Temperature Sensitivity of Wild-Type, Mutant and Genetic Sexing Strains of Ceratitis capitata

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
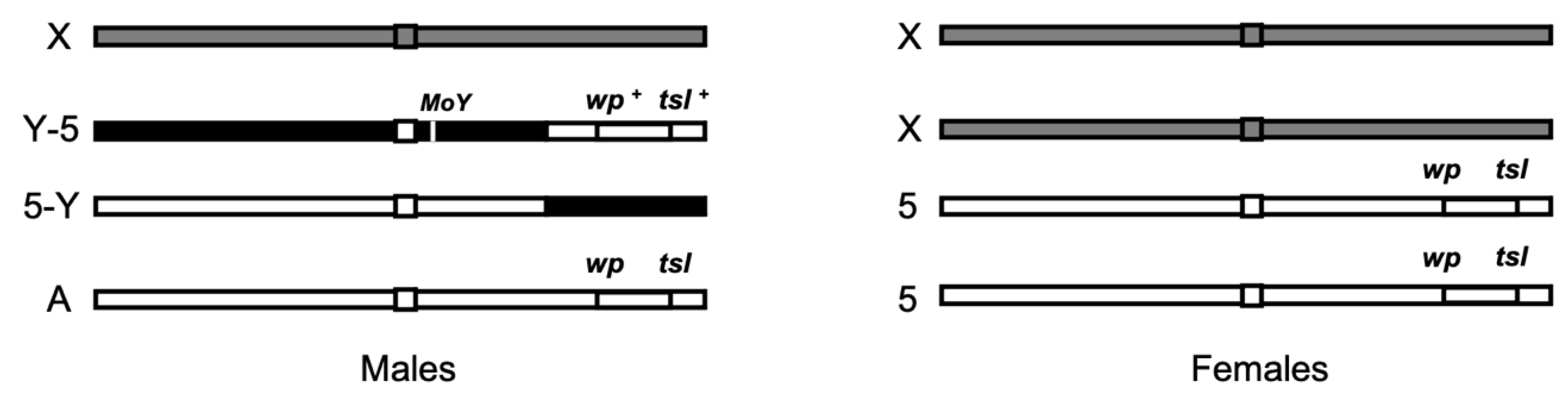
2.1. Ceratitis Capitata Strains and Rearing Conditions
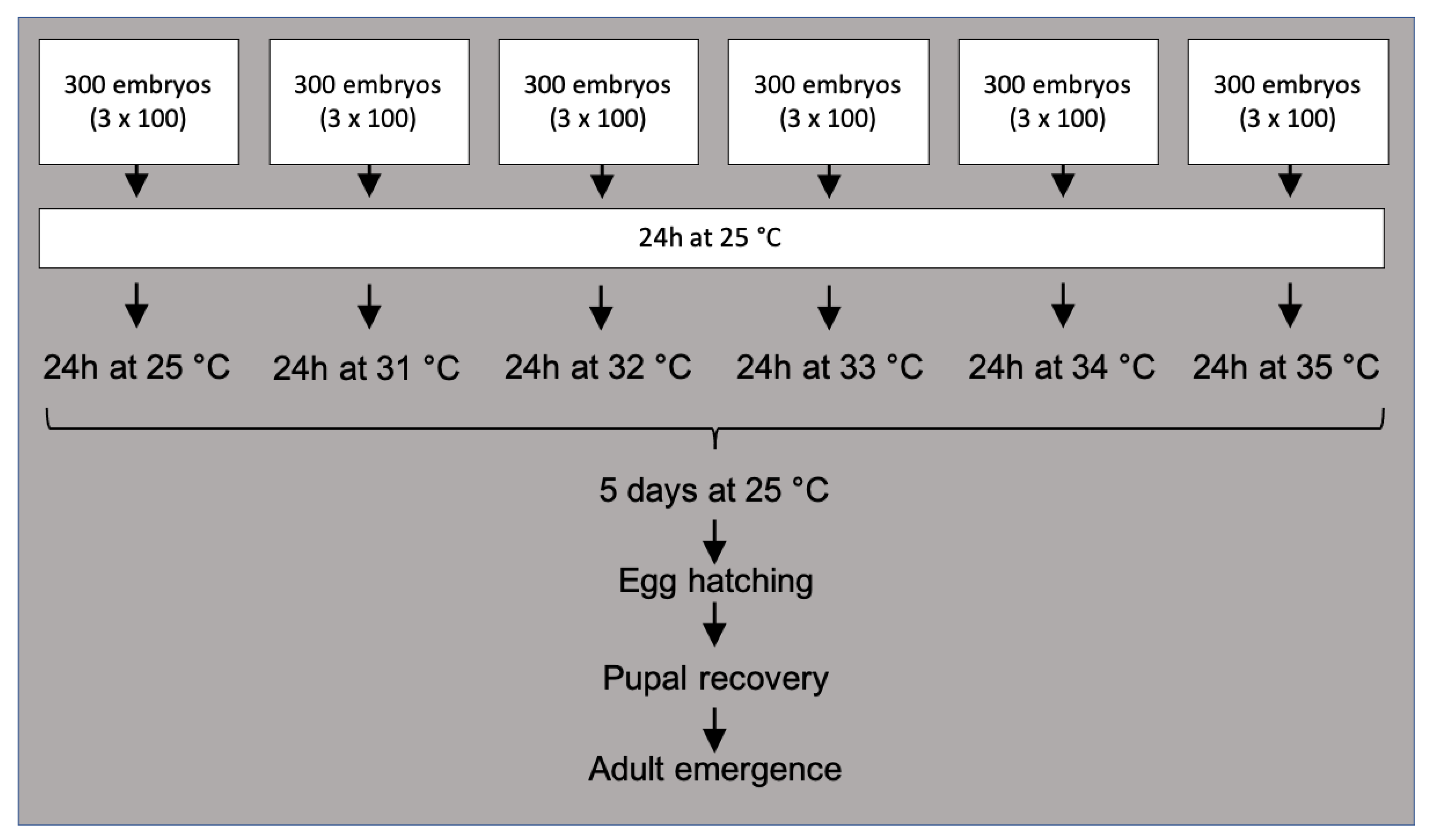
2.2. Temperature-Sensitive Lethal Test
2.3. Statistical Analysis
3. Results
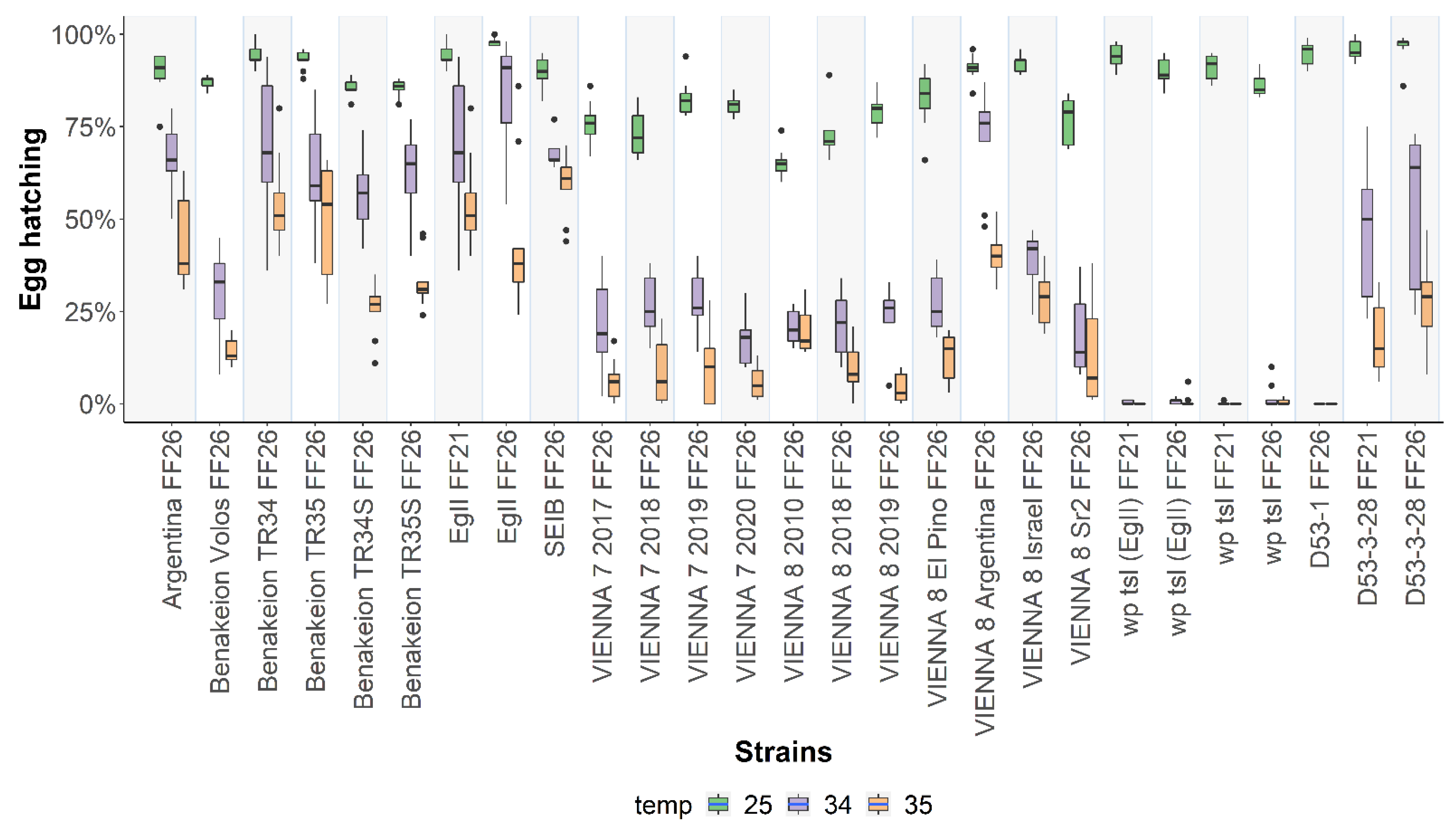
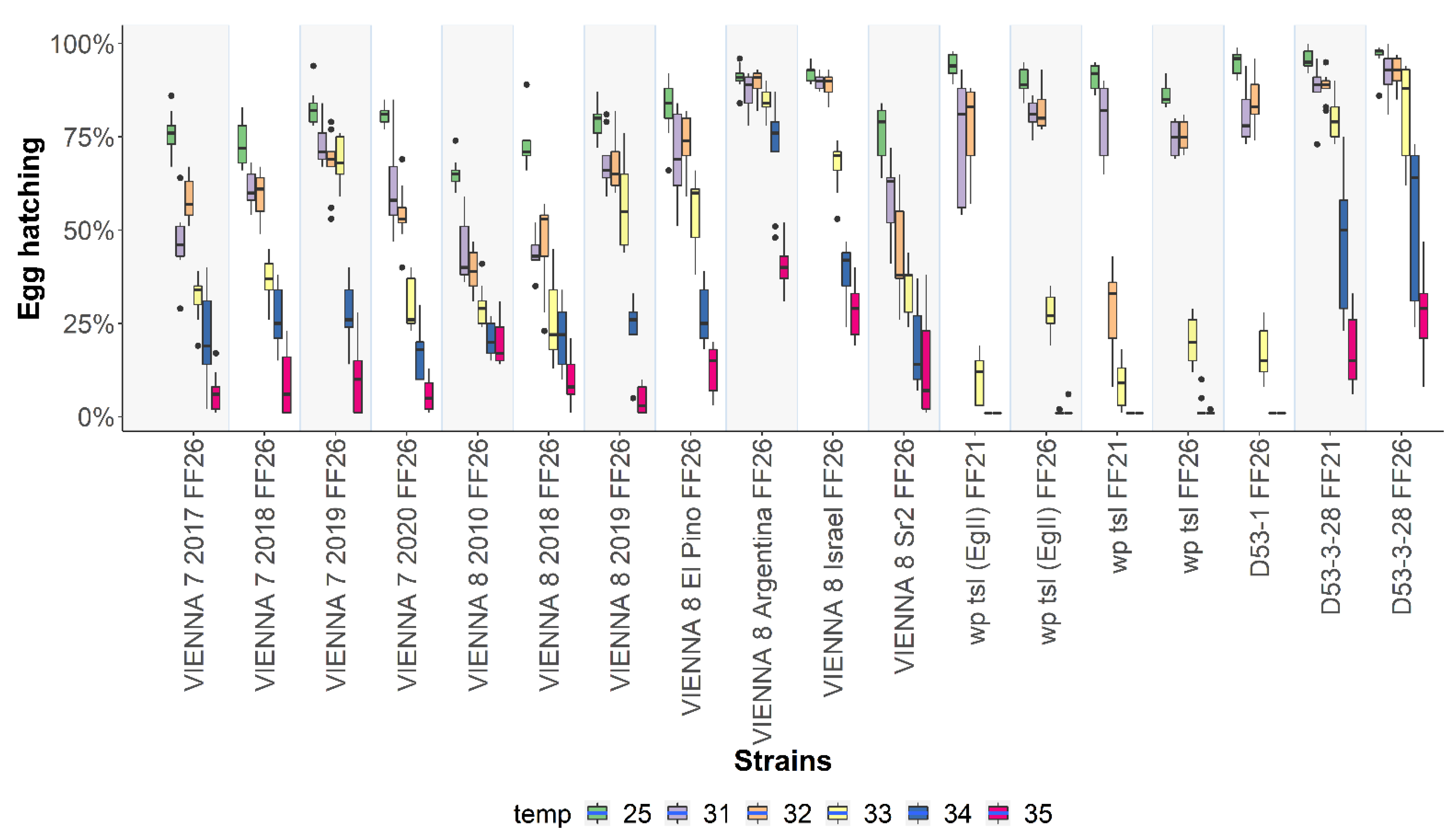
3.1. Egg Hatching at 25 °C, 34 °C, and 35 °C
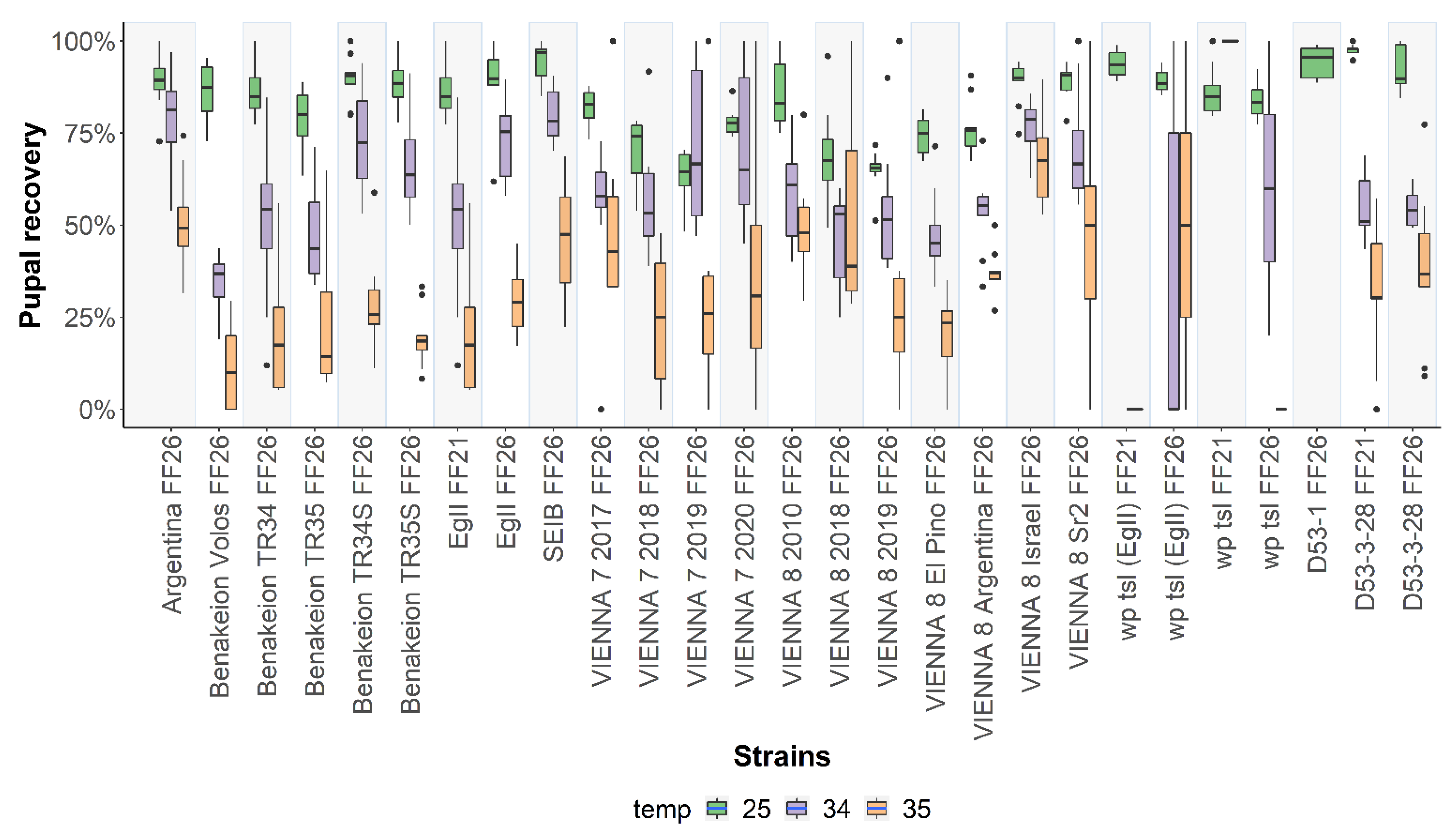
3.2. Pupal Recovery at 25 °C, 34 °C, and 35 °C
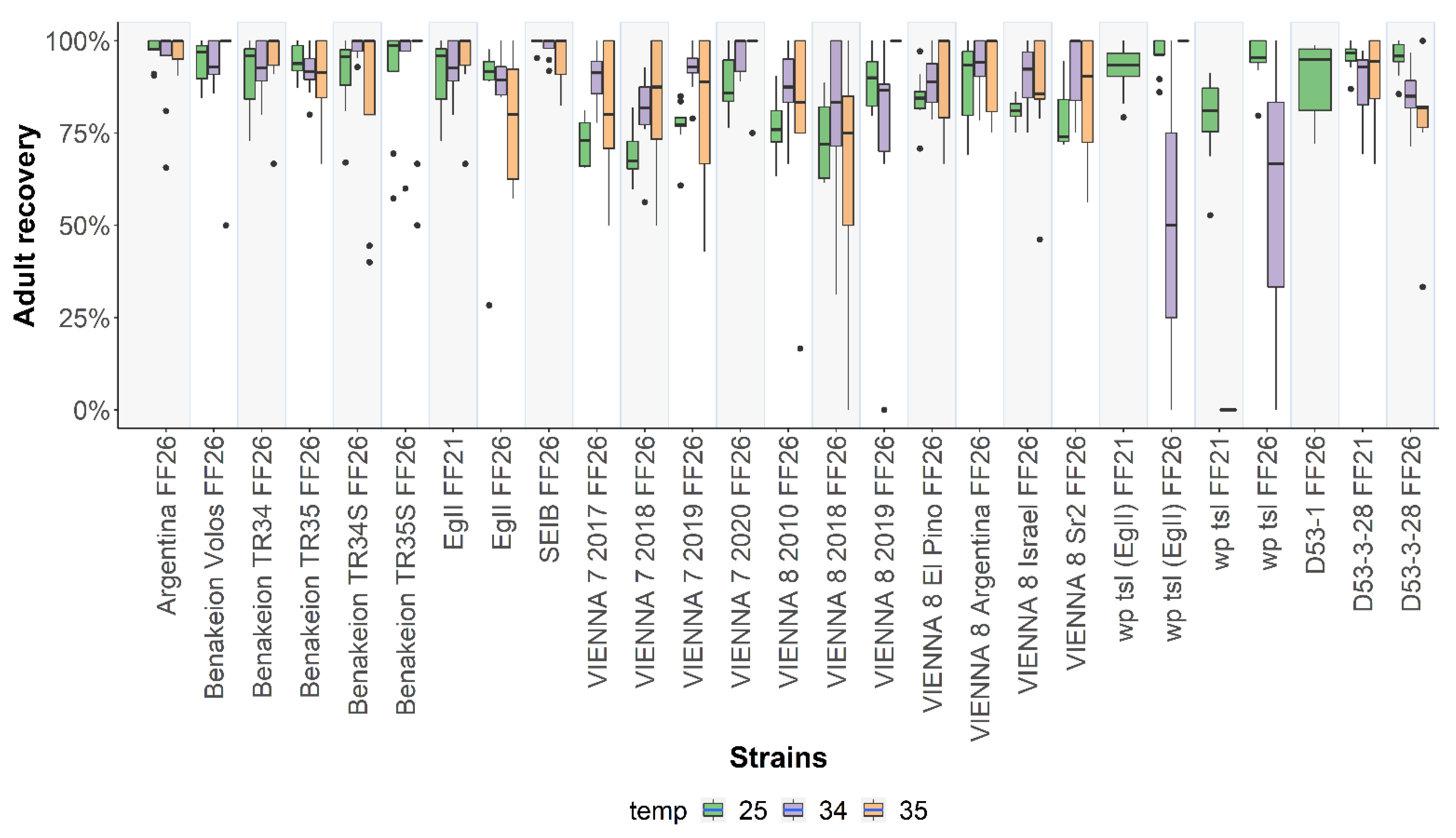
3.3. Adult Recovery at 25 °C, 34 °C, and 35 °C
3.4. Temperature-Sensitive Lethal Tests at Additional Temperatures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siebert, J.; Cooper, T. If Medfly Infestation Triggered a Trade Ban: Embargo on California Produce Would Cause Revenue, Job Loss. Calif. Agric. 1995, 49, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Deschepper, P.; Todd, T.N.; Virgilio, M.; De Meyer, M.; Barr, N.B.; Ruiz-Arce, R. Looking at the Big Picture: Worldwide Population Structure and Range Expansion of the Cosmopolitan Pest Ceratitis Capitata (Diptera, Tephritidae). Biol. Invasions 2021, 23, 3529–3543. [Google Scholar] [CrossRef]
- Papadopoulos, N.T. Mediterranean Fruit Fly, Ceratitis Capitata (Wiedemann) (Diptera: Tephritidae). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 2318–2322. ISBN 978-1-4020-6359-6. [Google Scholar]
- De Meyer, M.; Copeland, R.S.; Wharton, R.A.; McPheron, B.A.; Barnes, B.N. On the Geographic Origin of the Medfly Ceratitis Capitata (Wiedemann) (Diptera: Tephritidae). In Proceedings of the 6th International Fruit Fly Symposium, Stellenbosch, South Africa, 6–10 May 2002; pp. 45–53. [Google Scholar]
- Knipling, E.F. Possibilities of Insect Control or Eradication through the Use of Sexually Sterile Males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Taylor & Francis: Vienna, Austria, 2021. [Google Scholar]
- Klassen, W.; Vreysen, M.J.B. Area-Wide Integrated Pest Management and the Sterile Insect Technique. In Sterile Insect Technique; CRC Press: Boca Raton, FL, USA, 2021; pp. 75–112. ISBN 1-00-303557-4. [Google Scholar]
- Enkerlin, W. Economics of Area-Wide Sit Control Programs; Research Institute for Subtropics: Naha, Japan, 2003; pp. 1–10. [Google Scholar]
- Millo, E.P.; Herrero, R.A.; Alfaro-Lassala, F. Plan de Actuación Contra La Mosca de Las Frutas (Ceratitis Capitata) En La Comunidad Valenciana. Phytoma Esp. Rev. Prof. Sanid. Veg. 2003, 153, 127–130. [Google Scholar]
- Cayol, J.P.; Rössler, Y.; Weiss, M.; Bahdousheh, M.; Omari, M.; Hamalawi, M.; Almughayyar, A.; Barnes, B. Fruit Fly Control and Monitoring in the Near East: Shared Concern in a Regional Transboundary Problem. In Proceedings of the Symposium: 6th International Symposium on Fruit Flies of Economic Importance, Stellenbosch, South Africa, 6–10 May 2002; pp. 6–10. [Google Scholar]
- Porras-Reyes, L.A. Control Microbiano de Ceratitis Capitata (Wiedmann) (Mosca Del Mediterráneo) Con El Hongo Entomopatógeno Beauveria Bassiana (Balsamo) Vuillemin. In Proyecto de Graduación (Bachillerato en Ingeniería en Biotecnología) Instituto Tecnológico de Costa Rica, Escuela de Biología; RepositorioTEC: Cartago, Costa Rica, 2007; pp. 15–46. Available online: https://hdl.handle.net/2238/455 (accessed on 11 October 2022).
- Enkerlin, W.R. Impact of Fruit Fly Control Programmes Using the Sterile Insect Technique. In Sterile Insect Technique; CRC Press: Boca Raton, FL, USA, 2021; pp. 979–1006. ISBN 1-00-303557-4. [Google Scholar]
- Zavala-López, J.L.; Enkerlin, W.R. Guideline for Packing, Shipping, Holding and Release of Sterile Flies in Area-Wide Fruit Fly Control Programmes; FAO/IAEA: Vienna, Austria, 2017; pp. 1–140. [Google Scholar]
- Hendrichs, J.; Franz, G.; Rendon, P. Increased Effectiveness and Applicability of the Sterile Insect Technique through Male-Only Releases for Control of Mediterranean Fruit Flies during Fruiting Seasons. J. Appl. Entomol. 1995, 119, 371–377. [Google Scholar] [CrossRef]
- Caceres, C. Mass Rearing of Temperature Sensitive Genetic Sexing Strains in the Mediterranean Fruit Fly (Ceratitis Capitata). Genetica 2002, 116, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Rendón, P.; McInnis, D.; Lance, D.; Stewart, J. Medfly (Diptera:Tephritidae) Genetic Sexing: Large-Scale Field Comparison of Males-Only and Bisexual Sterile Fly Releases in Guatemala. J. Econ. Entomol. 2004, 97, 1547–1553. [Google Scholar] [CrossRef]
- Franz, G.; Bourtzis, K.; Caceres, C. Practical and Operational Genetic Sexing Systems Based on Classical Genetic Approaches in Fruit Flies, an Example for Other Species Amenable to Large-Scale Rearing for the Sterile Insect Technique. In Sterile Insect Technique; CRC Press: Boca Raton, FL, USA, 2021; pp. 575–604. ISBN 1-00-303557-4. [Google Scholar]
- Augustinos, A.A.; Targovska, A.; Cancio-Martinez, E.; Schorn, E.; Franz, G.; Cáceres, C.; Zacharopoulou, A.; Bourtzis, K.C. Eratitis Capitata Genetic Sexing Strains: Laboratory Evaluation of Strains from Mass-rearing Facilities Worldwide. Entomol. Exp. Appl. 2017, 164, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Zacharopoulou, A.; Augustinos, A.A.; Drosopoulou, E.; Tsoumani, K.T.; Gariou-Papalexiou, A.; Franz, G.; Mathiopoulos, K.D.; Bourtzis, K.; Mavragani-Tsipidou, P. A Review of More than 30 Years of Cytogenetic Studies of T Ephritidae in Support of Sterile Insect Technique and Global Trade. Entomol. Exp. Appl. 2017, 164, 204–225. [Google Scholar] [CrossRef] [Green Version]
- Fisher, K.; Caceres, C. A Filter Rearing System for Mass Reared Genetic Sexing Strains of Mediterranean Fruit Fly (Diptera: Tephritidae). In Proceedings of the Area-Wide Control of Fruit Flies and Other Insect Pests. Joint Proceedings of the International Conference on Area-Wide Control of Insect Pests, 28 May–2 June 1998 and the Fifth International Symposium on Fruit Flies of Economic Importance, Penang, Malaysia, 1–5 June 1998; Penerbit Universiti Sains Malaysia: Gelugor, Malaysia, 2000; pp. 543–550. [Google Scholar]
- Tanaka, N.; Steiner, L.F.; Ohinata, K.; Okamoto, R. Low-Cost Larval Rearing Medium for Mass Production of Oriental and Mediterranean Fruit Flies. J. Econ. Entomol. 1969, 62, 967–968. [Google Scholar] [CrossRef]
- Niyazi, N.; Caceres, C.; Delprat, A.; Wornoayporn, V.; Santos, E.R.; Franz, G.; Robinson, A.S. Genetics and Mating Competitiveness of Ceratitis Capitata (Diptera: Tephritidae) Strains Carrying the Marker Sergeant, Sr 2. Ann. Entomol. Soc. Am. 2005, 98, 119–125. [Google Scholar] [CrossRef]
- Dunn, P.K.; Smyth, G.K. Generalized Linear Models with Examples in R.; Springer: Berlin/Heidelberg, Germany, 2018; ISBN 1-4419-0118-3. [Google Scholar]
- Hartig, F.; Hartig, M.F. Package ‘DHARMa’; R Package: Vienna, Austria, 2017; pp. 1–53. [Google Scholar]
- Demétrio, C.G.B.; Hinde, J.; Moral, R.A. Models for Overdispersed Data in Entomology. In Ecological Modelling Applied to Entomology; Ferreira, C.P., Godoy, W.A.C., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 219–259. ISBN 978-3-319-06876-3. [Google Scholar]
- Nelder, J.A.; Wedderburn, R.W.M. Generalized Linear Models. J. R. Stat. Soc. Ser. Gen. 1972, 135, 370. [Google Scholar] [CrossRef]
- Moral, R.A.; Hinde, J.; Demétrio, C.G.B. Half-Normal Plots and Overdispersed Models in R: The Hnp Package. J. Stat. Softw. 2017, 81, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Searle, S.R.; Speed, F.M.; Milliken, G.A. Population Marginal Means in the Linear Model: An Alternative to Least Squares Means. Am. Stat. 1980, 34, 216–221. [Google Scholar] [CrossRef]
- Gilchrist, A.S.; Cameron, E.C.; Sved, J.A.; Meats, A.W. Genetic Consequences of Domestication and Mass Rearing of Pest Fruit Fly Bactrocera Tryoni (Diptera: Tephritidae). J. Econ. Entomol. 2012, 105, 1051–1056. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Hallas, R.; Sinclair, C.; Partridge, L. Rapid Loss of Stress Resistance in Drosophila Melanogaster under Adaptation to Laboratory Culture. Evolution 2001, 55, 436–438. [Google Scholar] [CrossRef]
- Raphael, K.A.; Shearman, D.C.; Gilchrist, A.S.; Sved, J.A.; Morrow, J.L.; Sherwin, W.B.; Riegler, M.; Frommer, M. Australian Endemic Pest Tephritids: Genetic, Molecular and Microbial Tools for Improved Sterile Insect Technique. BMC Genet. 2014, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zygouridis, N.E.; Argov, Y.; Nemny-Lavy, E.; Augustinos, A.A.; Nestel, D.; Mathiopoulos, K.D. Genetic Changes during Laboratory Domestication of an Olive Fly SIT Strain. J. Appl. Entomol. 2014, 138, 423–432. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Ross, P.A. Rates and Patterns of Laboratory Adaptation in (Mostly) Insects. J. Econ. Entomol. 2018, 111, 501–509. [Google Scholar] [CrossRef]
- Gaire, S.K.; Pokhrel, S.; Biswas, M.J.H.; Mainali, B.P.; Mendez, V.; Pérez, J.; Taylor, P.W.; Rempoulakis, P. Effects of Domestication on Quality Control Parameters of the Queensland Fruit Fly Bactrocera Tryoni (Diptera: Tephritidae). J. Appl. Entomol. 2022, 146, 687–699. [Google Scholar] [CrossRef]
- Reynolds, O.; Jessup, A.; Dominiak, B.; Smallridge, C.; Cockington, V.; Penrose, L.; Taylor, P.; Collins, S. Enhancing Emergence and Release Methods of the Sterile Insect Technique (SIT) to Improve Market Access. Horticulture Australia Ltd.: Sydney, Australia, 2012; pp. 116–129. [Google Scholar]
- Ward, C.M.; Aumann, R.A.; Whitehead, M.A.; Nikolouli, K.; Leveque, G.; Gouvi, G.; Fung, E.; Reiling, S.J.; Djambazian, H.; Hughes, M.A. White Pupae Phenotype of Tephritids Is Caused by Parallel Mutations of a MFS Transporter. Nat. Commun. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Porras, M.F.; Meza, J.S.; Rajotte, E.G.; Bourtzis, K.; Cáceres, C. Improving the Phenotypic Properties of the Ceratitis Capitata (Diptera: Tephritidae) Temperature-Sensitive Lethal Genetic Sexing Strain in Support of Sterile Insect Technique Applications. J. Econ. Entomol. 2020, 113, 2688–2694. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Staples, D.; Díaz-Fleischer, F.; Montoya, P. The Sterile Insect Technique: Success and Perspectives in the Neotropics. Neotrop. Entomol. 2021, 50, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Malacrida, A.R.; Gomulski, L.M.; Bonizzoni, M.; Bertin, S.; Gasperi, G.; Guglielmino, C. Al Globalization and Fruitfly Invasion and Expansion: The Medfly Paradigm. Genetica 2007, 131, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lemic, D.; Bjeliš, M.; Ninčević, P.; Živković, I.P.; Popović, L.; Gašparić, H.V.; Benitez, H.A. Medfly Phenotypic Plasticity as A Prerequisite for Invasiveness and Adaptation. Sustainability 2021, 13, 12510. [Google Scholar] [CrossRef]
- Weldon, C.W.; Nyamukondiwa, C.; Karsten, M.; Chown, S.L.; Terblanche, J.S. Geographic Variation and Plasticity in Climate Stress Resistance among Southern African Populations of Ceratitis Capitata (Wiedemann)(Diptera: Tephritidae). Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Weldon, C.W.; Díaz-Fleischer, F.; Pérez-Staples, D. Desiccation Resistance of Tephritid Flies: Recent Research Results and Future Directions. In Area-Wide Manag Fruit Fly Pests; CRC Press: Boca Raton, FL, USA, 2019; pp. 27–43. [Google Scholar]
- Pörtner, H.-O.; Roberts, D.C.; Adams, H.; Adler, C.; Aldunce, P.; Ali, E.; Begum, R.A.; Betts, R.; Kerr, R.B.; Biesbroek, R. Climate Change 2022: Impacts, Adaptation and Vulnerability. In IPCC Sixth Assessment Report; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Group | |
|---|---|---|
| 1 | EgII FF21 a | wild-type |
| 2 | EgII FF26 a | wild-type |
| 3 | Benakeion Volos FF26 | wild-type |
| 4 | Seibersdorf (SEIB) FF26 | wild-type |
| 5 | Argentina FF26 | wild-type |
| 6 | Benakeion TR 34 FF26 b | wild-type |
| 7 | Benakeion TR 35 FF26 b | wild-type |
| 8 | Benakeion TR 34S FF26 c | wild-type |
| 9 | Benakeion TR 35S FF26 c | wild-type |
| 10 | VIENNA 8 2010 FF26 d | GSS |
| 11 | VIENNA 8 2018 FF26 d | GSS |
| 12 | VIENNA 8 2019 FF26 d | GSS |
| 13 | VIENNA 8 Sr2 FF26 e | GSS |
| 14 | VIENNA 8 “El Pino” FF26 | GSS |
| 15 | VIENNA 8 Israel FF26 | GSS |
| 16 | VIENNA 8 Argentina FF26 | GSS |
| 17 | VIENNA 7 2017 FF26 d | GSS |
| 18 | VIENNA 7 2018 FF26 d | GSS |
| 19 | VIENNA 7 2019 FF26 d | GSS |
| 20 | VIENNA 7 2020 FF26 d | GSS |
| 21 | wp tsl FF21 a | mutant |
| 22 | wp tsl FF26 a | mutant |
| 23 | wp tsl (EgII) FF21 a | mutant |
| 24 | wp tsl (EgII) FF26 a | mutant |
| 25 | D53-3-28 FF21 a | mutant |
| 26 | D53-3-28 FF26 a | mutant |
| 27 | D53-1 FF26 | mutant |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sollazzo, G.; Gouvi, G.; Nikolouli, K.; Cancio Martinez, E.I.; Schetelig, M.F.; Bourtzis, K. Temperature Sensitivity of Wild-Type, Mutant and Genetic Sexing Strains of Ceratitis capitata. Insects 2022, 13, 943. https://doi.org/10.3390/insects13100943
Sollazzo G, Gouvi G, Nikolouli K, Cancio Martinez EI, Schetelig MF, Bourtzis K. Temperature Sensitivity of Wild-Type, Mutant and Genetic Sexing Strains of Ceratitis capitata. Insects. 2022; 13(10):943. https://doi.org/10.3390/insects13100943
Chicago/Turabian StyleSollazzo, Germano, Georgia Gouvi, Katerina Nikolouli, Elena I. Cancio Martinez, Marc F. Schetelig, and Kostas Bourtzis. 2022. "Temperature Sensitivity of Wild-Type, Mutant and Genetic Sexing Strains of Ceratitis capitata" Insects 13, no. 10: 943. https://doi.org/10.3390/insects13100943