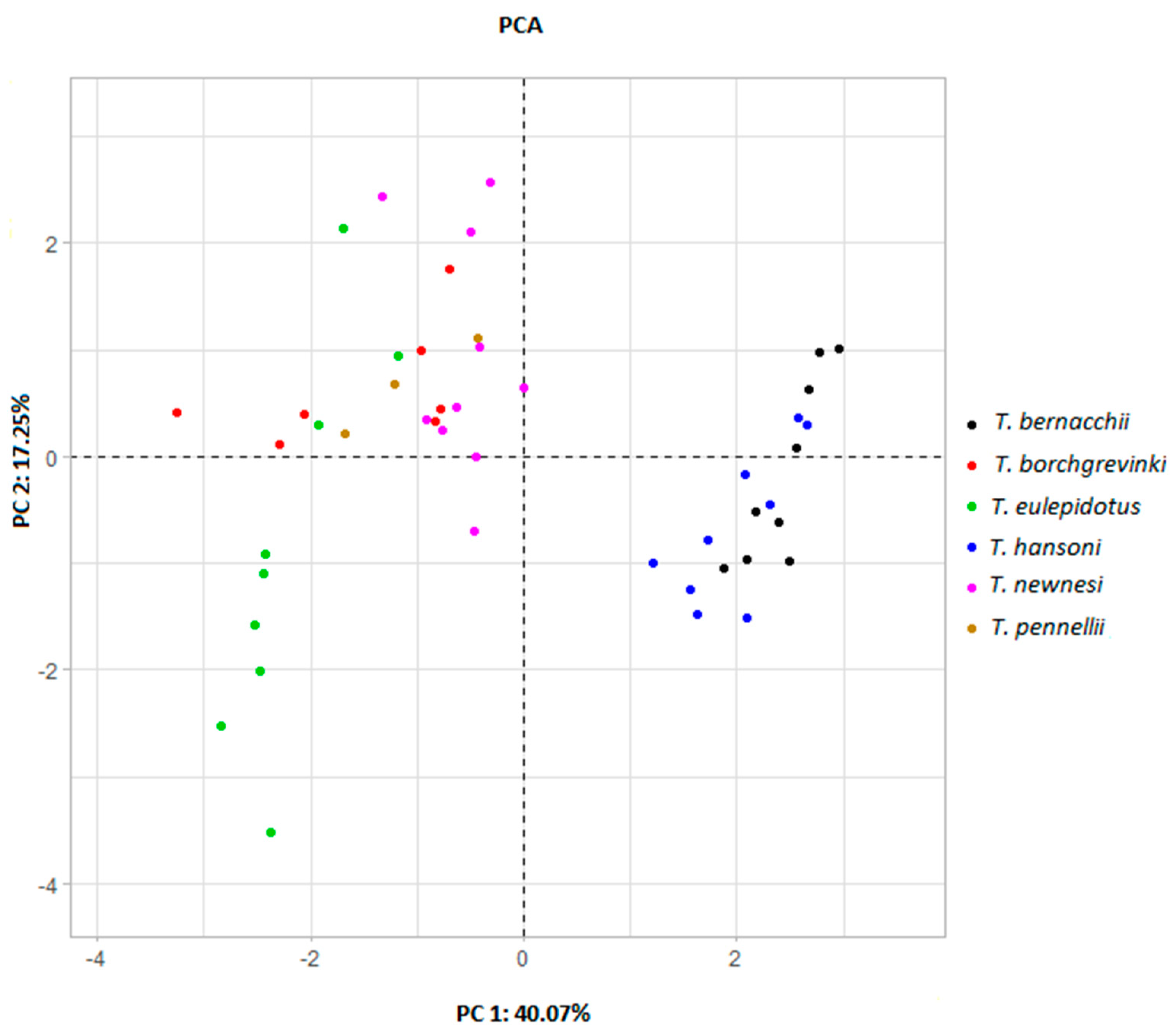
| PC1 | PC2 | PC3 | |
|---|---|---|---|
| Variance | 3.606 | 1.552 | 1.347 |
| % of Variance | 40.069 | 17.246 | 14.962 |
| Cumulative % of Variance | 40.069 | 57.315 | 72.277 |
| Traits | |||
| ED | −0.575 | 0.404 | −0.027 |
| HL | 0.271 | 0.458 | 0.607 |
| GW | −0.021 | 0.552 | 0.065 |
| BL | −0.868 | 0.361 | −0.085 |
| CSAepaxialis | 0.734 | 0.538 | −0.280 |
| LinSI | 0.714 | 0.533 | −0.393 |
| LoutSI | 0.911 | −0.130 | −0.066 |
| LinMA | 0.754 | −0.313 | −0.337 |
| LoutMA | 0.052 | 0.237 | 0.784 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carlig, E.; Di Blasi, D.; Pisano, E.; Vacchi, M.; Santovito, G.; Ghigliotti, L. Ecomorphological Differentiation of Feeding Structures within the Antarctic Fish Species Flock Trematominae (Notothenioidei) from Terra Nova Bay (Ross Sea). J. Mar. Sci. Eng. 2022, 10, 1876. https://doi.org/10.3390/jmse10121876
Carlig E, Di Blasi D, Pisano E, Vacchi M, Santovito G, Ghigliotti L. Ecomorphological Differentiation of Feeding Structures within the Antarctic Fish Species Flock Trematominae (Notothenioidei) from Terra Nova Bay (Ross Sea). Journal of Marine Science and Engineering. 2022; 10(12):1876. https://doi.org/10.3390/jmse10121876
Chicago/Turabian StyleCarlig, Erica, Davide Di Blasi, Eva Pisano, Marino Vacchi, Gianfranco Santovito, and Laura Ghigliotti. 2022. "Ecomorphological Differentiation of Feeding Structures within the Antarctic Fish Species Flock Trematominae (Notothenioidei) from Terra Nova Bay (Ross Sea)" Journal of Marine Science and Engineering 10, no. 12: 1876. https://doi.org/10.3390/jmse10121876