
Ozone Efficiency on Two Coleopteran Insect Pests and Its Effect on Quality and Germination of Barley
by
 , , , and
, , , and
Xue Dong
1,2,† ,
,
Manjree Agarwal
2,†,
Yu Xiao
2,
Yonglin Ren
2,
Garth Maker
2,* and
Xiangyang Yu
1,2,* 1
Institute of Agricultural Resources and Environment, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China
2
College of Science, Health, Engineering and Education, Murdoch University, Perth 6150, Australia
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this manuscript.
Insects 2022, 13(4), 318; https://doi.org/10.3390/insects13040318
Submission received: 8 February 2022
/
Revised: 17 March 2022
/
Accepted: 22 March 2022
/
Published: 24 March 2022
(This article belongs to the Section Insect Pest and Vector Management)
Abstract
:Simple Summary
Rhyzopertha dominica (Fabricius) and Tribolium castaneum (Herbst) are notorious global pests, destroying various stored grains, including barley, wheat, oats, maize, and rice. Ozone (O3) is a promising fumigant to control pests in stored grain since it can safely and rapidly auto-decompose without leaving residues, however, relatively few studies have focused on the toxicity of O3 on stored grain pests in stored barley. In this study we not only explored the susceptibility of all life stages of R. dominica and T. castaneum in barley seeds to different durations of gaseous O3, but also investigated the effect of O3 on germination ability, seedling growth, and quality of barley. O3 was effective against all life stages of two species in barley under sufficient exposure times without negative impacts affecting the commercial quality of barley. However, the germination ability and seedling growth were adversely impacted at longer O3 exposure times. Thus, it is imperative to select an optimal O3 exposure time to achieve the desired functional outcome, such as malting, animal feeding, and human consumption.
Abstract
Ozone (O3) is a potential fumigant to control pests in stored grain since it can safely and rapidly auto-decompose without leaving residues. In this study, the efficacy of O3 on all life stages of Rhyzopertha dominica (Fabricius) and Tribolium castaneum (Herbst) in barley and the physiological effects on barley and its quality were investigated. Complete control of all life stages of pests was obtained at 700 ppm for 1440 min of ozone exposure without negatively impacting the contents of soluble protein, moisture content, seed colour, hardness, and the weight of thousand barley seeds. The eggs and pupae of these two insects were the more tolerant stages than their larvae and adults. Prolonged exposure times (40 to 1440 min) and mortality assessment intervals (1, 2, and 7 days) increased O3 efficacy due to the reaction characteristics and delayed toxicity. Aging barley seeds appeared to be more sensitive to prolonged ozone duration than new seeds. A total of 20 and 40 min could promote germination rate, and longer O3 exposure (1440 min) was unfavourable for germination and seedling growth. Thus, it is imperative to select an optimal O3 exposure time to transfer ozone into quality contributors of final products and achieve the desired functional outcomes.
1. Introduction
Postharvest loss has become a crucial issue of the grain supply chain. It has been estimated that one-quarter to one-third of the world’s grain crop is lost every year during storage, with the majority attributed to insect attack [1]. The lesser grain borer, Rhyzopertha dominica (Fabricius), and the red flour beetle, Tribolium castaneum (Herbst), are notorious global pests, destroying various stored grains, including barley, wheat, oats, maize, and rice. Both species can cause severe economic loss by decreasing the quantity and quality of stored grain. As a primary grain insect, R. dominica has the ability to attack whole kernels, while T. castaneum is a secondary pest and can only attack damaged grain, dust, and milled products [2]. Fumigation is often the cheapest and most effective process that is available, playing a major role world-wide in preserving commodities [3]. Therefore, currently, fumigation is a common method to eliminate grain storage insect pests. In actual practice, however, most gases have been eliminated owing to unfavourable properties, the most important being chemical instability and destructive effects on commodities [4]. Methyl bromide is a fast-acting fumigant, but it has been removed from fumigant registration because it caused depletion of ozone layer. Phosphine is the widely used fumigant, however, continuous use has led to the evolution of resistant populations and environmental contamination [5]. R. dominica was the first pest with strong resistance to phosphine recorded in Australia [6]. Nayak analyzed 20 years of resistance data in Australia, suggesting that resistance to phosphine had increased significantly over this period and is currently between 60–80% (depending on species). In North America, there was a significant increase in the frequency of resistance in both T. castaneum and R. dominica over two decades [7,8]. Opit suggested that the most resistant T. castaneum population was 119-fold more resistant than the susceptible strain and the most resistant R. dominica population was over 1500-fold more resistant in North America [9]. Strongly resistant populations of T. castaneum and R. dominica were also reported in Asia, including China [10,11], Pakistan [12], and India [13,14]. Ethyl formate is an alternative one, but it has high water solubility which could be rapid absorbed by grain, thus it just has been registered as a fumigant in dried fruit in Australia, not in grain industry [15]. Diatomaceous earths are used to control stored grain, causing insects death through absorbing the epicuticular lipids of the insect cuticle, but it has a negative effect on the physical properties of grains, particularly bulk density [16,17,18]. Controlled atmosphere with inert gases such as CO2 is an alternative non-chemical method to control grain pests, however, it requires long exposure times (more than 10 days) [19]. Entomopathogenic fungi (EF) are considered the most promising biocontrol agents, which infect insects through mycelium penetrating into the insects cuticle and then growing in the haemocoel causing insects death [20]. However, there are no commercial biopesticides based on EF bioagents that are registered against the stored-grain insects [20]. The main limitations of the application of EF in insect control are the high moisture requirement for conidial germination and subsequent sporulation, more time consuming to kill insects, and the potential risk to immunodepressive or immunocompromised people [21,22]. Other non-chemical methods such as cold or low temperature treatment and irradiation have also been reported to eliminate the insects in stored grain, however, they are costly for commercial treatment of large-scale bulk grain or need long-term treatment [23,24,25]. Therefore, alternative methods are needed to control grain storage pests.
Ozone (O3) displays distinct advantages which could overcome the deficiencies of other fumigants. O3 occurs naturally in the atmosphere and could be generated on-site without transportation problems. Ozone can rapid auto-decompose to oxygen without leaving residues, along with a wide spectrum of activity against microorganisms, making it an attractive approach for medical sterilisation, food processing, and grain storage [26,27,28]. There are more than 100 years of using ozone in food industry and it was firstly used as food preservative of frozen meat in 1910 [29]. In 2001, The United States Food and Drug Administration (FDA) approved O3 as a direct additive for food treatment, storage, and processing [30]. For fruit processing, Sadeghi et al. [31] applied ozone to control Oryzaephilus surinamensis (Coleoptera: Silvanidae) and Ephestia kuehniella (Lepidoptera: Pyralidae) in stored dried figs, reporting that increasing the ozone concentration and times had no significant influence on the quality parameters of dried figs and raisins, including colour, sweetness, sourness, brittleness, hardness, and general acceptance. Ozone is also effective at eliminating storage pests, microbes, and degrading mycotoxins in grain [32,33,34]. There has been growing emphasis on the application of O3 to control R. dominica and T. castaneum. Sousa et al. [35] reported that 150 ppm gaseous O3 presented a high efficacy against both susceptible and phosphine-resistant insects from 16 populations of T. castaneum and 11 populations of R. dominica. O3 was also lethal to different stages of R. dominica and T. castaneum [36,37]. O3 presented a different efficacy against insects inside or outside the grain, mainly because grains can increase the decomposition of O3 [27]. The reaction of O3 within a grain can be divided into two phases. The first phase is that O3 interacts with active sites that are present in or on the kernel surface, causing the decomposition of O3. The second phase is the free movement of O3 through the grain layers once the reactive sites are saturated, with O3 concentration gradually increasing to an effective dosage for target pests [38]. The penetration of ozone into the bulk of grains also depends on gas diffusion, air speed within the grain layer, initial ozone concentration, bed thickness, temperature, and adsorption by the grain surface [39,40].Various grains have distinct effects on O3 insecticidal efficacy, which is attributed to differences in the surface and size of kernels.
The successful application of O3 against stored grain insects requires sufficient concentration and exposure time, however it may affect the grain quality. Zhu [41] reviewed that moderate O3 treatment could facilitate the dough strength, enhance the storage stability, and the whiteness of wheat flour. The malting quality of barley, such as alpha-amylase, fine grind extract, dynamic viscosity, and soluble protein were not impacted by O3 treatment of 26 mg/g [42]. Conversely, as a strong oxidant, O3 can react with some substrates in grains directly or via producing reactive oxygen species. Starch and lipid oxidation, protein modifications, grain discolouration, odour alteration, and germination loss may result from excessive use of O3 [43].
There were discrepancies in the effect of O3 on grain quality when the dosage of O3 was sufficient for eliminating insects pests in grain [43,44]. These discrepancies might be mainly caused by the differences of dose, duration, and grains. While some studies have assessed the efficacy of O3 against pests in maize and wheat, relatively few studies have focused on the toxicity of O3 on stored grain pests with barley. Thus, this study aims to explore the susceptibility of all life stages of R. dominica and T. castaneum in barley seeds to different durations of gaseous O3, and to investigate the effect of gaseous O3 on germination ability, seedling growth, and quality of barley with different storage times. Additionally, the different response of old and new barley seeds that were subjected to ozone treatment was also examined.
2. Materials and Methods
2.1. Insect Rearing
There were two insect species of stored product insects that were used for bioassays, these were Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae) and Rhyzopertha dominica (F.) (Coleoptera: Bostrychidae). They were the phosphine-susceptible MUWTC-6000 and MUWRD-7 strains, respectively, that were held at the Postharvest Biosecurity and Food Safety Laboratory, Murdoch University, Australia. The narrow-aged insects (2–3 days) were obtained by incubating 3000 adult insects with 1000 g of food, broken wheat Australian Standard White (ASW) for R. dominica and wheat flour/yeast 12:1 ratio for T. castaneum in 2 L jars that were sealed with meshed lids. The parent insects were removed after 3 days and the remaining culture medium were incubated in 2 L glass jars at 28 ± 1 °C and 70 ± 2% relative humidity (RH) with a 0:24 (L:D) h photoperiod. The newly emerged adults were narrow aged and transferred to a 2 L jar containing fresh food. The insects that were used in the experiments were one month old.
2.2. Barley Samples
Barley samples belonging to Sparticus CL were harvested in 2013/2014 and 2019/2020 from Lake Grace, Western Australia. The sample that was harvested in 2013/2014 was stored at 4 °C for 6 years. In the grain production business, the long-term storage of grain often occurs due to the relatively limited demand for consumption, cultivation, unexpected meteorological disasters, and also to prevent a change in specific characters [45]. The moisture content of the 2013/2014 and 2019/2020 harvest barley were 12.3% and 10.9%, determined with a Graintec HE 50 electronic moisture meter (Graintec Pty Ltd., Toowoomba, Australia). The results that were obtained were calculated from four replicate measurements. The insect-free samples were stored at 4 °C and equilibrated to room temperature before germination tests and quality measurements were conducted to room temperature at 25 ± 1 °C.
2.3. Fumigation Procedures with Ozone
A commercial O3 generator (Model FH-CYJ1520A-20 g/h, Shanghai Fenghua Optoelectronics Technology Co. Ltd., Shanghai, China) was utilized to treat the barley seeds. This generator continuously produced 700 ppm O3 from atmospheric air. The ozonation of barley seeds was carried out in a 2.4 L and 16.6-cm diameter Hysil semi-batch reactor with a desiccation glass chamber that was fitted with a lid containing a 2-cm central hole for ozonation. The gas was introduced from the ozone generator directly to the bottom of the reactor containing 1 kg of barley and passed through the seeds using a 1-cm diameter plastic tubing. Prior to introducing the O3, the barley seeds (1 kg) were mixed with 3 g adults (about 2000) and 7 g egg. The pupae and larvae were placed on a supporting metal mesh with 13-cm diameter, and 5 cm from the bottom to enable gas circulation. The ozone concentration to and from the reactor was continuously measured by an ozone monitor (Shenzhen Yuan Technology Co., Ltd., Shanghai, China). A total of nine treatments were designed for this study, comprising of a control (fresh air pass through the glass chamber), 700 ppm O3 exposure for 0, 10, 20, 40, 120, 240, 480, 960, and 1440 min, with three replicates. All the treatments were performed in ambient conditions at 20 ± 5 °C and 55 ± 3% RH.
2.4. Assessment of Insect Mortality
For the bioassay samples, all the adults, larvae, pupae, and eggs of the treated and untreated controls were retrieved at the end of the fumigation period from the above-treated and untreated barley with 710 and 180 μm sieves. The adults were removed and placed into 250 mL vials with food and incubated at 28 ± 1 °C and 70 ± 2% RH. The live and dead adult insects were counted at different assessment intervals, including immediately (0DAT), 1, 2, and 7 days after O3 treatment (1DAT, 2DAT, 7DAT). The remaining mixed-age cultures were incubated at 28 ± 1 °C and 70 ± 2% RH. The subsequent emerging adult insects were counted weekly for a period of 5 weeks, with live and dead adults removed at each count. The mortality was calculated based on a comparison of the emerging adults between the treated and untreated control samples. Once all the control insects emerged, the experiment was terminated.
2.5. Germination Test
The germination test was carried out based on the between-paper (BP) method of the International Seed Testing Association methods [46]. A total of 424 seeds (eight replicates of 53 seeds) in each harvest year were randomly selected. A large filter paper was saturated with 60 mL distilled water and folded in half, creating a double thickness. A steel template (290 × 580 mm) was placed over this wet paper, with holes in this stencil allowing the barley seeds to be placed 30 mm apart. Once the seeds were positioned correctly, the template was removed and the upper half of the filter paper was folded over the seeded area. The folded paper containing the seeds was then loosely rolled from one side perpendicular to the base and tied with elastic bands. All the treatments were stored individually in polythene zip bags and incubated for 7 days at 25 ± 1 °C in the dark.
The germination rate was detected 7 days after the start of the germination test. The morphological characteristics were tested, including the length of the shoot, the length of root, and the ratio of root length to shoot length on Day 7. The seedling vigour index was calculated using the below formula according to Islam et al. [47].
where: Y is Seedling Vigour Index (SVI); a is the mean root length (cm); b is the mean shoot length (cm); and c is the percentage of germination rate (%).
Y = (a + b) c/100
2.6. Grain Quality Measurement
Duplicate barley samples were randomly taken from each treatment for quality measurement. A near infrared range (NIR) instrument (Infratec™ 1241 Grain Analyser, Foss, Melbourne, VIC, Australia) was utilised for analysing the grain quality. The functioning of this analyser is based on the measurement of transmission spectra of samples in the near-infrared region [48,49]. The instrument had an extended wavelength range of 570–1100 nm. A total of 10 subsamples of whole kernels were scanned for each grain sample. The parameters we assessed in this research included the total soluble protein content, moisture content (MC), colour, hardness, and thousand weight (TW) of grain. The colour results were reported as lightness, which was determined by L* values (0 = black and 100 = white). TW was estimated by weighing 1000 seeds in grams.
2.7. Statistical Analysis
Data from the time-response bioassays were subjected to Probit analysis (IBM SPSS Statistics, version25.0). Time-mortality curves and LT50 and LT95 were generated. A one-way analysis of variance (ANOVA) and Tukey’s Honestly Significant Difference (HSD) test were employed to evaluate if there was a significant difference between the O3 treatments and control. The results were considered significant only when p ≤ 0.05. Origin 2019b was used for curve figures.
3. Results
3.1. Toxicity of O3 on R. dominica and T. castaneum Adults at Different Assessment Intervals
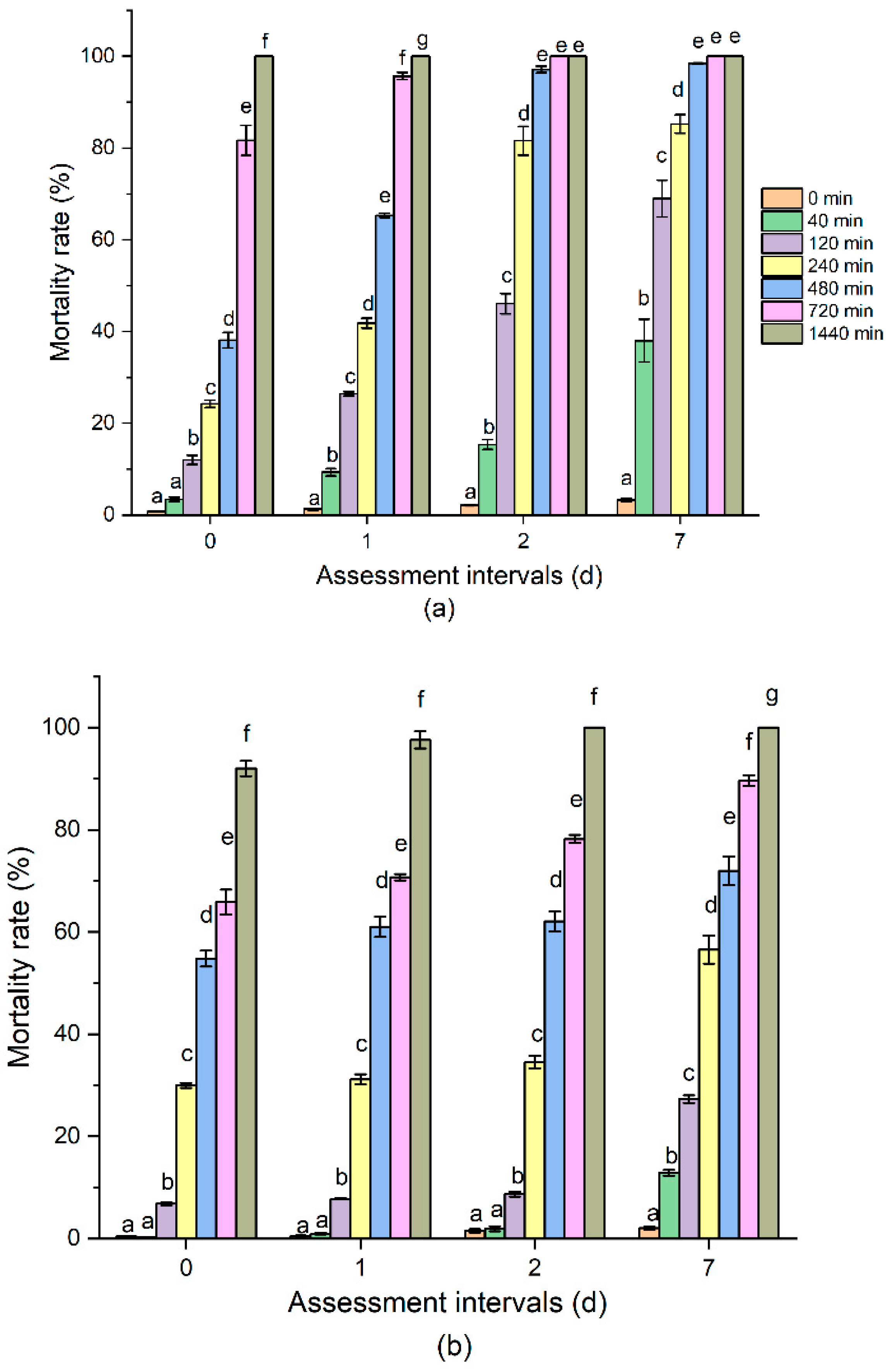
The mortality rate of both the insect species increased with prolonged O3 exposure time and assessment interval (Figure 1). In terms of R. dominica, the mortality rate significantly increased with extended O3 exposure time. Adults of R. dominica and T. castaneum in barley could be knocked down immediately at 700 ppm for 2 h exposure, but 90% of them can be recovered after treatment. However, 1440 min of O3 treatment could kill 92% adults immediately, 97% at 1DAT, and 100% adults at 2 and 7DAT. T. castaneum displayed a similar tendency when subjected to different O3 exposure times and assessed at intervals after O3 treatment. The mortality rate was 81% at 0DAT, and then increased to 95% at 1DAT, and 100% at 2 and 7DAT when subjected to 960-min treatment. Time–mortality regression for the populations of R. dominica and T. castaneum that were exposed to O3 at different assessment intervals are represented in Table 1. A total of 700 ppm ozone of each treatment time (0, 40, 120, 240, 480, 960, and 1440 min) for the two species was conducted. The Pearson goodness-of-fit Chi-square (χ2) test showed that the probit model fit to the data was significant (p < 0.05), thus, a heterogeneity factor (χ2/df) was used in the calculation of the confidence limits. The LC50 values were estimated for each species at different assessment intervals. At 700 ppm O3, the median lethal time (LT50) of R. dominica on 0DAT was 472.91 (343–653.55) min and that of T. castaneum on 0DAT was 453.07 (228.14–769.72) min. The LT90 values for R. dominica and T. castaneum on 0DAT were 2419.93 (1465.95–6336.68) and 1823.12 (972.31–27,304.4) min, respectively. The data indicated that R. dominica was more tolerant to the O3 treatment. Additionally, the LT50 and LT90 of both insects at O3 treatment dramatically decreased on 1DAT, 2DAT, and 7DAT compared with 0DAT. The LC50 of R. dominica and T. castaneum on 7DAT were the lowest, which were 213.50 and 63.60 min, respectively. The relative potency for R. dominica and T. castaneum was 2.22 and 7.12, respectively. Our results indicated the delayed toxicity of O3 and that it is better to evaluate the O3 efficacy against these two species more than 1 day after treatment.
3.2. Toxicity of O3 Duration on All Life Stages of R. dominica and T. castaneum
The mortality rate of all life stages for R. dominica and T. castaneum that were exposed to different O3 durations is summarised in Table 2. Based on our preliminary study, 10 and 20 min O3 exposure could not kill the insects, so an O3 duration that was longer than 40 min was investigated. A total of 1440 min of O3 treatment could kill all stages of both species. The mortality rate of R. dominica adults significantly increased with extended O3 durations and reached 89.62% at 960-min exposure (F5, 12 = 29.334, p < 0.001), while T. castaneum reached 91.39% at 240-min exposure and 100% at 960-min treatment (F5, 12 = 209.197, p < 0.001). The mortality rate of both R. dominica and T. castaneum eggs, larvae and pupae also considerably increased with extended O3 durations and reached 83.33%, 91.67%, and 71.30% at 960-min exposure, while those of T. castaneum reached 81.11%, 100%, and 93.33% at 960-min treatment, respectively. In Table 2, the different letters in each life stage indicate significant mortality differences of the ozone treatment times between the two species. The results indicate that except for the egg stage, T. castaneum was more sensitive to the O3 treatment than R. dominica. Considerable variation was observed in susceptibility among all the life stages of R. dominica and T. castaneum. In terms of R. dominica, the adults and larvae were more susceptible to the O3 treatment than the pupae and eggs. The adults and larvae achieved 86.92% and 91.67% mortality with 960 min O3 treatment, however the pupae and eggs reached 74.30% and 83.33% mortality. Regarding T. castaneum, the observed mortality was 100% for the larvae and adults and 93.33% for the pupae, compared to 81.11% for the eggs with 960 min exposure.
3.3. Effect of O3 Treatment Duration on Germination of Barley
The germination rate is one of the most critical parameters that contributes to predicting final crop yield, which was measured 7 days after sowing. The effect of O3 on the germination of barley seeds are presented in Table 3. The seeds that were harvested in 2013/2014 and 2019/2020 are presented as old and new barley in this article. The germination rate of the old harvest seeds increased from 94.34% to 97.17% at 40 min ozonation, while that of the new barley seeds increased from 98.35% to 99.06% at 10 and 20 min ozonation. However, the long duration of the O3 treatment, such as 480, 960, and 1440 min has been observed to have a considerably negative effect on the germination rate of old harvest seeds. Regarding new seeds, the germination rate started to significantly decrease under 960 and 1440-min O3 exposure compared with the untreated seeds (p < 0.05). In both old and new barley seeds, O3 application times of 1440 min were accompanied by dramatical decreases in the germination rates to 14.39 % and 20.28%, respectively.
3.4. Effect of O3 Treatment Duration on Seedling Growth
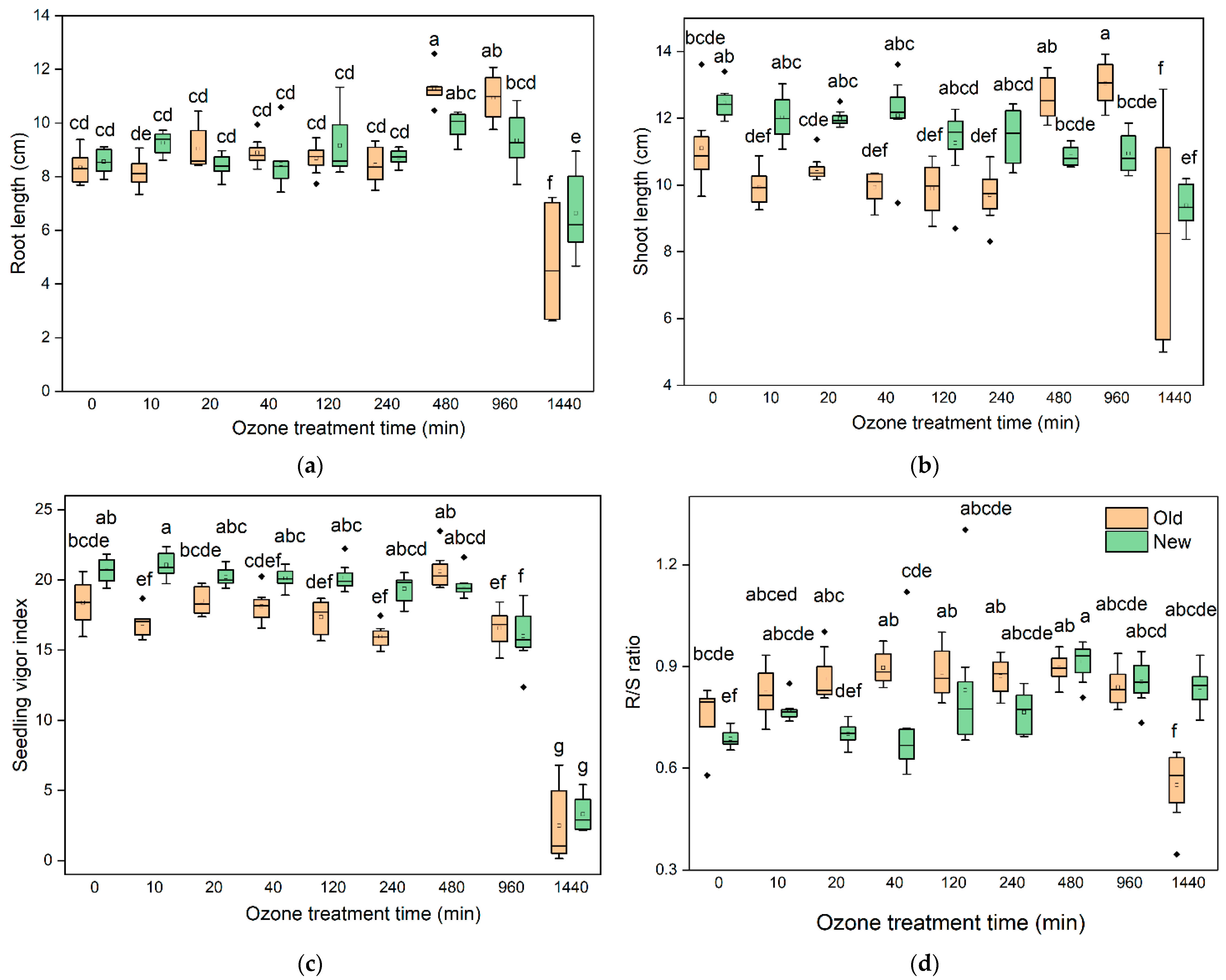
The effect of O3 on the morphological parameters (seedling length and vigour index) of barley seeds are presented in Figure 2. In terms of the total root length of new barley, there was no significant difference between 10, 20, 40, 120, 240, 480, and 960 min ozonation and the untreated samples. However, after 1440 min treatment, the shoot length decreased (6.65 cm) compared to the untreated samples (8.56 cm). For the old barley, 480 and 960 min O3 treatment significantly increased the root length to 11.28 and 10.95 cm from 8.34 cm, whereas the root length was significantly decreased to 4.79 cm at a longer exposure time (1440 min) compared with that of the control (8.34 cm) (Figure 2a). Regarding the shoot length, the new barley shoot length was not significantly inhibited by 10, 20, 40, 120, 240, 480, or 960-min O3 treatment, but was significantly inhibited at 1440 min ozonation. For the old barley seeds, there was a positive effect of O3 on the shoot length at 480- and 960-min treatment, which increased the shoot length to 12.61 cm and 13.05 cm compared with the untreated sample (11.11 cm). The shoot length was inhibited (8.49 cm) with the extension of O3 to 1440 min (Figure 2b).
The seedling vigour index (SVI) is an important parameter to reflect the potential field performance of different seeds. The SVI data is presented in Figure 2c. Regarding the new barley seeds, in comparison to the control, the SVI showed no significant difference under short duration ozonation, including 10, 20, 40, 120, 240, and 480 min. However, it dramatically decreased as the duration time increased to 960 and 1440 min (p < 0.05 value). In terms of the old seeds, only 1440 min O3 treatment significantly decreased the SVI compared with the untreated samples (p < 0.05). The ratio of root-to-shoot length (R/S ratio) in both the new and old barley seeds significantly decreased at 1440 min O3 treatment (p < 0.05). In addition, the R/S ratio in new barley seeds increased when they were subjected to 480 and 960 min exposure, however, in the old barley seeds there was no significant difference between 10, 20, 40, 120, 240, 480, or 960 min of O3 treatment and the control (Figure 2d).
3.5. Effect of O3 on Barley Grain Quality
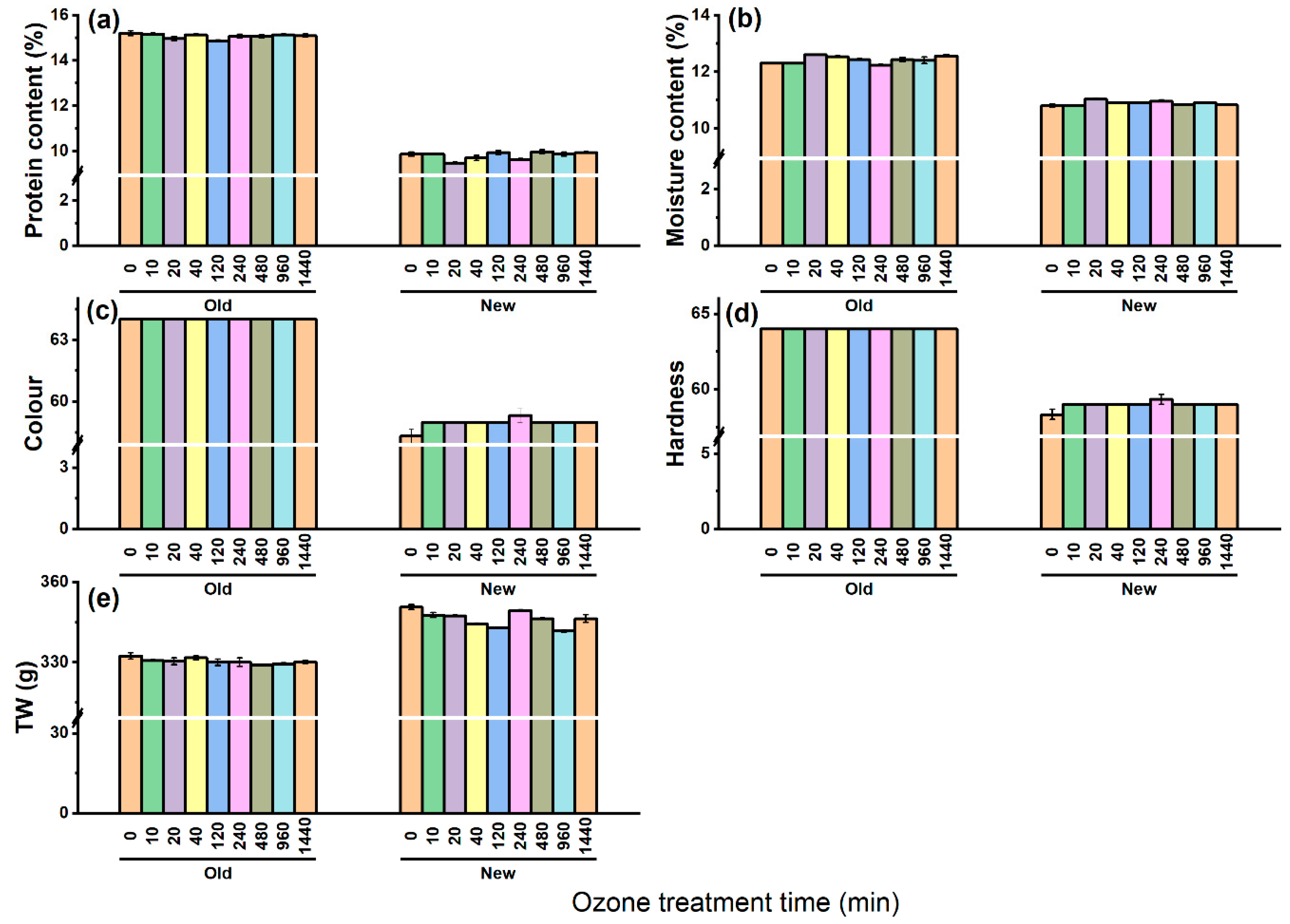
The soluble protein content of old and new barley seeds was 15.2% and 9.9%, respectively (Figure 3a). It can be observed that there was no significant effect of O3 on the soluble protein content in both samples, irrespective of the age. The moisture content of the old and new barley varied from 12.2% to 12.6% and 10.7% to 11%, respectively, and showed no change with increasing O3 duration in both the sample groups (Figure 3b). From Figure 3c, the old barley had a higher lightness value than the new barley. The colour of both the new and old barley was not affected by the O3 duration. As can be seen from Figure 3d, the hardness values for the old and new barley were 55.3 and 45.3, respectively. There was a less apparent trend towards increasing hardness of both the old and new seeds, characterised by small fluctuations up to 58 and 47.6. Generally, the TW of both the old and new barley was not considerably impacted by extended periods of O3 treatment. In terms of the old barley, the TW was decreased from 33.2 to 32.9 g, while the new barley was reduced from 35.1 to 34.2 g (Figure 3e). Overall, the value of the soluble protein content, moisture content, colour, and hardness of old barley were higher, and only thousand weight (TW) was lower than that of the new barley seeds (Figure 3).
4. Discussion
The results suggest that O3 has efficacy against both R. dominica and T. castaneum, with the toxicity data indicates that mortality occurs in a relatively short interval and increased with prolonged O3 treatment. Sufficient exposure time is required to kill insects since disinfestation action of O3 takes place mainly at the surface of the grain, unless saturation of the grain is reached [44]. It was observed that 960-min O3 treatment could kill 65% and 81% of R. dominica and T. castaneum, even if assessed immediately after O3 treatment. Lower exposure time led to a higher mortality rate of insects using longer assessment intervals. Thus, both exposure times and mortality assessment intervals should be considered to evaluate pesticide efficacy.
The assessment intervals of detecting insect mortality to pesticides should be varied based on insecticide mode of actions, namely 1–2 days for fast-acting insecticides, and 7–14 days for slow-acting insecticides. Our results indicated the delayed toxicity of O3 since the mortality of both species increased up to seven days, which is in accordance with Subramanyam et al. and Holmstrup et al. [50,51]. The mechanism of ozone killing insects is still elusive. There are two main hypothesised mechanisms. The first one is related to the respiratory rate. The respiratory system of insects is considered to be the first site of action for O3, as it enters insects through spiracles causing disrupted respiration in insects. Discontinuous gas exchange could lower insect metabolisms causing insect death [52]. The second hypothesis is based on strong the oxidative properties of O3, which could cause oxidative damage of essential components resulting in DNA strand breaks and cell membrane oxidation [53]. Another possible reason is that gene expression and damage to DNA exposures do not occur immediately after O3 exposure, but in the post-exposure period [51].
Toxicity data indicated a noticeable difference in the susceptibility between R. dominica and T. castaneum, especially at different life stages. In general, all life stages except for eggs of R. dominica have a higher tolerance to O3 than T. castaneum. In addition, longer O3 exposure times at 700 ppm were required for eggs and pupae of both species to reach higher mortality compared with larvae, which agrees with previous research [36,54]. Distinct responses of life stages to O3 could be attributed to their respiration rates. The respiratory system of insects is considered as the first site of action for O3. On the basis of phosphine studies [55], we suggest that the low respiration rate of pupae and eggs might reduce the O3 uptake resulting in greater tolerance [56]. Furthermore, the majority of the outer layer of eggs and pupae are made of lipids, sometimes covered with a waxy coat which could provide an additional barrier against O3.
The current results indicate that, to obtain 100% mortality of all stages of both species, the required exposure time of O3 at 700 ppm was 1440 min. The short duration O3 treatment did not significantly influence the germination ability in either the old or new barley. However, with increasing duration, the germination ability was strongly impacted. These observations are in agreement with Wu et al. [57], who suggested that the germination ability of stored wheat was damaged with prolonged O3 treatment. Peroxide or superoxide accumulation, which is elicited by O3, can damage antioxidant mechanisms, reduce the activity of related enzymes, and disturb macromolecule biosynthesis, resulting in suppression of the germination process. This is problematic, as quality of barley is crucial in determining its end utilisation. For example, the germination rate of barley seed must exceed 95% for malting [58]. Therefore, a high seed germination ability plays a pivotal role in improving crop production and shortening the manufacturing costs of the malting process. Some studies have indicated that moderate duration ozonation can accelerate the germination of tomato seeds because reactive oxygen species (ROS) that are produced by O3 promote cell signalling [59]. The ROS break seed dormancy and promote germination by regulating hormone levels and reducing abscisic acid levels [60]. In this study, the germination ability of two samples of different ages was enhanced under moderate ozonation time, especially for old seeds, which increased from 94.34% to 97.17% at 40 min O3 treatment, improving the germination ability of aging barley to meet the malting barley grade. Further research needs to be conducted, specifically utilising seeds with a lower germination ability to explore if moderate O3 processing time can significantly increase the germination ability.
This study also suggested that prolonged O3 exposure (1440 min) was unfavourable for seedling growth. Both the root and shoot length were decreased with extending O3 duration to 1440 min, which is consistent with reports that maize seedling length decreased with increasing O3 processing time [61]. Additionally, the root length of both old and new barley significantly increased under moderate O3 processing time. Seedling growth is a cell division and expansion process, and endogenous reactive oxygen species (ROS), such as hydrogen peroxide (H2O2) that are induced by O3, play essential roles in cell growth [62]. The root-to-shoot ratio is a good indicator of the overall health and physiology of plants. It is generally genetically fixed for each cultivar but can be modified by environmental factors [63]. The ratio of the root length to the shoot length increased when the seeds were subjected to longer O3 exposure time, signifying that plants reduce shoot growth and enhance root growth in response to O3 stress. Alexander et al. [64] suggested that the root-to-shoot ratio of spring wheat was increased from increasing O3 concentration. Plants apportioning more biomass to roots and less to stems when they encounter unfavourable conditions has been previously established [65].
Quality characteristics including the soluble protein content, colour, and TW may affect the final market price of grain. In this research, different O3 treatment times did not influence the soluble protein content, moisture content, TW, and colour of boththe old and new seeds, which closely correlated with previous studies [66,67]. Seed storage is one of the most important factors that affect seed quality. Seed reserves are lost during storage, as manifested by the lower TW of old seeds. The total soluble protein was higher in the old seeds compared with the new ones, consistent with Lozano et al. [68] who reported that the total soluble protein and amino acid content of Jatropha curcas L. seeds increased by 160% and 67% during storage, respectively. A possible explanation could be the consequence of soluble protein or amino acid that is converted from storage proteins to maintain respiration and respond to stress during storage [69]. As the energy resources are used by the embryo during germination, organic compounds in the seeds continually reduce due to respiration processes after harvest, resulting in an energy deficiency during germination and early seedling development. This explains why the old seeds in this study appeared to have a lower germination ability and seedling growth capacity compared to the new seeds. Additionally, old seeds tend to be more susceptible to O3 stress. The germination rate of the old seeds significantly reduced at 480 min ozonation, whereas that of the new barley did not significantly decrease until 960 min exposure.
Although the morphological characteristics and germination were adversely influenced by excess ozone exposure, that can be regulated to achieve desired outcomes if proper measures are taken. For instance, if the barley seeds are utilised for livestock feeding and human consumption, it is necessary to supply sufficient ozone treatment to eliminate pests rather than achieve high germination ability. Most farmers grow barley for sale as malting barley where germination ability and other malting quality parameters could also be maintained and even improved based on the proper application of ozone. This research found that short ozone exposure could enhance the germination ability of aging seeds to meet malting barley requirements. All these studies make it evident that ozone has great potential to improve the functionalities of grain products while ensuring food safety.
5. Conclusions
More than two days after ozone treatment should be considered as the endpoint to evaluate insects’ mortality rates due to delayed toxicity of O3. C × t product 36 mg h/L (700 ppm × 24 h) of ozone offered complete mortality for all stages of the two species of tested insects at an endpoint of dead, which is highly toxic than phosphine and enthyl formate and without quality parameters including soluble protein content, moisture content, colour, hardness, and TW. The pupae and eggs were more tolerant to O3 than the larvae and adults. Moderate ozone treatment time such as 20 min and 40 min increased the germination ability to greater than 95%, however, the germination ability and seedling growth of both the old and new barley seeds were impaired due to long exposure to O3 (1440 min), especially for the longer storage seeds. Overall, the data demonstrated that the multiple effects of ozone treatment on the parameters of barley seeds could be utilised to control the quality attributes of final food products, thus, it is imperative to select an optimal O3 exposure time to achieve the desired function, such as malting, animal feeding, and human consumption. Future investigations are needed to evaluate the O3 efficacy against insects in large-sized silos, as well as elaborate on how metabolites in barley seeds shift in response to O3 treatment.
Author Contributions
Conceptualization, X.D., G.M. and X.Y.; methodology, X.D., M.A. and Y.X.; software, X.D.; validation, X.D., Y.X. and M.A.; formal analysis, X.D.; investigation, X.D., M.A. and Y.X.; resources, Y.R.; data curation, X.D.; writing—original draft preparation, X.D.; writing—review and editing, X.D., G.M., M.A. and X.Y.; visualization, X.D. and M.A.; supervision, G.M., Y.R. and X.Y.; project administration, Y.R. and X.Y. All authors have read and agreed to the published version of the manuscript.
Funding
There is no funding report in this research.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mesterházy, Á.; Oláh, J.; Popp, J. Losses in the grain supply chain: Causes and solutions. Sustainability 2020, 12, 2342. [Google Scholar] [CrossRef] [Green Version]
- Shankar, U.; Abrol, D.P. Integrated Pest Management in Stored Grains. In Integrated Pest Management: Principles and Practice; CABI: Wallingford, UK, 2012; pp. 386–407. [Google Scholar]
- Banks, H. In Behaviour of gases in grain storages, Fumigation and Controlled Atmosphere Storage of Grain. In Proceedings of the International Conference, Singapore, 4–18 February 1989; pp. 96–107. [Google Scholar]
- Bond, E.J.; Monro, H.A.U. Manual of Fumigation for Insect Control; FAO: Rome, Italy, 1984; Volume 54. [Google Scholar]
- Nayak, M.K.; Daglish, G.J.; Phillips, T.W.; Ebert, P.R. Resistance to the fumigant phosphine and its management in insect pests of stored products: A global perspective. Annu. Rev. Entomol. 2020, 65, 333–350. [Google Scholar] [CrossRef] [Green Version]
- Collins, P.J. Resistance to grain protectants and fumigants in insect pests of stored products in Australia. In Proceedings of the Australian Postharvest Technical Conference, Canberra, Australia, 25–27 June 2003; pp. 55–57. [Google Scholar]
- Cato, A.; Elliott, B.; Nayak, M.K.; Phillips, T.W. Geographic variation in phosphine resistance among North American populations of the red flour beetle (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2017, 110, 1359–1365. [Google Scholar] [CrossRef]
- Gautam, S.; Opit, G.; Hosoda, E. Phosphine resistance in adult and immature life stages of Tribolium castaneum (Coleoptera: Tenebrionidae) and Plodia interpunctella (Lepidoptera: Pyralidae) populations in California. J. Econ. Entomol. 2016, 109, 2525–2533. [Google Scholar] [CrossRef]
- Opit, G.; Phillips, T.W.; Aikins, M.J.; Hasan, M. Phosphine resistance in Tribolium castaneum and Rhyzopertha dominica from stored wheat in Oklahoma. J. Econ. Entomol. 2012, 105, 1107–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Li, F.; Liu, M.; Wang, Y.; Shen, F.; Tang, P. Susceptibility of Tribolium castaneum to phosphine in China and functions of cytochrome P450s in phosphine resistance. J. Pest Sci. 2019, 92, 1239–1248. [Google Scholar] [CrossRef]
- Song, X.; Wang, P.; Zhang, H. Phosphine resistance in Rhyzopertha dominica (Fabricius) (Coleoptera: Bostrichidae) from different geographical populations in China. Afr. J. Biotechnol. 2011, 10, 16367–16373. [Google Scholar]
- Wakil, W.; Kavallieratos, N.G.; Usman, M.; Gulzar, S.; El-Shafie, H.A. Detection of phosphine resistance in field populations of four key stored-grain insect pests in Pakistan. Insects 2021, 12, 288. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, S.; Nayak, K.R.; Anjum, S.S. The action of phosphine against the eggs of phosphine-resistant and-susceptible strains of Rhyzopertha dominica F. Pest Manag. Sci. Former. Pestic. Sci. 2001, 57, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, S. Phosphine resistance in stored grain insect pests in India. In Proceedings of the 7th International Working Conference on Stored-Product Protection, Berlin, Germany, 7–11 October 2018; pp. 14–19. [Google Scholar]
- Finkelman, S.; Lendler, E.; Navarro, S.; Navarro, H.; Ashbell, G. New prospects for ethyl formate as a fumigant for the date industry. In Proceedings of the 10th International Working Conference on Stored Product Protection, Estoril, Portugal, 27 June–2 July 2010; pp. 359–364. [Google Scholar]
- Athanassiou, C.; Korunic, Z.; Kavallieratos, N.; Peteinatos, G.; Boukouvala, M.; Mikeli, N. New trends in the use of diatomaceous earth against stored-grain insects. In Proceedings of the 9th International Working Conference on Stored Product Protection, Campinas, Brazil, 15 October 2006; pp. 730–740. [Google Scholar]
- Korunic, Z.; Cenkowski, S.; Fields, P. Grain bulk density as affected by diatomaceous earth and application method. Postharvest Biol. Technol. 1998, 13, 81–89. [Google Scholar] [CrossRef]
- Ziaee, M.; Ebadollahi, A.; Wakil, W. Integrating inert dusts with other technologies in stored products protection. Toxin Rev. 2021, 40, 404–419. [Google Scholar] [CrossRef]
- Kumar, H.; Vijay, V.K.; Subbarao, P.M.; Chandra, R. Studies on the application of bio-carbon dioxide as controlled atmosphere on pest management in wheat grain storage. J. Stored Prod. Res. 2022, 95, 101911. [Google Scholar] [CrossRef]
- Batta, Y.A.; Kavallieratos, N.G. The use of entomopathogenic fungi for the control of stored-grain insects. Int. J. Pest Manag. 2018, 64, 77–87. [Google Scholar] [CrossRef]
- Singh, D.; Raina, T.K.; Singh, J. Entomopathogenic fungi: An effective biocontrol agent for management of insect populations naturally. J. Pharm. Sci. Res. 2017, 9, 833. [Google Scholar]
- Rumbos, C.I.; Athanassiou, C.G. Use of entomopathogenic fungi for the control of stored-product insects: Can fungi protect durable commodities? J. Pest Sci. 2017, 90, 839–854. [Google Scholar] [CrossRef]
- Sutar, S.A.; Thirumdas, R.; Chaudhari, B.B.; Deshmukh, R.R.; Annapure, U.S. Effect of cold plasma on insect infestation and keeping quality of stored wheat flour. J. Stored Prod. Res. 2021, 92, 101774. [Google Scholar] [CrossRef]
- Carocho, M.; Antonio, A.L.; Barreira, J.C.; Rafalski, A.; Bento, A.; Ferreira, I.C. Validation of gamma and electron beam irradiation as alternative conservation technology for European chestnuts. Food Bioprocess Technol. 2014, 7, 1917–1927. [Google Scholar] [CrossRef] [Green Version]
- Navarro, S. The use of modified and controlled atmospheres for the disinfestation of stored products. J. Pest Sci. 2012, 85, 301–322. [Google Scholar] [CrossRef]
- Dufresne, S.; Hewitt, A.; Robitaille, S. Ozone sterilization: Another option for healthcare in the 21st century. Am. J. Infect. Control. 2004, 32, E26–E27. [Google Scholar] [CrossRef]
- Isikber, A.A.; Athanassiou, C.G. The use of ozone gas for the control of insects and micro-organisms in stored products. J. Stored Prod. Res. 2015, 64, 139–145. [Google Scholar] [CrossRef]
- Rice, R.G.; Graham, D.M.; Lowe, M.T. Recent ozone applications in food processing and sanitation. Food Saf. Mag. 2002, 8, 10–17. [Google Scholar]
- Carletti, L.; Botondi, R.; Moscetti, R.; Stella, E.; Monarca, D.; Cecchini, M.; Massantini, R. Use of ozone in sanitation and storage of fresh fruits and vegetables. J. Food Agric. Environ. 2013, 11, 585–589. [Google Scholar]
- FDA, United States Food and Drug Administration. Rules and Regulations, Part 173-Secondary Direct Food Additives Permitted in Food for Human Consumption (21 CFR Part 173 Authority: 21 USC. 321, 342, 348). Fed. Regist. 2001, 66, 123. [Google Scholar]
- Sadeghi, R.; Mirabi Moghaddam, R.; Taghizadeh, M. Application of ozone to control dried fig pests—Oryzaephilus surinamensis (Coleoptera: Silvanidae) and Ephestia kuehniella (Lepidoptera: Pyralidae)—And its organoleptic properties. J. Econ. Entomol. 2017, 110, 2052–2055. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.; Wu, J.; Doan, H. Inactivation of fungi associated with barley grain by gaseous ozone. J. Environ. Sci. Health Part B 2003, 38, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Bonjour, E.; Opit, G.; Hardin, J.; Jones, C.; Payton, M.; Beeby, R. Efficacy of ozone fumigation against the major grain pests in stored wheat. J. Econ. Entomol. 2011, 104, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Savi, G.D.; Piacentini, K.C.; Bittencourt, K.O.; Scussel, V.M. Ozone treatment efficiency on Fusarium graminearum and deoxynivalenol degradation and its effects on whole wheat grains (Triticum aestivum L.) quality and germination. J. Stored Prod. Res. 2014, 59, 245–253. [Google Scholar] [CrossRef]
- Sousa, A.D.; Faroni, L.D.A.; Guedes, R.; Tótola, M.; Urruchi, W. Ozone as a management alternative against phosphine-resistant insect pests of stored products. J. Stored Prod. Res. 2008, 44, 379–385. [Google Scholar] [CrossRef]
- McDonough, M.X.; Mason, L.J.; Woloshuk, C.P. Susceptibility of stored product insects to high concentrations of ozone at different exposure intervals. J. Stored Prod. Res. 2011, 47, 306–310. [Google Scholar] [CrossRef]
- Mishra, G.; Palle, A.A.; Srivastava, S.; Mishra, H.N. Disinfestation of stored wheat grain infested with Rhyzopertha dominica by ozone treatment: Process optimization and impact on grain properties. J. Sci. Food Agric. 2019, 99, 5008–5018. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Faroni, L.; Sousa, A.; Prates, L.; Abreu, A. Kinetics of the ozone gas reaction in popcorn kernels. J. Stored Prod. Res. 2019, 83, 168–175. [Google Scholar] [CrossRef]
- Pandiselvam, R.; Thirupathi, V.; Anandakumar, S. Reaction kinetics of ozone gas in paddy grains. J. Food Process Eng. 2015, 38, 594–600. [Google Scholar] [CrossRef]
- Subramanyam, B.; Xinyi, E.; Savoldelli, S.; Sehgal, B. Efficacy of ozone against Rhyzopertha dominica adults in wheat. J. Stored Prod. Res. 2017, 70, 53–59. [Google Scholar] [CrossRef]
- Zhu, F. Effect of ozone treatment on the quality of grain products. Food Chem. 2018, 264, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Dodd, J.G.; Vegi, A.; Vashisht, A.; Tobias, D.; Schwarz, P.; Wolf-Hall, C.E. Effect of ozone treatment on the safety and quality of malting barley. J. Food Prot. 2011, 74, 2134–2141. [Google Scholar] [CrossRef]
- Tiwari, B.; Brennan, C.S.; Curran, T.; Gallagher, E.; Cullen, P.; O’Donnell, C. Application of ozone in grain processing. J. Cereal Sci. 2010, 51, 248–255. [Google Scholar] [CrossRef]
- Jian, F.; Jayas, D.S.; White, N.D. Can ozone be a new control strategy for pests of stored grain? Agric. Res. 2013, 2, 1–8. [Google Scholar] [CrossRef]
- Matsue, Y.; Uchimura, Y.; Sato, H. Confirmation of the productivity of the stored seeds of wheat and two-rowed barley. Plant Prod. Sci. 2002, 5, 187–193. [Google Scholar] [CrossRef] [Green Version]
- ISTA. International Rules for Seed Testing; International Seed Testing Association: Basserdorf, Switzerland, 2006. [Google Scholar]
- Islam, A.; Anuar, N.; Yaakob, Z. Effect of genotypes and pre-sowing treatments on seed germination behavior of Jatropha. Asian J. Plant Sci. 2009, 8, 433. [Google Scholar] [CrossRef]
- Pasynkov, A.; Pasynkova, E. Comparative evaluation of different methods to determine the crude gluten content in wheat grain. Russ. Agric. Sci. 2021, 47, 238–243. [Google Scholar] [CrossRef]
- Pojić, M.; Mastilović, J.; Majcen, N. The application of near infrared spectroscopy in wheat quality control. In Infrared Spectroscopy–Life and Biomedical Sciences; Theophanides, T., Ed.; Intech Publisher: Rijeka, Croatia, 2012; pp. 167–184. [Google Scholar]
- Subramanyam, B.; Li, B. Efficacy of ozone against phosphine susceptible and resistant strains of four stored-product insect species. Insects 2017, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Holmstrup, M.; Sørensen, J.G.; Heckmann, L.-H.; Slotsbo, S.; Hansen, P.; Hansen, L.S. Effects of ozone on gene expression and lipid peroxidation in adults and larvae of the red flour beetle (Tribolium castaneum). J. Stored Prod. Res. 2011, 47, 378–384. [Google Scholar] [CrossRef]
- Boopathy, B.; Rajan, A.; Radhakrishnan, M. Ozone: An Alternative Fumigant in Controlling the Stored Product Insects and Pests: A Status Report. Ozone Sci. Eng. 2022, 44, 79–95. [Google Scholar] [CrossRef]
- Ballinger, C.A.; Cueto, R.; Squadrito, G.; Coffin, J.F.; Velsor, L.W.; Pryor, W.A.; Postlethwait, E.M. Antioxidant-mediated augmentation of ozone-induced membrane oxidation. Free Radic. Biol. Med. 2005, 38, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Xinyi, E.; Li, B.; Subramanyam, B. Efficacy of ozone against adults and immature stages of phosphine susceptible and resistant strains of Rhyzopertha dominica. J. Stored Prod. Res. 2019, 83, 110–116. [Google Scholar]
- Pimentel, M.A.G.; Faroni, L.R.D.A.; Tótola, M.R.; Guedes, R.N.C. Phosphine resistance, respiration rate and fitness consequences in stored-product insects. Pest Manag. Sci. Former. Pestic. Sci. 2007, 63, 876–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emekci, M.; Navarro, S.; Donahaye, E.; Rindner, M.; Azrieli, A. Respiration of Rhyzopertha dominica (F.) at reduced oxygen concentrations. J. Stored Prod. Res. 2004, 40, 27–38. [Google Scholar] [CrossRef]
- Wu, J.; Doan, H.; Cuenca, M.A. Investigation of gaseous ozone as an anti-fungal fumigant for stored wheat. J. Chem. Technol. Biotechnol. Int. Res. Process Environ. Clean Technol. 2006, 81, 1288–1293. [Google Scholar] [CrossRef]
- Shrestha, R.K.; Lindsey, L.E. Agronomic management of malting barley and research needs to meet demand by the craft brew industry. Agron. J. 2019, 111, 1570–1580. [Google Scholar] [CrossRef] [Green Version]
- Monroy Vazquez, M.E.; Peña-Valdivia, C.B.; García, J.R.; Solano, E.; Campos, H.; García, E. Chemical scarification and ozone in seed dormancy alleviation of wild and domesticated Opuntia, Cactaceae. Ozone Sci. Eng. 2017, 39, 104–114. [Google Scholar] [CrossRef]
- Wang, H.; Qi, Q.; Schorr, P.; Cutler, A.J.; Crosby, W.L.; Fowke, L.C. ICK1, a cyclin-dependent protein kinase inhibitor from Arabidopsis thaliana interacts with both Cdc2a and CycD3, and its expression is induced by abscisic acid. Plant J. 1998, 15, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Normov, D.; Chesniuk, E.; Shevchenko, A.; Normova, T.; Goldman, R.; Pozhidaev, D.; Bohinc, T.; Trdan, S. Does ozone treatment of maize seeds influence their germination and growth energy? Acta Agric. Slov. 2019, 114, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Gapper, C.; Dolan, L. Control of plant development by reactive oxygen species. Plant Physiol. 2006, 141, 341–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bláha, L. Possible Experimental Mistakes in Agricultural Research. Agric. Sci. 2019, 10, 1423–1437. [Google Scholar] [CrossRef] [Green Version]
- Alexander, L.; Yuri, S.; Mikhail, P.; Olga, S.; Sergey, K.; Irina, L. Treatment of spring wheat seeds by ozone generated from humid air and dry oxygen. Res. Agric. Eng. 2018, 64, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Bonifas, K.D.; Walters, D.T.; Cassman, K.G.; Lindquist, J.L. Nitrogen supply affects root: Shoot ratio in corn and velvetleaf (Abutilon theophrasti). Weed Sci. 2005, 53, 670–675. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Shao, H.; Luo, X.; Wang, R.; Li, Y.; Li, Y.; Luo, Y.; Chen, Z. Effect of ozone treatment on deoxynivalenol and wheat quality. PLoS ONE 2016, 11, e0147613. [Google Scholar] [CrossRef] [PubMed]
- Sui, Z.; Yao, T.; Zhong, J.; Li, Y.; Kong, X.; Ai, L. Ozonation treatment improves properties of wheat flour and the baking quality of cake. Philipp. Agric. Sci. 2016, 99, 50–57. [Google Scholar]
- Lozano-Isla, F.; Campos, M.L.; Endres, L.; Bezerra-Neto, E.; Pompelli, M.F. Effects of seed storage time and salt stress on the germination of Jatropha curcas L. Ind. Crops Prod. 2018, 118, 214–224. [Google Scholar] [CrossRef]
- Azevedo Neto, A.D.; Prisco, J.T.; Gomes-Filho, E. Changes in soluble amino-N, soluble proteins and free amino acids in leaves and roots of salt-stressed maize genotypes. J. Plant Interact. 2009, 4, 137–144. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The mortality rate of (a) Rhyzopertha dominica and (b) Tribolium castaneum to different O3 exposure times at 0, 1, 2, and 7 days after O3 treatment. The bars with different letters (a–g) are significantly different (Tukey’s HSD test, p < 0.05).
Figure 1.
The mortality rate of (a) Rhyzopertha dominica and (b) Tribolium castaneum to different O3 exposure times at 0, 1, 2, and 7 days after O3 treatment. The bars with different letters (a–g) are significantly different (Tukey’s HSD test, p < 0.05).

Figure 2.
Changes in the morphological characteristics with O3 treatment duration on (a) barley root length; (b) barley shoot length; (c) seedling vigour index of barley seeds; (d) the ratio of root-to-shoot length. Significant differences (p < 0.05) between each treatment are indicated by letters. Groups do not have significant differences if they have identical marked letters. Statistical reliability of the differences was determined based on Tukey’s HSD test.
Figure 2.
Changes in the morphological characteristics with O3 treatment duration on (a) barley root length; (b) barley shoot length; (c) seedling vigour index of barley seeds; (d) the ratio of root-to-shoot length. Significant differences (p < 0.05) between each treatment are indicated by letters. Groups do not have significant differences if they have identical marked letters. Statistical reliability of the differences was determined based on Tukey’s HSD test.

Figure 3.
Effect of O3 duration on the quality of barley seeds. (a) The protein content; (b) moisture content; (c) colour; (d) hardness; (e) thousand seeds weight. There is no significant differences between each ozonetreatments based on Tukey’s HSD test.
Figure 3.
Effect of O3 duration on the quality of barley seeds. (a) The protein content; (b) moisture content; (c) colour; (d) hardness; (e) thousand seeds weight. There is no significant differences between each ozonetreatments based on Tukey’s HSD test.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Regression analysis for mortality rate of Rhyzopertha dominica and Tribolium castaneum at different assessment intervals. 700 ppm ozone of each treatment time (0, 40, 120, 240, 480, 960, and 1440 min) for two species were conducted.
Table 1.
Regression analysis for mortality rate of Rhyzopertha dominica and Tribolium castaneum at different assessment intervals. 700 ppm ozone of each treatment time (0, 40, 120, 240, 480, 960, and 1440 min) for two species were conducted.
| Insects | DAT † | Slope ‡ ± SE | Intercept ± SE | Relative Median Potency * | LT50 (95% CI §) (min) | LT95 (95% CI) (min) | χ2 ¶ | df |
|---|---|---|---|---|---|---|---|---|
| R. dominica | 0 | 2.32 ± 0.05 | −6.21 ± 0.15 | 1.00 | 472.91 (343.04–653.55) | 2419.93 (1465.95–6336.68) | 118.02 | 6 |
| 1 | 2.48 ± 0.06 | −6.48 ± 0.15 | 1.15 | 410.12 (278.81–587.64) | 1887.41 (1130.49–5594.71) | 162.50 | 6 | |
| 2 | 2.74 ± 0.06 | −7.01 ± 0.17 | 1.25 | 378.37 (265.87–515.29) | 1510.96 (976.73–3617.63) | 138.20 | 6 | |
| 7 | 2.06 ± 0.05 | −4.80 ± 0.12 | 2.22 | 213.50 (146.17–291.40) | 1342.06 (843.67–3033.99) | 95.58 | 6 | |
| T. castaneum | 0 | 2.72 ± 0.07 | −7.23 ± 0.20 | 1.00 | 453.07 (228.14–769.72) | 1823.12 (972.31–27304.4) | 322.13 | 6 |
| 1 | 2.31 ± 0.06 | −5.54 ± 0.14 | 1.80 | 252.04 (143.69–378.75) | 1300.80 (747.48–4884.78) | 198.01 | 6 | |
| 2 | 2.85 ± 0.07 | −5.88 ± 0.16 | 3.91 | 115.79 (92.92–139.38) | 437.12 (341.45–620.62) | 40.70 | 6 | |
| 7 | 2.30 ± 0.07 | −4.15± 0.14 | 7.12 | 63.60 (49.06–78.09) | 329.40 (256.77–463.99) | 27.22 | 6 |
† Day after O3 treatment; ‡ Mean ± standard error; * Relative potency = LC 50 of 0 DAT/LC50 of another DAT (1, 2, 7); § Confidence interval; ¶ Chi-square value for goodness-of-fit of probit model to data.
Table 2.
Efficacy of O3 duration at 700 ppm on all life stages of Rhyzopertha dominica and Tribolium castaneum.
Table 2.
Efficacy of O3 duration at 700 ppm on all life stages of Rhyzopertha dominica and Tribolium castaneum.
| Life Stage | O3 Treatment Time (min) | Rhyzopertha dominica | Tribolium castaneum |
|---|---|---|---|
| Mortality Rate (%) † | Mortality Rate (%) | ||
| Egg | 0 | 0.00 ± 0.00 g | 1.67 ± 1.67 g |
| 40 | 18.33 ± 3.33 fg | 17.92 ± 1.5 fg | |
| 120 | 38.33 ± 3.33 ef | 33.33 ± 3.85 ef | |
| 240 | 68.33 ± 4.41 bcd | 49.01 ± 5.21 de | |
| 480 | 70.63 ± 5.23 bc | 61.52 ± 9.34 cd | |
| 960 | 83.33 ± 6.01 ab | 81.11 ± 2.22 abc | |
| 1440 | 100.00 ± 0.00 a | 100.00 ± 0.00 a | |
| Larvae | 0 | 3.33 ± 1.92 g | 3.70 ± 0.93 g |
| 40 | 26.67 ± 3.33 f | 25.56 ± 2.94 f | |
| 120 | 53.33 ± 6.67 e | 51.38 ± 2.01 e | |
| 240 | 60.42 ± 5.51 de | 82.88 ± 1.93 bc | |
| 480 | 70.83 ± 2.08 cd | 99.67 ± 0.33 a | |
| 960 | 91.67 ± 2.08 ab | 100.00 ± 0.00 a | |
| 1440 | 100.00 ± 0.00 a | 100.00 ± 0.00 a | |
| Pupae | 0 | 0.00± 0.00 g | 2.78 ± 1.60 g |
| 40 | 12.95 ± 2.50 fg | 24.39 ± 3.43 ef | |
| 120 | 28.62 ± 3.10 de | 51.82 ± 4.30 cd | |
| 240 | 44.21 ± 4.89 c | 64.55 ± 2.92 b | |
| 480 | 63.82 ± 4.20 b | 74.24 ± 2.98 b | |
| 960 | 74.30 ± 1.86 b | 93.33 ± 3.33 a | |
| 1440 | 100.00 ± 0.00 a | 100.00 ± 0.00 a | |
| Adults | 0 | 2.06 ± 0.32 h | 3.21 ± 0.61 gh |
| 40 | 12.85 ± 0.58 g | 38.00 ± 8.09 e | |
| 120 | 27.29 ± 0.77 f | 68.97 ± 6.90 c | |
| 240 | 56.57 ± 2.74 d | 85.24 ± 3.51 b | |
| 480 | 71.95 ± 2.81 c | 98.45 ± 0.29 a | |
| 960 | 89.62 ± 1.06 ab | 100.00 ± 0.00 a | |
| 1440 | 100 ± 0.00 a | 100 ± 0.00 a |
† Mean ± standard error. Different letters in each life stage indicate significant mortality differences at ozone treatment times between the two species (p < 0.05, Tukey’s HSD test).
Table 3.
Effect of O3 duration at 700 ppm on the germination rate of old harvest and newly harvest barley seeds.
Table 3.
Effect of O3 duration at 700 ppm on the germination rate of old harvest and newly harvest barley seeds.
| O3 Treatment Duration (min) | Germination Rate (%) | |
|---|---|---|
| Old (2013/2014) † | New (2019/2020) | |
| 0 | 94.34 ± 1.67 ab | 98.35 ± 0.56 a |
| 10 | 94.11 ± 0.97 ab | 99.06 ± 0.36 a |
| 20 | 95.28 ± 0.87 ab | 99.06 ± 0.36 a |
| 40 | 97.17 ± 1.01 a | 98.59 ± 0.59 a |
| 120 | 95.28 ± 1.01 ab | 98.82 ± 0.79 a |
| 240 | 84.21 ± 1.12 bc | 96.23 ± 0.71 a |
| 480 | 86.32 ± 0.93 c | 94.34 ± 0.94 a |
| 960 | 68.16 ± 1.61 d | 78.54 ± 2.71 b |
| 1440 | 14.39 ± 4.55 e | 20.28 ± 1.84 c |
† Mean ± standard error. Means in the same column with different letters (a–e) are significantly different at p < 0.05 following Tukey’s HSD test.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dong, X.; Agarwal, M.; Xiao, Y.; Ren, Y.; Maker, G.; Yu, X. Ozone Efficiency on Two Coleopteran Insect Pests and Its Effect on Quality and Germination of Barley. Insects 2022, 13, 318. https://doi.org/10.3390/insects13040318
AMA Style
Dong X, Agarwal M, Xiao Y, Ren Y, Maker G, Yu X. Ozone Efficiency on Two Coleopteran Insect Pests and Its Effect on Quality and Germination of Barley. Insects. 2022; 13(4):318. https://doi.org/10.3390/insects13040318
Chicago/Turabian StyleDong, Xue, Manjree Agarwal, Yu Xiao, Yonglin Ren, Garth Maker, and Xiangyang Yu. 2022. "Ozone Efficiency on Two Coleopteran Insect Pests and Its Effect on Quality and Germination of Barley" Insects 13, no. 4: 318. https://doi.org/10.3390/insects13040318
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.