
Proteomic Analysis of HCC-1954 and MCF-7 Cell Lines Highlights Crosstalk between αv and β1 Integrins, E-Cadherin and HER-2
 , , and
, , and 
Abstract
:1. Introduction
2. Results
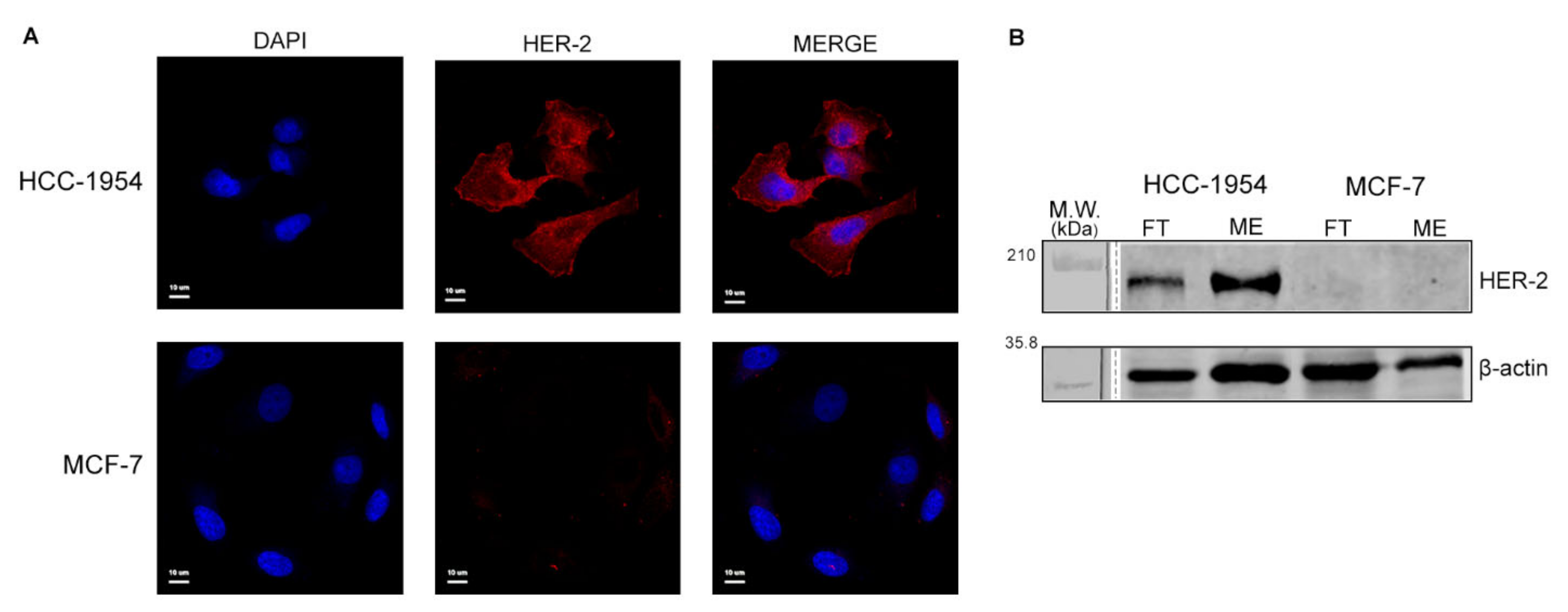
2.1. Analysis of HER-2 Expression in HCC-1954 and MCF-7 Cell Lines Confirm the Difference of Their Breast Cancer Phenotypes
2.2. Label-Free LC-MSE Proteomic Data Quality Analysis Shows Confidence in Protein Expression among the Different Samples
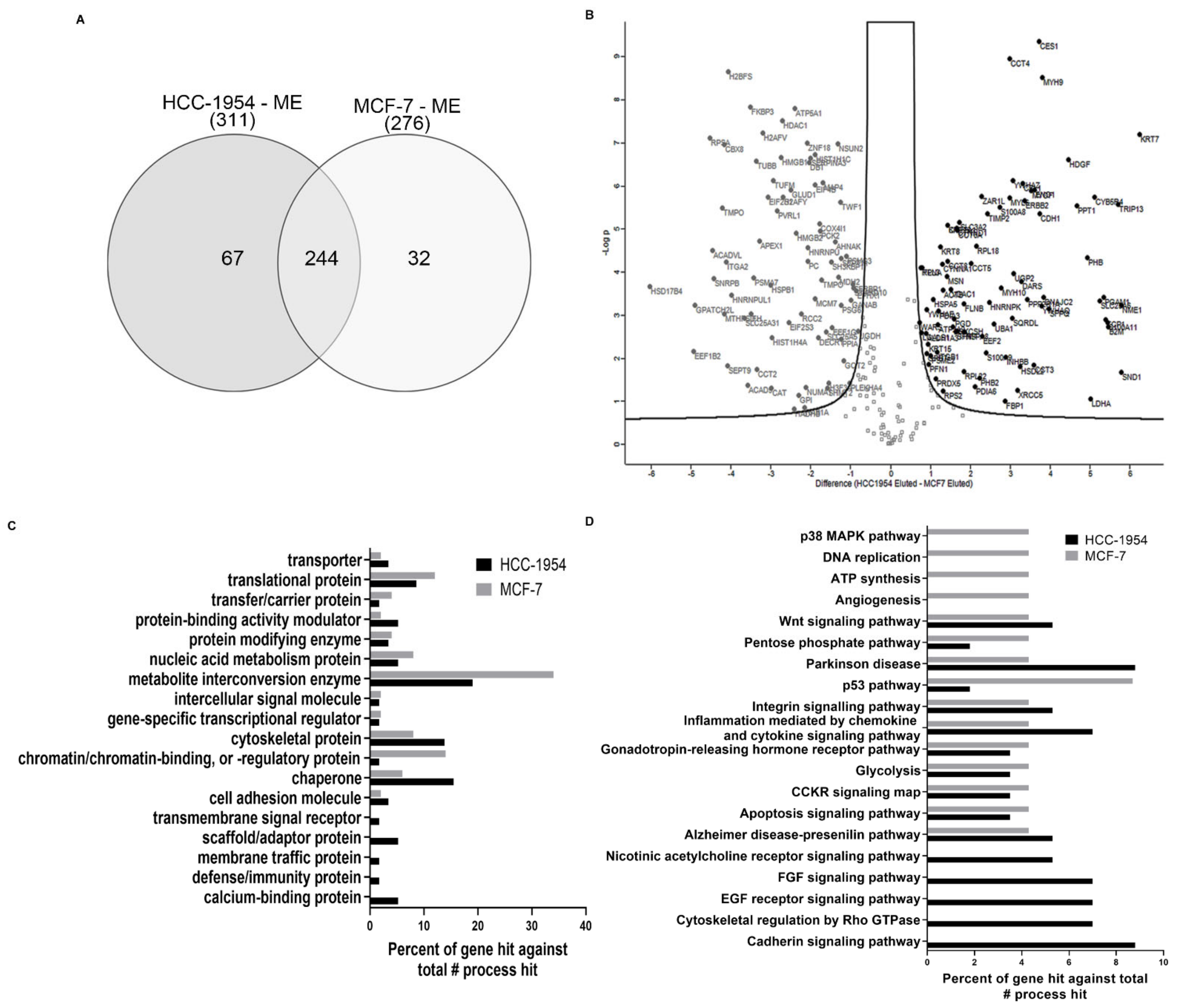
2.3. HCC-1954 and MCF-7 Label-Free LC-MSE Proteomic Statistical Analysis Filtered a Total of 450 Proteins
2.4. Proteins Identified and Quantified for HCC-1954 and MCF-7 Cell Line Fractions Show Statistically Significant Differences in Abundance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HCC-1954 | MCF-7 |
|---|---|
| 14-3-3 protein theta YWHAQ | 26S proteasome regulatory subunit 6A PSMC3 |
| 3-hydroxyacyl-CoA dehydrogenase type-2 HSD17B10 | 4-aminobutyrate aminotransferase, mitochondrial ABAT |
| 40S ribosomal protein S19 RPS19 | Acyl-CoA dehydrogenase family member 9, ACAD9 |
| 40S ribosomal protein S27 RPS27 | Alpha-1-antichymotrypsin SERPINA3 |
| 45 kDa calcium-binding protein SDF4 | Cell division cycle and apoptosis regulator protein 1 CCAR1 |
| 6-phosphogluconate dehydrogenase, decarboxylating PGD | Core histone macro-H2A.1 H2AFY |
| Acetyl-CoA acetyltransferase, mitochondrial ACAT1 | DNA-(apurinic or apyrimidinic site) lyase APEX1 |
| Actin, aortic smooth muscle ACTA2 | Elongation factor 1-delta EEF1D |
| Actin, cytoplasmic 2 ACTG1 | Enoyl-CoA hydratase, mitochondrial ECHS1 |
| Actin-related protein 2/3 complex subunit 1B ARPC1B | Eukaryotic translation initiation factor 2 subunit 3 EIF2S3 |
| Adenosylhomocysteinase AHCY | Eukaryotic translation initiation factor 4B EIF4B |
| ADP,ATP carrier protein, heart isoform T1 SLC25A4 | Heterogeneous nuclear ribonucleoprotein A3 HNRNPA3 |
| Aldehyde dehydrogenase family 1 member A3 ALDH1A3 | Histone H1.2 HIST1H1C |
| Alpha-actinin-1 ACTN1 | Histone H2A.V H2AFV |
| Alpha-enolase ENO1 | Kininogen-1 KNG1 PE = 1 SV = 2 |
| Beta-2-microglobulin B2M | Leucine--tRNA ligase, cytoplasmic LARS |
| Cadherin-1 CDH1 | Methyl-CpG-binding protein 2 MECP2 |
| Catenin alpha-1 CTNNA1 | Nectin-1 NECTIN1 |
| Catenin delta-1 CTNND1 | Neuroblast differentiation-associated protein AHNAK |
| CD166 antigen ALCAM | Phosphoenolpyruvate carboxykinase [GTP], PCK2 |
| CD44 antigen CD44 | Pleckstrin homology domain-containing family A member 4 PLEKHA4 |
| CD59 glycoprotein CD59 | Pregnancy-specific beta-1-glycoprotein 6 PSG6 |
| Coiled-coil domain-containing protein 170 CCDC170 | Pyruvate carboxylase, mitochondrial PC |
| Core histone macro-H2A.2 H2AFY2 | Septin-11 SEPTIN11 |
| Cytochrome b5 reductase 4 CYB5R4 | Splicing regulatory glutamine/lysine-rich protein 1 SREK1 |
| Cytochrome P450 4F11 CYP4F11 | START domain-containing protein 10 STARD10 |
| D-3-phosphoglycerate dehydrogenase PHGDH | Twinfilin-1 TWF1 |
| DnaJ homolog subfamily C member 2 DNAJC2 | |
| Elongation factor 2 EEF2 | |
| Far upstream element-binding protein 3 FUBP3 | |
| Fatty acid synthase FASN | |
| Filamin-A FLNA | |
| Filamin-B FLNB | |
| Fructose-bisphosphate aldolase C ALDOC | |
| Galectin-1 LGALS1 | |
| Heat shock 70 kDa protein 1 (HSP70-1) | |
| Hepatoma-derived growth factor HDGF | |
| Inactive caspase-12 CASP12 | |
| Integrin alpha-V ITGAV | |
| Integrin beta-1 ITGB1 | |
| Keratin, type I cytoskeletal 15 KRT15 | |
| Keratin, type I cytoskeletal 18 KRT18 | |
| Keratin, type II cytoskeletal 6B KRT6B | |
| Keratin, type II cytoskeletal 7 KRT7 | |
| Keratin, type II cytoskeletal 8 KRT8 | |
| Kunitz-type protease inhibitor 1 SPINT1 | |
| Metalloproteinase inhibitor 2 TIMP2 | |
| Mitochondrial import receptor subunit TOM70 TOMM70 | |
| MORN repeat-containing protein 1 MORN1 | |
| Myosin light polypeptide 6 MYL6 | |
| Myosin-10 MYH10 | |
| Myosin-9 MYH9 | |
| NKAP-like protein NKAPL | |
| Nucleolin NCL | |
| Nucleoside diphosphate kinase A NME1 | |
| Pachytene checkpoint protein 2 homolog TRIP13 | |
| POTE ankyrin domain family member F POTEF | |
| Protein FAM118B FAM118B | |
| Protein-glutamine gamma-glutamyltransferase 2 TGM2 | |
| Putative Ras-related protein Rab-1C RAB1C | |
| Receptor tyrosine-protein kinase erbB-2 ERBB2 | |
| Serine/threonine-protein phosphatase 2A 65 kDa regulatory subunit A alpha isoform PPP2R1A | |
| Short-chain dehydrogenase/reductase 3 DHRS3 | |
| Sulfide:quinone oxidoreductase, mitochondrial SQOR | |
| T-box brain protein 1 TBR1 | |
| T-complex protein 1 subunit alpha TCP1 | |
| T-complex protein 1 subunit delta CCT4 | |
| T-complex protein 1 subunit epsilon CCT5 | |
| T-complex protein 1 subunit theta CCT8 | |
| T-complex protein 1 subunit zeta CCT6A | |
| Tripartite motif-containing protein 77 TRIM77 | |
| Tropomyosin alpha-3 chain TPM3 | |
| Tropomyosin alpha-4 chain TPM4 | |
| UTP–glucose-1-phosphate uridylyltransferase UGP2 | |
| Y-box-binding protein 3 YBX3 | |
| ZAR1-like protein ZAR1L |
2.5. Gene Ontology Analysis of Proteins Upregulated in HCC-1954 or MCF-7 ME Fractions Indicates the Cell Line Characteristics That Account for Their Phenotype Differences
2.6. Highly Expressed Proteins of HCC-1954 Could Be Associated with Cell Mechanobiology
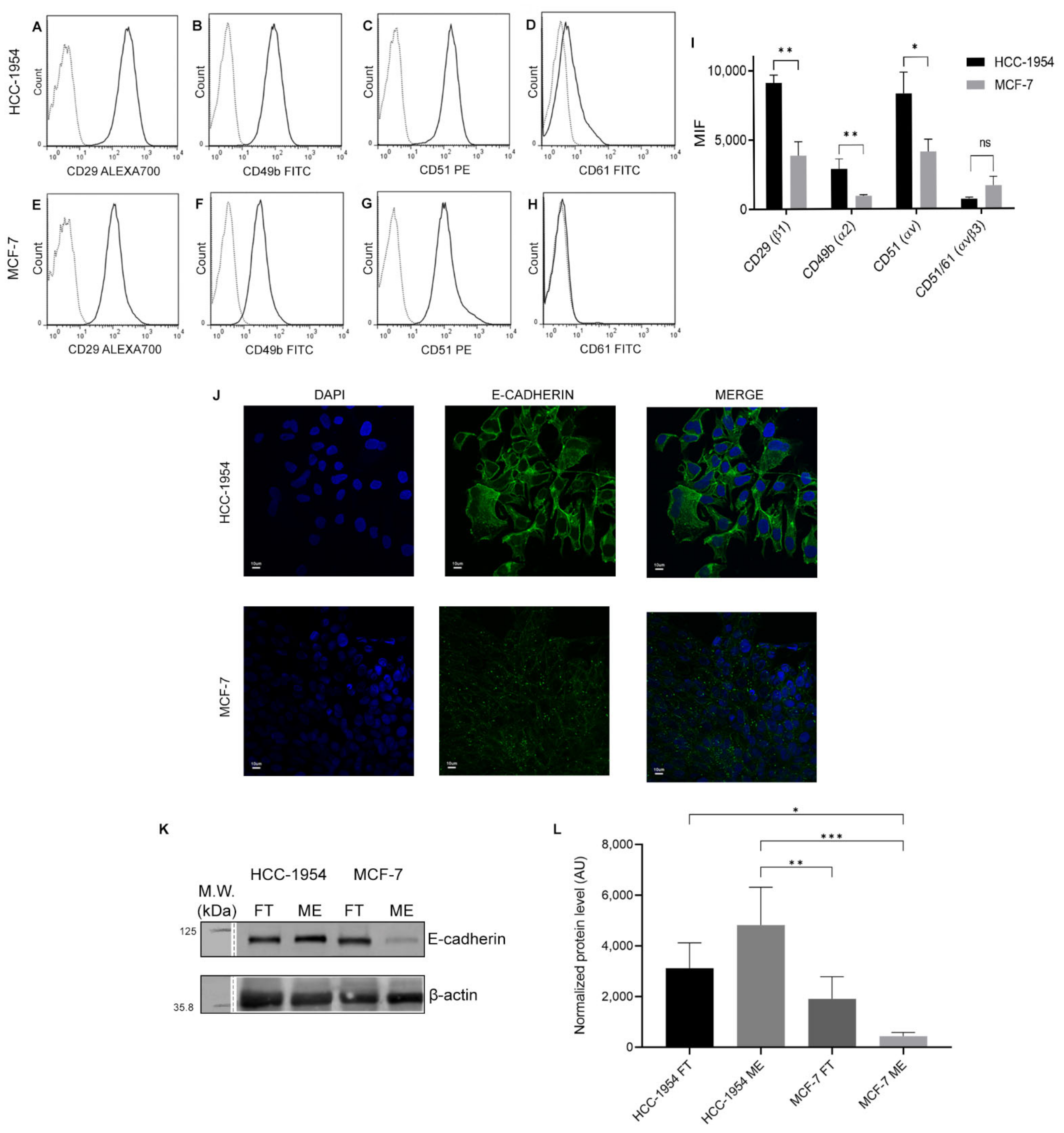
2.7. Integrin β1 and Cadherin-1 Expression in HCC-1954 and MCF-7 Cell Lines Are in Accordance with Cells’ Characteristic Phenotype

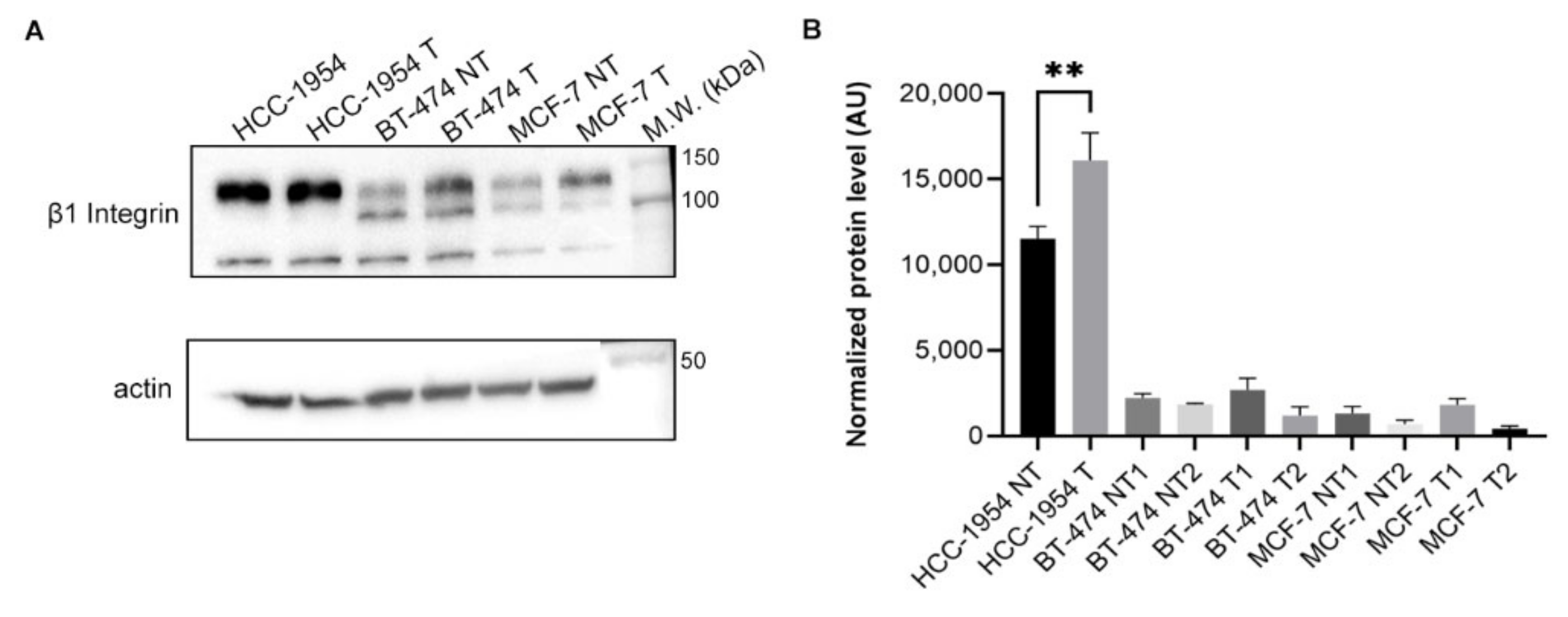
2.8. HCC-1954 Cell Line Presents Higher β1 Integrin Plasma Membrane Expression
2.9. β1 Integrin Presents a Significatively Higher Signal in the HCC-1954 Cell Line after Trastuzumab Treatment
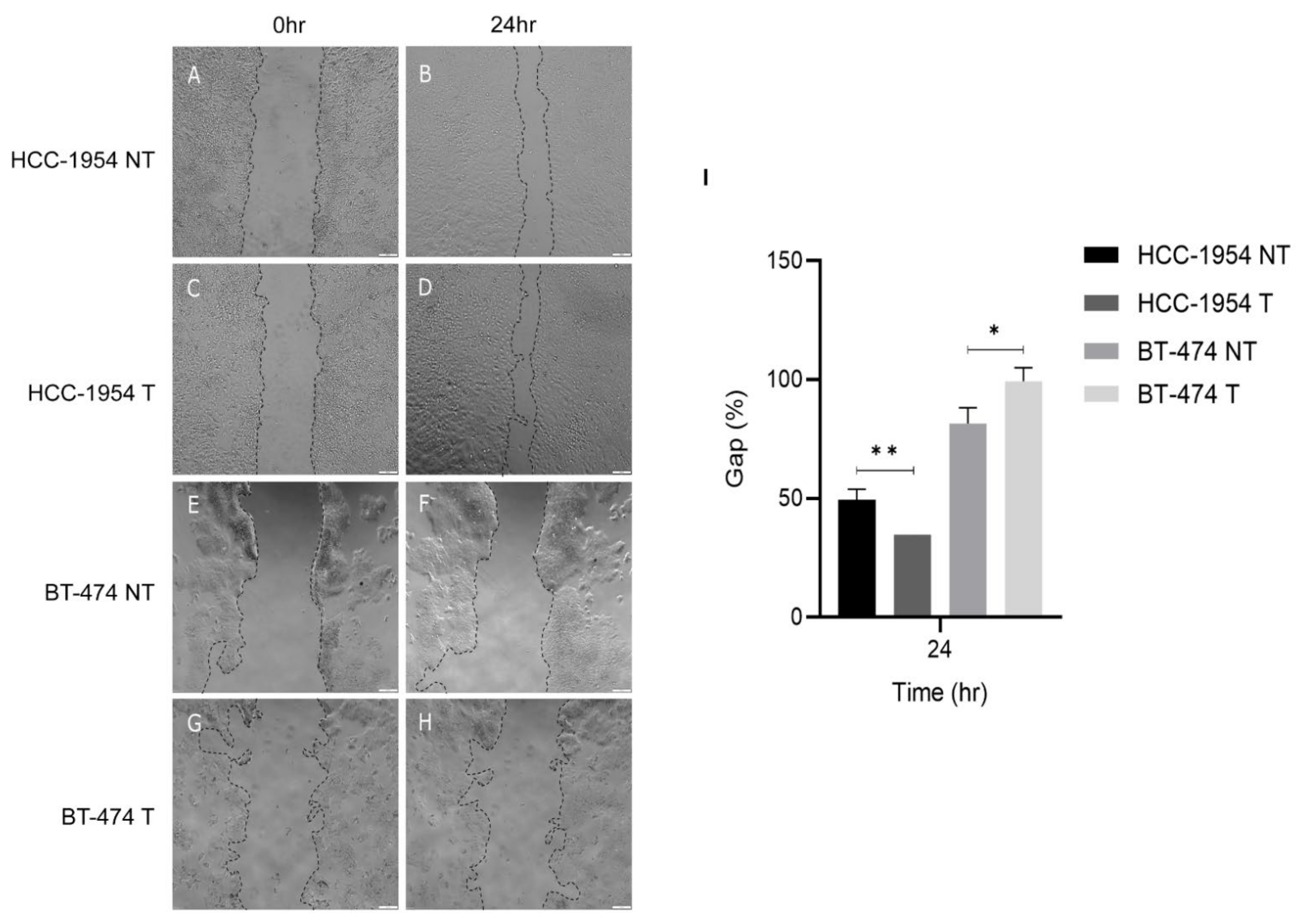
2.10. Trastuzumab Treatment Enhanced HCC-1954 Cell Migration
2.11. HER-2 and β1 Integrin Expression Presented a Trend in Correlation in HER-2+ but No Correlation in Luminal A Breast Cancer Subtypes
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Enrichment of Membrane Proteins, Quantification, and Protein Digestion
4.3. Peptide Desalting
4.4. Label-Free LC-MSE Analysis
4.5. Mass Spectra Data Analysis and Protein Quantification
4.6. Bioinformatic Analysis
4.7. Immunofluorescence
4.8. Flow Cytometry Analysis (FACS)
4.9. Western Blotting of HER-2 and E-Cadherin in Fractions Obtained from the Cell Surface Protein Isolation Kit from Pierce®
4.10. Western Blotting of β1 Integrin from Cell Lines Treated or Not with Trastuzumab and from Cell Line Fractions
4.11. Wound-Healing Assay
4.12. Gene Expression Correlation Analysis
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azamjah, N.; Soltan-Zadeh, Y.; Zayeri, F. Global trend of breast cancer mortality rate: A 25-year study. Asian Pac. J. Cancer Prev. 2019, 20, 2015–2020. [Google Scholar] [CrossRef]
- Zurrida, S.; Veronesi, U. Milestones in breast cancer treatment. Breast J. 2015, 21, 3–12. [Google Scholar] [CrossRef]
- Sørlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef]
- Eliyatkin, N.; Yalcin, E.; Zengel, B.; Aktaş, S.; Vardar, E. Molecular Classification of Breast Carcinoma: From Traditional, Old-Fashioned Way to A New Age, and A New Way. J. Breast Health 2015, 11, 59–66. [Google Scholar] [CrossRef] [PubMed]
- McCubrey, J.A.; Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Wong, E.W.T.; Chang, F.; Lehmann, B.; Terrian, D.M.; Milella, M.; Tafuri, A.; et al. Roles of the Raf/MEK/ERK pathway in cell growth, malignant transformation and drug resistance. Biochim. Biophys. Acta Mol. Cell Res. 2007, 1773, 1263–1284. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, A.; Grela-Wojewoda, A.; Domagała-Haduch, M.; Ambicka, A.; Harazin-Lechowska, A.; Janecka-Widła, A.; Cedrych, I.; Majchrzyk, K.; Kruczak, A.; Ryś, J.; et al. Proteins involved in HER2 signalling pathway, their relations and influence on metastasis-free survival in HER2-positive breast cancer patients treated with trastuzumab in adjuvant setting. J. Cancer 2017, 8, 131–139. [Google Scholar] [CrossRef]
- Lu, W.; Kang, Y. Epithelial-Mesenchymal Plasticity in Cancer Progression and Metastasis. Dev. Cell 2019, 49, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Kute, T.; Lack, C.M.; Willingham, M.; Bishwokama, B.; Williams, H.; Barrett, K.; Mitchell, T.; Vaughn, J.P. Development of Herceptin Resistance in Breast Cancer Cells. Cytom. Part A 2004, 57, 86–93. [Google Scholar] [CrossRef]
- Tóth, G.; Szöőr, Á.; Simon, L.; Yarden, Y.; Szöllősi, J.; Vereb, G. The combination of trastuzumab and pertuzumab administered at approved doses may delay development of trastuzumab resistance by additively enhancing antibody-dependent cell-mediated cytotoxicity. MAbs 2016, 8, 1361–1370. [Google Scholar] [CrossRef]
- Valabrega, G.; Montemurro, F.; Aglietta, M. Trastuzumab: Mechanism of action, resistance and future perspectives in HER2-overexpressing breast cancer. Ann. Oncol. 2007, 18, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Kunte, S.; Abraham, J.; Montero, A.J. Novel HER2–targeted therapies for HER2–positive metastatic breast cancer. Cancer 2020, 126, 4278–4288. [Google Scholar] [CrossRef]
- Henriques, B.; Mendes, F.; Martins, D. Immunotherapy in breast cancer: When, how, and what challenges? Biomedicines 2021, 9, 1687. [Google Scholar] [CrossRef]
- Padmanaban, V.; Krol, I.; Suhail, Y.; Szczerba, B.M.; Aceto, N.; Bader, J.S.; Ewald, A.J. E-cadherin is required for metastasis in multiple models of breast cancer. Nature 2019, 573, 439–444. [Google Scholar] [CrossRef]
- Canel, M.; Serrels, A.; Frame, M.C.; Brunton, V.G. E-cadherin-integrin crosstalk in cancer invasion and metastasis. J. Cell Sci. 2013, 126, 393–401. [Google Scholar] [CrossRef]
- Hamidi, H.; Pietilä, M.; Ivaska, J. The complexity of integrins in cancer and new scopes for therapeutic targeting. Br. J. Cancer 2016, 115, 1017–1023. [Google Scholar] [CrossRef]
- Nagano, M.; Hoshino, D.; Koshikawa, N.; Akizawa, T.; Seiki, M. Turnover of focal adhesions and cancer cell migration. Int. J. Cell Biol. 2012, 2012, 310616. [Google Scholar] [CrossRef] [PubMed]
- Popielarski, M.; Ponamarczuk, H.; Stasiak, M.; Watała, C.; Świątkowska, M. Modifications of disulfide bonds in breast cancer cell migration and invasiveness. Am. J. Cancer Res. 2019, 9, 1554–1582. [Google Scholar]
- Gazdar, A.F.; Kurvari, V.; Virmani, A.; Gollahon, L.; Sakaguchi, M.; Westerfield, M.; Kodagoda, D.; Stasny, V.; Cunningham, H.T.; Wistuba, I.I.; et al. Characterization of paired tumor and non-tumor cell lines established from patients with breast cancer. Int. J. Cancer 1998, 78, 766–774. [Google Scholar] [CrossRef]
- Bignell, G.R.; Santarius, T.; Pole, J.C.; Butler, A.P.; Perry, J.; Pleasance, E.; Greenman, C.; Menzies, A.; Taylor, S.; Edkins, S.; et al. Architectures of somatic genomic rearrangement in human cancer amplicons at sequence-level resolution. Genome Res. 2007, 17, 1296–1303. [Google Scholar] [CrossRef]
- Zhao, Q.; Caballero, O.L.; Levy, S.; Stevenson, B.J.; Iseli, C.; de Souza, S.J.; Galante, P.A.; Busam, D.; Leversha, M.A.; Chadalavada, K.; et al. Transcriptome-guided characterization of genomic rearrangements in a breast cancer cell line. Proc. Natl. Acad. Sci. USA 2009, 106, 1886–1891. [Google Scholar] [CrossRef]
- Soule, H.D.; Vazquez, J.; Long, A.; Albert, S.; Brennan, M. A human cell line from a pleural effusion derived from a breast carcinoma1,2. J. Natl. Cancer Inst. 1973, 51, 1409–1416. [Google Scholar] [CrossRef]
- Shim, J.S.; Rao, R.; Beebe, K.; Neckers, L.; Han, I.; Nahta, R.; Liu, J.O. Selective inhibition of HER2-positive breast cancer cells by the HIV protease inhibitor nelfinavir. J. Natl. Cancer Inst. 2012, 104, 1576–1590. [Google Scholar] [CrossRef]
- Su, Y.; Pogash, T.J.; Nguyen, T.D.; Russo, J. Development and characterization of two human triple-negative breast cancer cell lines with highly tumorigenic and metastatic capabilities. Cancer Med. 2016, 5, 558–573. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- McSherry, E.A.; Brennan, K.; Hudson, L.; Hill, A.D.K.; Hopkins, A.M. Breast cancer cell migration is regulated through junctional adhesion molecule-A-mediated activation of Rap1 GTPase. Breast Cancer Res. 2011, 13, R31. [Google Scholar] [CrossRef]
- Lawson, J.C.; Blatch, G.L.; Edkins, A.L. Cancer stem cells in breast cancer and metastasis. Breast Cancer Res. Treat. 2009, 118, 241–254. [Google Scholar] [CrossRef]
- Von der Heyde, S.; Wagner, S.; Czerny, A.; Nietert, M.; Ludewig, F.; Salinas-Riester, G.; Arlt, D.; Beißbarth, T. mRNA profiling reveals determinants of trastuzumab efficiency in HER2-positive breast cancer. PLoS ONE 2015, 10, e0117818. [Google Scholar] [CrossRef]
- Britton, K.M.; Kirby, J.A.; Lennard, T.W.J.; Meeson, A.P. Cancer stem cells and side population cells in breast cancer and metastasis. Cancers 2011, 3, 2106–2130. [Google Scholar] [CrossRef]
- Trapé, A.P.; Gonzalez-Angulo, A.M. Breast cancer and metastasis: On the way toward individualized therapy. Cancer Genom. Proteom. 2012, 9, 297–310. [Google Scholar]
- De Gagliato, D.M.; Jardim, D.L.F.; Marchesi, M.S.P.; Hortobagyi, G.N. Mechanisms of resistance and sensitivity to anti-HER2 therapies in HER2+ breast cancer. Oncotarget 2016, 7, 64431–64446. [Google Scholar] [CrossRef]
- Henjes, F.; Bender, C.; von der Heyde, S.; Braun, L.; Mannsperger, H.A.; Schmidt, C.; Wiemann, S.; Hasmann, M.; Aulmann, S.; Beissbarth, T.; et al. Strong EGFR signaling in cell line models of ERBB2-amplified breast cancer attenuates response towards ERBB2-targeting drugs. Oncogenesis 2012, 1, e16–e19. [Google Scholar] [CrossRef]
- Hampton, O.A.; Miller, C.A.; Koriabine, M.; Li, J.; Den Hollander, P.; Carbone, L.; Nefedov, M.; Ten Hallers, B.F.; Lee, A.V.; De Jong, P.J.; et al. Long-range massively parallel mate pair sequencing detects distinct mutations and similar patterns of structural mutability in two breast cancer cell lines. Cancer Genet. 2011, 204, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Asif, H.M.; Sultana, S.; Ahmed, S.; Akhtar, N.; Tariq, M. MINI-REVIEW HER-2 Positive Breast Cancer—A Mini-Review. Asian Pac. J. Cancer Prev. 2016, 17, 1609–1615. [Google Scholar] [CrossRef]
- Huang, C.; Park, C.C.; Hilsenbeck, S.G.; Ward, R.; Rimawi, M.F.; Wang, Y.-C.; Shou, J.; Bissell, M.J.; Osborne, C.K.; Schiff, R. Β1 integrin mediates an alternative survival pathway in breast cancer cells resistant to lapatinib. Breast Cancer Res. 2011, 13, R84. [Google Scholar] [CrossRef]
- Li, H.; Meng, F.; Jiang, L.; Ren, Y.; Qiu, Z.; Yu, P.; Peng, J. Comparison of LC-MS/MS-based targeted proteomics and conventional analytical methods for monitoring breast cancer resistance protein expression. Life Sci. 2019, 231, 116548. [Google Scholar] [CrossRef]
- Jiang, P.; Enomoto, A.; Takahashi, M. Cell biology of the movement of breast cancer cells: Intracellular signalling and the actin cytoskeleton. Cancer Lett. 2009, 284, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Escrivá-de-Romaní, S.; Arumí, M.; Bellet, M.; Saura, C. HER2-positive breast cancer: Current and new therapeutic strategies. Breast 2018, 39, 80–88. [Google Scholar] [CrossRef]
- Altundag, K. Her2 positive subtype and breast cancer brain metastasis: Any effect of anti-Her2 targeted therapy? J. Neuro-Oncol. 2017, 135, 639. [Google Scholar] [CrossRef]
- Kast, K.; Link, T.; Friedrich, K.; Petzold, A.; Niedostatek, A.; Schoffer, O.; Werner, C.; Klug, S.J.; Werner, A.; Gatzweiler, A.; et al. Impact of breast cancer subtypes and patterns of metastasis on outcome. Breast Cancer Res. Treat. 2015, 150, 621–629. [Google Scholar] [CrossRef]
- Toscani, A.M.; Sampayo, R.G.; Barabas, F.M.; Fuentes, F.; Simian, M.; Leskow, F.C. Distinct ErbB2 receptor populations differentially interact with beta1 integrin in breast cancer cell models. PLoS ONE 2017, 12, e0174230. [Google Scholar] [CrossRef]
- Lesniak, D.; Xu, Y.; Deschenes, J.; Lai, R.; Thoms, J.; Murray, D.; Gosh, S.; Mackey, J.R.; Sabri, S.; Abdulkarim, B. β1-integrin circumvents the antiproliferative effects of trastuzumab in human epidermal growth factor receptor-2-positive breast cancer. Cancer Res. 2009, 69, 8620–8628. [Google Scholar] [CrossRef]
- Le, A.V.P.; Szaumkessel, M.; Tan, T.Z.; Thiery, J.P.; Thompson, E.W.; Dobrovic, A. DNA methylation profiling of breast cancer cell lines along the epithelial mesenchymal spectrum—Implications for the choice of circulating tumour DNA methylation markers. Int. J. Mol. Sci. 2018, 19, 2553. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.Z.; Miow, Q.H.; Miki, Y.; Noda, T.; Mori, S.; Huang, R.Y.-J.; Thiery, J.P. Epithelial-mesenchymal transition spectrum quantification and its efficacy in deciphering survival and drug responses of cancer patients. EMBO Mol. Med. 2014, 6, 1279–1293. [Google Scholar] [CrossRef]
- Lee, G.; Han, S.B.; Lee, J.H.; Kim, H.W.; Kim, D.H. Cancer Mechanobiology: Microenvironmental Sensing and Metastasis. ACS Biomater. Sci. Eng. 2019, 5, 3735–3752. [Google Scholar] [CrossRef]
- Mendonsa, A.M.; Na, T.Y.; Gumbiner, B.M. E-cadherin in contact inhibition and cancer. Oncogene 2018, 37, 4769–4780. [Google Scholar] [CrossRef]
- Williams, E.D.; Gao, D.; Redfern, A.; Thompson, E.W. Controversies around epithelial–mesenchymal plasticity in cancer metastasis. Nat. Rev. Cancer 2019, 19, 716–732. [Google Scholar] [CrossRef]
- Matsui, C.; Takatani-Nakase, T.; Maeda, S.; Takahashi, K. High-glucose conditions promote anchorage-independent colony growth in human breast cancer MCF-7 cells. Biol. Pharm. Bull. 2018, 41, 1379–1383. [Google Scholar] [CrossRef]
- Darvishi, B.; Boroumandieh, S.; Majidzadeh, A.K.; Salehi, M.; Jafari, F.; Farahmand, L. The role of activated leukocyte cell adhesion molecule (ALCAM) in cancer progression, invasion, metastasis and recurrence: A novel cancer stem cell marker and tumor-specific prognostic marker. Exp. Mol. Pathol. 2020, 115, 104443. [Google Scholar] [CrossRef]
- Tachezy, M.; Zander, H.; Gebauer, F.; Marx, A.; Kaifi, J.T.; Izbicki, J.R.; Bockhorn, M. Activated leukocyte cell adhesion molecule (CD166)—Its prognostic power for colorectal cancer patients. J. Surg. Res. 2012, 177, e15–e20. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Mann, M. Combination of FASP and StageTip-based fractionation allows in-depth analysis of the hippocampal membrane proteome. J. Proteome Res. 2009, 8, 5674–5678. [Google Scholar] [CrossRef]
- Qi, D.; Brownridge, P.; Xia, D.; Mackay, K.; Gonzalez-Galarza, F.F.; Kenyani, J.; Harman, V.; Beynon, R.; Jones, A. A software toolkit and interface for performing stable isotope labeling and top3 quantification using progenesis LC-MS. OMICS A J. Integr. Biol. 2012, 16, 489–495. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]





| Gene 1 | Gene 2 | Spearman r | p Value | |
|---|---|---|---|---|
| Luminal A | ITGB1 | HER2 | −0.035 | 0.4201 |
| HER-2+ | ITGB1 | HER2 | 0.214 | 0.0617 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Abreu Pereira, D.; Sandim, V.; Fernandes, T.F.B.; Almeida, V.H.; Rocha, M.R.; do Amaral, R.J.F.C.; Rossi, M.I.D.; Kalume, D.E.; Zingali, R.B. Proteomic Analysis of HCC-1954 and MCF-7 Cell Lines Highlights Crosstalk between αv and β1 Integrins, E-Cadherin and HER-2. Int. J. Mol. Sci. 2022, 23, 10194. https://doi.org/10.3390/ijms231710194
de Abreu Pereira D, Sandim V, Fernandes TFB, Almeida VH, Rocha MR, do Amaral RJFC, Rossi MID, Kalume DE, Zingali RB. Proteomic Analysis of HCC-1954 and MCF-7 Cell Lines Highlights Crosstalk between αv and β1 Integrins, E-Cadherin and HER-2. International Journal of Molecular Sciences. 2022; 23(17):10194. https://doi.org/10.3390/ijms231710194
Chicago/Turabian Stylede Abreu Pereira, Denise, Vanessa Sandim, Thais F. B. Fernandes, Vitor Hugo Almeida, Murilo Ramos Rocha, Ronaldo J. F. C. do Amaral, Maria Isabel D. Rossi, Dário Eluan Kalume, and Russolina B. Zingali. 2022. "Proteomic Analysis of HCC-1954 and MCF-7 Cell Lines Highlights Crosstalk between αv and β1 Integrins, E-Cadherin and HER-2" International Journal of Molecular Sciences 23, no. 17: 10194. https://doi.org/10.3390/ijms231710194