
Endurance Training Increases the Running Performance of Untrained Men without Changing the Mitochondrial Volume Density in the Gastrocnemius Muscle
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Physical Characteristics and HRmax
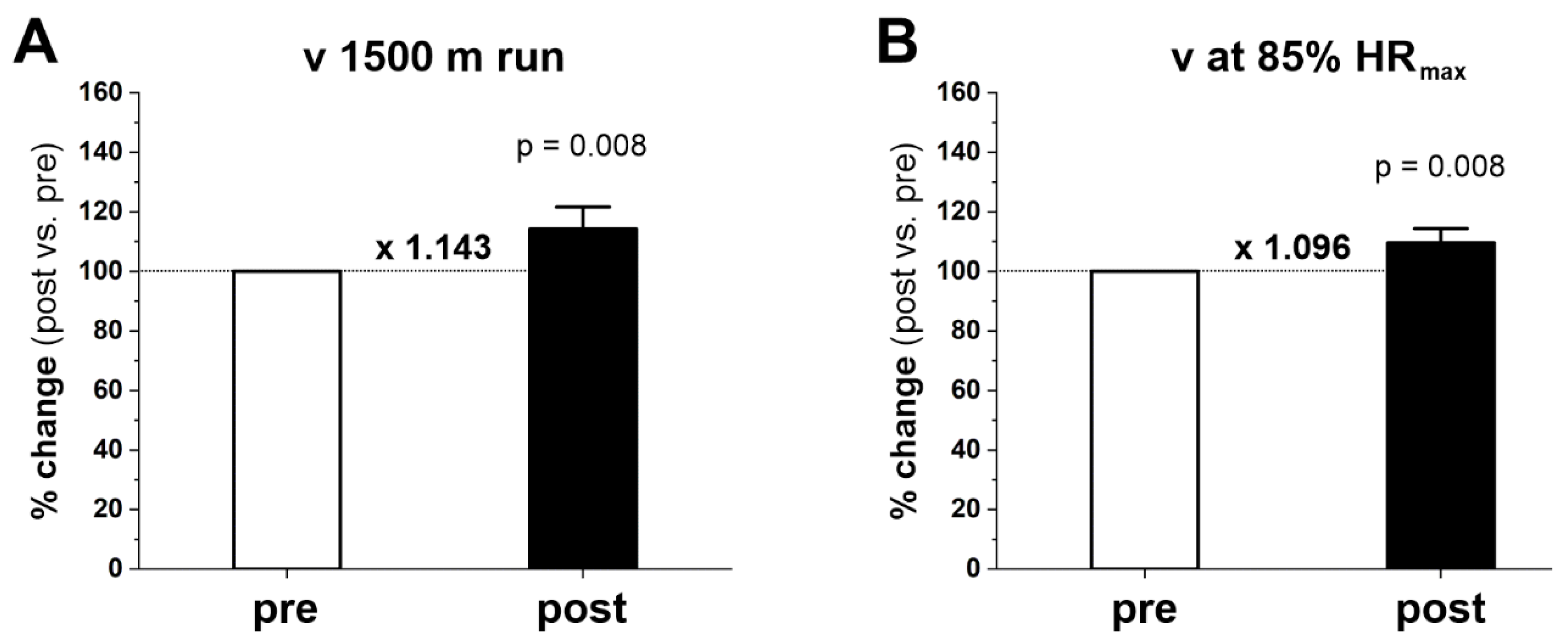
2.2. Running Performance
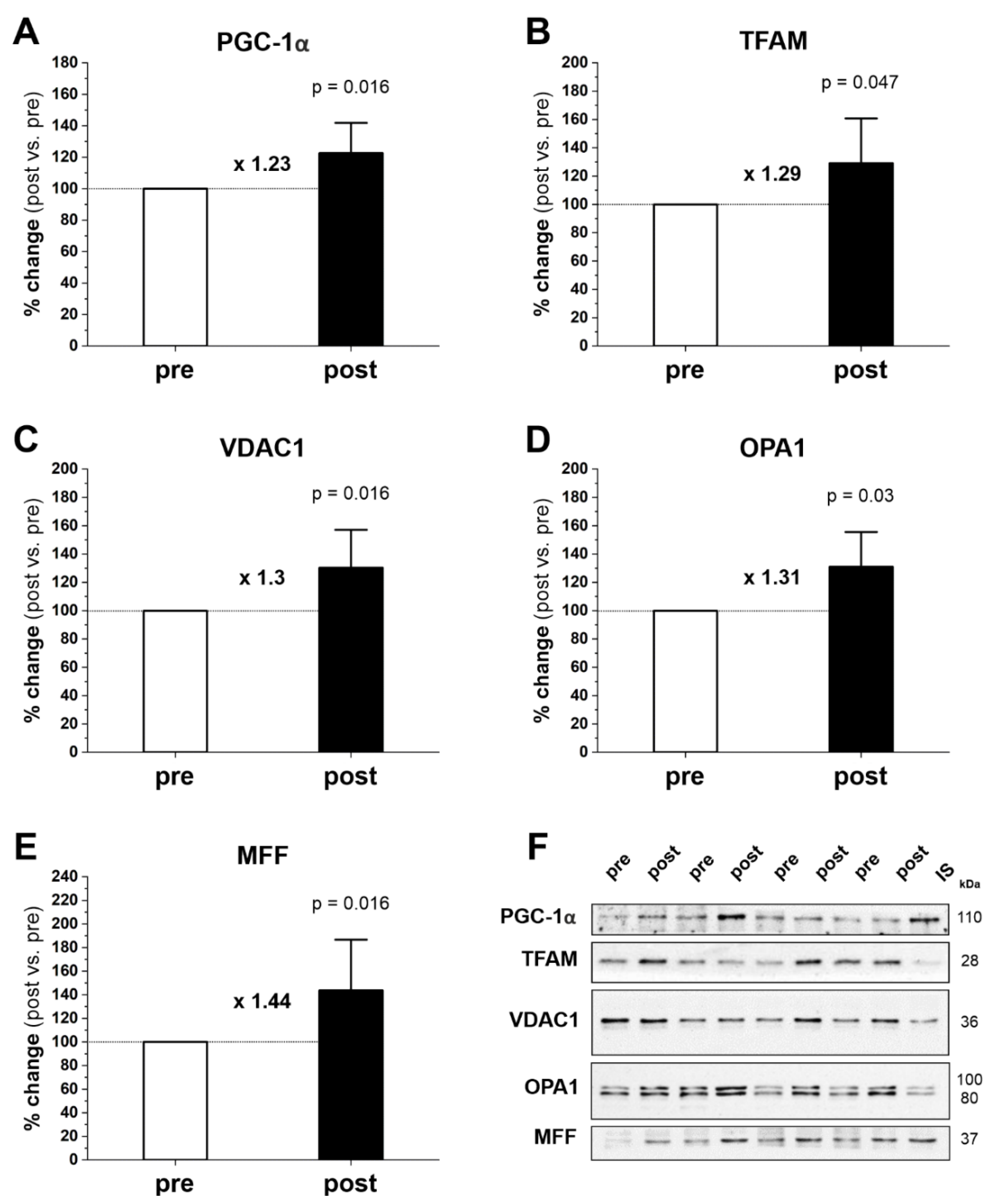
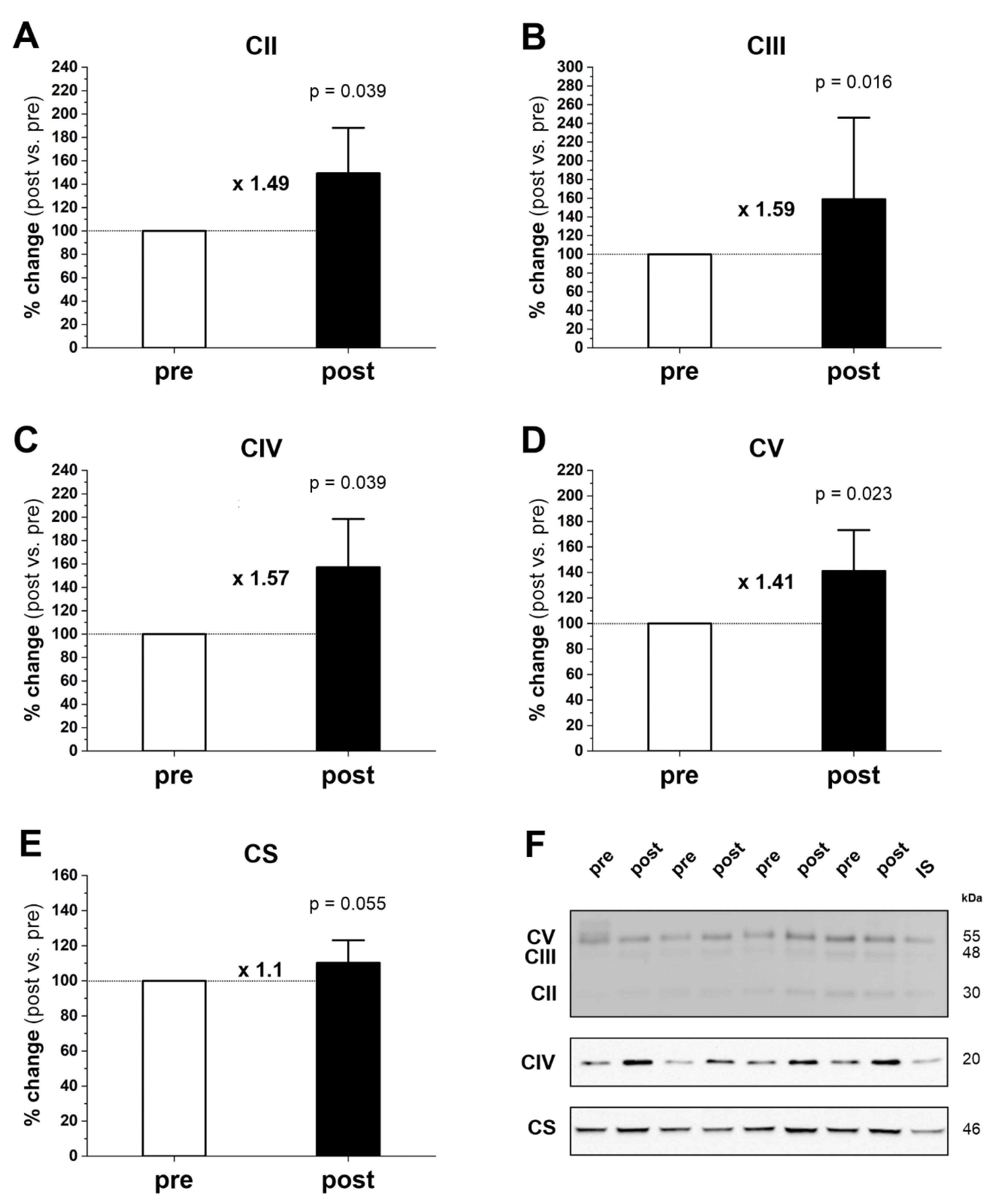
2.3. Mitochondrial Turnover
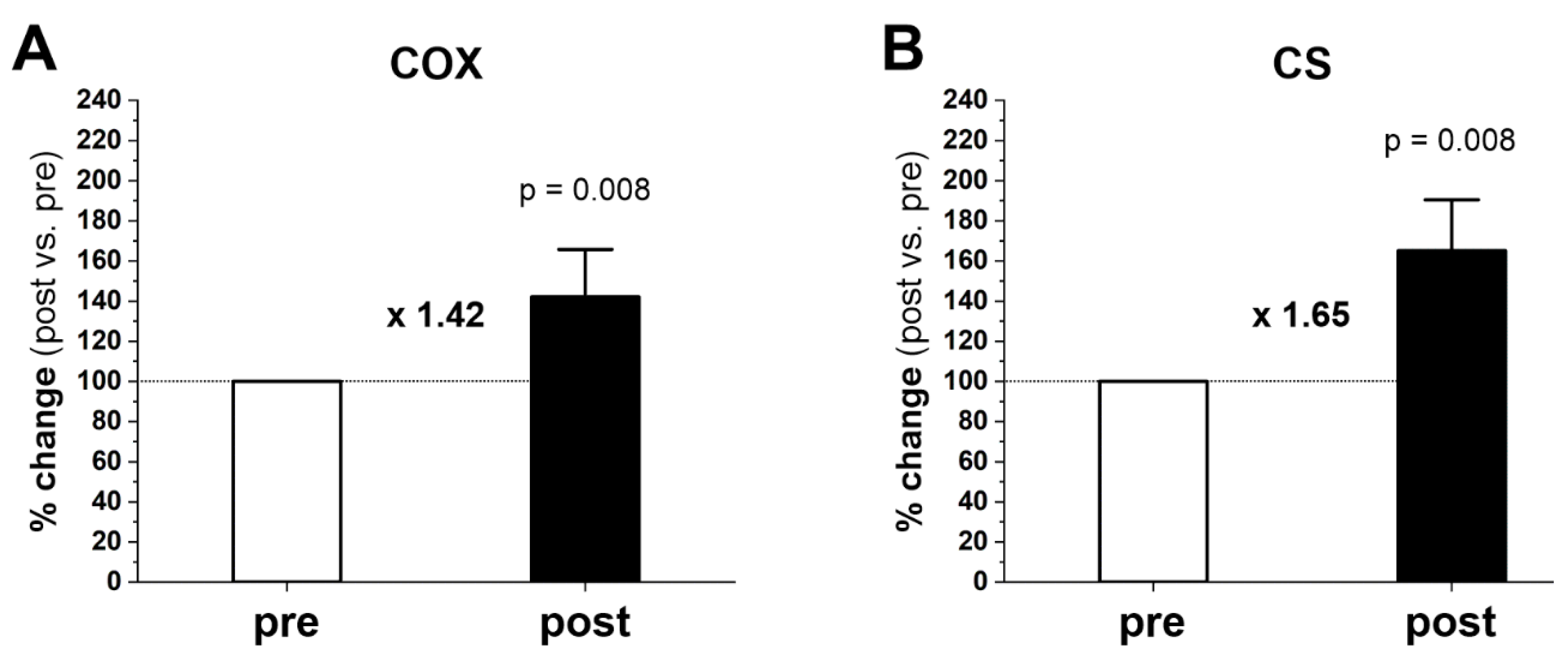
2.4. Muscle COX and CS Activities
2.5. Mitochondrial Content in the Gastrocnemius Muscle
3. Discussion
3.1. Training and VO2max
3.2. Training and Running Economy
3.3. Mitochondrial Volume Density
3.4. Muscle COX and CS Activities
3.5. The Effect of Muscle OXPHOS Activity on Running Performance
3.6. Other Potential Training-Induced Factors Important for Improving Running Speed
3.7. Summing Up
4. Materials and Methods
4.1. Subjects
4.2. Running Performance
4.3. Muscle Biopsy and Analyses
4.4. Muscle Mitochondrial Protein Content
4.5. Maximal CS and COX Activities
4.6. Muscle Morphometry
4.7. Endurance Training Program
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pedersen, B.K.; Saltin, B. Exercise as medicine—Evidence for prescribing exercise as therapy in 26 different chronic diseases. Scand. J. Med. Sci. Sports 2015, 25 (Suppl. S3), 1–72. [Google Scholar] [CrossRef] [PubMed]
- Lundby, C.; Montero, D.; Joyner, M. Biology of VO2max: Looking under the physiology lamp. Acta Physiol. 2017, 220, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Joyner, M.J.; Lundby, C. Concepts about VO2max and trainability are context dependent. Exerc. Sport Sci. Rev. 2018, 46, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Holloszy, J.O. Biochemical adaptations in muscle. Effects of exercise on mitochondrial oxygen uptake and respiratory enzyme activity in skeletal muscle. J. Biol. Chem. 1967, 242, 2278–2282. [Google Scholar] [CrossRef]
- Morgan, T.E.; Cobb, L.A.; Short, F.A.; Ross, R.; Gunn, D.R. Effects of long-term exercise on human muscle mitochondria. In Muscle Metabolism during Exercise; Pernow, B., Saltin, B., Eds.; Springer: Boston, MA, USA, 1971; pp. 87–95. [Google Scholar]
- Henriksson, J.; Reitman, J.S. Time course of changes in human skeletal muscle succinate dehydrogenase and cytochrome oxidase activities and maximal oxygen uptake with physical activity and inactivity. Acta Physiol. Scand. 1977, 99, 91–97. [Google Scholar] [CrossRef]
- Wibom, R.; Hultman, E.; Johansson, M.; Matherei, K.; Constantin-Teodosiu, D.; Schantz, P.G. Adaptation of mitochondrial ATP production in human skeletal muscle to endurance training and detraining. J. Appl. Physiol. 1992, 73, 2004–2010. [Google Scholar] [CrossRef]
- Baldwin, K.; Haddad, F. Effects of exercise training on the biochemical pathways associated with sarcopenia. In Muscle and Exercise Physiology; Zoladz, J.A., Ed.; Elsevier Inc. Academic Press: London, UK, 2019; pp. 347–377. [Google Scholar] [CrossRef]
- Hoppeler, H.; Howald, H.; Conley, K.; Lindstedt, S.L.; Claassen, H.; Vock, P.; Weibel, E.R. Endurance training in humans: Aerobic capacity and structure of skeletal muscle. J. Appl. Physiol. 1985, 59, 320–327. [Google Scholar] [CrossRef]
- Suter, E.; Hoppeler, H.; Claassen, H.; Billeter, R.; Aebi, U.; Horber, F.; Jaeger, P.; Marti, B. Ultrastructural modification of human skeletal muscle tissue with 6-month moderate-intensity exercise training. Int. J. Sports Med. 1995, 16, 160–166. [Google Scholar] [CrossRef]
- Turner, D.L.; Hoppeler, H.; Claassen, H.; Vock, P.; Kayser, B.; Schena, F.; Ferretti, G. Effects of endurance training on oxidative capacity and structural composition of human arm and leg muscles. Acta Physiol. Scand. 1997, 161, 459–464. [Google Scholar] [CrossRef]
- Montero, D.; Cathomen, A.; Jacobs, R.A.; Flück, D.; de Leur, J.; Keiser, S.; Bonne, T.; Kirk, N.; Lundby, A.-K.; Lundby, C. Haematological rather than skeletal muscle adaptations contribute to the increase in peak oxygen uptake induced by moderate endurance training. J. Physiol. 2015, 593, 4677–4688. [Google Scholar] [CrossRef]
- Meinild Lundby, A.-K.; Jacobs, R.A.; Gehrig, S.; de Leur, J.; Hauser, M.; Bonne, T.C.; Flück, D.; Dandanell, S.; Kirk, N.; Kaech, A.; et al. Exercise training increases skeletal muscle mitochondrial volume density by enlargement of existing mitochondria and not de novo biogenesis. Acta Physiol. 2018, 222, e12976. [Google Scholar] [CrossRef] [PubMed]
- Hoppeler, H.; Lüthi, P.; Claassen, H.; Weibel, E.R.; Howald, H. The ultrastructure of the normal human skeletal muscle. A morphometric analysis on untrained men, women and well-trained orienteers. Pflug. Arch. 1973, 344, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Zoladz, J.A.; Koziel, A.; Woyda-Ploszczyca, A.; Celichowski, J.; Jarmuszkiewicz, W. Endurance training increases the efficiency of rat skeletal muscle mitochondria. Pflug. Arch. 2016, 468, 1709–1724. [Google Scholar] [CrossRef] [PubMed]
- Lundby, C.; Jacobs, R.A. Adaptations of skeletal muscle mitochondria to exercise training. Exp. Physiol. 2016, 101, 17–22. [Google Scholar] [CrossRef]
- Larsen, S.; Nielsen, J.; Hansen, C.N.; Nielsen, L.B.; Wibrand, F.; Stride, N.; Schroder, H.D.; Boushel, R.; Helge, J.W.; Dela, F.; et al. Biomarkers of mitochondrial content in skeletal muscle of healthy young human subjects. J. Physiol. 2012, 590, 3349–3360. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Grassi, B.; Majerczak, J.; Szkutnik, Z.; Korostyński, M.; Grandys, M.; Jarmuszkiewicz, W.; Korzeniewski, B. Mechanisms responsible for the acceleration of pulmonary O2 on-kinetics in humans after prolonged endurance training. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R1101–R1114. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Grassi, B.; Szkutnik, Z. Metabolic transitions and muscle metabolic stability. effects of exercise training. In Muscle and Excercise Physiology; Zoladz, J.A., Ed.; Elsevier Inc. Academic Press: London, UK, 2019; pp. 391–422. [Google Scholar] [CrossRef]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef]
- Scarpulla, R.C. Nuclear control of respiratory gene expression in mammalian cells. J. Cell. Biochem. 2006, 97, 673–683. [Google Scholar] [CrossRef]
- Ngo, H.B.; Lovely, G.A.; Phillips, R.; Chan, D.C. Distinct structural features of TFAM drive mitochondrial DNA packaging versus transcriptional activation. Nat. Commun. 2014, 5, 3077. [Google Scholar] [CrossRef]
- Kukat, C.; Davies, K.M.; Wurm, C.A.; Spåhr, H.; Bonekamp, N.A.; Kühl, I.; Joos, F.; Polosa, P.L.; Park, C.B.; Posse, V.; et al. Cross-strand binding of TFAM to a single mtDNA molecule forms the mitochondrial nucleoid. Proc. Natl. Acad. Sci. USA 2015, 112, 11288–11293. [Google Scholar] [CrossRef]
- Long, Q.; Zhou, Y.; Wu, H.; Du, S.; Hu, M.; Qi, J.; Li, W.; Guo, J.; Wu, Y.; Yang, L.; et al. Phase separation drives the self-assembly of mitochondrial nucleoids for transcriptional modulation. Nat. Struct. Mol. Biol. 2021, 28, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Granata, C.; Jamnick, N.A.; Bishop, D.J. Principles of exercise prescription, and how they influence exercise-induced changes of transcription factors and other regulators of mitochondrial biogenesis. Sports Med. 2018, 48, 1541–1559. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-H.; Kim, K.M.; Oh, H.T.; Yoo, G.D.; Jeong, M.G.; Lee, H.; Park, J.; Jeong, K.; Kim, Y.K.; Ko, Y.-G.; et al. TAZ links exercise to mitochondrial biogenesis via mitochondrial transcription factor A. Nat. Commun. 2022, 13, 653. [Google Scholar] [CrossRef] [PubMed]
- Moitra, A.; Rapaport, D. The biogenesis process of VDAC—From early cytosolic events to its final membrane integration. Front. Physiol. 2021, 12, 732742. [Google Scholar] [CrossRef]
- Slavin, M.B.; Memme, J.M.; Oliveira, A.N.; Moradi, N.; Hood, D.A. Regulatory networks controlling mitochondrial quality control in skeletal muscle. Am. J. Physiol. Cell Physiol. 2022, 322, C913–C926. [Google Scholar] [CrossRef]
- Pesta, D.; Hoppel, F.; Macek, C.; Messner, H.; Faulhaber, M.; Kobel, C.; Parson, W.; Burtscher, M.; Schocke, M.; Gnaiger, E. Similar qualitative and quantitative changes of mitochondrial respiration following strength and endurance training in normoxia and hypoxia in sedentary humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1078–R1087. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Grassi, B.; Majerczak, J.; Szkutnik, Z.; Korostyński, M.; Karasiński, J.; Kilarski, W.; Korzeniewski, B. Training-induced acceleration of O2 uptake on-kinetics precedes muscle mitochondrial biogenesis in humans. Exp. Physiol. 2013, 98, 883–898. [Google Scholar] [CrossRef]
- Léger, L.; Mercier, D. Gross energy cost of horizontal treadmill and track running. Sports Med. 1984, 1, 270–277. [Google Scholar] [CrossRef]
- Billat, V.L.; Morton, R.H.; Blondel, N.; Berthoin, S.; Bocquet, V.; Koralsztein, J.P.; Barstow, T.J. Oxygen kinetics and modelling of time to exhaustion whilst running at various velocities at maximal oxygen uptake. Eur. J. Appl. Physiol. 2000, 82, 178–187. [Google Scholar] [CrossRef]
- Gollnick, P.D.; Armstrong, R.B.; Saltin, B.; Saubert, C.W., 4th; Sembrowich, W.L.; Shepherd, R.E. Effect of training on enzyme activity and fiber composition of human skeletal muscle. J. Appl. Physiol. 1973, 34, 107–111. [Google Scholar] [CrossRef]
- Korzeniewski, B.; Rossiter, H.B. Factors determining training-induced changes in O(2max), critical power, and O(2) on-kinetics in skeletal muscle. J. Appl. Physiol. 2021, 130, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, J.; Nordesjö, L.O.; Jorfeldt, L.; Saltin, B. Muscle lactate, ATP, and CP levels during exercise after physical training in man. J. Appl. Physiol. 1972, 33, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Barnes, K.R.; Kilding, A.E. Running economy: Measurement, norms, and determining factors. Sport. Med.-Open 2015, 1, 8. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Kirby, B.S.; Clark, I.E.; Rice, H.M.; Fulkerson, E.; Wylie, L.J.; Wilkerson, D.P.; Vanhatalo, A.; Wilkins, B.W. Physiological demands of running at 2-hour marathon race pace. J. Appl. Physiol. 2021, 130, 369–379. [Google Scholar] [CrossRef]
- Majerczak, J.; Korostynski, M.; Nieckarz, Z.; Szkutnik, Z.; Duda, K.; Zoladz, J.A. Endurance training decreases the non-linearity in the oxygen uptake-power output relationship in humans. Exp. Physiol. 2012, 97, 386–399. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Rademaker, A.C.; Sargeant, A.J. Non-linear relationship between O2 uptake and power output at high intensities of exercise in humans. J. Physiol. 1995, 488, 211–217. [Google Scholar] [CrossRef]
- Ellis, R.G.; Sumner, B.J.; Kram, R. Muscle contributions to propulsion and braking during walking and running: Insight from external force perturbations. Gait Posture 2014, 40, 594–599. [Google Scholar] [CrossRef]
- Nielsen, J.; Gejl, K.D.; Hey-Mogensen, M.; Holmberg, H.-C.; Suetta, C.; Krustrup, P.; Elemans, C.P.H.; Ørtenblad, N. Plasticity in mitochondrial cristae density allows metabolic capacity modulation in human skeletal muscle. J. Physiol. 2017, 595, 2839–2847. [Google Scholar] [CrossRef] [Green Version]
- Larsen, S.W.; Helge, J.; Dela, F. Is there plasticity in mitochondrial cristae density with endurance training? J. Physiol. 2017, 595, 2985. [Google Scholar] [CrossRef]
- Zhang, Z.-Y.; Chen, B.; Zhao, D.-J.; Kang, L. Functional modulation of mitochondrial cytochrome c oxidase underlies adaptation to high-altitude hypoxia in a Tibetan migratory locust. Proc. Biol. Sci. 2013, 280, 20122758. [Google Scholar] [CrossRef]
- Antunes, F.; Boveris, A.; Cadenas, E. On the mechanism and biology of cytochrome oxidase inhibition by nitric oxide. Proc. Natl. Acad. Sci. USA 2004, 101, 16774–16779. [Google Scholar] [CrossRef] [PubMed]
- Whipp, B.J.; Wasserman, K. Oxygen uptake kinetics for various intensities of constant-load work. J. Appl. Physiol. 1972, 33, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Zoladz, J.A.; Gladden, L.B.; Hogan, M.C.; Nieckarz, Z.; Grassi, B. Progressive recruitment of muscle fibers is not necessary for the slow component of VO2 kinetics. J. Appl. Physiol. 2008, 105, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Woledge, R.C. Possible effects of fatigue on muscle efficiency. Acta Physiol. Scand. 1998, 162, 267–273. [Google Scholar] [CrossRef]
- Fitts, R.H. The role of acidosis in fatigue: Pro perspective. Med. Sci. Sports Exerc. 2016, 48, 2335–2338. [Google Scholar] [CrossRef]
- Westerblad, H. Acidosis is not a significant cause of skeletal muscle fatigue. Med. Sci. Sports Exerc. 2016, 48, 2339–2342. [Google Scholar] [CrossRef]
- Grassi, B.; Rossiter, H.B.; Zoladz, J.A. Skeletal muscle fatigue and decreased efficiency: Two sides of the same coin? Exerc. Sport Sci. Rev. 2015, 43, 75–83. [Google Scholar] [CrossRef]
- Korzeniewski, B.; Zoladz, J.A. A model of oxidative phosphorylation in mammalian skeletal muscle. Biophys. Chem. 2001, 92, 17–34. [Google Scholar] [CrossRef]
- Korzeniewski, B.; Zoladz, J.A. Factors determining the oxygen consumption rate (VO2) on-kinetics in skeletal muscles. Biochem. J. 2004, 379, 703–710. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Majerczak, J.; Grassi, B.; Szkutnik, Z.; Korostyński, M.; Gołda, S.; Grandys, M.; Jarmuszkiewicz, W.; Kilarski, W.; Karasinski, J.; et al. Mechanisms of attenuation of pulmonary V’O2 slow component in humans after prolonged endurance training. PLoS ONE 2016, 11, e0154135. [Google Scholar] [CrossRef]
- Chang, K.-V.; Wu, W.-T.; Huang, K.-C.; Han, D.-S. Effectiveness of early versus delayed exercise and nutritional intervention on segmental body composition of sarcopenic elders—A randomized controlled trial. Clin. Nutr. 2021, 40, 1052–1059. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.L.; Paul, L.; Steultjens, M.P.M.; Smith, S.L. Biomarkers associated with lower limb muscle function in individuals with sarcopenia: A systematic review. J. Cachexia. Sarcopenia Muscle 2022. [Google Scholar] [CrossRef] [PubMed]
- Bilski, J.; Pierzchalski, P.; Szczepanik, M.; Bonior, J.; Zoladz, J.A. Multifactorial mechanism of sarcopenia and sarcopenic obesity. Role of physical exercise, microbiota and myokines. Cells 2022, 11, 160. [Google Scholar] [CrossRef]
- Bartlett, M.F.; Fitzgerald, L.F.; Kent, J.A. Rates of oxidative ATP synthesis are not augmented beyond the pH threshold in human vastus lateralis muscles during a stepwise contraction protocol. J. Physiol. 2021, 599, 1997–2013. [Google Scholar] [CrossRef]
- Weibel, E.R. Stereological Methods: Practical Methods for Biological Morphometry; Academic Press: New York, NY, USA, 1979; Volume 1. [Google Scholar]
- Dahl, R.; Larsen, S.; Dohlmann, T.L.; Qvortrup, K.; Helge, J.W.; Dela, F.; Prats, C. Three-dimensional reconstruction of the human skeletal muscle mitochondrial network as a tool to assess mitochondrial content and structural organization. Acta Physiol. 2015, 213, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.E.; White, K.; Davey, T.; Philips, J.; Ogden, R.T.; Lawless, C.; Warren, C.; Hall, M.G.; Ng, Y.S.; Falkous, G.; et al. Quantitative 3D Mapping of the Human Skeletal Muscle Mitochondrial Network. Cell Rep. 2019, 26, 996–1009. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Sargeant, A.J.; Emmerich, J.; Stoklosa, J.; Zychowski, A. Changes in acid-base status of marathon runners during an incremental field test. Relationship to mean competitive marathon velocity. Eur. J. Appl. Physiol. Occup. Physiol. 1993, 67, 71–76. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Majerczak, J.; Filipowska, J.; Tylko, G.; Guzik, M.; Karasinski, J.; Piechowicz, E.; Pyza, E.; Chlopicki, S.; Zoladz, J.A. Impact of long-lasting spontaneous physical activity on bone morphogenetic protein 4 in the heart and tibia in murine model of heart failure. Physiol. Rep. 2020, 8, e14412. [Google Scholar] [CrossRef]
- Majerczak, J.; Kij, A.; Drzymala-Celichowska, H.; Kus, K.; Karasinski, J.; Nieckarz, Z.; Grandys, M.; Celichowski, J.; Szkutnik, Z.; Hendgen-Cotta, U.B.; et al. Nitrite concentration in the striated muscles is reversely related to myoglobin and mitochondrial proteins content in rats. Int. J. Mol. Sci. 2022, 23, 2686. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Status | Volume Density of Total Mitochondria in % | Volume Density of IMF Mitochondria in % | Volume Density of SS Mitochondria in % |
|---|---|---|---|
| Before training | 9.78 ± 1.13 | 6.32 ± 1.04 | 3.30 ± 0.47 |
| After training | 8.54 ± 0.81 n.s. | 5.14 ± 0.64 (p = 0.017) | 2.94 ± 0.79 n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoladz, J.A.; Majerczak, J.; Galganski, L.; Grandys, M.; Zapart-Bukowska, J.; Kuczek, P.; Kołodziejski, L.; Walkowicz, L.; Szymoniak-Chochół, D.; Kilarski, W.; et al. Endurance Training Increases the Running Performance of Untrained Men without Changing the Mitochondrial Volume Density in the Gastrocnemius Muscle. Int. J. Mol. Sci. 2022, 23, 10843. https://doi.org/10.3390/ijms231810843
Zoladz JA, Majerczak J, Galganski L, Grandys M, Zapart-Bukowska J, Kuczek P, Kołodziejski L, Walkowicz L, Szymoniak-Chochół D, Kilarski W, et al. Endurance Training Increases the Running Performance of Untrained Men without Changing the Mitochondrial Volume Density in the Gastrocnemius Muscle. International Journal of Molecular Sciences. 2022; 23(18):10843. https://doi.org/10.3390/ijms231810843
Chicago/Turabian StyleZoladz, Jerzy A., Joanna Majerczak, Lukasz Galganski, Marcin Grandys, Justyna Zapart-Bukowska, Piotr Kuczek, Leszek Kołodziejski, Lucyna Walkowicz, Dorota Szymoniak-Chochół, Wincenty Kilarski, and et al. 2022. "Endurance Training Increases the Running Performance of Untrained Men without Changing the Mitochondrial Volume Density in the Gastrocnemius Muscle" International Journal of Molecular Sciences 23, no. 18: 10843. https://doi.org/10.3390/ijms231810843