
Role of microRNAs in Congenital Diaphragmatic Hernia-Associated Pulmonary Hypertension
 , , ,
, , ,
Abstract
:1. Introduction
2. Role of miRNAs in CDH Related Pulmonary Hypertension
2.1. Animal CDH Models
2.2. Human CDH Studies
3. Discussion and Future Perspective
3.1. Understanding the Etiopathogenesis of CDH-Related PH
3.2. Contribution of miRNA in CDH Related PH
3.3. miRNAs as Diagnostic and Prognostic Biomarkers
3.4. miRNAs as Therapeutic Targets and Tools
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Paoletti, M.; Raffler, G.; Gaffi, M.S.; Antounians, L.; Lauriti, G.; Zani, A. Prevalence and Risk Factors for Congenital Diaphragmatic Hernia: A Global View. J. Pediatr. Surg. 2020, 55, 2297–2307. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.S.; Harting, M.T.; Lally, P.A.; Miller, C.C.; Hirschl, R.B.; Davis, C.F.; Dassinger, M.S.; Buchmiller, T.L.; Van Meurs, K.P.; Yoder, B.A.; et al. Mortality in Congenital Diaphragmatic Hernia: A Multicenter Registry Study of Over 5000 Patients Over 25 Years. Ann. Surg. 2021, 277, 520–527. [Google Scholar] [CrossRef]
- Putnam, L.R.; Harting, M.T.; Tsao, K.; Morini, F.; Yoder, B.A.; Luco, M.; Lally, P.A.; Lally, K.P.; on behalf of the Congenital Diaphragmatic Hernia Study Group. Congenital Diaphragmatic Hernia Defect Size and Infant Morbidity at Discharge. Pediatrics 2016, 138, e20162043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, V.S.; Harting, M.T. Congenital Diaphragmatic Hernia-Associated Pulmonary Hypertension. Semin. Perinatol. 2020, 44, 151167. [Google Scholar] [CrossRef]
- Kitagawa, M.; Hislop, A.; Boyden, E.A.; Reid, L. Lung Hypoplasia in Congenital Diaphragmatic Hernia a Quantitative Study of Airway, Artery, and Alveolar Development. Br. J. Surg. 1971, 58, 342–346. [Google Scholar] [CrossRef]
- Levin, D.L. Morphologic Analysis of the Pulmonary Vascular Bed in Congenital Left-Sided Diaphragmatic Hernia. J. Pediatr. 1978, 92, 805–809. [Google Scholar] [CrossRef]
- Hedrick, H.L.; Danzer, E.; Merchant, A.; Bebbington, M.W.; Zhao, H.; Flake, A.W.; Johnson, M.P.; Liechty, K.W.; Howell, L.J.; Wilson, R.D.; et al. Liver Position and Lung-to-Head Ratio for Prediction of Extracorporeal Membrane Oxygenation and Survival in Isolated Left Congenital Diaphragmatic Hernia. Am. J. Obstet. Gynecol. 2007, 197, 422.e1–422.e4. [Google Scholar] [CrossRef]
- Jani, J.; Nicolaides, K.H.; Keller, R.L.; Benachi, A.; Peralta, C.F.A.; Favre, R.; Moreno, O.; Tibboel, D.; Lipitz, S.; Eggink, A.; et al. Observed to Expected Lung Area to Head Circumference Ratio in the Prediction of Survival in Fetuses with Isolated Diaphragmatic Hernia. Ultrasound Obstet. Gynecol. 2007, 30, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Jani, J.C.; Benachi, A.; Nicolaides, K.H.; Allegaert, K.; Gratacós, E.; Mazkereth, R.; Matis, J.; Tibboel, D.; Van Heijst, A.; Storme, L.; et al. Prenatal Prediction of Neonatal Morbidity in Survivors with Congenital Diaphragmatic Hernia: A Multicenter Study. Ultrasound Obstet. Gynecol. 2009, 33, 64–69. [Google Scholar] [CrossRef]
- Kipfmueller, F.; Akkas, S.; Pugnaloni, F.; Bo, B.; Lemloh, L.; Schröder, L.; Gembruch, U.; Geipel, A.; Berg, C.; Heydweiller, A.; et al. Echocardiographic Assessment of Pulmonary Hypertension in Neonates with Congenital Diaphragmatic Hernia Using Pulmonary Artery Flow Characteristics. J. Clin. Med. 2022, 11, 3038. [Google Scholar] [CrossRef]
- DeKoninck, P.; Gratacos, E.; Van Mieghem, T.; Richter, J.; Lewi, P.; Ancel, A.M.; Allegaert, K.; Nicolaides, K.; Deprest, J. Results of Fetal Endoscopic Tracheal Occlusion for Congenital Diaphragmatic Hernia and the Set up of the Randomized Controlled TOTAL Trial. Early Hum. Dev. 2011, 87, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Deprest, J.A.; Nicolaides, K.H.; Benachi, A.; Gratacos, E.; Ryan, G.; Persico, N.; Sago, H.; Johnson, A.; Wielgoś, M.; Berg, C.; et al. Randomized Trial of Fetal Surgery for Severe Left Diaphragmatic Hernia. N. Engl. J. Med. 2021, 385, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Deprest, J.A.; Benachi, A.; Gratacos, E.; Nicolaides, K.H.; Berg, C.; Persico, N.; Belfort, M.; Gardener, G.J.; Ville, Y.; Johnson, A.; et al. Randomized Trial of Fetal Surgery for Moderate Left Diaphragmatic Hernia. N. Engl. J. Med. 2021, 385, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Croce, C.M. Causes and Consequences of MicroRNA Dysregulation in Cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Banerjee, S.; de Freitas, A.; Sanders, Y.Y.; Ding, Q.; Matalon, S.; Thannickal, V.J.; Abraham, E.; Liu, G. Participation of MiR-200 in Pulmonary Fibrosis. Am. J. Pathol. 2012, 180, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Khoshgoo, N.; Visser, R.; Falk, L.; Day, C.A.; Ameis, D.; Iwasiow, B.M.; Zhu, F.; Öztürk, A.; Basu, S.; Pind, M.; et al. MicroRNA-200b Regulates Distal Airway Development by Maintaining Epithelial Integrity. Sci. Rep. 2017, 7, 6382. [Google Scholar] [CrossRef] [Green Version]
- Eastwood, M.P.; Deprest, J.; Russo, F.M.; Wang, H.; Mulhall, D.; Iwasiow, B.; Mahood, T.H.; Keijzer, R. MicroRNA 200b Is Upregulated in the Lungs of Fetal Rabbits with Surgically Induced Diaphragmatic Hernia. Prenat. Diagn. 2018, 38, 645–653. [Google Scholar] [CrossRef]
- Zhu, S.; He, Q.; Zhang, R.; Wang, Y.; Zhong, W.; Xia, H.; Yu, J. Decreased Expression of MiR-33 in Fetal Lungs of Nitrofen-Induced Congenital Diaphragmatic Hernia Rat Model. J. Pediatr. Surg. 2016, 51, 1096–1100. [Google Scholar] [CrossRef]
- Khoshgoo, N.; Kholdebarin, R.; Pereira-Terra, P.; Mahood, T.H.; Falk, L.; Day, C.A.; Iwasiow, B.M.; Zhu, F.; Mulhall, D.; Fraser, C.; et al. Prenatal MicroRNA MiR-200b Therapy Improves Nitrofen-Induced Pulmonary Hypoplasia Associated With Congenital Diaphragmatic Hernia. Ann. Surg. 2019, 269, 979–987. [Google Scholar] [CrossRef]
- Fabietti, I.; Nardi, T.; Favero, C.; Dioni, L.; Cantone, L.; Pergoli, L.; Hoxha, M.; Pinatel, E.; Mosca, F.; Bollati, V.; et al. Extracellular Vesicles and Their MiRNA Content in Amniotic and Tracheal Fluids of Fetuses with Severe Congenital Diaphragmatic Hernia Undergoing Fetal Intervention. Cells 2021, 10, 1493. [Google Scholar] [CrossRef]
- Pereira-Terra, P.; Deprest, J.A.; Kholdebarin, R.; Khoshgoo, N.; DeKoninck, P.; Munck, A.A.B.-D.; Wang, J.; Zhu, F.; Rottier, R.J.; Iwasiow, B.M.; et al. Unique Tracheal Fluid MicroRNA Signature Predicts Response to FETO in Patients With Congenital Diaphragmatic Hernia. Ann. Surg. 2015, 262, 1130. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Rivero, M.; Zhang, R.; Heilmann-Heimbach, S.; Mueller, A.; Bagci, S.; Dresbach, T.; Schröder, L.; Holdenrieder, S.; Reutter, H.M.; Kipfmueller, F. Circulating MicroRNAs Are Associated with Pulmonary Hypertension and Development of Chronic Lung Disease in Congenital Diaphragmatic Hernia. Sci. Rep. 2018, 8, 10735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piersigilli, F.; Syed, M.; Lam, T.T.; Dotta, A.; Massoud, M.; Vernocchi, P.; Quagliariello, A.; Putignani, L.; Auriti, C.; Salvatori, G.; et al. An Omic Approach to Congenital Diaphragmatic Hernia: A Pilot Study of Genomic, MicroRNA, and Metabolomic Profiling. J. Perinatol. 2020, 40, 952–961. [Google Scholar] [CrossRef]
- Costlow, R.D.; Manson, J.M. The Heart and Diaphragm: Target Organs in the Neonatal Death Induced by Nitrofen (2,4-Dichlorophenyl-p-Nitrophenyl Ether). Toxicology 1981, 20, 209–227. [Google Scholar] [CrossRef] [PubMed]
- Belik, J.; Davidge, S.T.; Zhang, W.; Pan, J.; Greer, J.J. Airway Smooth Muscle Changes in the Nitrofen-Induced Congenital Diaphragmatic Hernia Rat Model. Pediatr. Res. 2003, 53, 737–743. [Google Scholar] [CrossRef] [Green Version]
- Keijzer, R.; Liu, J.; Deimling, J.; Tibboel, D.; Post, M. Dual-Hit Hypothesis Explains Pulmonary Hypoplasia in the Nitrofen Model of Congenital Diaphragmatic Hernia. Am. J. Pathol. 2000, 156, 1299–1306. [Google Scholar] [CrossRef] [Green Version]
- Iritani, I. Experimental Study on Embryogenesis of Congenital Diaphragmatic Hernia. Anat. Embryol. (Berl.) 1984, 169, 133–139. [Google Scholar] [CrossRef]
- Kluth, D.; Tenbrinck, R.; von Ekesparre, M.; Kangah, R.; Reich, P.; Brandsma, A.; Tibboel, D.; Lambrecht, W. The Natural History of Congenital Diaphragmatic Hernia and Pulmonary Hypoplasia in the Embryo. J. Pediatr. Surg. 1993, 28, 456–462, discussion 462–463. [Google Scholar] [CrossRef] [PubMed]
- van Loenhout, R.B.; Tibboel, D.; Post, M.; Keijzer, R. Congenital Diaphragmatic Hernia: Comparison of Animal Models and Relevance to the Human Situation. Neonatology 2009, 96, 137–149. [Google Scholar] [CrossRef]
- Mendelsohn, C.; Lohnes, D.; Decimo, D.; Lufkin, T.; LeMeur, M.; Chambon, P.; Mark, M. Function of the Retinoic Acid Receptors (RARs) during Development (II). Multiple Abnormalities at Various Stages of Organogenesis in RAR Double Mutants. Development 1994, 120, 2749–2771. [Google Scholar] [CrossRef]
- Noble, B.R.; Babiuk, R.P.; Clugston, R.D.; Underhill, T.M.; Sun, H.; Kawaguchi, R.; Walfish, P.G.; Blomhoff, R.; Gundersen, T.E.; Greer, J.J. Mechanisms of Action of the Congenital Diaphragmatic Hernia-Inducing Teratogen Nitrofen. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, L1079–L1087. [Google Scholar] [CrossRef]
- Howe, E.N.; Cochrane, D.R.; Richer, J.K. The MiR-200 and MiR-221/222 MicroRNA Families: Opposing Effects on Epithelial Identity. J. Mammary Gland Biol. Neoplasia 2012, 17, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Kurashige, J.; Kamohara, H.; Watanabe, M.; Hiyoshi, Y.; Iwatsuki, M.; Tanaka, Y.; Kinoshita, K.; Saito, S.; Baba, Y.; Baba, H. MicroRNA-200b Regulates Cell Proliferation, Invasion, and Migration by Directly Targeting ZEB2 in Gastric Carcinoma. Ann. Surg. Oncol. 2012, 19, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Gorelova, A.; Berman, M.; Al Ghouleh, I. Endothelial-to-Mesenchymal Transition in Pulmonary Arterial Hypertension. Antioxid. Redox. Signal. 2021, 34, 891–914. [Google Scholar] [CrossRef] [PubMed]
- Dingemann, J.; Doi, T.; Ruttenstock, E.; Puri, P. Abnormal Platelet-Derived Growth Factor Signaling Accounting for Lung Hypoplasia in Experimental Congenital Diaphragmatic Hernia. J. Pediatr. Surg. 2010, 45, 1989–1994. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, J.J.; Briot, A.; Enciso, J.; Zovein, A.C.; Ren, S.; Zhang, Z.W.; Radtke, F.; Simons, M.; Wang, Y.; Iruela-Arispe, M.L. Endothelial Deletion of Murine Jag1 Leads to Valve Calcification and Congenital Heart Defects Associated with Alagille Syndrome. Development 2012, 4460, 4449–4460. [Google Scholar] [CrossRef] [Green Version]
- Takayasu, H.; Nakazawa, N.; Montedonico, S.; Puri, P. Down-Regulation of Wnt Signal Pathway in Nitrofen-Induced Hypoplastic Lung. J. Pediatr. Surg. 2007, 42, 426–430. [Google Scholar] [CrossRef]
- Ambalavanan, N.; Bulger, A.; Murphy-Ullrich, J.; Oparil, S.; Chen, Y.-F. Endothelin-A Receptor Blockade Prevents and Partially Reverses Neonatal Hypoxic Pulmonary Vascular Remodeling. Pediatr. Res. 2005, 57, 631–636. [Google Scholar] [CrossRef] [Green Version]
- Keller, R.L.; Tacy, T.A.; Hendricks-Munoz, K.; Xu, J.; Moon-Grady, A.J.; Neuhaus, J.; Moore, P.; Nobuhara, K.K.; Hawgood, S.; Fineman, J.R. Congenital Diaphragmatic Hernia: Endothelin-1, Pulmonary Hypertension, and Disease Severity. Am. J. Respir. Crit. Care Med. 2010, 182, 555–561. [Google Scholar] [CrossRef] [Green Version]
- El Agha, E.; Schwind, F.; Ruppert, C.; Günther, A.; Bellusci, S.; Schermuly, R.T.; Kosanovic, D. Is the Fibroblast Growth Factor Signaling Pathway a Victim of Receptor Tyrosine Kinase Inhibition in Pulmonary Parenchymal and Vascular Remodeling? Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 315, L248–L252. [Google Scholar] [CrossRef]
- Kim, J.; Kang, Y.; Kojima, Y.; Lighthouse, J.K.; Hu, X.; Aldred, M.A.; McLean, D.L.; Park, H.; Comhair, S.A.; Greif, D.M.; et al. Endothelial Apelin-FGF Link Mediated by MicroRNAs 424 and 503 Is Disrupted in Pulmonary Arterial Hypertension. Nat. Med. 2013, 19, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Zhang, X.; Kang, K.; Chen, J.; Wu, Z.; Huang, J.; Lu, W.; Chen, Y.; Zhang, J.; Wang, Z.; et al. MicroRNA-223 Attenuates Hypoxia-Induced Vascular Remodeling by Targeting RhoB/MLC2 in Pulmonary Arterial Smooth Muscle Cells. Sci. Rep. 2016, 6, 24900. [Google Scholar] [CrossRef]
- Fiedler, J.; Thum, T. New Insights into MiR-17–92 Cluster Regulation and Angiogenesis. Circ. Res. 2016, 118, 9–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veenma, D.C.M.; de Klein, A.; Tibboel, D. Developmental and Genetic Aspects of Congenital Diaphragmatic Hernia. Pediatr. Pulmonol. 2012, 47, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Schulz, F.; Jenetzky, E.; Zwink, N.; Bendixen, C.; Kipfmueller, F.; Rafat, N.; Heydweiller, A.; Wessel, L.; Reutter, H.; Mueller, A.; et al. Parental Risk Factors for Congenital Diaphragmatic Hernia—A Large German Case-Control Study. BMC Pediatr. 2021, 21, 278. [Google Scholar] [CrossRef] [PubMed]
- Zani, A.; Chung, W.K.; Deprest, J.; Harting, M.T.; Jancelewicz, T.; Kunisaki, S.M.; Patel, N.; Antounians, L.; Puligandla, P.S.; Keijzer, R. Congenital Diaphragmatic Hernia. Nat. Rev. Dis. Primers. 2022, 8, 1–20. [Google Scholar] [CrossRef] [PubMed]
- van den, H.L.; Reiss, I.; Felix, J.F.; Hop, W.C.J.; Lally, P.A.; Lally, K.P.; Tibboel, D.; for the Congenital Diaphragmatic Hernia Study Group. Risk Factors for Chronic Lung Disease and Mortality in Newborns with Congenital Diaphragmatic Hernia. Neonatology 2010, 98, 370–380. [Google Scholar] [CrossRef]
- Acker, S.N.; Seedorf, G.J.; Abman, S.H.; Nozik-Grayck, E.; Partrick, D.A.; Gien, J. Pulmonary Artery Endothelial Cell Dysfunction and Decreased Populations of Highly Proliferative Endothelial Cells in Experimental Congenital Diaphragmatic Hernia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 305, L943–L952. [Google Scholar] [CrossRef] [Green Version]
- Rhinn, M.; Dollé, P. Retinoic Acid Signalling during Development. Development 2012, 139, 843–858. [Google Scholar] [CrossRef] [Green Version]
- Sood, B.G.; Wykes, S.; Landa, M.; De Jesus, L.; Rabah, R. Expression of ENOS in the Lungs of Neonates with Pulmonary Hypertension. Exp. Mol. Pathol. 2011, 90, 9–12. [Google Scholar] [CrossRef]
- Shehata, S.M.K.; Mooi, W.J.; Okazaki, T.; El-Banna, I.; Sharma, H.S.; Tibboel, D. Enhanced Expression of Vascular Endothelial Growth Factor in Lungs of Newborn Infants with Congenital Diaphragmatic Hernia and Pulmonary Hypertension. Thorax 1999, 54, 427–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benincasa, G.; DeMeo, D.L.; Glass, K.; Silverman, E.K.; Napoli, C. Epigenetics and Pulmonary Diseases in the Horizon of Precision Medicine: A Review. Eur. Respir. J. 2021, 57, 2003406. [Google Scholar] [CrossRef]
- Wagner, R.; Montalva, L.; Zani, A.; Keijzer, R. Basic and Translational Science Advances in Congenital Diaphragmatic Hernia. Semin. Perinatol. 2020, 44, 151170. [Google Scholar] [CrossRef]
- Mulhall, D.; Khoshgoo, N.; Visser, R.; Iwasiow, B.; Day, C.; Zhu, F.; Eastwood, P.; Keijzer, R. MiR-200 Family Expression during Normal and Abnormal Lung Development Due to Congenital Diaphragmatic Hernia at the Later Embryonic Stage in the Nitrofen Rat Model. Pediatr. Surg. Int. 2020, 36, 1429–1436. [Google Scholar] [CrossRef]
- Garcia-Barceló, M.M.; Wong, K.K.Y.; Lui, V.C.H.; Yuan, Z.W.; So, M.T.; Ngan, E.S.W.; Miao, X.P.; Chung, P.H.Y.; Khong, P.L.; Tam, P.K.H. Identification of a HOXD13 Mutation in a VACTERL Patient. Am. J. Med. Genet. Part A 2008, 146, 3181–3185. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hoyle, G.W. Overexpression of PDGF-A in the Lung Epithelium of Transgenic Mice Produces a Lethal Phenotype Associated with Hyperplasia of Mesenchymal Cells. Dev. Biol. 2001, 239, 338–349. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, M.; Möröy, T. The Serine/Threonine Kinase Pim-1. Int. J. Biochem. Cell Biol. 2005, 37, 726–730. [Google Scholar] [CrossRef]
- Keil, T.W.; Baldassi, D.; Merkel, O.M. TH2-Cell Targeted Pulmonary SiRNA Delivery for the Treatment of Asthma. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2020, 12, e1634. [Google Scholar] [CrossRef] [Green Version]
- Asha, K.; Kumar, P.; Sanicas, M.; Meseko, C.A.; Khanna, M.; Kumar, B. Advancements in Nucleic Acid Based Therapeutics against Respiratory Viral Infections. J. Clin. Med. 2019, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christopher Boyd, A.; Guo, S.; Huang, L.; Kerem, B.; Oren, Y.S.; Walker, A.J.; Hart, S.L. New Approaches to Genetic Therapies for Cystic Fibrosis. J. Cyst. Fibros. 2020, 19, S54–S59. [Google Scholar] [CrossRef] [Green Version]
- Naghizadeh, S.; Mohammadi, A.; Baradaran, B.; Mansoori, B. Overcoming Multiple Drug Resistance in Lung Cancer Using SiRNA Targeted Therapy. Gene 2019, 714, 143972. [Google Scholar] [CrossRef]
- Patel, N.; Lally, P.A.; Kipfmueller, F.; Massolo, A.C.; Luco, M.; Van Meurs, K.P.; Lally, K.P.; Harting, M.T. Ventricular Dysfunction Is a Critical Determinant of Mortality in Congenital Diaphragmatic Hernia. Am. J. Respir. Crit. Care Med. 2019, 200, 1522–1530. [Google Scholar] [CrossRef]
- Patel, N.; Massolo, A.C.; Kipfmueller, F. Congenital Diaphragmatic Hernia-Associated Cardiac Dysfunction. Semin. Perinatol. 2020, 44, 151168. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Massolo, A.C.; Kraemer, U.S.; Kipfmueller, F. The Heart in Congenital Diaphragmatic Hernia: Knowns, Unknowns, and Future Priorities. Front. Pediatr. 2022, 10, 890422. [Google Scholar] [CrossRef]
- Patel, N.; Kipfmueller, F. Cardiac Dysfunction in Congenital Diaphragmatic Hernia: Pathophysiology, Clinical Assessment, and Management. Semin. Pediatr. Surg. 2017, 26, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Tian, R. Mitochondrial Dysfunction in Pathophysiology of Heart Failure. J. Clin. Investig. 2018, 128, 3716–3726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhaorigetu, S.; Gupta, V.S.; Jin, D.; Harting, M.T. Cardiac Energy Metabolism May Play a Fundamental Role in Congenital Diaphragmatic Hernia-Associated Ventricular Dysfunction. J. Mol. Cell. Cardiol. 2021, 157, 14–16. [Google Scholar] [CrossRef]


{kind=link}
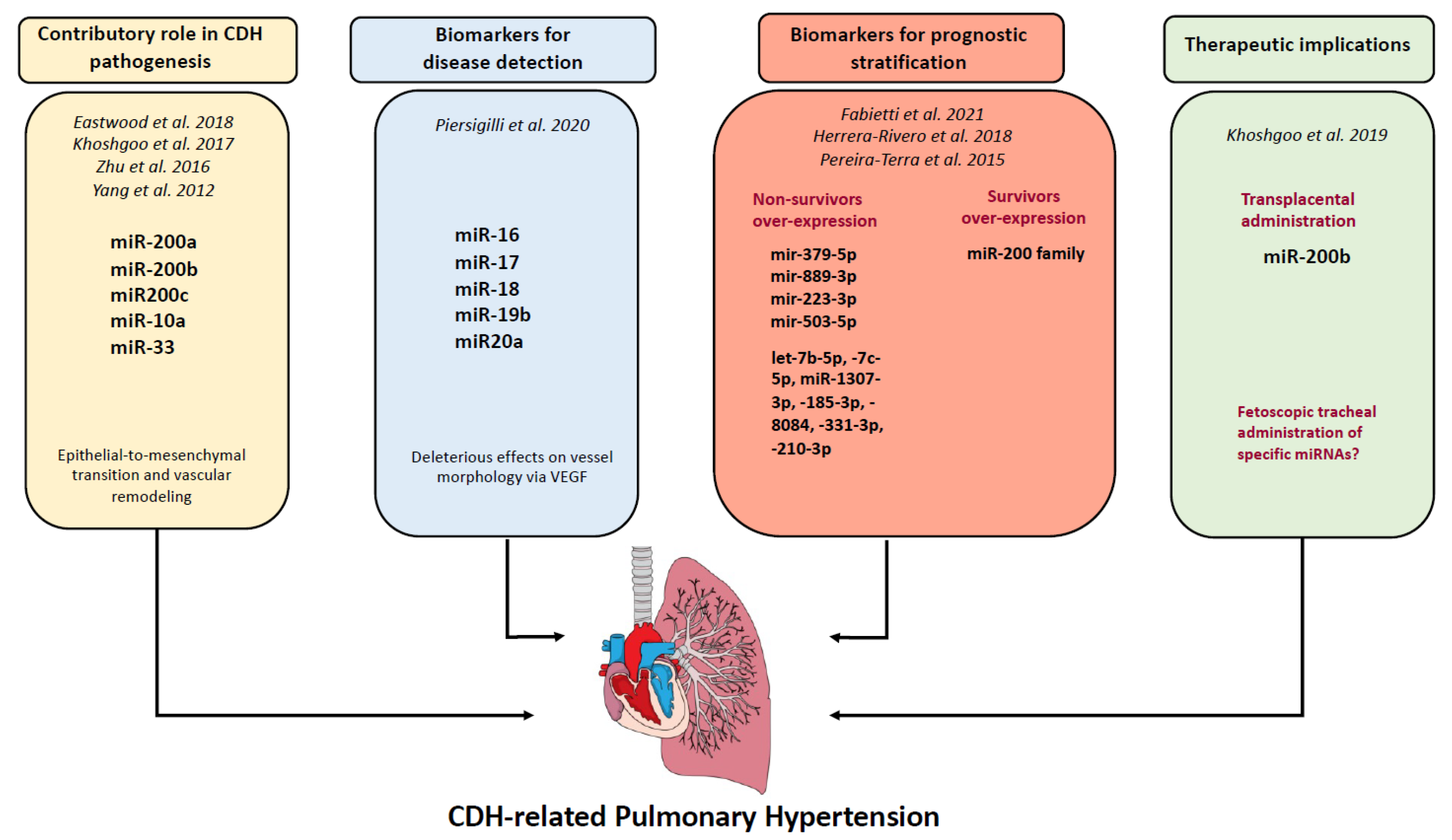
| Study, Year | Material | MiRNAs | Results | Possible Effect/Affected Pathways |
|---|---|---|---|---|
| Animal Models | ||||
| Yang et al., 2012 [15] | Nitrofen rat model | miR-200a; miR-200b; miR200c | Downregulated | Fibrotic lung disease |
| Khoshgoo et al., 2017 [16] | Nitrofen rat model | miR-200b | miR-200b−/− knockout model | Altered airway branching, alveolar wall structure and downregulation of epithelial cell differentiation, fibrotic lung |
| Eastwood et al., 2018 [17] | Rabbit model | miR-200b-3p | Overexpressed | Overexpression in hypoplastic lung |
| Zhu et al., 2016 [18] | Nitrofen rat model | miR-33 | Decreased expression | Decreased expression in CDH lungs, altered lung epithelial–mesenchymal interaction |
| Khoshgoo et al., 2019 [19] | Nitrofen rat model | miR-200b | Compensatory upregulation of miR-200b in less hypoplastic lungs | Transplacental miR-200b therapy corrects abnormal branching morphogenesis |
| Study, Year | Material | MiRNAs | Results | Possible Effect/Affected Pathways |
|---|---|---|---|---|
| Human studies | ||||
| Fabietti et al., 2021 [20] | Amniotic and tracheal fluids of fetuses with CDH undergoing FETO | mir-379-5p; mir-889-3p | Overexpression in pre-FETO amniotic fluid samples of non-survivors | Vascular remodeling, epithelial cell protection and renewal |
| Fabietti et al., 2021 [20] | Amniotic and tracheal fluids of fetuses with CDH undergoing FETO | mir-223-3p; mir-503-5p | Overexpression in tracheal fluid of post-FETO non-survivors | Pulmonary smooth muscle cell proliferation and migration |
| Pereira-Terra et al., 2015 [21] | Amniotic and tracheal fluids of fetuses with CDH undergoing FETO | miR200b; miR-10a | Upregulation in fetal lungs of CDH. Increased expression of miR-200 in FETO responders | Decreased TGF-β/SMAD signaling in human bronchial epithelial cells |
| Herrera-Rivero et al., 2018 [22] | Blood of CDH newborns, collected 24 h after birth | let-7b-5p, -7c-5p, miR-1307-3p, -185-3p, -8084, -331-3p, -210-3p | Differentially expressed in CDH non survivor or developing chronic lung disease | Cell cycle regulation, inflammation processes and tissue morphogenesis |
| Piersigilli et al., 2020 [23] | Tracheal aspirates of CDH newborn | miR-16, miR-17, miR-18, miR-19b, miR20a | Differentially expressed in CDH patients compared with healthy controls | (TGF)-β signaling pathway, early lung branching, morphogenesis and alveologenesis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pugnaloni, F.; Capolupo, I.; Patel, N.; Giliberti, P.; Dotta, A.; Bagolan, P.; Kipfmueller, F. Role of microRNAs in Congenital Diaphragmatic Hernia-Associated Pulmonary Hypertension. Int. J. Mol. Sci. 2023, 24, 6656. https://doi.org/10.3390/ijms24076656
Pugnaloni F, Capolupo I, Patel N, Giliberti P, Dotta A, Bagolan P, Kipfmueller F. Role of microRNAs in Congenital Diaphragmatic Hernia-Associated Pulmonary Hypertension. International Journal of Molecular Sciences. 2023; 24(7):6656. https://doi.org/10.3390/ijms24076656
Chicago/Turabian StylePugnaloni, Flaminia, Irma Capolupo, Neil Patel, Paola Giliberti, Andrea Dotta, Pietro Bagolan, and Florian Kipfmueller. 2023. "Role of microRNAs in Congenital Diaphragmatic Hernia-Associated Pulmonary Hypertension" International Journal of Molecular Sciences 24, no. 7: 6656. https://doi.org/10.3390/ijms24076656