
Primary Composition and Pigments of 11 Red Seaweed Species from the Center of Portugal

 ,
,  ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biomass Harvesting and Processing
2.2. Yield of Moisture Content and Ash Quantification
2.3. Crude Protein Determination
2.4. Lipid Content Quantification
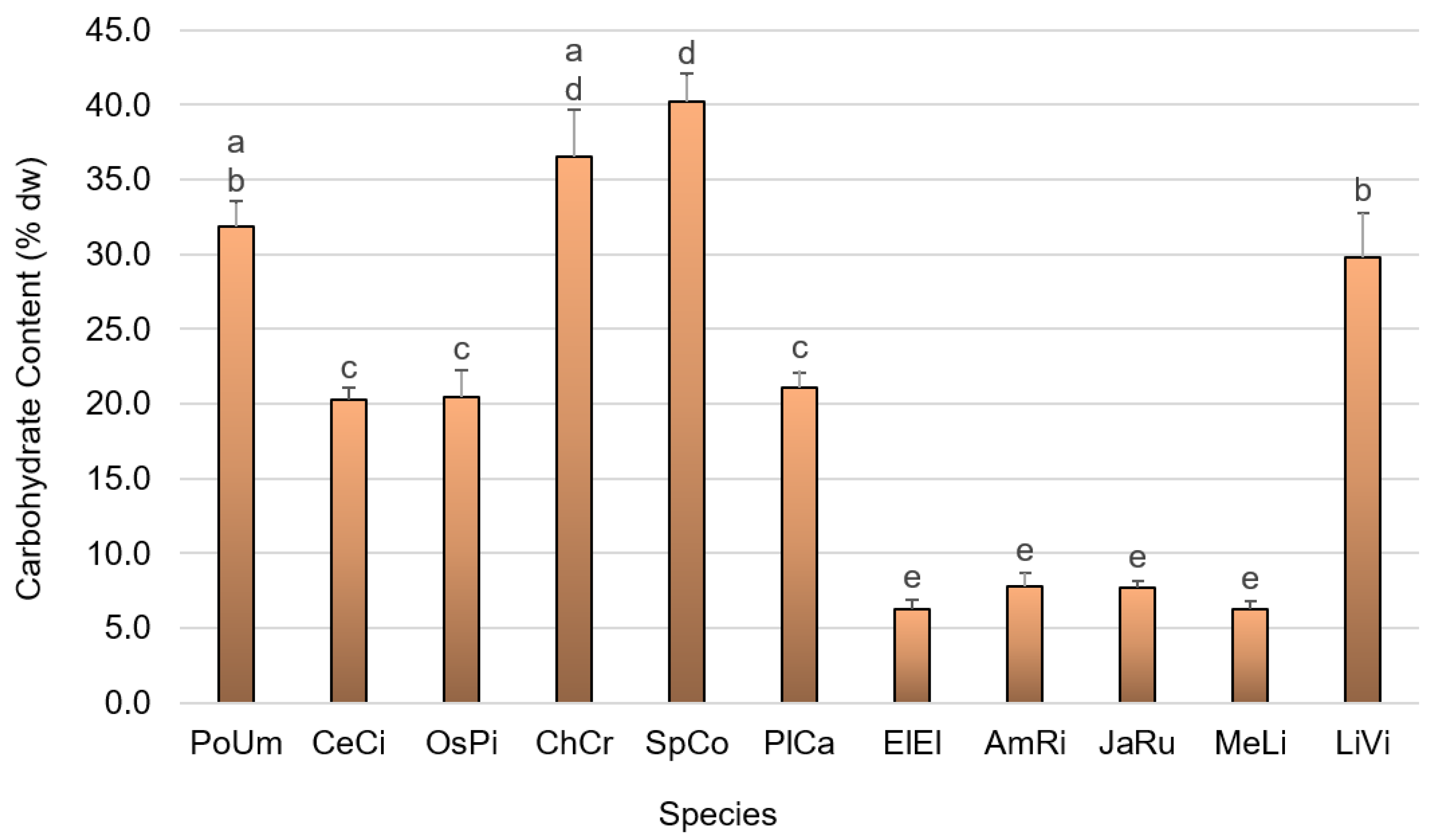
2.5. Carbohydrate Content Quantification
2.6. Pigment Content Determination
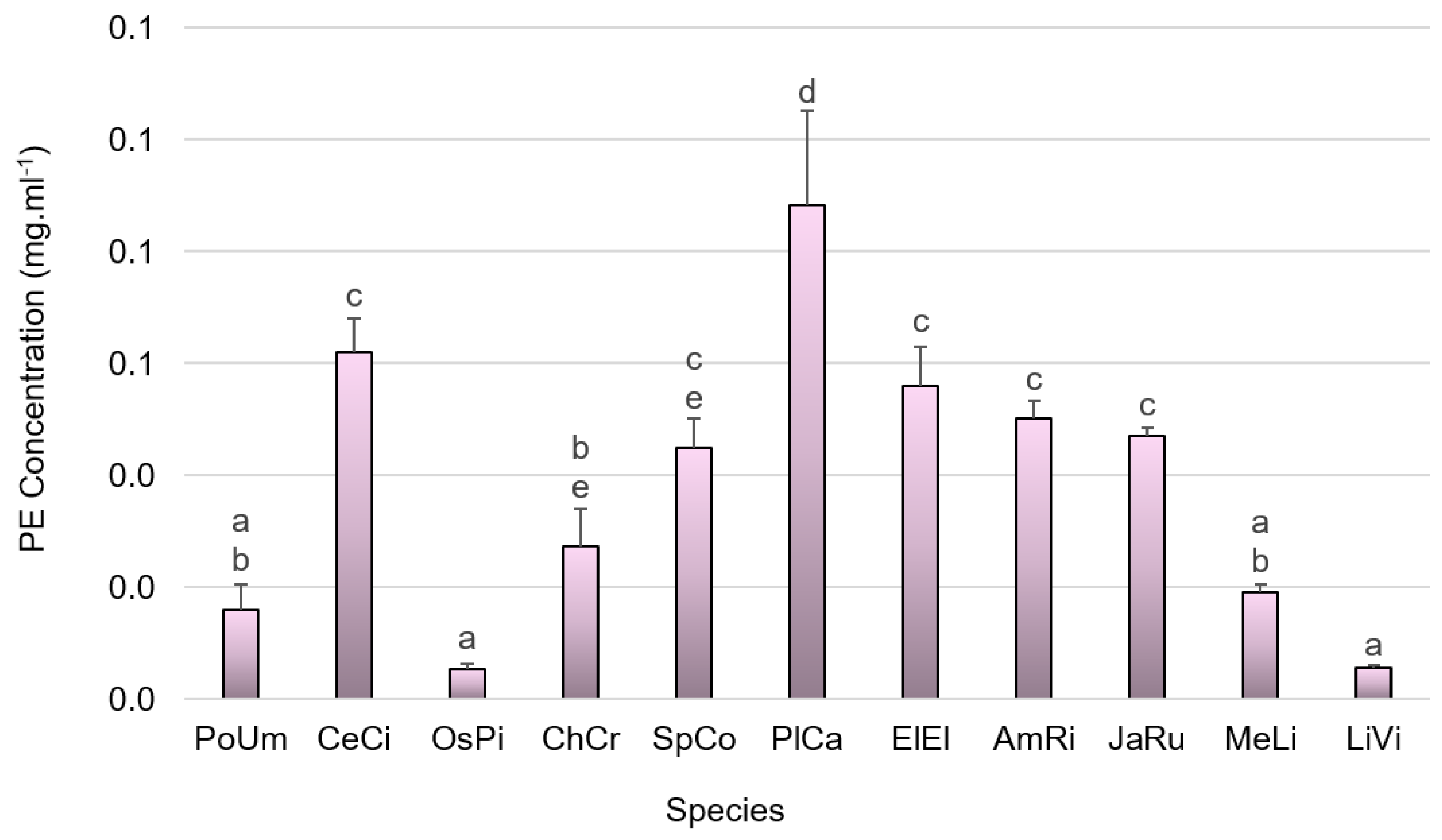
2.6.1. Phycobiliproteins
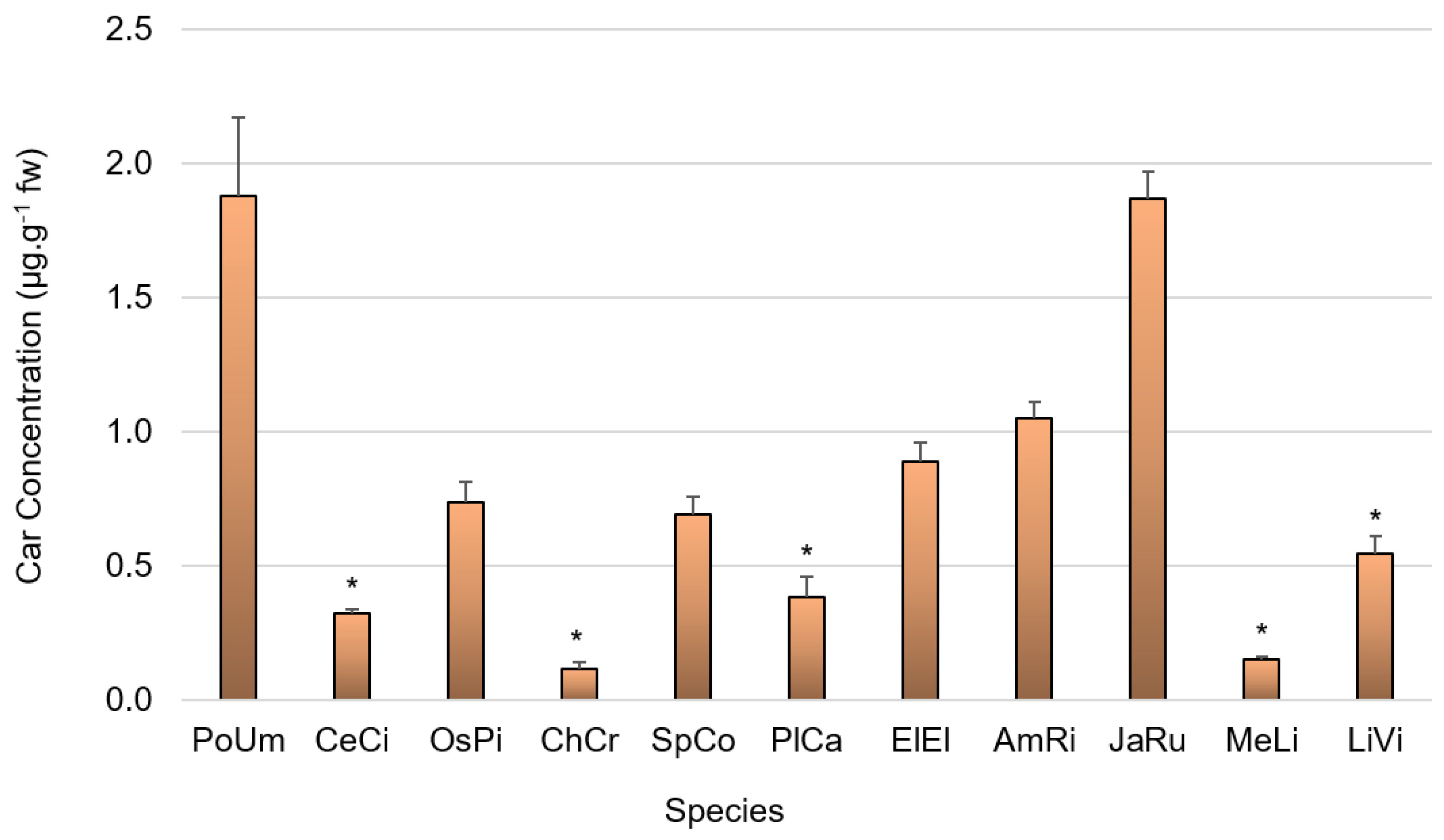
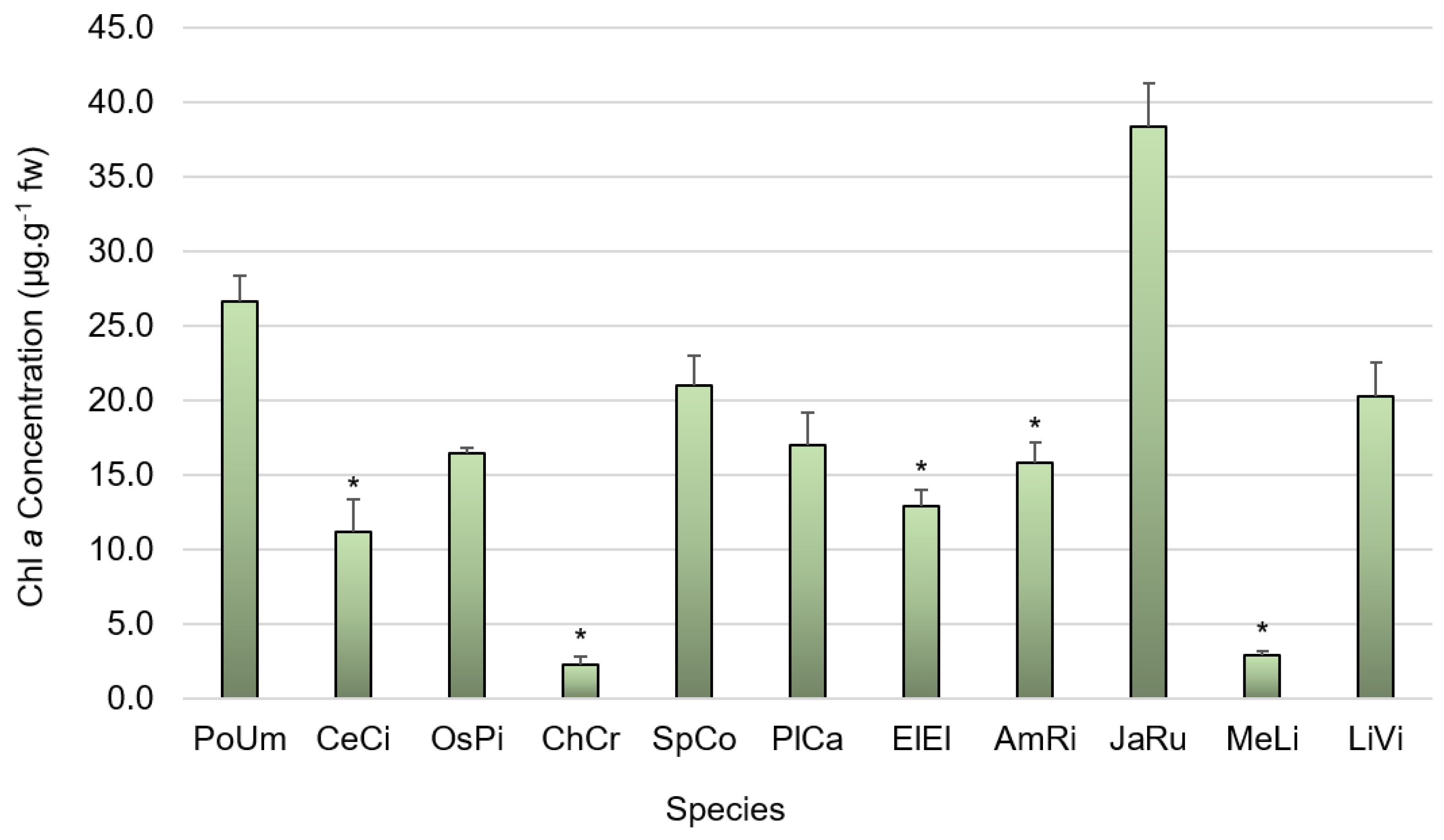
2.6.2. Carotenoid and Chlorophyll a Content Determination
2.7. Statistical Analysis
3. Results
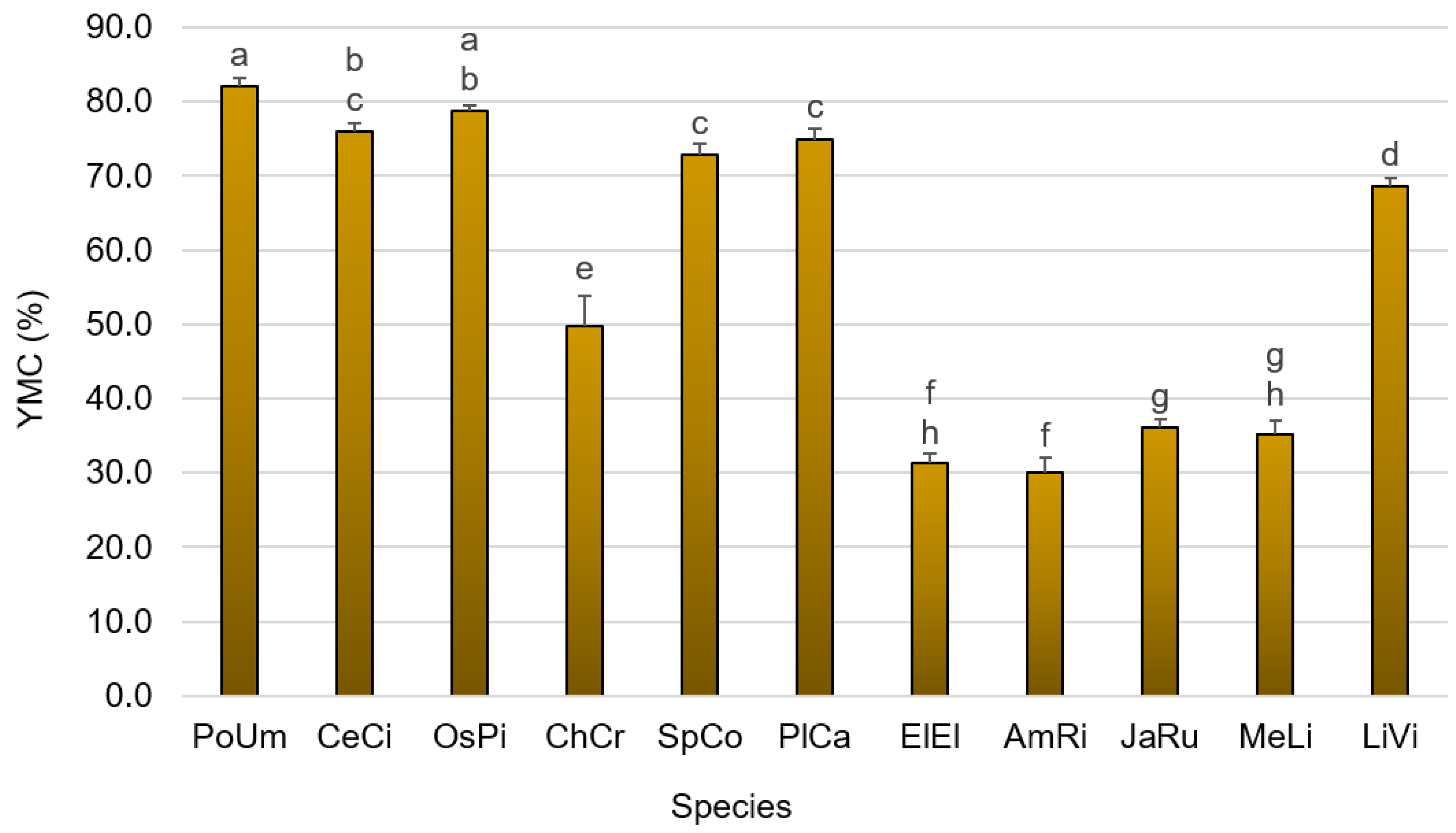
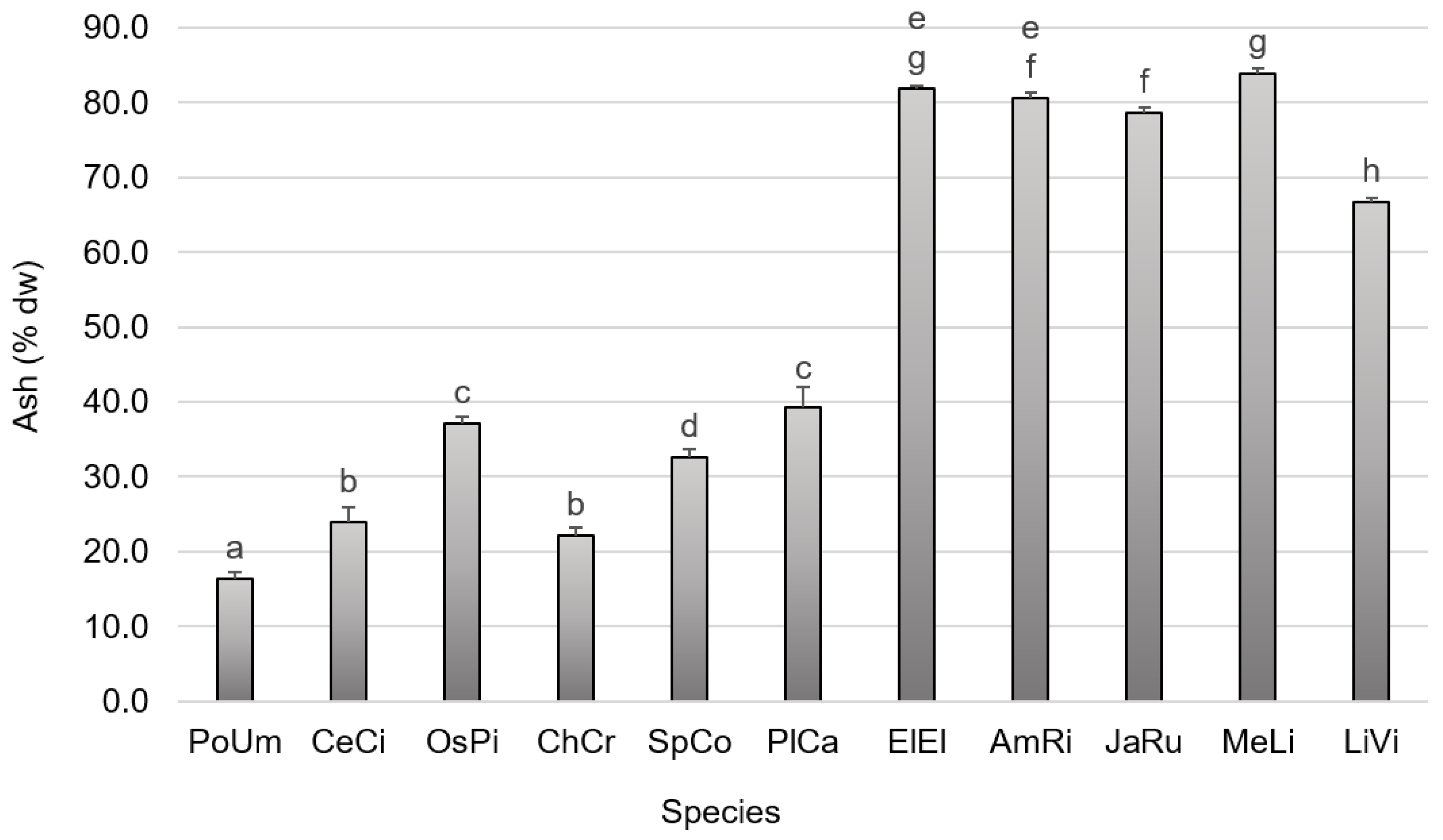
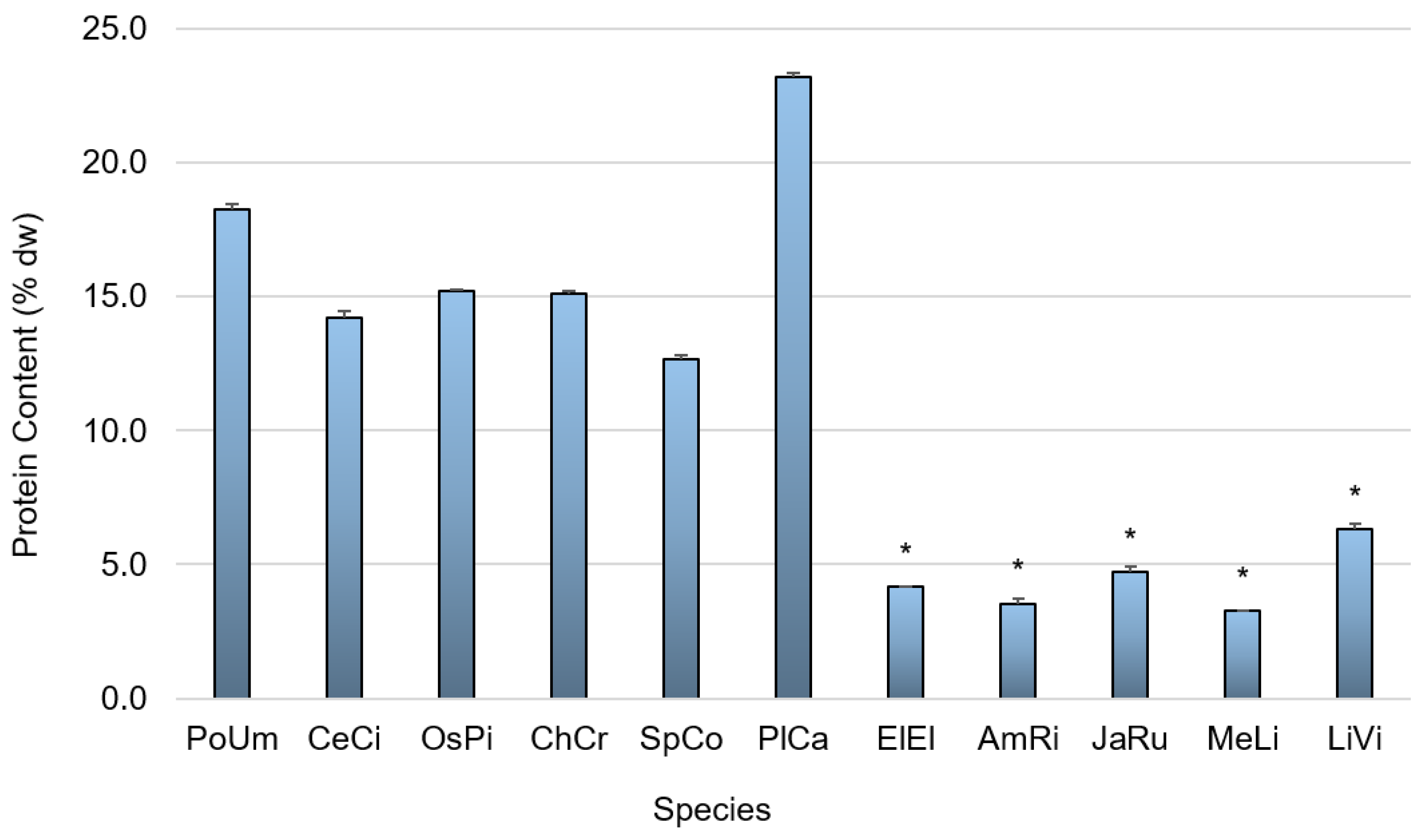
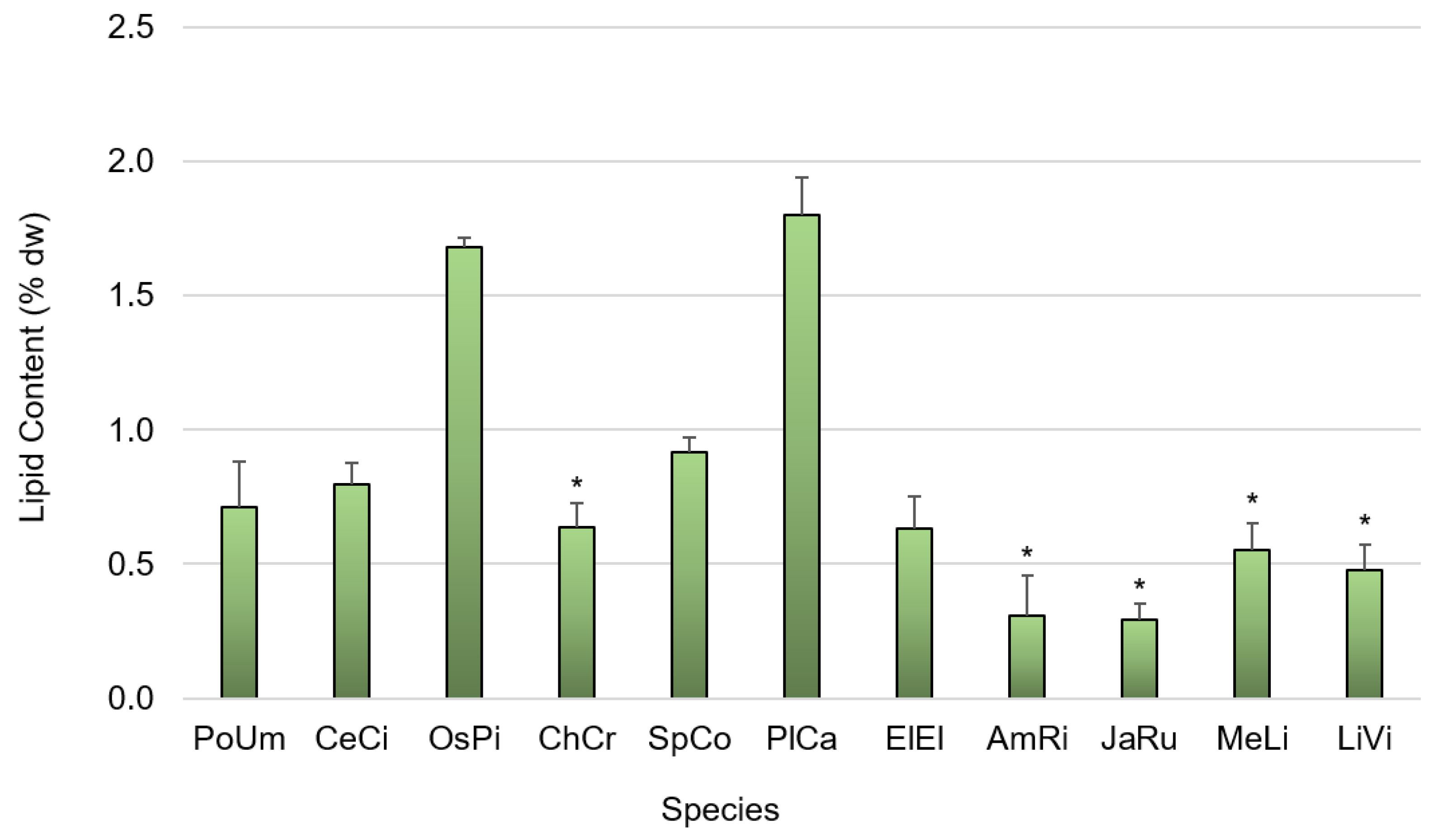
3.1. Primary Composition
3.2. Pigment Composition
4. Discussion
4.1. Primary Composition
4.1.1. Yield of Moisture Content and Ash Content
4.1.2. Protein Content
4.1.3. Lipid Content
4.1.4. Carbohydrate Content
4.2. Pigment Composition
4.2.1. Phycobiliprotein Content
4.2.2. Carotenoid Content
4.2.3. Chlorophyll a Content
4.2.4. Additional Considerations
4.3. Species-Specific Notes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ariede, M.B.; Candido, T.M.; Jacome, A.L.M.; Velasco, M.V.R.; de Carvalho, J.C.M.; Baby, A.R. Cosmetic attributes of algae—A review. Algal Res. 2017, 25, 483–487. [Google Scholar] [CrossRef]
- Hagen Rødde, R.S.; Vårum, K.M.; Larsen, B.A.; Myklestad, S.M. Seasonal and geographical variation in the chemical composition of the red alga Palmaria palmata (L.) Kuntze. Bot. Mar. 2004, 47, 125–133. [Google Scholar] [CrossRef]
- Young, R.M.; Von Salm, J.L.; Amsler, M.O.; Lopez-Bautista, J.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Site-specific variability in the chemical diversity of the Antarctic red alga Plocamium cartilagineum. Mar. Drugs 2013, 11, 2126–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeshi, S.; Yumiko, Y.-S.; Joko, S. Mineral Components and Anti-oxidant Activities of Tropical Seaweeds. J. Ocean Univ. China 2005, 4, 205–208. [Google Scholar] [CrossRef]
- Afonso, C.; Correia, A.P.; Freitas, M.V.; Baptista, T.; Neves, M.; Mouga, T. Seasonal changes in the nutritional composition of Agarophyton vermiculophyllum (Rhodophyta, Gracilariales) from the center of Portugal. Foods 2021, 10, 1145. [Google Scholar] [CrossRef]
- Cavaco, M.; Duarte, A.; Freitas, M.V.; Afonso, C.; Bernardino, S.; Pereira, L.; Martins, M.; Mouga, T. Seasonal nutritional profile of Gelidium corneum (Rhodophyta, Gelidiaceae) from the center of Portugal. Foods 2021, 10, 2394. [Google Scholar] [CrossRef]
- Kumar, K.S.; Kumari, S.; Singh, K.; Kushwaha, P. Influence of Seasonal Variation on Chemical Composition and Nutritional Profiles of Macro- and Microalgae. In Recent Advances in Micro and Macroalgal Processing; Rajauria, G., Yuan, Y.V., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2021; pp. 14–71. [Google Scholar]
- Rodrigues, D.; Freitas, A.C.; Pereira, L.; Rocha-Santos, T.A.P.; Vasconcelos, M.W.; Roriz, M.; Rodríguez-Alcalá, L.M.; Gomes, A.M.P.; Duarte, A.C. Chemical composition of red, brown and green macroalgae from Buarcos bay in Central West Coast of Portugal. Food Chem. 2015, 183, 197–207. [Google Scholar] [CrossRef]
- Kreissig, K.J.; Hansen, L.T.; Jensen, P.E.; Wegeberg, S.; Geertz-Hansen, O.; Sloth, J.J. Characterisation and chemometric evaluation of 17 elements in ten seaweed species from Greenland. PLoS ONE 2021, 16, e0243672. [Google Scholar] [CrossRef]
- Patarra, R.F.; Paiva, L.; Neto, A.I.; Lima, E.; Baptista, J. Nutritional value of selected macroalgae. J. Appl. Phycol. 2011, 23, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Nisizawa, K.; Noda, H.; Kikuchi, R.; Watanabe, T. The main seaweed foods in Japan. Hydrobiologia 1987, 151–152, 5–29. [Google Scholar] [CrossRef]
- Santos, M.A.Z.; Colepicolo, P.; Pupo, D.; Fujii, M.T.; de Pereira, C.M.P.; Mesko, M.F. Antarctic red macroalgae: A source of polyunsaturated fatty acids. J. Appl. Phycol. 2017, 29, 759–767. [Google Scholar] [CrossRef]
- Cotas, J.; Leandro, A.; Pacheco, D.; Gonçalves, A.M.M.; Pereira, L. A Comprehensive Review of the Nutraceutical and Therapeutic Applications of Red Seaweeds (Rhodophyta). Life 2020, 10, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, M.M.; Alotaibi, B.S.; EL-Sheekh, M.M. Therapeutic Uses of Red Macroalgae. Molecules 2020, 25, 4411. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L. Edible Seaweeds of the World, 1st ed.; Pereira, L., Ed.; CRC Press: Boca Raton, FL, USA, 2016; ISBN 978-1-4987-3047-1. [Google Scholar]
- Isaka, S.; Cho, K.; Nakazono, S.; Abu, R.; Ueno, M.; Kim, D.; Oda, T. Antioxidant and anti-inflammatory activities of porphyran isolated from discolored nori (Porphyra yezoensis). Int. J. Biol. Macromol. 2015, 74, 68–75. [Google Scholar] [CrossRef]
- Pereira, L. A review of the nutrient composition of selected edible seaweeds. In Seaweed: Ecology, Nutrient Composition and Medicinal Uses; Pomin, V.H., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2011; pp. 15–47. ISBN 978-1-61470-878-0. [Google Scholar]
- Mouritsen, O.G.; Dawczynski, C.; Duelund, L.; Jahreis, G.; Vetter, W.; Schröder, M. On the human consumption of the red seaweed dulse (Palmaria palmata (L.) Weber & Mohr). J. Appl. Phycol. 2013, 25, 1777–1791. [Google Scholar] [CrossRef]
- Gioele, C.; Marilena, S.; Valbona, A.; Nunziacarla, S.; Andrea, S.; Antonio, M. Gracilaria gracilis, Source of Agar: A Short Review. Curr. Org. Chem. 2017, 21, 380–386. [Google Scholar] [CrossRef]
- López-López, I.; Bastida, S.; Ruiz-Capillas, C.; Bravo, L.; Larrea, M.T.; Sánchez-Muniz, F.; Cofrades, S.; Jiménez-Colmenero, F. Composition and antioxidant capacity of low-salt meat emulsion model systems containing edible seaweeds. Meat Sci. 2009, 83, 492–498. [Google Scholar] [CrossRef]
- Marsham, S.; Scott, G.W.; Tobin, M.L. Comparison of nutritive chemistry of a range of temperate seaweeds. Food Chem. 2007, 100, 1331–1336. [Google Scholar] [CrossRef]
- Harrysson, H.; Hayes, M.; Eimer, F.; Carlsson, N.-G.; Toth, G.B.; Undeland, I. Production of protein extracts from Swedish red, green, and brown seaweeds, Porphyra umbilicalis Kützing, Ulva lactuca Linnaeus, and Saccharina latissima (Linnaeus) J. V. Lamouroux using three different methods. J. Appl. Phycol. 2018, 30, 3565–3580. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Wang, J.; Wang, S.; Xu, X. Porphyra species: A mini-review of its pharmacological and nutritional properties. J. Med. Food 2016, 19, 111–119. [Google Scholar] [CrossRef]
- Yanshin, N.; Kushnareva, A.; Lemesheva, V.; Birkemeyer, C.; Tarakhovskaya, E. Chemical Composition and Potential Practical Application of 15 Red Algal Species from the White Sea Coast (the Arctic Ocean). Molecules 2021, 26, 2489. [Google Scholar] [CrossRef] [PubMed]
- Osório, C.; Machado, S.; Peixoto, J.; Bessada, S.; Pimentel, F.B.; Alves, R.C.; Oliveira, M.B.P.P. Pigments content (Chlorophylls, fucoxanthin and phycobiliproteins) of different commercial dried algae. Separations 2020, 7, 33. [Google Scholar] [CrossRef]
- Carpena, M.; Caleja, C.; Pereira, E.; Pereira, C.; Ćirić, A.; Soković, M.; Soria-Lopez, A.; Fraga-Corral, M.; Simal-Gandara, J.; Ferreira, I.C.F.R.; et al. Red Seaweeds as a Source of Nutrients and Bioactive Compounds: Optimization of the Extraction. Chemosensors 2021, 9, 132. [Google Scholar] [CrossRef]
- Pereira, L.; Gheda, S.F.; Ribeiro-Claro, P.J.A. Analysis by Vibrational Spectroscopy of Seaweed Polysaccharides with Potential Use in Food, Pharmaceutical, and Cosmetic Industries. Int. J. Carbohydr. Chem. 2013, 2013, 1–7. [Google Scholar] [CrossRef]
- Santos, S.A.O.; Vilela, C.; Freire, C.S.R.; Abreu, M.H.; Rocha, S.M.; Silvestre, A.J.D. Chlorophyta and Rhodophyta macroalgae: A source of health promoting phytochemicals. Food Chem. 2015, 15, 122–128. [Google Scholar] [CrossRef]
- Nunes, N.; Ferraz, S.; Valente, S.; Barreto, M.C.; Pinheiro de Carvalho, M.A.A. Biochemical composition, nutritional value, and antioxidant properties of seven seaweed species from the Madeira Archipelago. J. Appl. Phycol. 2017, 29, 2427–2437. [Google Scholar] [CrossRef]
- Mendes, M.; Pereira, R.; Sousa Pinto, I.; Carvalho, A.P.; Gomes, A.M. Antimicrobial activity and lipid profile of seaweed extracts from the North Portuguese Coast. Int. Food Res. J. 2013, 20, 3337–3345. [Google Scholar]
- Rupérez, P. Mineral content of edible marine seaweeds. Food Chem. 2002, 79, 23–26. [Google Scholar] [CrossRef]
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Fatty acid contents and profiles of 16 macroalgae collected from the Irish Coast at two seasons. J. Appl. Phycol. 2014, 26, 451–463. [Google Scholar] [CrossRef]
- Lopes, D.; Melo, T.; Meneses, J.; Abreu, M.H.; Pereira, R.; Domingues, P.; Lillebø, A.I.; Calado, R.; Rosário Domingues, M. A New Look for the Red Macroalga Palmaria palmata: A seafood with polar lipids rich in EPA and with antioxidant properties. Mar. Drugs 2019, 17, 533. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Nishizawa, M.; Shimizu, Y.; Saeki, H. Anti-inflammatory effects of dulse (Palmaria palmata) resulting from the simultaneous water-extraction of phycobiliproteins and chlorophyll a. Food Res. Int. 2017, 100, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Galland-Irmouli, A.V.; Fleurence, J.; Lamghari, R.; Luçon, M.; Rouxel, C.; Barbaroux, O.; Bronowicki, J.P.; Villaume, C.; Guéant, J.L. Nutritional value of proteins from edible seaweed Palmaria palmata (Dulse). J. Nutr. Biochem. 1999, 10, 353–359. [Google Scholar] [CrossRef]
- Mishra, V.K.; Temelli, F.; Ooraikul, B.; Shacklock, P.F.; Craigie, J.S. Lipids of the Red Alga, Palmaria palmata. Bot. Mar. 2009, 36, 169–174. [Google Scholar] [CrossRef]
- De Almeida, C.L.F.; Falcão, H.D.S.; Lima, G.R.D.M.; Montenegro, C.D.A.; Lira, N.S.; de Athayde-Filho, P.F.; Rodrigues, L.C.; de Souza, M.F.V.; Barbosa-Filho, J.M.; Batista, L.M. Bioactivities from marine algae of the genus Gracilaria. Int. J. Mol. Sci. 2011, 12, 4550–4573. [Google Scholar] [CrossRef] [PubMed]
- Francavilla, M.; Franchi, M.; Monteleone, M.; Caroppo, C. The red seaweed Gracilaria gracilis as a multi products source. Mar. Drugs 2013, 11, 3754–3776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonso, C.; Correia, A.P.; Freitas, M.V.; Mouga, T.; Baptista, T. In Vitro evaluation of the antibacterial and antioxidant activities of extracts of Gracilaria gracilis with a view into its potential use as an additive in fish feed. Appl. Sci. 2021, 11, 6642. [Google Scholar] [CrossRef]
- Freitas, M.V.; Mouga, T.; Correia, A.P.; Afonso, C.; Baptista, T.; Patr, A.; Baptista, T. New Insights on the Sporulation, Germination, and Nutritional Profile of Gracilaria gracilis (Rhodophyta) Grown under Controlled Conditions. J. Mar. Sci. Eng. 2021, 9, 562. [Google Scholar] [CrossRef]
- Zepeda, E.; Freile-Pelegrín, Y.; Robledo, D. Nutraceutical assessment of Solieria filiformis and Gracilaria cornea (Rhodophyta) under light quality modulation in culture. J. Appl. Phycol. 2020, 32, 2363–2373. [Google Scholar] [CrossRef]
- Mantri, V.A.; Veeragurunthan, V.; Sambhwani, K.; Anisoddin Kazi, M. Concise review of industrially important red seaweed Gracilaria dura (C. Agardh) J. Agardh. J. Appl. Phycol. 2022, 34, 1825–1841. [Google Scholar] [CrossRef]
- Matias, M.; Pinteus, S.; Martins, A.; Silva, J.; Alves, C.; Mouga, T.; Gaspar, H.; Pedrosa, R. Gelidiales Are Not Just Agar—Revealing the Antimicrobial Potential of Gelidium corneum for Skin Disorders. Antibiotics 2022, 11, 481. [Google Scholar] [CrossRef]
- Mittal, R.; Raghavarao, K.S.M.S. Extraction of R-Phycoerythrin from marine macro-algae, Gelidium pusillum, employing consortia of enzymes. Algal Res. 2018, 34, 1–11. [Google Scholar] [CrossRef]
- Kang, M.C.; Kang, N.; Kim, S.Y.; Lima, I.S.; Ko, S.C.; Kim, Y.T.; Kim, Y.B.; Jeung, H.D.; Choi, K.S.; Jeon, Y.J. Popular edible seaweed, Gelidium amansii prevents against diet-induced obesity. Food Chem. Toxicol. 2016, 90, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.; Serrano, E.; Silva, J.; Rodrigues, C.; Pinteus, S.; Gaspar, H.; Botana, L.M.; Alpoim, M.C.; Pedrosa, R. Sphaerococcus coronopifolius bromoterpenes as potential cancer stem cell-targeting agents. Biomed. Pharmacother. 2020, 128, 110275. [Google Scholar] [CrossRef] [PubMed]
- Pinteus, S.; Alves, C.; Rodrigues, A.; Mouga, T.; Pedrosa, R. Algae from the Peniche coast (Portugal) exhibit new promising antibacterial activities against fish pathogenic bacteria. Curr. Opin. Biotechnol. 2011, 22, 33–34. [Google Scholar] [CrossRef]
- Valentão, P.; Trindade, P.; Gomes, D.; Guedes de Pinho, P.; Mouga, T.; Andrade, P.B. Codium tomentosum and Plocamium cartilagineum: Chemistry and antioxidant potential. Food Chem. 2010, 119, 1359–1368. [Google Scholar] [CrossRef]
- Alves, C.; Pinteus, S.; Rodrigues, A.; Horta, A.; Pedrosa, R. Algae from Portuguese Coast Presented High Cytotoxicity and Antiproliferative Effects on an in vitro Model of Human Colorectal Cancer. Pharmacognosy Res. 2018, 10, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.M.; Matos, J.; Afonso, C.; Gomes, R.; Bandarra, N.M.; Cardoso, C. Azorean macroalgae (Petalonia binghamiae, Halopteris scoparia and Osmundea pinnatifida) bioprospection: A study of fatty acid profiles and bioactivity. Int. J. Food Sci. Technol. 2019, 54, 880–890. [Google Scholar] [CrossRef]
- Hegazi, M.M.; Pérez-Ruzafa, A.; Almela, L.; Candela, M.E. Separation and identification of chlorophylls and carotenoids from Caulerpa prolifera, Jania rubens and Padina pavonica by reversed-phase high-performance liquid chromatography. J. Chromatogr. A 1998, 829, 153–159. [Google Scholar] [CrossRef]
- Polat, S.; Ozogul, Y. Seasonal proximate and fatty acid variations of some seaweeds from the northeastern Mediterranean coast. Oceanologia 2013, 55, 375–391. [Google Scholar] [CrossRef] [Green Version]
- Mohy El-Din, S.M.; El-Ahwany, A.M.D. Bioactivity and phytochemical constituents of marine red seaweeds (Jania rubens, Corallina mediterranea and Pterocladia capillacea). J. Taibah Univ. Sci. 2016, 10, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Aslam, M.N.; Kreider, J.M.; Paruchuri, T.; Bhagavathula, N.; DaSilva, M.; Zernicke, R.F.; Goldstein, S.A.; Varani, J. A mineral-rich extract from the red marine algae Lithothamnion calcareum preserves bone structure and function in female mice on a western-style diet. Calcif. Tissue Int. 2010, 86, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, O.; Stenson, B.; Widaa, A.; O′Gorman, D.M.; O′Brien, F.J. Incorporation of the natural marine multi-mineral dietary supplement Aquamin enhances osteogenesis and improves the mechanical properties of a collagen-based bone graft substitute. J. Mech. Behav. Biomed. Mater. 2015, 47, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Pereira Da Silva, R.; Sayuri, G.; Kawai, D.; Dias De Andrade, F.R.; Danilo, V.; Bezzon, N.; Gomes Ferraz, H. Characterisation and Traceability of Calcium Carbonate from the Seaweed Lithothamnium calcareum. Solids 2021, 2, 192–211. [Google Scholar] [CrossRef]
- Cavaco, M.; Duarte, A.; Bernardino, S.; Afonso, C.; Mouga, T. Sustainable use of seaweeds from S. Martinho do Porto Portugal – past present and future perspective. In Proceedings of the International Conference on Water, Energy, Food and Sustainability; Galvão, J., Ed.; Springer: Leiria, Portugal, 2021; pp. 1–10. [Google Scholar]
- Santos, R.; Duarte, P. Marine plant harvest in Portugal. J. Appl. Phycol. 1991, 3, 11–18. [Google Scholar] [CrossRef]
- Mabeau, S.; Fleurence, J. Seaweed in Food Products: Biochemical and Nutritional Aspects. Trends Food Sci. Technol. 1993, 4, 103–107. [Google Scholar] [CrossRef]
- IMARC Group. Seaweed Market: Global Industry Trends, Share, Size, Growth, Opportunity and Forecast 2022–2027; IMARC: Uttar Pradesh, India, 2022. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 20th ed.; Latimer, G.W., Jr., Ed.; Association of Official Analytical Chemists: Rockville, Maryland, 2016; ISBN 0935584870. [Google Scholar]
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern. J. Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef] [Green Version]
- Angell, A.R.; Mata, L.; de Nys, R.; Paul, N.A. The protein content of seaweeds: A universal nitrogen-to-protein conversion factor of five. J. Appl. Phycol. 2016, 28, 511–524. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Dumay, J.; Morançais, M.; Nguyen, H.P.T.; Fleurence, J. Extraction and Purification of R-phycoerythrin from Marine Red Algae. In Natural Products From Marine Algae. Methods in Molecular Biology; Stengel, D., Connan, S., Eds.; Humana Press: New York, NY, USA, 2015; Volume 1308. [Google Scholar]
- Beattie, S.W.; Morançais, M.; Déléris, P.; Fleurence, J.; Dumay, J. Extraction of phycocyanin and phycoerythrin pigments. In Protocols for Macroalgae Research; Charrier, B., Wichard, T., Reddy, C.R.K., Eds.; CRC Press: Boca Raton, FL, USA, 2018; pp. 249–265. [Google Scholar]
- Beer, S.; Eshel, A. Determining Phycoerythrin and Phycocyanin Concentrations in Aqueous Crude Extracts of Red Algae. Mar. Freshw. Res. 1985, 36, 785–792. [Google Scholar] [CrossRef]
- Román, R.B.; Alvárez-Pez, J.M.; Fernández, F.G.A.; Grima, E.M. Recovery of pure B-phycoerythrin from the microalga Porphyridium cruentum. J. Biotechnol. 2002, 93, 73–85. [Google Scholar] [CrossRef]
- Kirk, J.T.O.; Allen, R.L. Dependence of chloroplast pigment synthesis on protein synthesis: Effect of actidione. Biochem. Biophys. Res. Commun. 1965, 21, 523–530. [Google Scholar] [CrossRef]
- Ritchie, R.J. Universal chlorophyll equations for estimating chlorophylls a, b, c, and d and total chlorophylls in natural assemblages of photosynthetic organisms using acetone, methanol, or ethanol solvents. Photosynthetica 2008, 46, 115–126. [Google Scholar] [CrossRef]
- Pereira, L. Nutritional Composition of the Main Edible Algae. In Therapeutic and Nutritional Uses of Algae; Pereira, L., Ed.; CRC Press: Boca Raton, FL, 2018; pp. 65–127. [Google Scholar]
- Cofrades, S.; López-López, I.; Bravo, L.; Ruiz-Capillas, C.; Bastida, S.; Larrea, M.T.; Jiménez-Colmenero, F. Nutritional and Antioxidant Properties of Different Brown and Red Spanish Edible Seaweeds. Food Sci. Technol. Int. 2010, 16, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Paiva, L.; Lima, E.; Patarra, R.F.; Neto, A.I.; Baptista, J. Edible Azorean macroalgae as source of rich nutrients with impact on human health. Food Chem. 2014, 164, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Frikha, F.; Kammoun, M.; Hammami, N.; Mchirgui, R.; Belbahri, L.; Gargouri, Y.; Miled, N.; Ben–Rebah, F. Composición química y algunas actividades biológicas de algas marinas recolectadas en Túnez. Ciencias Mar. 2011, 37, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Rupérez, P.; Saura-Calixto, F.; Rupérez, P.; Saura-Calixto, F. Dietary fibre and physicochemical properties of edible Spanish seaweeds. Eur. Food Res. Technol. 2001, 212, 349–354. [Google Scholar] [CrossRef]
- Fleurence, J. Seaweed proteins: Biochemical, nutritional aspects and potential uses. Trends Food Sci. Technol. 1999, 10, 25–28. [Google Scholar] [CrossRef]
- Sánchez-Machado, D.I.; López-Cervantes, J.; López-Hernández, J.; Paseiro-Losada, P. Fatty acids, total lipid, protein and ash contents of processed edible seaweeds. Food Chem. 2004, 85, 439–444. [Google Scholar] [CrossRef]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Lordan, S.; Ross, R.P.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamero-Vega, G.; Palacios-Palacios, M.; Quitral, V. Nutritional Composition and Bioactive Compounds of Red Seaweed: A Mini-Review. J. Food Nutr. Res. 2020, 8, 431–440. [Google Scholar] [CrossRef]
- Bocanegra, A.; Bastida, S.; Benedi, J.; Rodenas, S.; Sanchez-Muniz, F.J. Characteristics and Nutritional and Cardiovascular-Health Properties of Seaweeds. J. Med. Food 2013, 12, 236–258. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L. Therapeutic Uses of Phycocolloids. In Therapeutic and Nutritional Uses of Algae; Pereira, L., Ed.; CRC Press: Boca Raton, FL, USA, 2017; pp. 128–142. ISBN 9781315152844. [Google Scholar]
- Li, W.; Su, H.N.; Pu, Y.; Chen, J.; Liu, L.N.; Liu, Q.; Qin, S. Phycobiliproteins: Molecular structure, production, applications, and prospects. Biotechnol. Adv. 2019, 37, 340–353. [Google Scholar] [CrossRef] [PubMed]
- Fleurence, J. R-phycoerythrin from red macroalgae: Strategies for extraction and potential application in biotechnology. Appl. Biotechnol. Food Sci. Policy 2003, 1, 1–6. [Google Scholar]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef]
- Dagnino-Leone, J.; Figueroa, C.P.; Castañeda, M.L.; Youlton, A.D.; Vallejos-Almirall, A.; Agurto-Muñoz, A.; Pavón Pérez, J.; Agurto-Muñoz, C. Phycobiliproteins: Structural aspects, functional characteristics, and biotechnological perspectives. Comput. Struct. Biotechnol. J. 2022, 20, 1506–1527. [Google Scholar] [CrossRef]
- Fernández-Rojas, B.; Hernández-Juárez, J.; Pedraza-Chaverri, J. Nutraceutical properties of Phycocyanin. J. Funct. Foods 2014, 11, 375–392. [Google Scholar] [CrossRef]
- Eriksen, N.T. Production of phycocyanin - A pigment with applications in biology, biotechnology, foods and medicine. Appl. Microbiol. Biotechnol. 2008, 80, 1–14. [Google Scholar] [CrossRef]
- Ramu Ganesan, A.; Kannan, M.; Karthick Rajan, D.; Pillay, A.; Shanmugam, M.; Sathishkumar, P.; Johansen, J.; Tiwari, B.K. Phycoerythrin: A pink pigment from red sources (Rhodophyta) for a greener biorefining approach to food applications. Crit. Rev. Food Sci. Nutr. 2022, 0, 1–19. [Google Scholar] [CrossRef]
- Kannaujiya, V.K.; Kumar, D.; Richa, J.P.; Sonker, A.S.; Rajneesh, V.S.; Sundaram, S.; Sinha, R.P. Recent advances in production and the biotechnological significance of phycobiliproteins. In New Approaches in Biological Research; Sinha, R.P., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2017. [Google Scholar]
- Rapoport, A.; Guzhova, I.; Bernetti, L.; Buzzini, P.; Kieliszek, M.; Kot, A.M. Carotenoids and some other pigments from fungi and yeasts. Metabolites 2021, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Martínez, A.J.; Mandić, A.I.; Bantis, F.; Böhm, V.; Borge, G.I.A.; Brnčić, M.; Bysted, A.; Cano, M.P.; Dias, M.G.; Elgersma, A.; et al. A comprehensive review on carotenoids in foods and feeds: Status quo, applications, patents, and research needs. Crit. Rev. Food Sci. Nutr. 2020, 5, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Pangestuti, R.; Kim, S.K. Biological activities and health benefit effects of natural pigments derived from marine algae. J. Funct. Foods 2011, 3, 255–266. [Google Scholar] [CrossRef]
- Soni, B.; Kalavadia, B.; Trivedi, U.; Madamwar, D. Extraction, purification and characterization of phycoerythrin from Michrochaete and its biological activities. Biocatal. Agric. Biotechnol. 2018, 13, 84–89. [Google Scholar] [CrossRef]
- Yabuta, Y.; Fujimura, H.; Kwak, C.S.; Enomoto, T.; Watanabe, F. Antioxidant activity of the phycoerythrobilin compound formed from a dried Korean purple laver (Porphyra sp.) during in vitro digestion. Food Sci. Technol. Res. 2010, 16, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Nagaraj, S.; Arulmurugan, P.; Rajaram, M.G.; Karuppasamy, K.; Jayappriyan, K.R.; Sundararaj, R.; Vijayanand, N.; Rengasamy, R. Hepatoprotective and antioxidative effects of C-phycocyanin from Arthrospira maxima SAG 25780 in CCl4-induced hepatic damage rats. Biomed. Prev. Nutr. 2012, 2, 81–85. [Google Scholar] [CrossRef]
- Lanfer-Marquez, U.M.; Barros, R.M.C.; Sinnecker, P. Antioxidant activity of chlorophylls and their derivatives. Food Res. Int. 2005, 38, 885–891. [Google Scholar] [CrossRef]
- Arun Kumar, S.; Brown, L. Seaweeds as potential therapeutic interventions for the metabolic syndrome. Rev. Endocr. Metab. Disord. 2013, 14, 299–308. [Google Scholar] [CrossRef]
- Cikoš, A.-M.; Šubarić, D.; Roje, M.; Babić, J.; Jerković, I.; Jokić, S. Recent advances on macroalgal pigments and their biological activities (2016–2021). Algal Res. 2022, 65, 102748. [Google Scholar] [CrossRef]
- Lee, H.G.; Lu, Y.A.; Je, J.G.; Jayawardena, T.U.; Kang, M.C.; Lee, S.H.; Kim, T.H.; Lee, D.S.; Lee, J.M.; Yim, M.J.; et al. Effects of Ethanol Extracts from Grateloupia elliptica, a Red Seaweed, and Its Chlorophyll Derivative on 3T3-L1 Adipocytes: Suppression of Lipid Accumulation through Downregulation of Adipogenic Protein Expression. Mar. Drugs 2021, 19, 91. [Google Scholar] [CrossRef]
- Kannaujiya, V.K.; Kumar, D.; Singh, V.; Sinha, R.P. Advances in Phycobiliproteins Research: Innovations and Commercialization. In Natural Bioactive Compounds: Technological Advancements; Sinha, R.P., Häder, D.-P., Eds.; Academic Press: London, UK, 2021; pp. 57–81. [Google Scholar]
- Niu, J.; Xu, M.; Wang, G.; Zhang, K.; Peng, G. Comprehensive extraction of agar and R-phycoerythrin from Gracilaria lemaneiformis (Bangiales, Rhodophyta). Indian J. Mar. Sci. 2013, 42, 21–28. [Google Scholar]
- Leney, A.C.; Tschanz, A.; Heck, A.J.R. Connecting color with assembly in the fluorescent B-phycoerythrin protein complex. FEBS J. 2018, 285, 178–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bei, H.; Guang-Ce, W.; Chen-Kui, Z.; Zhen-Gang, L. The experimental research of R-phycoerythrin subunits on cancer treatment: A new photosensitizer in PDT. Cancer Biother. Radiopharm. 2002, 17, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Dagnino-Leone, J.; Figueroa, M.; Uribe, E.; Hinrichs, M.V.; Ortiz-López, D.; Martínez-Oyanedel, J.; Bunster, M. Biosynthesis and characterization of a recombinant eukaryotic allophycocyanin using prokaryotic accessory enzymes. Microbiologyopen 2020, 9, e989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, L. Guia ilustrado das macroalgas: Conhecer e reconhecer algumas espécies da flora portuguesa; Pereira, L., Ed.; Imprensa da Universidade de Coimbra: Coimbra, Portugal, 2009; ISBN 978-989-26-0397-1. [Google Scholar]
- Huang, D.; Boxin, O.U.; Prior, R.L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Kim, J.K.; Kraemer, G.P.; Neefus, C.D.; Chung, I.K.; Yarish, C. Effects of temperature and ammonium on growth, pigment production and nitrogen uptake by four species of Porphyra (Bangiales, Rhodophyta) native to the New England coast. J. Appl. Phycol. 2007, 19, 431–440. [Google Scholar] [CrossRef]
- Spectral Data for B-PE, R-PE and APC—Table 6.2 | Thermo Fisher Scientific—PT. Available online: https://www.thermofisher.com/pt/en/home/references/molecular-probes-the-handbook/tables/spectral-data-for-b-pe-r-pe-and-apc.html (accessed on 29 June 2022).
- Kaixian, Q.; Franklin, M.; Borowitzka, M.A. The study for isolation and purification of R-phycoerythrin from a red alga. Appl. Biochem. Biotechnol. 1993, 43, 133–139. [Google Scholar] [CrossRef]
- Saluri, M.; Kaldmäe, M.; Tuvikene, R. Reliable quantification of R-phycoerythrin from red algal crude extracts. J. Appl. Phycol. 2020, 32, 1421–1428. [Google Scholar] [CrossRef]
- Ismail, M.M.; Osman, M.E.H. Seasonal fluctuation of photosynthetic pigments of most common red seaweeds species collected from Abu Qir, Alexandria, Egypt. Rev. Biol. Mar. Oceanogr. 2016, 51, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Bhat, I.; Haripriya, G.; Jogi, N.; Mamatha, B.S. Carotenoid composition of locally found seaweeds of Dakshina Kannada district in India. Algal Res. 2021, 53, 102154. [Google Scholar] [CrossRef]
- Bjørnland, T.; Aguilar-Martinez, M. Carotenoids in red algae. Phytochemistry 1976, 15, 291–296. [Google Scholar] [CrossRef]
- Takaichi, S.; Yokoyama, A.; Mochimaru, M.; Uchida, H.; Murakami, A. Carotenogenesis diversification in phylogenetic lineages of Rhodophyta. J. Phycol. 2016, 52, 329–338. [Google Scholar] [CrossRef]
- Esteban, R.; Martínez, B.; Fernández-Marín, B.; Becerril, J.M.; García-Plazaola, J.I. Carotenoid composition in Rhodophyta: Insights into xanthophyll regulation in Corallina elongata. Eur. J. Phycol. 2009, 44, 221–230. [Google Scholar] [CrossRef]
- Halim, R.; Hosikian, A.; Lim, S.; Danquah, M.K. Chlorophyll Extraction from Microalgae: A Review on the Process Engineering Aspects. Int. J. Chem. Eng. 2010, 2010, 391632. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Roca, M. In Vitro bioavailability of chlorophyll pigments from edible seaweeds. J. Funct. Foods 2018, 41, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Cikoš, A.M.; Flanjak, I.; Bojanić, K.; Babić, S.; Čižmek, L.; Čož-Rakovac, R.; Jokić, S.; Jerković, I. Bioprospecting of Coralline Red Alga Amphiroa rigida J.V. Lamouroux: Volatiles, Fatty Acids and Pigments. Molecules 2021, 26, 520. [Google Scholar] [CrossRef]
- Yang, F.; Wei, Z.; Long, L. Transcriptomic and Physiological Responses of the Tropical Reef Calcified Macroalga Amphiroa fragilissima to Elevated Temperature. J. Phycol. 2021, 57, 1254–1265. [Google Scholar] [CrossRef]
- Brawley, S.H.; Blouin, N.A.; Ficko-Blean, E.; Wheeler, G.L.; Lohr, M.; Goodson, H.V.; Jenkins, J.W.; Blaby-Haas, C.E.; Helliwell, K.E.; Chan, C.X.; et al. Insights into the red algae and eukaryotic evolution from the genome of Porphyra umbilicalis (Bangiophyceae, Rhodophyta). Proc. Natl. Acad. Sci. USA 2017, 114, E6361–E6370. [Google Scholar] [CrossRef] [Green Version]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Paiva, L.; Patarra, R.; Neto, A.; Lima, E.; Baptista, J. Antioxidant activity of macroalgae from the Azores. Arquipelago. Life Mar. Sci. 2012, 29, 1–6. [Google Scholar]
- Pinteus, S.; Alves, C.; Monteiro, H.; Araújo, E.; Horta, A.; Pedrosa, R. Asparagopsis armata and Sphaerococcus coronopifolius as a natural source of antimicrobial compounds. World J. Microbiol. Biotechnol. 2015, 31, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.; Alves, C.; Horta, A.; Pinteus, S.; Silva, J.; Culioli, G.; Thomas, O.P.; Pedrosa, R. Antitumor and antimicrobial potential of bromoditerpenes isolated from the red alga, Sphaerococcus coronopifolius. Mar. Drugs 2015, 13, 713–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Figueroa, F.; Niell, F.X. Effects of light quality on chlorophyll and biliprotein accumulation in seaweeds. Mar. Biol. 1990, 104, 321–327. [Google Scholar] [CrossRef]
- Williamson, C.J.; Brodie, J.; Goss, B.; Yallop, M.; Lee, S.; Perkins, R. Corallina and Ellisolandia (Corallinales, Rhodophyta) photophysiology over daylight tidal emersion: Interactions with irradiance, temperature and carbonate chemistry. Mar. Biol. 2014, 161, 2051–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, W.A. Calcified macroalgae critical to coastal ecosystems and vulnerable to change: A review. Mar. Freshw. Res. 2009, 60, 787–801. [Google Scholar] [CrossRef]
- Adluri, R.S.; Zhan, L.; Bagchi, M.; Maulik, N.; Maulik, G. Comparative effects of a novel plant-based calcium supplement with two common calcium salts on proliferation and mineralization in human osteoblast cells. Mol. Cell. Biochem. 2010, 340, 73–80. [Google Scholar] [CrossRef]
- Frestedt, J.L.; Kuskowski, M.A.; Zenk, J.L. A natural seaweed derived mineral supplement (Aquamin F) for knee osteoarthritis: A randomised, placebo controlled pilot study. Nutr. J. 2009, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.H.T.; Jung, W.K.; Kim, S.K. Marine algae possess therapeutic potential for CA-mineralization via osteoblastic differentiation. Adv. Food Nutr. Res. 2011, 64, 429–441. [Google Scholar] [CrossRef]
- Walsh, P.J.; Buchanan, F.J.; Dring, M.; Maggs, C.; Bell, S.; Walker, G.M. Low-pressure synthesis and characterisation of hydroxyapatite derived from mineralise red algae. Chem. Eng. J. 2008, 137, 173–179. [Google Scholar] [CrossRef]
- Sayın, S. Antimicrobial Activities of Some Marine Macroalgae Species from Iskenderun Bay. Turkish J. Agric-Food Sci. Technol. 2021, 9, 792–796. [Google Scholar] [CrossRef]
- Caria Mendes, M.; Navalho, S.; Ferreira, A.; Paulino, C.; Figueiredo, D.; Silva, D.; Gao, F.; Gama, F.; Bombo, G.; Jacinto, R.; et al. Algae as Food in Europe: An Overview of Species Diversity and Their Application. Foods 2022, 11, 1871. [Google Scholar] [CrossRef] [PubMed]
- European Commision EU Novel Food Catalogue. Available online: https://webgate.ec.europa.eu/fip/novel_food_catalogue/ (accessed on 29 June 2022).
- Freitas, M.V.; Pereira, L.; Afonso, C.; Mouga, T. Sabores do Mar: Algas Edíveis do Centro de Portugal; Politécnico de Leiria: Leiria, Portugal, 2021; ISBN 9789895339006. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Order | Code | Harvest Season | Coordinates |
|---|---|---|---|---|
| Porphyra umbilicalis (Linnaeus) J.Agadh | Bangiales | PoUm | Winter | 39°19′10″ N, 9°21′24″ W |
| Ceramium ciliatum (J.Ellis) Ducluzeau | Ceramiales | CeCi | Summer | 39°19′27″ N, 9°21′39″ W |
| Osmundea pinnatifida (Hudson) Stackhouse | Ceramiales | OsPi | Winter | 39°22′07″ N, 9°22′41″ W |
| Chondrus crispus Stackhouse | Gigartinales | ChCr | Spring | 40°09′57″ N, 8°53′05″ W |
| Sphaerococcus coronopifolius Stackhouse | Gigartinales | SpCo | Summer | 39°22′3″ N, 9°22′26″ W |
| Plocamium cartilagineum (Linnaeus) P.S.Dixon | Plocamiales | PlCa | Winter | 39°19′10″ N, 9°21′24″ W |
| Ellisolandia elongata (J.Ellis & Solander) K.R.Hind & G.W.Saunders | Corallinales | ElEl | Winter | 39°19′10″ N, 9°21′24″ W |
| Amphiroa rigida J.V.Lamouroux | Corallinales | AmRi | Winter | 39°19′10″ N, 9°21′24″ W |
| Jania rubens (Linnaeus) J.V.Lamouroux | Corallinales | JaRu | Winter | 39°19′10″ N, 9°21′24″ W |
| Mesophyllum lichenoides (J.Ellis) Me.Lemoine | Hapalidiales | MeLi | Winter | 39°19′10″ N, 9°21′24″ W |
| Liagora viscida (Forsskål) C.Agardh | Nemaliales | LiVi | Summer | 39°22′3″ N, 9°22′26″ W |
| Species | PC (mg.mL−1) |
|---|---|
| Porphyra umbilicalis | 0.009 ± 0.003 |
| Ceramium ciliatum | 0.006 ± 0.001 |
| Osmundea pinnatifida | 0.008 ± 0.001 |
| Chondrus crispus | 0.004 ± 0.001 |
| Sphaerococcus coronopifolius | 0.001 ± 0.000 |
| Plocamium cartilagineum | 0.006 ± 0.001 |
| Ellisolandia elongata | 0 |
| Amphiroa rigida | 0 |
| Jania rubens | 0 |
| Mesophyllum lichenoides | 0 |
| Liagora viscida | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freitas, M.V.; Inácio, L.G.; Martins, M.; Afonso, C.; Pereira, L.; Mouga, T. Primary Composition and Pigments of 11 Red Seaweed Species from the Center of Portugal. J. Mar. Sci. Eng. 2022, 10, 1168. https://doi.org/10.3390/jmse10091168
Freitas MV, Inácio LG, Martins M, Afonso C, Pereira L, Mouga T. Primary Composition and Pigments of 11 Red Seaweed Species from the Center of Portugal. Journal of Marine Science and Engineering. 2022; 10(9):1168. https://doi.org/10.3390/jmse10091168
Chicago/Turabian StyleFreitas, Marta V., Leonardo G. Inácio, Mendelson Martins, Clélia Afonso, Leonel Pereira, and Teresa Mouga. 2022. "Primary Composition and Pigments of 11 Red Seaweed Species from the Center of Portugal" Journal of Marine Science and Engineering 10, no. 9: 1168. https://doi.org/10.3390/jmse10091168