
Curcumin Ameliorates Particulate Matter-Induced Pulmonary Injury through Bimodal Regulation of Macrophage Inflammation via NF-κB and Nrf2

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
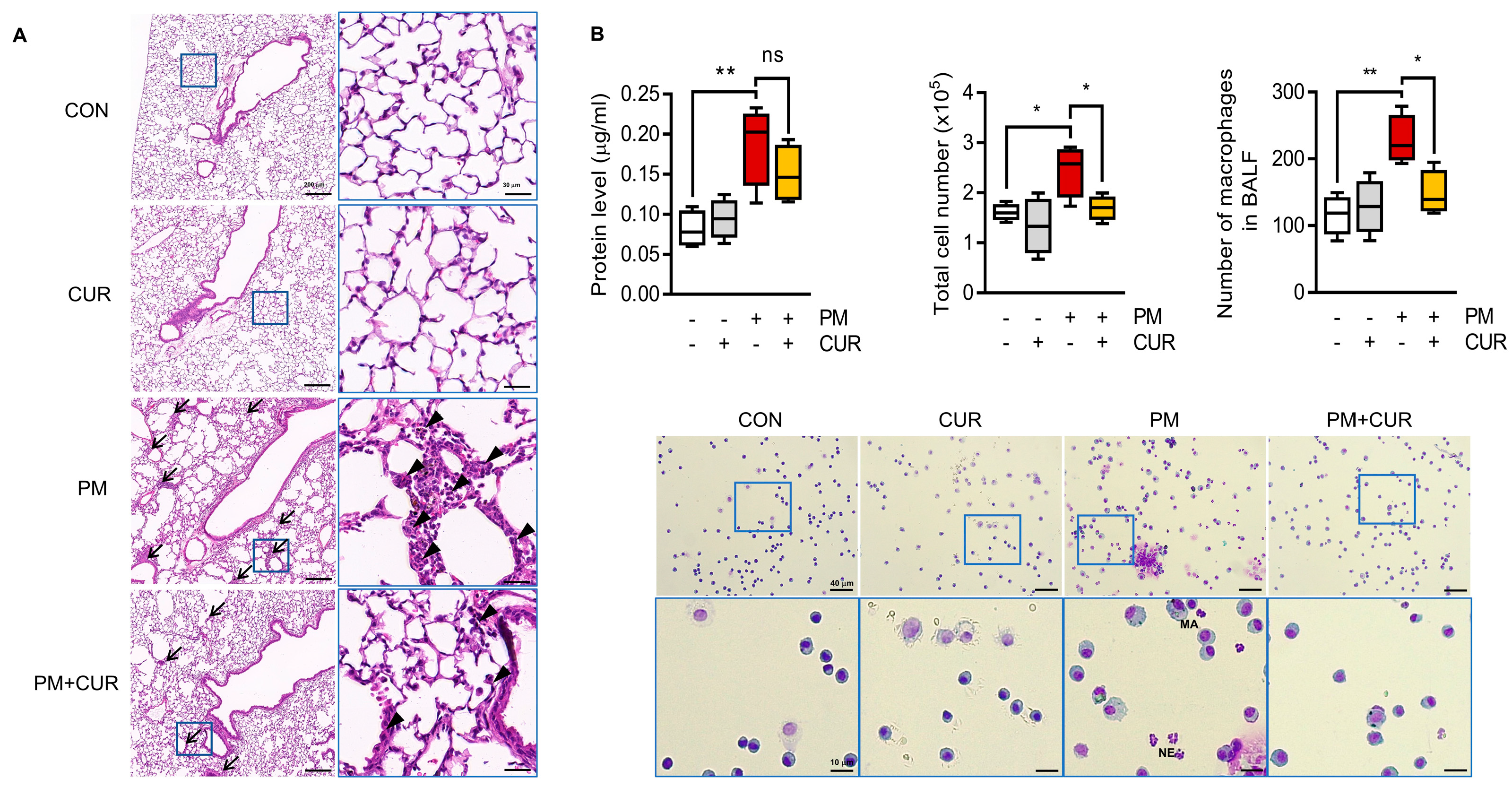
2.1. Curcumin Alleviated Histological Changes and Immune Cell Infiltration in PM-Treated Mouse Lung Tissue and Bronchoalveolar Lavage Fluid (BALF)
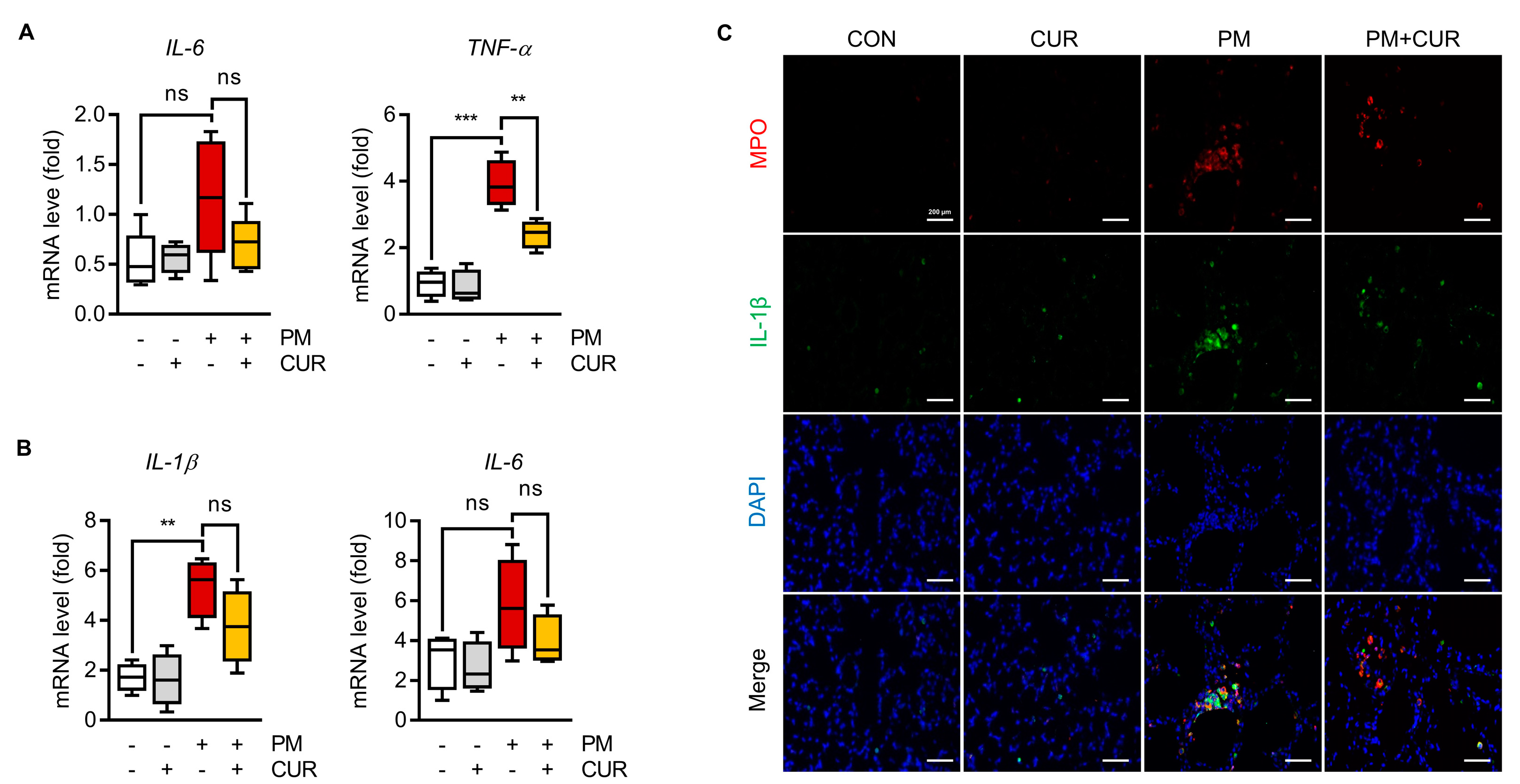
2.2. Curcumin Downregulated the Expression of IL-1β, IL-6, and TNF-α in the Lung Tissue and BALF of Mice with PM-Induced Lung Inflammation
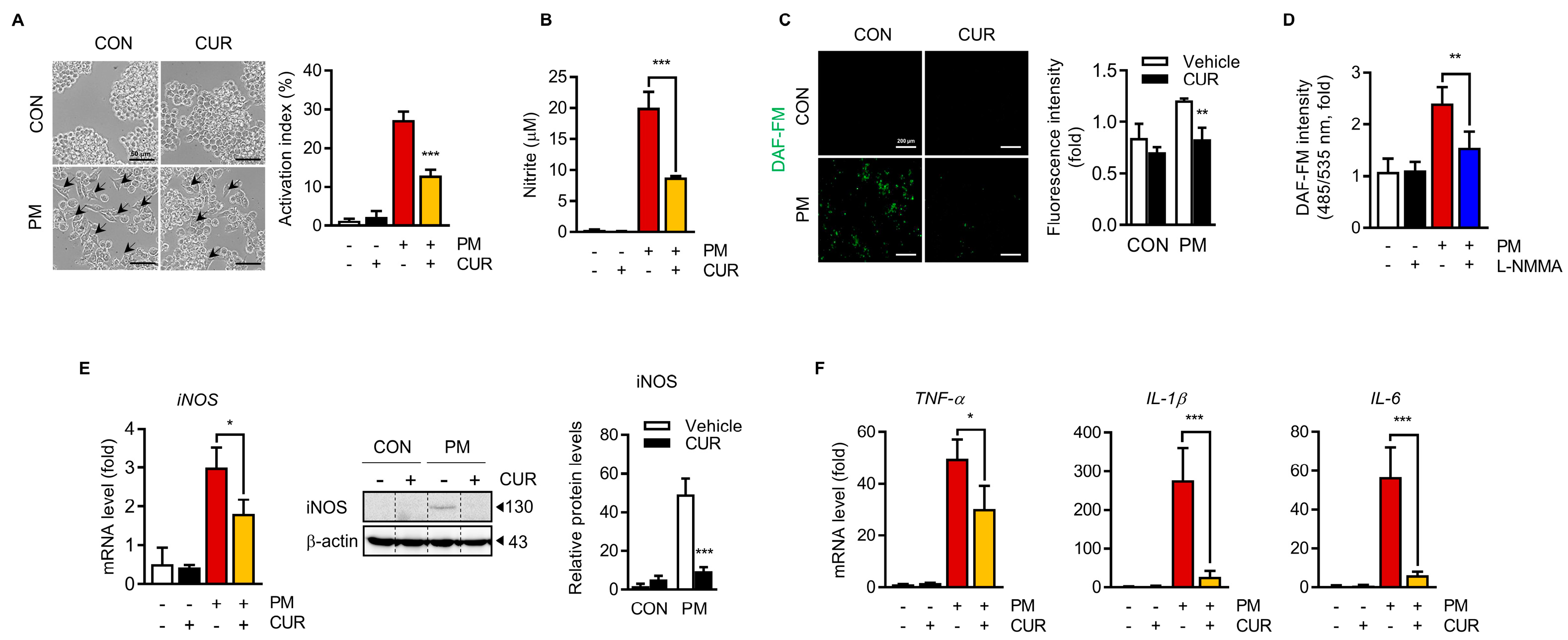
2.3. Curcumin Attenuated the PM-Induced NO Production and Inflammatory Cytokine Expression in RAW264.7 Macrophage Cells
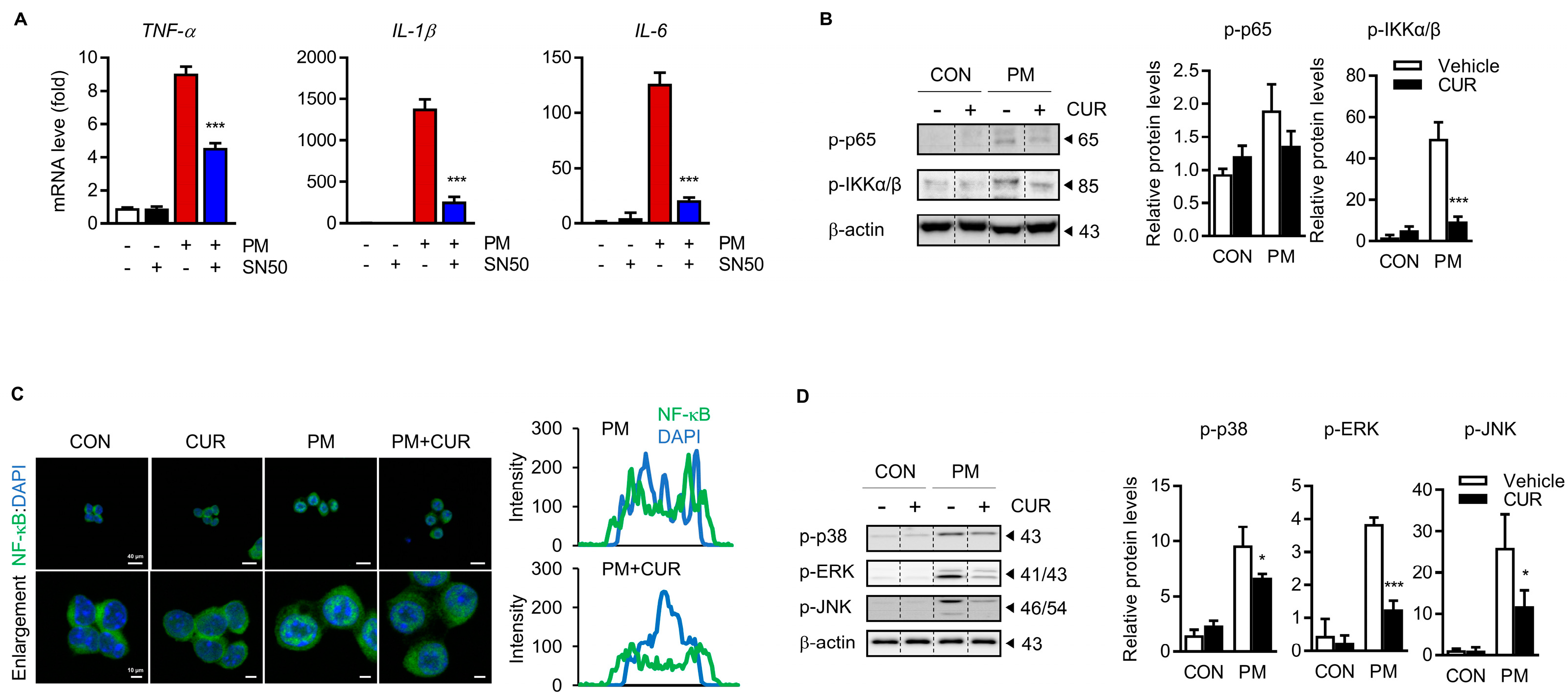
2.4. The Inhibitory Effect of Curcumin on PM-Induced Cytokine Production Was Dependent on the Mitogen-Activated Protein Kinase (MAPK) and NF-kB Signaling Pathways
2.5. Curcumin Reduced the PM-Induced Oxidative Stress through the Nrf2/HO-1 Pathway with Inhibition of Keap1
3. Discussion
4. Materials and Methods
4.1. Animals and Reagents
4.2. PM Inhalation and Curcumin Treatment
4.3. Collection and Analysis of BALF
4.4. Lung Histopathology and Immunofluorescence Analysis
4.5. RNA Extraction and qPCR Analysis
4.6. Cell Culture
4.7. Nitrite Assay
4.8. Intracellular Detection of Oxidative Nitrosylation
4.9. Intracellular ROS Assessment
4.10. Immunoblot Analysis
4.11. Quantification and Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Di, Q.; Wang, Y.; Zanobetti, A.; Wang, Y.; Koutrakis, P.; Choirat, C.; Dominici, F.; Schwartz, J.D. Air Pollution and Mortality in the Medicare Population. N. Engl. J. Med. 2017, 376, 2513–2522. [Google Scholar] [CrossRef] [PubMed]
- Schraufnagel, D.E.; Balmes, J.R.; Cowl, C.T.; De Matteis, S.; Jung, S.-H.; Mortimer, K.; Perez-Padilla, R.; Rice, M.B.; Riojas-Rodriguez, H.; Sood, A.; et al. Air Pollution and Noncommunicable Diseases: A Review by the Forum of International Respiratory Societies’ Environmental Committee, Part 2: Air Pollution and Organ Systems. Chest 2019, 155, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Lelieveld, J.; Klingmüller, K.; Pozzer, A.; Pöschl, U.; Fnais, M.; Daiber, A.; Münzel, T. Cardiovascular Disease Burden from Ambient Air Pollution in Europe Reassessed Using Novel Hazard Ratio Functions. Eur. Heart J. 2019, 40, 1590–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.S.V.; Langrish, J.P.; Nair, H.; McAllister, D.A.; Hunter, A.L.; Donaldson, K.; Newby, D.E.; Mills, N.L. Global Association of Air Pollution and Heart Failure: A Systematic Review and Meta-Analysis. Lancet 2013, 382, 1039–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, A.S.; Newman, L.S. Genetic and Environmental Modulation of Chronic Obstructive Pulmonary Disease. Respir. Physiol. 2001, 128, 3–11. [Google Scholar] [CrossRef]
- Gavett, S.H.; Koren, H.S. The Role of Particulate Matter in Exacerbation of Atopic Asthma. Int. Arch. Allergy Immunol. 2001, 124, 109–112. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, F.; Roepstorff, P.; Yang, F.; Lu, Z.; Ding, W. TMT-Based Quantitative Proteomics Analysis Reveals Airborne PM2.5-Induced Pulmonary Fibrosis. Int. J. Environ. Res. Public Health 2018, 16, 98. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.R. The Cardiovascular Effects of Air Pollution: Prevention and Reversal by Pharmacological Agents. Pharmacol. Ther. 2022, 232, 107996. [Google Scholar] [CrossRef]
- Münzel, T.; Gori, T.; Al-Kindi, S.; Deanfield, J.; Lelieveld, J.; Daiber, A.; Rajagopalan, S. Effects of Gaseous and Solid Constituents of Air Pollution on Endothelial Function. Eur. Heart J. 2018, 39, 3543–3550. [Google Scholar] [CrossRef] [Green Version]
- Shiraiwa, M.; Selzle, K.; Pöschl, U. Hazardous Components and Health Effects of Atmospheric Aerosol Particles: Reactive Oxygen Species, Soot, Polycyclic Aromatic Compounds and Allergenic Proteins. Free Radic. Res. 2012, 46, 927–939. [Google Scholar] [CrossRef]
- Miller, M.R. Oxidative Stress and the Cardiovascular Effects of Air Pollution. Free Radic. Biol. Med. 2020, 151, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Glencross, D.A.; Ho, T.-R.; Camiña, N.; Hawrylowicz, C.M.; Pfeffer, P.E. Air Pollution and Its Effects on the Immune System. Free Radic. Biol. Med. 2020, 151, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Schins, R.P.; Knaapen, A.M.; Weishaupt, C.; Winzer, A.; Borm, P.J. Cytotoxic and Inflammatory Effects of Coarse and Fine Particulate Matter in Macrophages and Epithelial Cells. Ann. Occup. Hyg. 2002, 46, 203–206. [Google Scholar] [CrossRef] [Green Version]
- Shukla, A.; Timblin, C.; BeruBe, K.; Gordon, T.; McKinney, W.; Driscoll, K.; Vacek, P.; Mossman, B.T. Inhaled Particulate Matter Causes Expression of Nuclear Factor (NF)-KappaB-Related Genes and Oxidant-Dependent NF-KappaB Activation in Vitro. Am. J. Respir. Cell Mol. Biol. 2000, 23, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Oh, E.-Y.; Lee, J.-H.; Park, J.-W.; Park, H.J. Effects of Particulate Matter 10 Inhalation on Lung Tissue RNA Expression in a Murine Model. Tuberc. Respir. Dis. 2021, 84, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Tsai, D.-H.; Amyai, N.; Marques-Vidal, P.; Wang, J.-L.; Riediker, M.; Mooser, V.; Paccaud, F.; Waeber, G.; Vollenweider, P.; Bochud, M. Effects of Particulate Matter on Inflammatory Markers in the General Adult Population. Part. Fibre Toxicol. 2012, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Shakoor, H.; Feehan, J.; Apostolopoulos, V.; Platat, C.; Al Dhaheri, A.S.; Ali, H.I.; Ismail, L.C.; Bosevski, M.; Stojanovska, L. Immunomodulatory Effects of Dietary Polyphenols. Nutrients 2021, 13, 728. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxidative Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Bloedon, T.K.; Braithwaite, R.E.; Carson, I.A.; Klimis-Zacas, D.; Lehnhard, R.A. Impact of Anthocyanin-Rich Whole Fruit Consumption on Exercise-Induced Oxidative Stress and Inflammation: A Systematic Review and Meta-Analysis. Nutr. Rev. 2019, 77, 630–645. [Google Scholar] [CrossRef]
- Ola, M.S.; Al-Dosari, D.; Alhomida, A.S. Role of Oxidative Stress in Diabetic Retinopathy and the Beneficial Effects of Flavonoids. Curr. Pharm. Des. 2018, 24, 2180–2187. [Google Scholar] [CrossRef]
- Moghadamtousi, S.Z.; Kadir, H.A.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A Review on Antibacterial, Antiviral, and Antifungal Activity of Curcumin. BioMed Res. Int. 2014, 2014, 186864. [Google Scholar] [CrossRef] [Green Version]
- Ramezani, M.; Hatamipour, M.; Sahebkar, A. Promising Anti-Tumor Properties of Bisdemethoxycurcumin: A Naturally Occurring Curcumin Analogue. J. Cell. Physiol. 2018, 233, 880–887. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yue, Y.; Zheng, X.; Zhang, K.; Chen, S.; Du, Z. Curcumin, Inflammation, and Chronic Diseases: How Are They Linked? Molecules 2015, 20, 9183–9213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, W.; Qian, K.; Xiong, J.; Ma, K.; Wang, A.; Zou, Y. Curcumin Alleviates Lipopolysaccharide Induced Sepsis and Liver Failure by Suppression of Oxidative Stress-Related Inflammation via PI3K/AKT and NF-ΚB Related Signaling. Biomed. Pharmacother. 2016, 83, 302–313. [Google Scholar] [CrossRef]
- Li, R.; Fang, H.; Shen, J.; Jin, Y.; Zhao, Y.; Wang, R.; Fu, Y.; Tian, Y.; Yu, H.; Zhang, J. Curcumin Alleviates LPS-Induced Oxidative Stress, Inflammation and Apoptosis in Bovine Mammary Epithelial Cells via the NFE2L2 Signaling Pathway. Toxins 2021, 13, 208. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Wu, X. Curcumin Protects Trabecular Meshwork Cells From Oxidative Stress. Investig. Ophthalmol. Vis. Sci. 2016, 57, 4327–4332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-S.; Oh, J.-M.; Choi, H.; Kim, S.W.; Kim, S.W.; Kim, B.G.; Cho, J.H.; Lee, J.; Lee, D.C. Activation of the Nrf2/HO-1 Pathway by Curcumin Inhibits Oxidative Stress in Human Nasal Fibroblasts Exposed to Urban Particulate Matter. BMC Complement. Med. Ther. 2020, 20, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Huang, X.-L.; Chen, J.; Mao, L.-N.; Liu, X.; Yuan, W.-S.; Wu, X.-J.; Luo, G.-W. Curcumin Protects BEAS-2B Cells from PM2.5-induced Oxidative Stress and Inflammation by Activating NRF2/Antioxidant Response Element Pathways. Int. J. Mol. Med. 2021, 47, 45. [Google Scholar] [CrossRef] [PubMed]
- Hogg, J.C.; van Eeden, S. Pulmonary and Systemic Response to Atmospheric Pollution. Respirology 2009, 14, 336–346. [Google Scholar] [CrossRef]
- Hiraiwa, K.; van Eeden, S.F. Contribution of Lung Macrophages to the Inflammatory Responses Induced by Exposure to Air Pollutants. Mediat. Inflamm. 2013, 2013, 619523. [Google Scholar] [CrossRef]
- Basnet, P.; Skalko-Basnet, N. Curcumin: An Anti-Inflammatory Molecule from a Curry Spice on the Path to Cancer Treatment. Molecules 2011, 16, 4567–4598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadus, M.C.; Lau, C.; Bikhchandani, J.; Lynch, H.T. Curcumin: An Age-Old Anti-Inflammatory and Anti-Neoplastic Agent. J. Tradit. Complement. Med. 2017, 7, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lelli, D.; Sahebkar, A.; Johnston, T.P.; Pedone, C. Curcumin Use in Pulmonary Diseases: State of the Art and Future Perspectives. Pharmacol. Res. 2017, 115, 133–148. [Google Scholar] [CrossRef]
- Yuan, J.; Liu, R.; Ma, Y.; Zhang, Z.; Xie, Z. Curcumin Attenuates Airway Inflammation and Airway Remolding by Inhibiting NF-ΚB Signaling and COX-2 in Cigarette Smoke-Induced COPD Mice. Inflammation 2018, 41, 1804–1814. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Shi, C.; Min, J.; Li, L.; Zhu, T.; Yu, H.; Deng, H. Study on the Mechanism of Curcumin Regulating Lung Injury Induced by Outdoor Fine Particulate Matter (PM2.5). Mediat. Inflamm. 2019, 2019, 8613523. [Google Scholar] [CrossRef] [Green Version]
- Byrne, A.J.; Mathie, S.A.; Gregory, L.G.; Lloyd, C.M. Pulmonary Macrophages: Key Players in the Innate Defence of the Airways. Thorax 2015, 70, 1189–1196. [Google Scholar] [CrossRef] [Green Version]
- Boorsma, C.E.; Draijer, C.; Melgert, B.N. Macrophage Heterogeneity in Respiratory Diseases. Mediat. Inflamm. 2013, 2013, 769214. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith, C.A.; Frevert, C.; Imrich, A.; Sioutas, C.; Kobzik, L. Alveolar Macrophage Interaction with Air Pollution Particulates. Environ. Health Perspect. 1997, 105 (Suppl. S5), 1191–1195. [Google Scholar] [CrossRef] [Green Version]
- Michael, S.; Montag, M.; Dott, W. Pro-Inflammatory Effects and Oxidative Stress in Lung Macrophages and Epithelial Cells Induced by Ambient Particulate Matter. Environ. Pollut. 2013, 183, 19–29. [Google Scholar] [CrossRef]
- Rao, K.M. MAP Kinase Activation in Macrophages. J. Leukoc. Biol. 2001, 69, 3–10. [Google Scholar] [CrossRef]
- Jayakumar, T.; Lin, K.-C.; Chang, C.-C.; Hsia, C.-W.; Manubolu, M.; Huang, W.-C.; Sheu, J.-R.; Hsia, C.-H. Targeting MAPK/NF-ΚB Pathways in Anti-Inflammatory Potential of Rutaecarpine: Impact on Src/FAK-Mediated Macrophage Migration. Int. J. Mol. Sci. 2021, 23, 92. [Google Scholar] [CrossRef] [PubMed]
- Dorrington, M.G.; Fraser, I.D.C. NF-ΚB Signaling in Macrophages: Dynamics, Crosstalk, and Signal Integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef] [PubMed]
- Bagaev, A.V.; Garaeva, A.Y.; Lebedeva, E.S.; Pichugin, A.V.; Ataullakhanov, R.I.; Ataullakhanov, F.I. Elevated Pre-Activation Basal Level of Nuclear NF-ΚB in Native Macrophages Accelerates LPS-Induced Translocation of Cytosolic NF-ΚB into the Cell Nucleus. Sci. Rep. 2019, 9, 4563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 Signaling Pathway: Pivotal Roles in Inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-Antioxidant Response Element Signaling Pathway and Its Activation by Oxidative Stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef] [Green Version]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting Molecular Cross-Talk between Nrf2 and NF-ΚB Response Pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Rayamajhi, M.; Redente, E.F.; Condon, T.V.; Gonzalez-Juarrero, M.; Riches, D.W.H.; Lenz, L.L. Non-Surgical Intratracheal Instillation of Mice with Analysis of Lungs and Lung Draining Lymph Nodes by Flow Cytometry. J. Vis. Exp. 2011, 51, e2702. [Google Scholar] [CrossRef]
- Limjunyawong, N.; Mock, J.; Mitzner, W. Instillation and Fixation Methods Useful in Mouse Lung Cancer Research. J. Vis. Exp. 2015, 102, e52964. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Xue, X. Detection of Total Reactive Oxygen Species in Adherent Cells by 2′,7′-Dichlorodihydrofluorescein Diacetate Staining. J. Vis. Exp. 2020, 160, e60682. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.K.; Kim, H.D.; Lee, S.H.; Lee, J.H. Curcumin Ameliorates Particulate Matter-Induced Pulmonary Injury through Bimodal Regulation of Macrophage Inflammation via NF-κB and Nrf2. Int. J. Mol. Sci. 2023, 24, 1858. https://doi.org/10.3390/ijms24031858
Lee MK, Kim HD, Lee SH, Lee JH. Curcumin Ameliorates Particulate Matter-Induced Pulmonary Injury through Bimodal Regulation of Macrophage Inflammation via NF-κB and Nrf2. International Journal of Molecular Sciences. 2023; 24(3):1858. https://doi.org/10.3390/ijms24031858
Chicago/Turabian StyleLee, Min Kook, Hyo Dam Kim, Suk Hee Lee, and Jin Hyup Lee. 2023. "Curcumin Ameliorates Particulate Matter-Induced Pulmonary Injury through Bimodal Regulation of Macrophage Inflammation via NF-κB and Nrf2" International Journal of Molecular Sciences 24, no. 3: 1858. https://doi.org/10.3390/ijms24031858