Genetic Variation of the Host Plant Species Matters for Interactions with Above- and Belowground Herbivores

Abstract
:1. Introduction
2. Experimental Section
2.1. Model Plant
2.2. Study Organisms
2.3. Herbivore Treatment
2.4. Root Material and Sample Extraction for Total Protein and PI Analysis
2.5. Total Protein Content Measurement
2.6. Protease Inhibitor Measurement
2.7. FT-IR Spectroscopy and Sample Preparation
2.8. Chemometrics
2.9. Carbon and Nitrogen Concentration Measurement
2.10. Choice and Non-Choice Feeding Bioassay
2.11. Statistical Analysis
3. Results and Discussion
3.1. Results
3.1.1. Herbivore Performance
3.1.1.1. Spodoptera Exigua

3.1.1.2. Agriotes spp. Larvae

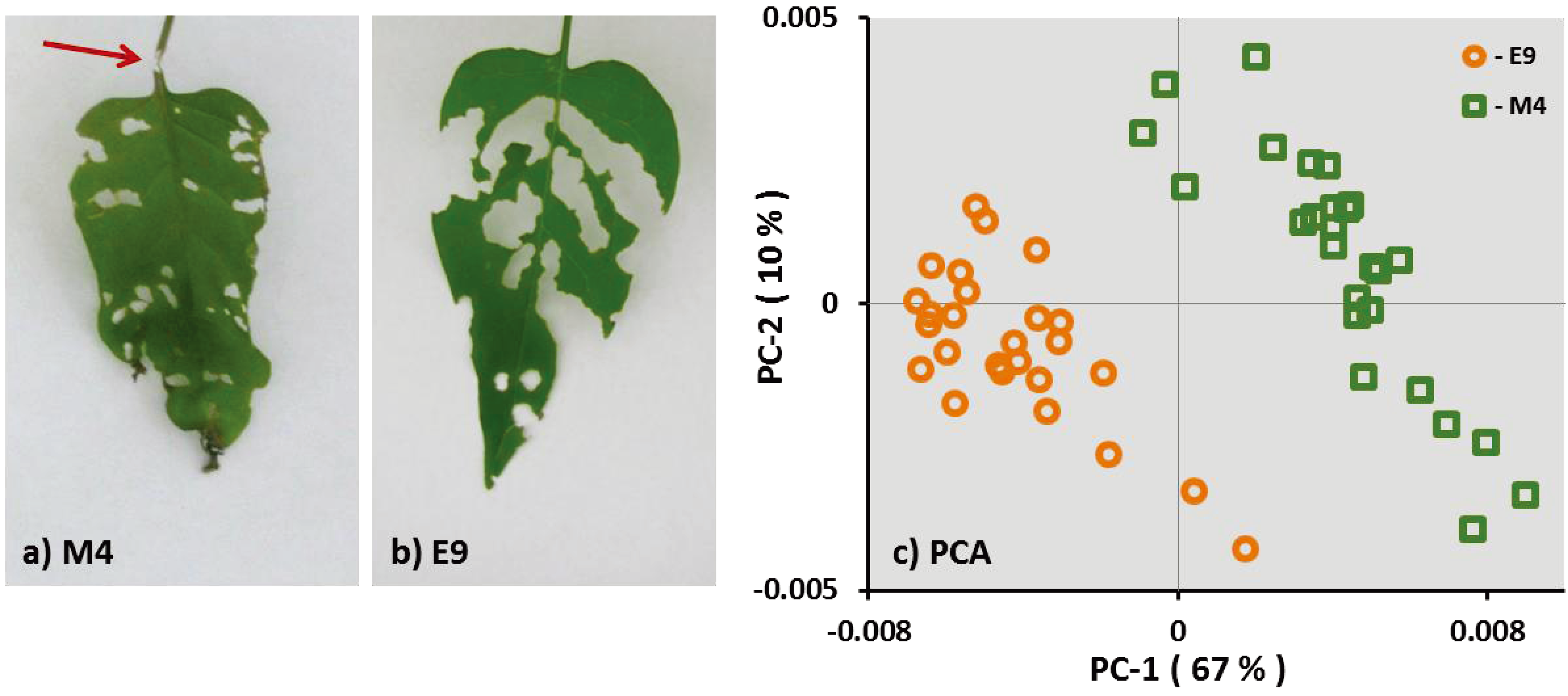{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | BGH | AGH+BGH |
|---|---|---|
| M4 | 2.00 ± 1.13 | 0.71 ± 0.93 |
| E9 | 1.35 ± 1.00 | 0.58 ± 0.57 |
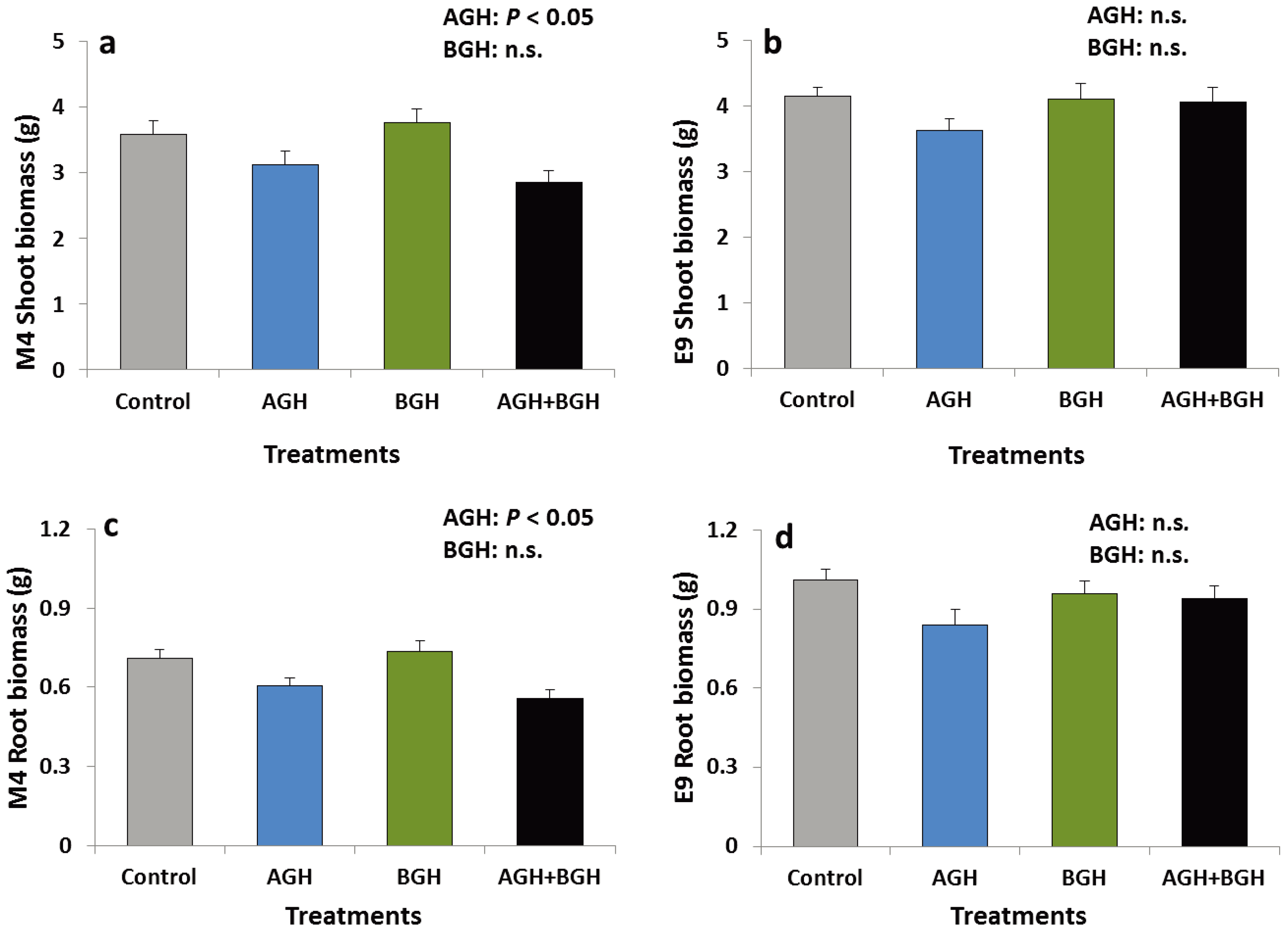
3.1.2. Plant Biomass

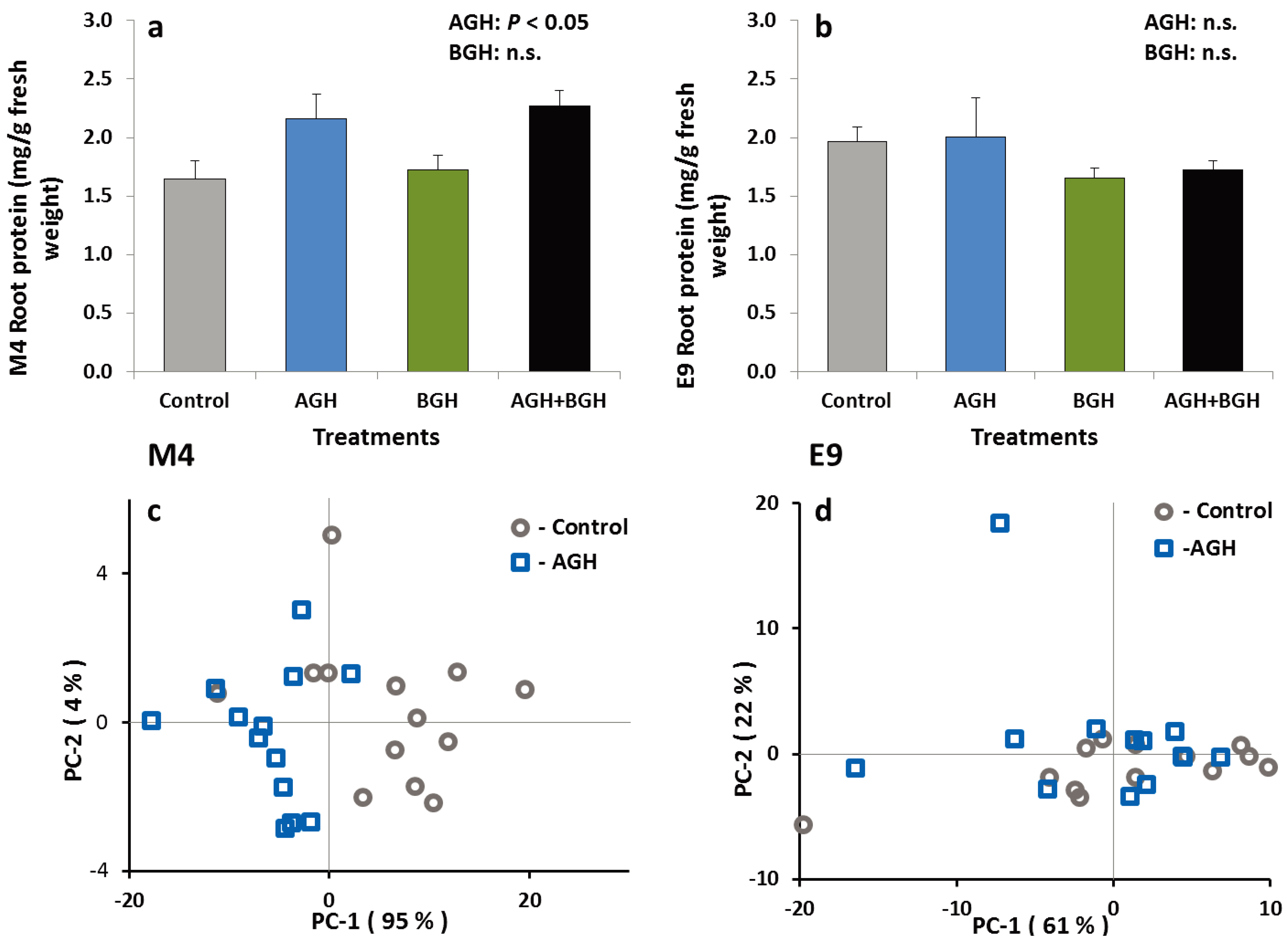
3.1.3. PI and Protein Content in the Root Tissue

3.1.4. Carbon and Nitrogen Concentration
3.1.5. Feeding Bioassay

| Bioassay | Genotypes | Root eaten (mg) | Aggregate | |||
|---|---|---|---|---|---|---|
| Control | AGH | BGH | AGH+BGH | |||
| Non-Choice | M4 | 3.6 ± 0.20 | 3.62 ± 0.08 | 3.73 ± 0.11 | 3.53 ± 0.18 | 3.62 ± 0.07 |
| E9 | 3.57 ± 0.15 | 3.55 ± 0.17 | 3.27 ± 0.09 | 3.33 ± 0.14 | 3.43 ± 0.07 | |
| Choice | M4 | 2.03 ± 0.16 | 2.22 ± 0.16 | 1.95 ± 0.12 | 2.03 ± 0.09 | 2.06 ± 0.07 |
| E9 | 2.20 ± 0.10 | 2.21 ± 0.19 | 1.78 ± 0.19 | 1.85 ± 0.08 | 2.01 ± 0.08 | |
| Bioassay | Genotypes | Weight gain of the larvae (mg) | ||||
| Control | AGH | BGH | AGH+BGH | |||
| Non-Choice | M4 | 0.43 ± 0.14 | 0.46 ± 0.16 | 1.17 ± 0.35 | 0.79 ± 0.30 | 0.71 ± 0.13 |
| E9 | 0.31 ± 0.24 | 0.85 ± 0.19 | 0.45 ± 0.21 | 0.33 ± 0.24 | 0.48 ± 0.11 | |
| Choice | M4 | E9 | ||||
| 1.59 ± 0.41 | 0.51 ± 0.32 | |||||
3.2. Discussion
3.2.1. AG Herbivory Effects on Plant Traits
3.2.2. Effects of Changes in Plant Traits by AG Herbivory on BG Herbivores
3.2.3. Differences among the Plant Genotypes
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Masters, G.J.; Brown, V.K.; Gange, A.C. Plant mediated interactions between aboveground and belowground insect herbivores. Oikos 1993, 66, 148–151. [Google Scholar] [CrossRef]
- Blossey, B.; Hunt-Joshi, T.R. Belowground herbivory by insects: Influence on plants and aboveground herbivores. Ann. Rev. Entomol. 2003, 48, 521–547. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, I.; Halitschke, R.; Kessler, A.; Rehill, B.J.; Sardanelli, S.; Denno, R.F. Physiological integration of roots and shoots in plant defense strategies links above- and belowground herbivory. Ecol. Lett. 2008, 11, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, I.; Halitschke, R.; Kessler, A.; Sardanelli, S.; Denno, R.F. Constitutive and induced defenses to herbivory in above- and belowground plant tissues. Ecology 2008, 89, 392–406. [Google Scholar] [CrossRef] [PubMed]
- Wurst, S.; van der Putten, W.H. Root herbivore identity matters in plant-mediated interactions between root and shoot herbivores. Basic Appl. Ecol. 2007, 8, 491–499. [Google Scholar] [CrossRef]
- Erb, M.; Ton, J.; Degenhardt, J.; Turlings, T.C.J. Interactions between arthropod-induced aboveground and belowground defenses in plants. Plant Physiol. 2008, 146, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Robert, C.A.M.; Hibbard, B.E.; Turlings, T.C.J. Sequence of arrival determines plant-mediated interactions between herbivores. J. Ecol. 2011, 99, 7–15. [Google Scholar] [CrossRef]
- Johnson, S.N.; Clark, K.E.; Hartley, S.E.; Jones, T.H.; McKenzie, S.W.; Koricheva, J. Aboveground-belowground herbivore interactions: A meta-analysis. Ecology 2012, 93, 2208–2215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutyniok, M.; Müller, C. Plant-mediated interactions between shoot-feeding aphids and root-feeding nematodes depend on nitrate fertilization. Oecologia 2013, 173, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Gatehouse, J.A. Plant resistance towards insect herbivores: A dynamic interaction. New Phytol. 2002, 156, 145–169. [Google Scholar] [CrossRef]
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [PubMed]
- Bezemer, T.M.; van Dam, N.M. Linking aboveground and belowground interactions via induced plant defenses. Trends Ecol. Evol. 2005, 20, 617–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezemer, T.M.; Wagenaar, R.; van Dam, N.M.; Wackers, F.L. Interactions between above- and belowground insect herbivores as mediated by the plant defense system. Oikos 2003, 101, 555–562. [Google Scholar] [CrossRef]
- Vandermeijden, E.; Wijn, M.; Verkaar, H.J. Defense and regrowth, alternative plant strategies in the struggle against herbivores. Oikos 1988, 51, 355–363. [Google Scholar] [CrossRef]
- Strauss, S.Y.; Agrawal, A.A. The ecology and evolution of plant tolerance to herbivory. Trends Ecol. Evol. 1999, 14, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, N.M. Belowground herbivory and plant defenses. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 373–391. [Google Scholar]
- Babst, B.A.; Ferrieri, R.A.; Gray, D.W.; Lerdau, M.; Schlyer, D.J.; Schueller, M.; Thorpe, M.R.; Orians, C.M. Jasmonic acid induces rapid changes in carbon transport and partitioning in populus. New Phytol. 2005, 167, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.A.R.; Ferrieri, A.P.; Robert, C.A.M.; Glauser, G.; Kallenbach, M.; Baldwin, I.T.; Erb, M. Leaf-herbivore attack reduces carbon reserves and regrowth from the roots via jasmonate and auxin signaling. New Phytol. 2013, 200, 1234–1246. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.N.; Hawes, C.; Karley, A.J. Reappraising the role of plant nutrients as mediators of interactions between root- and foliar-feeding insects. Funct. Ecol. 2009, 23, 699–706. [Google Scholar] [CrossRef]
- Ohgushi, T. Indirect interaction webs: Herbivore-induced effects through trait change in plants. Ann. Rev. Ecol. Evol. Syst. 2005, 36, 81–105. [Google Scholar] [CrossRef]
- Wurst, S.; Wagenaar, R.; Biere, A.; van der Putten, W.H. Microorganisms and nematodes increase levels of secondary metabolites in roots and root exudates of Plantago lanceolata. Plant Soil 2010, 329, 117–126. [Google Scholar] [CrossRef]
- Rasmann, S.; Agrawal, A.A. In defense of roots: A research agenda for studying plant resistance to belowground herbivory. Plant Physiol. 2008, 146, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Glauser, G.; Robert, C.A.M. Induced immunity against belowground insect herbivores-activation of defenses in the absence of a jasmonate burst. J. Chem. Ecol. 2012, 38, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, I.T. Mechanism of damage-induced alkaloid production in wild tobacco. J. Chem. Ecol. 1989, 15, 1661–1680. [Google Scholar] [CrossRef] [PubMed]
- Karban, R. Community organization of erigeron-glaucus folivores—Effects of competition, predation, and host plant. Ecology 1989, 70, 1028–1039. [Google Scholar] [CrossRef]
- Johnson, M.T.J.; Agrawal, A.A. Plant genotype and environment interact to shape a diverse arthropod community on evening primrose (Oenothera biennis). Ecology 2005, 86, 874–885. [Google Scholar] [CrossRef]
- Smith, R.A.; Mooney, K.A.; Agrawal, A.A. Coexistence of three specialist aphids on common milkweed, asclepias syriaca. Ecology 2008, 89, 2187–2196. [Google Scholar] [CrossRef] [PubMed]
- English-Loeb, G.; Karban, R.; Walker, M.A. Genotypic variation in constitutive and induced resistance in grapes against spider mite (Acari: Tetranychidae) herbivores. Environ. Entomol. 1998, 27, 297–304. [Google Scholar]
- Bingham, R.A.; Agrawal, A.A. Specificity and trade-offs in the induced plant defence of common milkweed Asclepias syriaca to two lepidopteran herbivores. J. Ecol. 2010, 98, 1014–1022. [Google Scholar] [CrossRef]
- Uesugi, A.; Poelman, E.H.; Kessler, A. A test of genotypic variation in specificity of herbivore-induced responses in Solidago altissima L. (Asteraceae). Oecologia 2013, 173, 1387–1396. [Google Scholar]
- Hol, W.H.G.; Macel, M.; van Veen, J.A.; van der Meijden, E. Root damage and aboveground herbivory change concentration and composition of pyrrolizidine alkaloids of Senecio jacobaea. Basic Appl. Ecol. 2004, 5, 253–260. [Google Scholar] [CrossRef]
- Wurst, S.; van Dam, N.M.; Monroy, F.; Biere, A.; van der Putten, W.H. Intraspecific variation in plant defense alters effects of root herbivores on leaf chemistry and aboveground herbivore damage. J. Chem. Ecol. 2008, 34, 1360–1367. [Google Scholar] [CrossRef] [PubMed]
- Pegtel, D.M. Germination in populations of Solanum dulcamara L. from contrasting habitats. New Phytol. 1985, 100, 671–679. [Google Scholar] [CrossRef]
- Viswanathan, D.V.; Narwani, A.J.T.; Thaler, J.S. Specificity in induced plant responses shapes patterns of herbivore occurrence on Solanum dulcamara. Ecology 2005, 86, 886–896. [Google Scholar] [CrossRef]
- Viswanathan, D.V.; Lifchits, O.A.; Thaler, J.S. Consequences of sequential attack for resistance to herbivores when plants have specific induced responses. Oikos 2007, 116, 1389–1399. [Google Scholar] [CrossRef]
- Sonnemann, I.; Grunz, S.; Wurst, S. Horizontal migration of click beetle (Agriotes spp.) larvae depends on food availability. Entomol. Exp. Appl. 2014, 150, 174–178. [Google Scholar]
- Bode, R.F.; Halitschke, R.; Kessler, A. Herbivore damage-induced production and specific anti-digestive function of serine and cysteine protease inhibitors in tall goldenrod, Solidago altissima L. (Asteraceae). Planta 2013, 237, 1287–1296. [Google Scholar]
- Dixon, R.A.; Strack, D. Phytochemistry meets genome analysis, and beyond. Phytochemistry 2003, 62, 815–816. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Ellis, D.I. Metabolomics: Current analytical platforms and methodologies. Trac-Trends Anal. Chem. 2005, 24, 285–294. [Google Scholar]
- Ellis, D.I.; Goodacre, R. Metabolic fingerprinting in disease diagnosis: Biomedical applications of infrared and raman spectroscopy. Analyst 2006, 131, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Socaciu, C.; Fetea, F.; Ranga, F. IR and Raman spectroscopy-advanced and versatile techniques for agrifood quality and authenticity assessment. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca Agric. 2009, 66, 459–464. [Google Scholar]
- Schulz, H.; Schrader, B.; Quilitzsch, R.; Pfeffer, S.; Krüger, H. Rapid classification of basil chemotypes by various vibrational spectroscopy methods. J. Agric. Food Chem. 2003, 51, 2475–2481. [Google Scholar] [CrossRef] [PubMed]
- Gidman, E.A.; Stevens, C.J.; Goodacre, R.; Broadhurst, D.; Emmett, B.; Gwynn-Jones, D. Using metabolic fingerprinting of plants for evaluating nitrogen deposition impacts on the landscape level. Global Change Biol. 2006, 12, 1460–1465. [Google Scholar] [CrossRef]
- Krähmer, A.; Gudi, G.; Weiher, N.; Gierus, M.; Schütze, W.; Schulz, H. Characterization and quantification of secondary metabolite profiles in leaves of red and white clover species by nir and atr-ir spectroscopy. Vib. Spectrosc. 2013, 68, 96–103. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. (Eds.) Mixed Effects Models and Extensions in Ecology with R; Springer: Berlin, Heidelberg, Germany, 2009; ISBN: 978-0-387-87457-9 (Print) 978-0-387-87458-6 (Online).
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Siemann, E.; Yang, X.F.; Wheeler, G.S.; Ding, J.Q. Facilitation and inhibition: Changes in plant nitrogen and secondary metabolites mediate interactions between above-ground and below-ground herbivores. Proc. R. Soc. B Biol. Sci. 2013, 280, 7. [Google Scholar]
- Bronner, R.; Westphal, E.; Dreger, F. Enhanced peroxidase-activity associated with the hypersensitive response of Solanum dulcamara to the gall mite Aceria cladophthirus (Acari, Eriophyoidea). Can. J. Bot.-Rev. Can. Botanique 1991, 69, 2192–2196. [Google Scholar]
- Viswanathan, D.V.; Thaler, J.S. Plant vascular architecture and within-plant spatial patterns in resource quality following herbivory. J. Chem. Ecol. 2004, 30, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Zavala, J.A.; Patankar, A.G.; Gase, K.; Baldwin, I.T. Constitutive and inducible trypsin proteinase inhibitor production incurs large fitness costs in Nicotiana attenuata. Proc. Natl. Acad. Sci. USA 2004, 101, 1607–1612. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.W.; Yi, H.S.; Kim, H.; Lee, B.; Lee, S.; Ghim, S.Y.; Ryu, C.M. Whitefly infestation of pepper plants elicits defence responses against bacterial pathogens in leaves and roots and changes the below-ground microflora. J. Ecol. 2011, 99, 46–56. [Google Scholar] [CrossRef]
- Furlan, L. The biology of Agriotes ustulatus Schäller (Col., Elateridae). II. Larval development, pupation, whole cycle description and practical implications. J. Appl. Entomol. Z. Angew. Entomol. 1998, 122, 71–78. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kafle, D.; Krähmer, A.; Naumann, A.; Wurst, S. Genetic Variation of the Host Plant Species Matters for Interactions with Above- and Belowground Herbivores. Insects 2014, 5, 651-667. https://doi.org/10.3390/insects5030651
Kafle D, Krähmer A, Naumann A, Wurst S. Genetic Variation of the Host Plant Species Matters for Interactions with Above- and Belowground Herbivores. Insects. 2014; 5(3):651-667. https://doi.org/10.3390/insects5030651
Chicago/Turabian StyleKafle, Dinesh, Andrea Krähmer, Annette Naumann, and Susanne Wurst. 2014. "Genetic Variation of the Host Plant Species Matters for Interactions with Above- and Belowground Herbivores" Insects 5, no. 3: 651-667. https://doi.org/10.3390/insects5030651