
Extracellular Cysteines Are Critical to Form Functional Cx46 Hemichannels

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
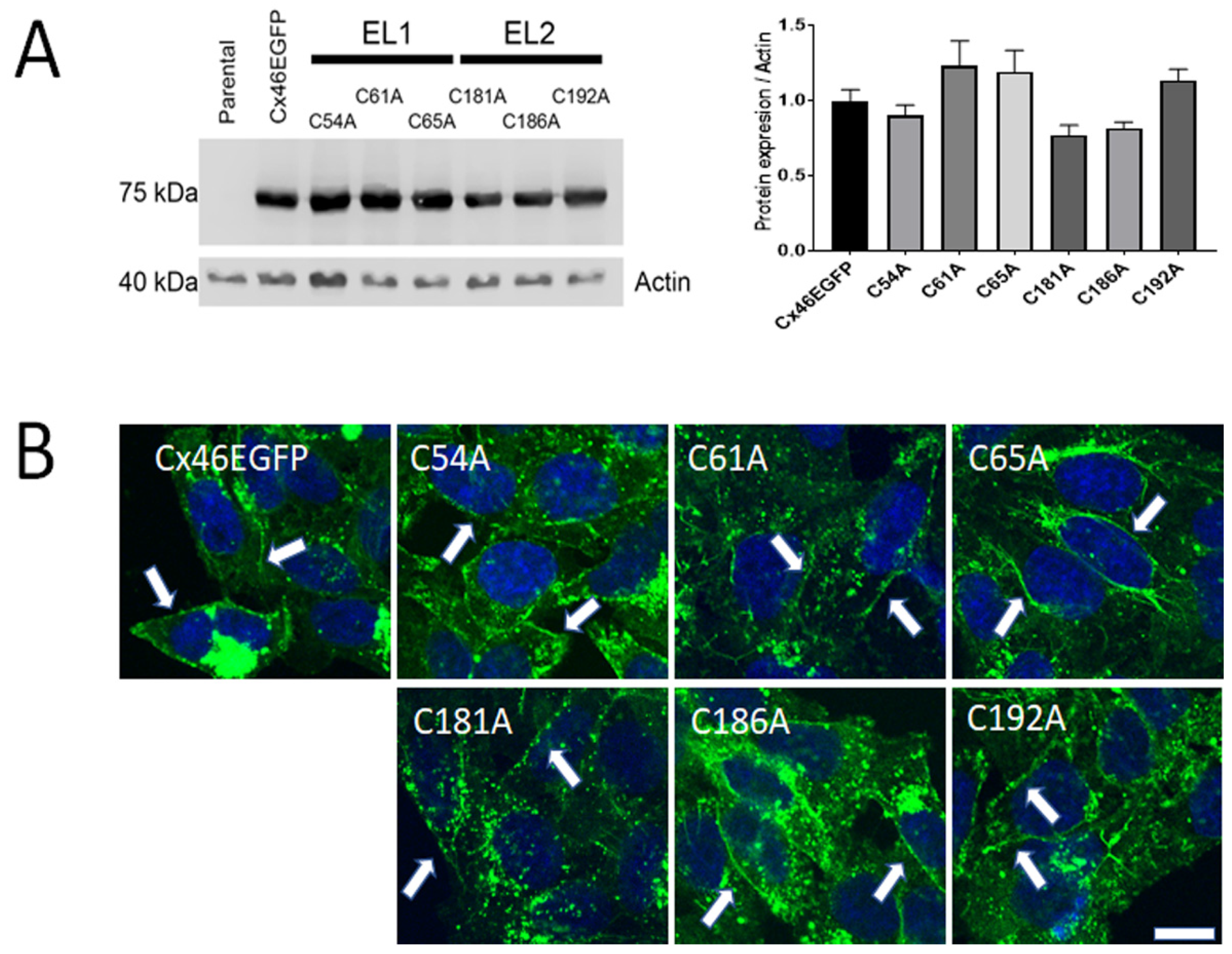
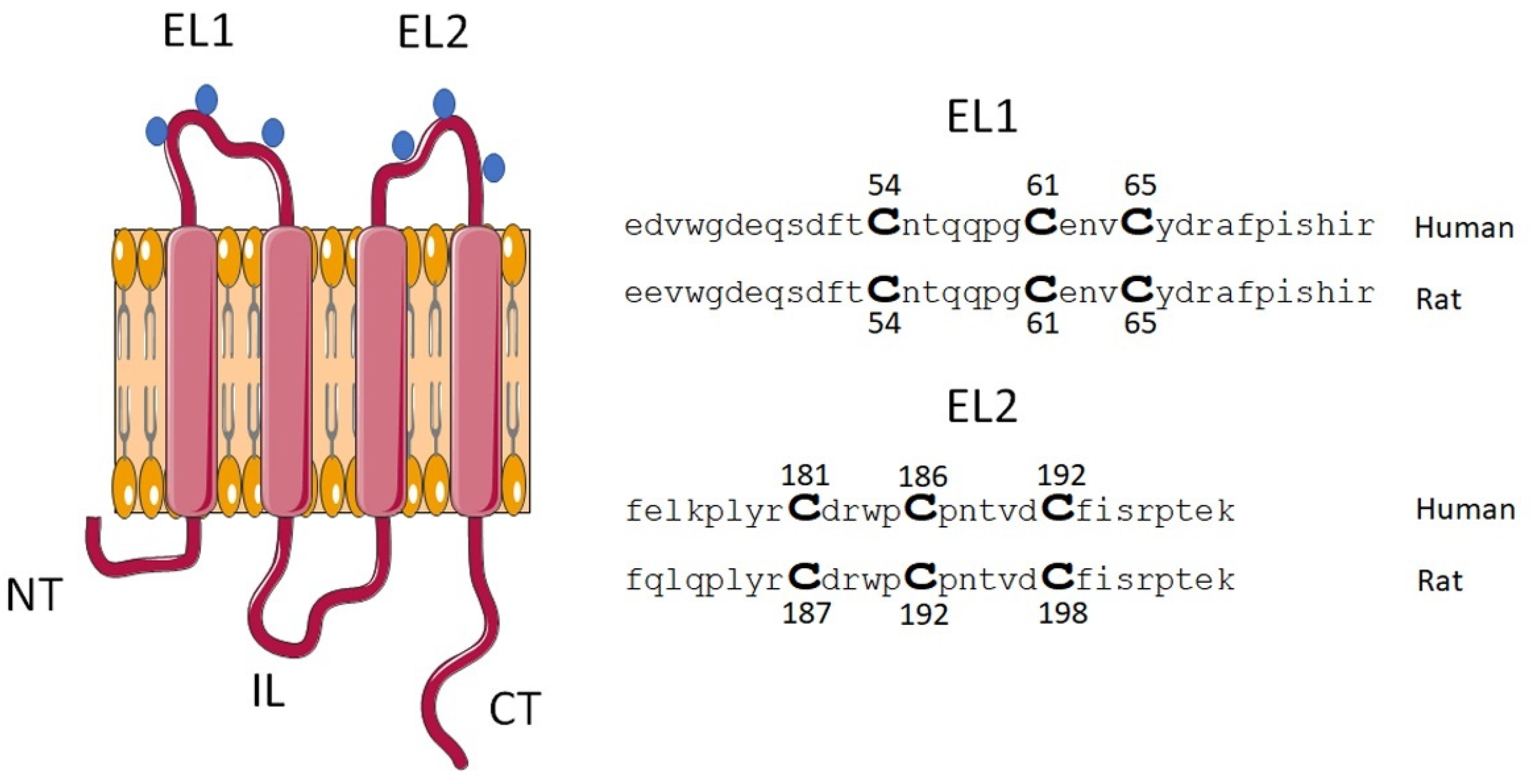
2.1. Mutations of Cx46 Extracellular Cys Do Not Alter Their Expression and Cellular Localization
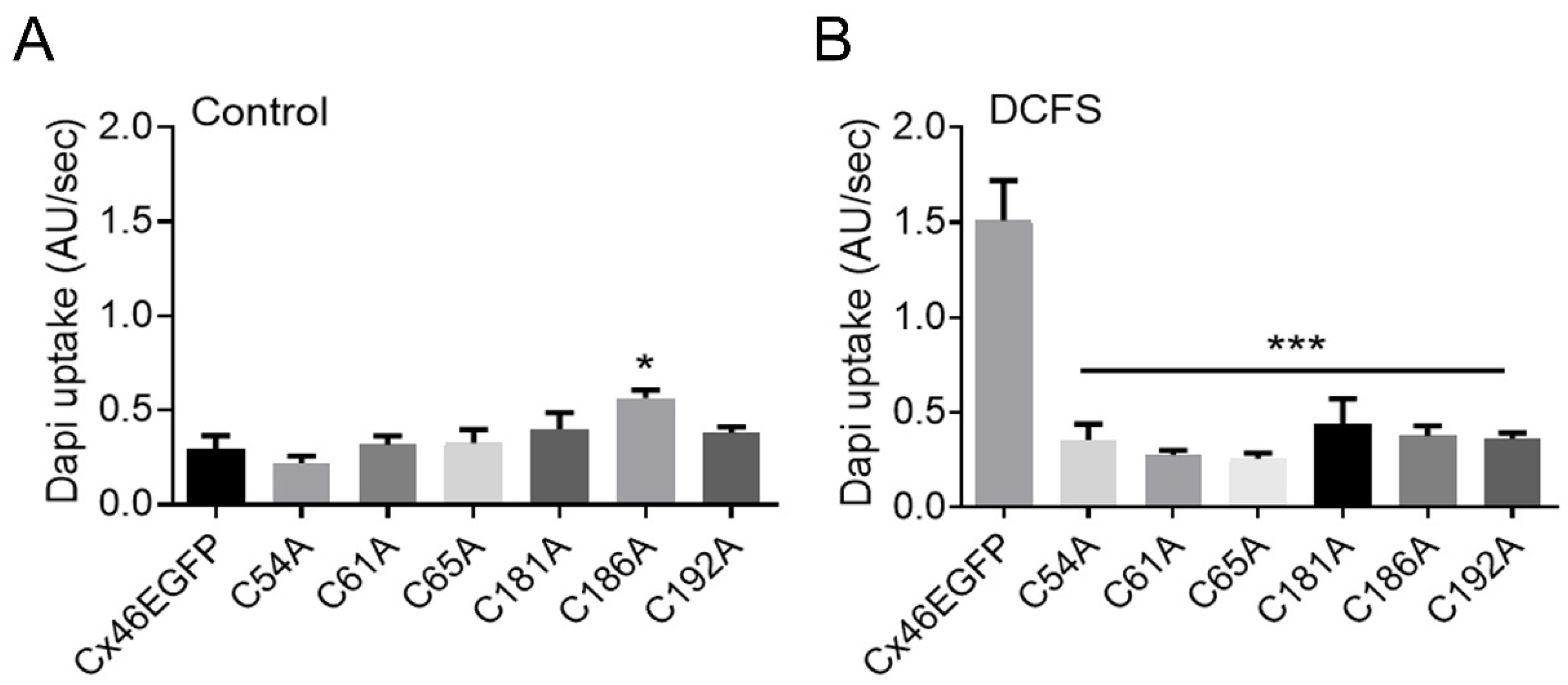
2.2. Mutation of Extracellular Cys of Cx46 Eliminates Hemichannel Sensitivity to Removal of Divalent Cations
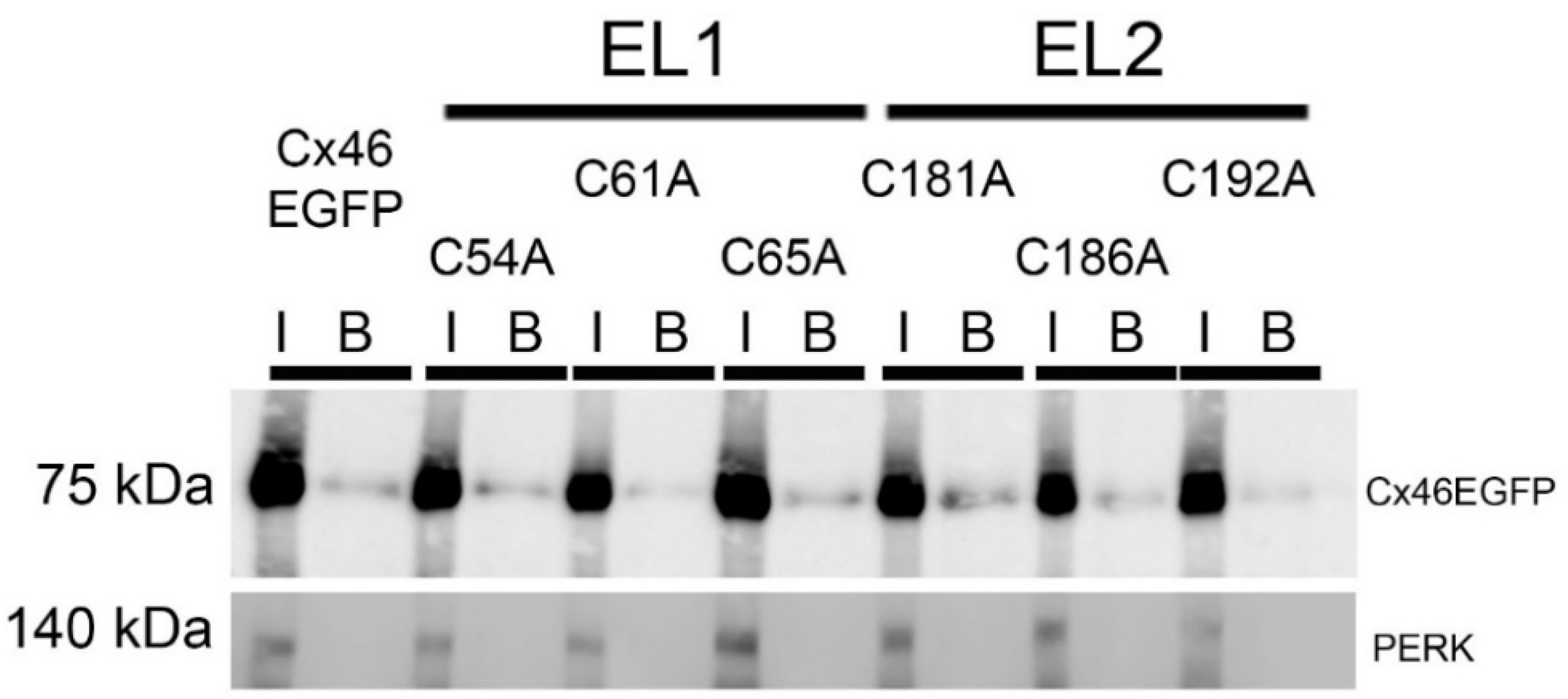
2.3. Extracellular Cys Mutants of Cx46 Are Targeted to the Plasma Membrane
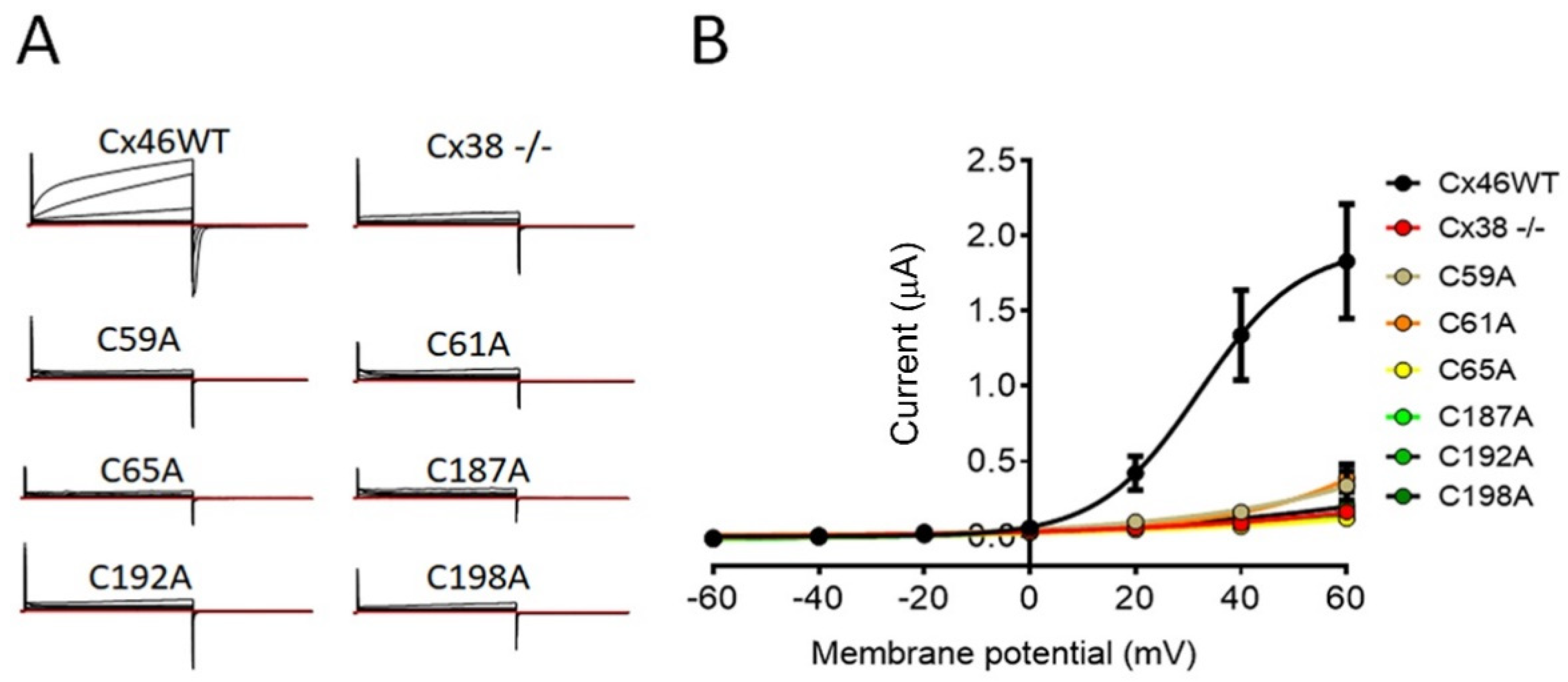
2.4. Mutation of Extracellular Cys of Cx46 Eliminates Hemichannel Sensitivity to Plasma-Membrane Depolarization
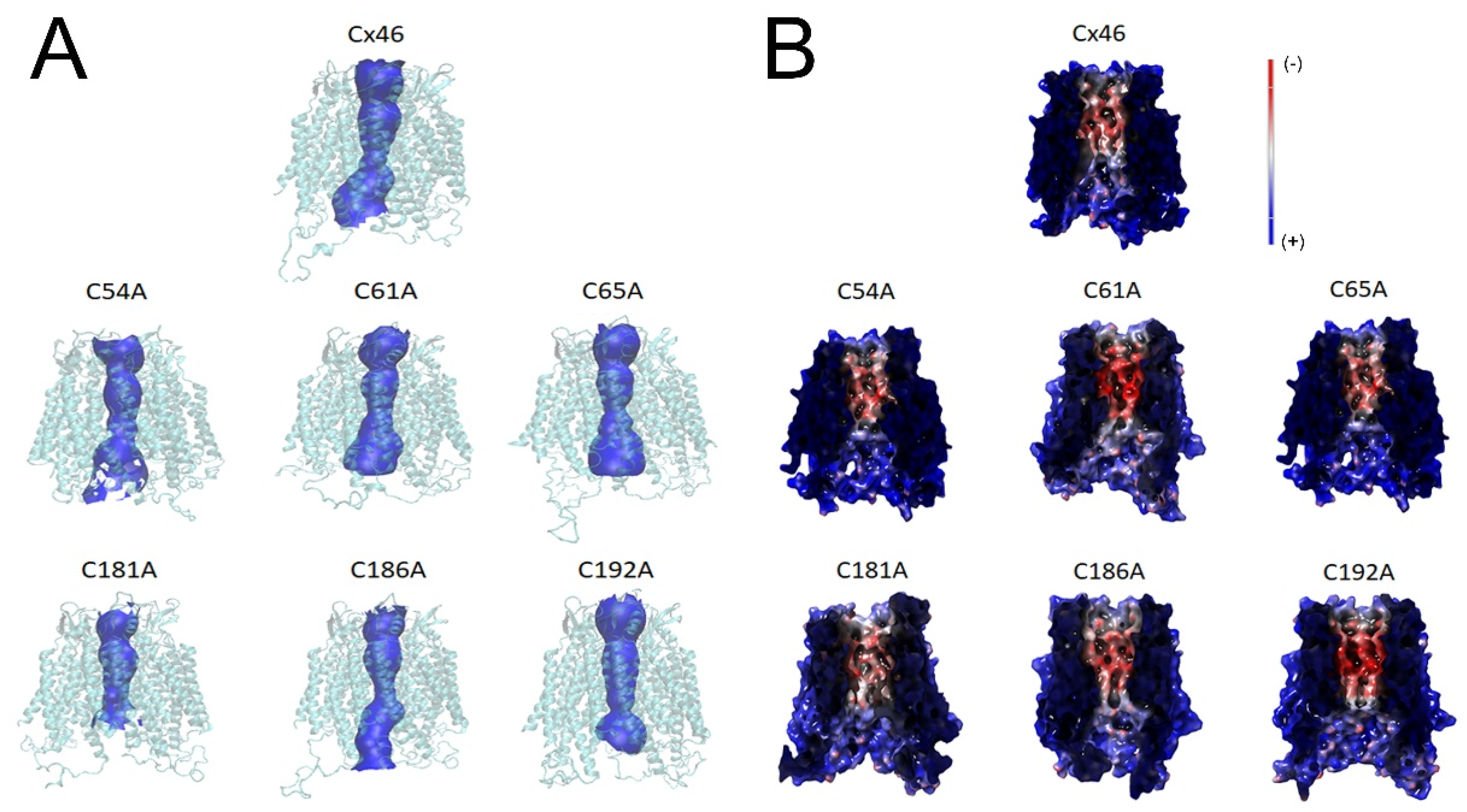
2.5. In Silico Modeling of Hemichannels Formed by Extracellular Cys Mutants of Cx46 Identify Small Changes in Pore Shape and Electrostatic Potential
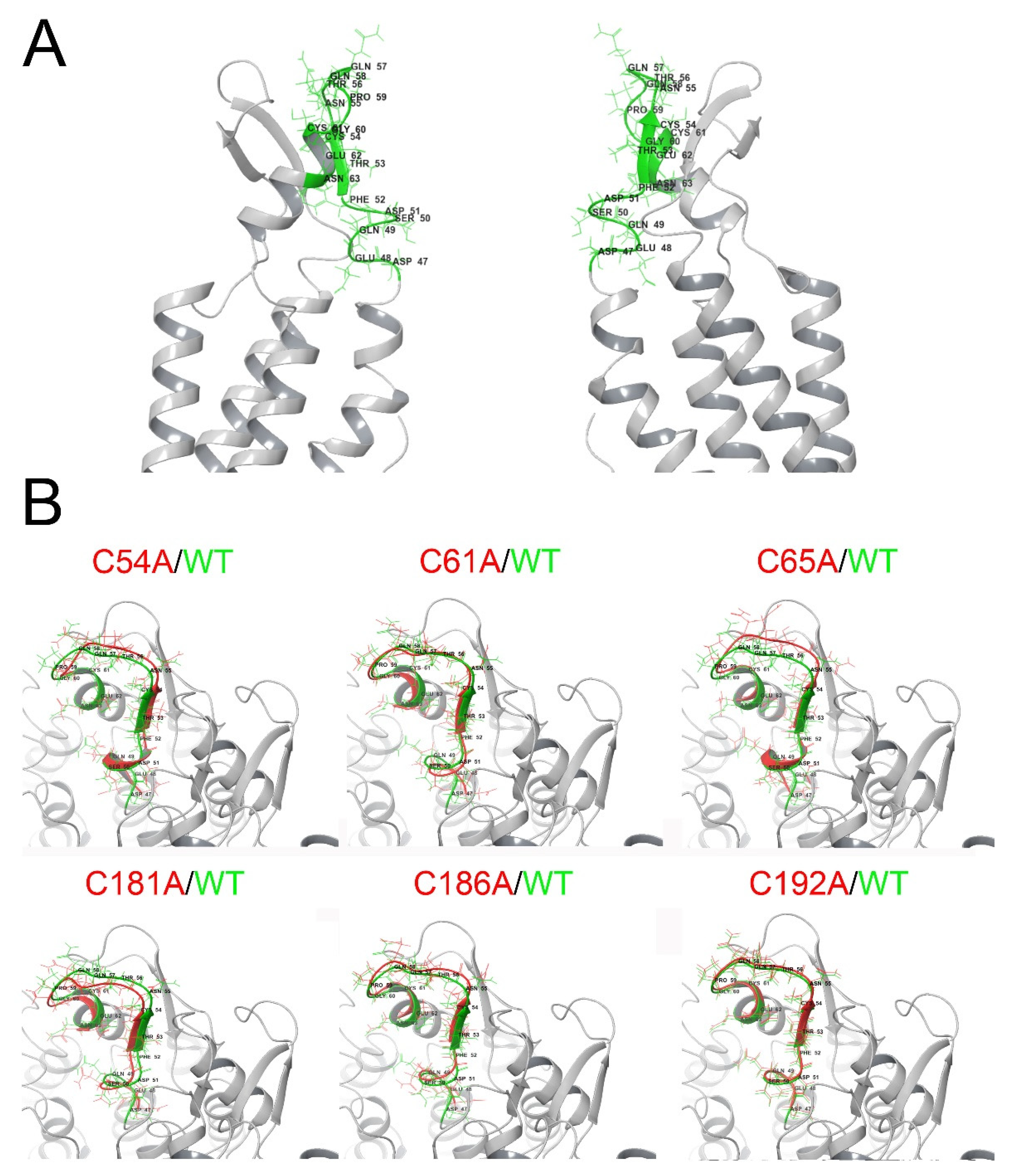
2.6. Extracellular Cys Mutants Display Changes in the Parahelix 47–61 Segment In Silico
3. Discussion
4. Materials and Methods
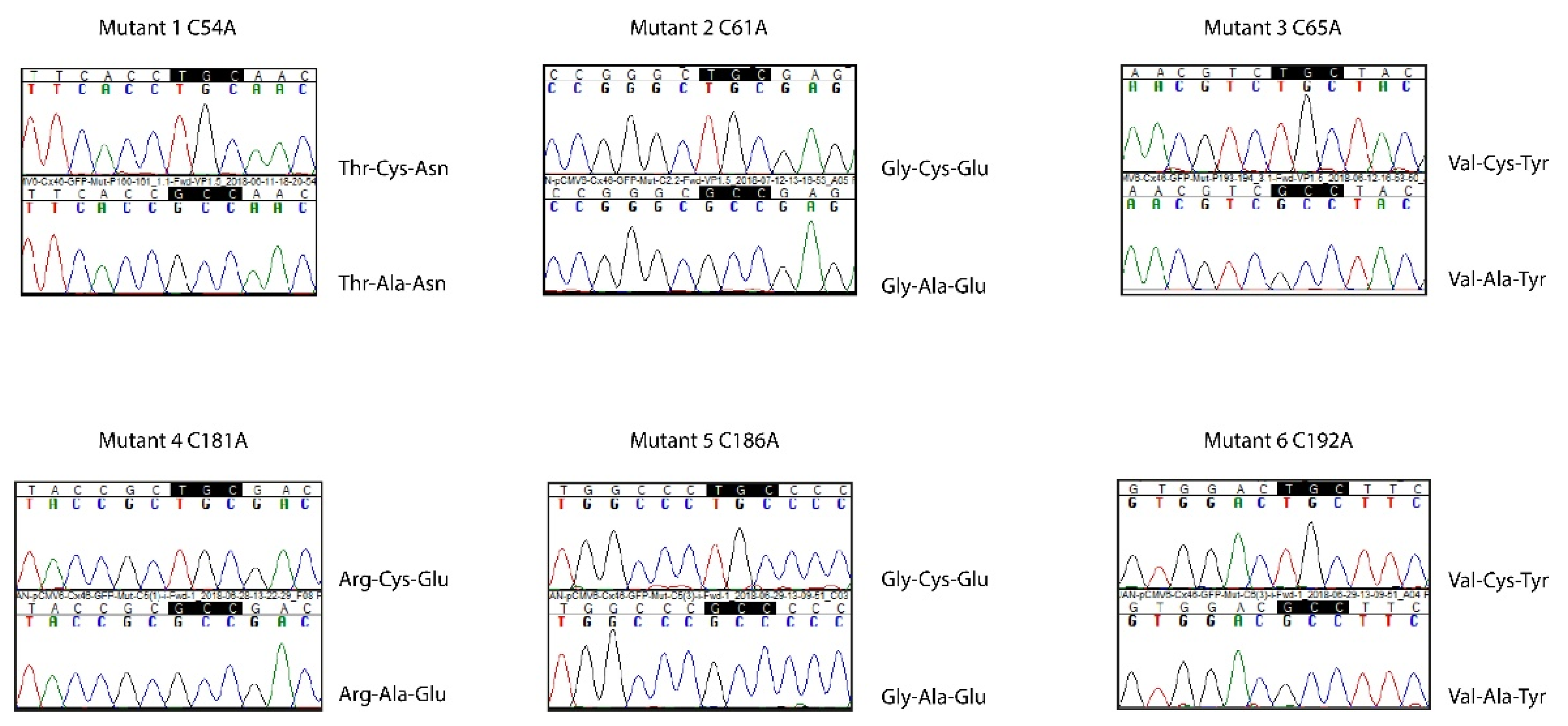
4.1. Plasmid Engineering
4.2. Xenopus laevis Oocytes Preparation
4.3. cRNA Preparation and Injection into Xenopus laevis Oocytes
4.4. Whole-Cell Electrophysiological Recordings
4.5. Transfection of HeLa Cells
4.6. Dye Uptake
4.7. Biotinylation
4.8. Western Blot Analysis
4.9. EGFP Cellular Localization
4.10. Molecular Dynamics
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sáez, J.C.; Retamal, M.; Basilio, D.; Bukauskas, F.F.; Bennett, M.V. Connexin-based gap junction hemichannels: Gating mechanisms. Biochim. Biophys. Acta (BBA)-Biomembr. 2005, 1711, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, X.-Q.; Nicholson, B.J. Size Selectivity between Gap Junction Channels Composed of Different Connexins. Cell Commun. Adhes. 2001, 8, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.; Haddad, B.G.; O’Neill, S.E.; Chorev, D.; Yoshioka, C.C.; Robinson, C.; Zuckerman, D.M.; Reichow, S.L. Structure of native lens connexin 46/50 intercellular channels by cryo-EM. Nature 2018, 564, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Nakagawa, S.; Suga, M.; Yamashita, E.; Oshima, A.; Fujiyoshi, Y.; Tsukihara, T. Structure of the connexin 26 gap junction channel at 3.5 Å resolution. Nature 2009, 458, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Sáez, J.C.; Schalper, K.A.; Retamal, M.; Orellana, J.A.; Shoji, K.F.; Bennett, M.V. Cell membrane permeabilization via connexin hemichannels in living and dying cells. Exp. Cell Res. 2010, 316, 2377–2389. [Google Scholar] [CrossRef]
- Stout, C.E.; Costantin, J.L.; Naus, C.C.G.; Charles, A.C. Intercellular Calcium Signaling in Astrocytes via ATP Release through Connexin Hemichannels. J. Biol. Chem. 2002, 277, 10482–10488. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.-C.; Wyeth, M.; Baltan-Tekkok, S.; Ransom, B.R. Functional Hemichannels in Astrocytes: A Novel Mechanism of Glutamate Release. J. Neurosci. 2003, 23, 3588–3596. [Google Scholar] [CrossRef] [Green Version]
- Tong, X.; Lopez, W.; Ramachandran, J.; Ayad, W.A.; Liu, Y.; Lopez-Rodriguez, A.; Harris, A.L.; Contreras, J.E. Glutathione release through connexin hemichannels: Implications for chemical modification of pores permeable to large molecules. J. Gen. Physiol. 2015, 146, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Rana, S.; Dringen, R. Gap junction hemichannel-mediated release of glutathione from cultured rat astrocytes. Neurosci. Lett. 2007, 415, 45–48. [Google Scholar] [CrossRef]
- Franco, L.; Zocchi, E.; Usai, C.; Guida, L.; Bruzzone, S.; Costa, A.; De Flora, A. Paracrine roles of NAD+ and cyclic ADP-ribose in increasing intracellular calcium and enhancing cell proliferation of 3T3 fibroblasts. J. Biol. Chem. 2001, 276, 21642–21648. [Google Scholar] [CrossRef] [Green Version]
- Retamal, M.A.; Froger, N.; Palacios-Prado, N.; Ezan, P.; Sáez, P.J.; Sáez, J.C.; Giaume, C. Cx43 Hemichannels and Gap Junction Channels in Astrocytes Are Regulated Oppositely by Proinflammatory Cytokines Released from Activated Microglia. J. Neurosci. 2007, 27, 13781–13792. [Google Scholar] [CrossRef] [PubMed]
- Schalper, K.A.; Sánchez, H.A.; Lee, S.C.; Altenberg, G.; Nathanson, M.H.; Sáez, J.C. Connexin 43 hemichannels mediate the Ca2+ influx induced by extracellular alkalinization. Am. J. Physiol. Physiol. 2010, 299, C1504–C1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linsambarth, S.; Carvajal, F.J.; Moraga-Amaro, R.; Mendez, L.; Tamburini, G.; Jimenez, I.; Verdugo, D.A.; Gómez, G.I.; Jury, N.; Martínez, P.; et al. Astroglial gliotransmitters released via Cx43 hemichannels regulate NMDAR-dependent transmission and short-term fear memory in the basolateral amygdala. FASEB J. 2022, 36, e22134. [Google Scholar] [CrossRef] [PubMed]
- Stehberg, J.; Moraga-Amaro, R.; Salazar, C.; Becerra, A.; Echeverría, C.; Orellana, J.A.; Bultynck, G.; Ponsaerts, R.; Leybaert, L.; Simon, F.; et al. Release of gliotransmitters through astroglial connexin 43 hemichannels is necessary for fear memory consolidation in the basolateral amygdala. FASEB J. 2012, 26, 3649–3657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orellana, J.A. Physiological Functions of Glial Cell Hemichannels. In Glial Cells in Health and Disease of the CNS; von Bernhardi, R., Ed.; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2016; Volume 949, pp. 93–108. [Google Scholar] [CrossRef]
- Kamermans, M.; Fahrenfort, I.; Schultz, K.; Janssen-Bienhold, U.; Sjoerdsma, T.; Weiler, R. Hemichannel-Mediated Inhibition in the Outer Retina. Science 2001, 292, 1178–1180. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, M.; Xue, T.; Li, G.; Wang, D.; Shang, P.; Jiang, J.X.; Xu, H. Osteocytic connexin 43 channels affect fracture healing. J. Cell. Physiol. 2019, 234, 19824–19832. [Google Scholar] [CrossRef]
- Ma, L.; Hua, R.; Tian, Y.; Cheng, H.; Fajardo, R.J.; Pearson, J.J.; Guda, T.; Shropshire, D.B.; Gu, S.; Jiang, J.X. Connexin 43 hemichannels protect bone loss during estrogen deficiency. Bone Res. 2019, 7, 11. [Google Scholar] [CrossRef]
- Roux, L.; Madar, A.; Lacroix, M.M.; Yi, C.; Benchenane, K.; Giaume, C. Astroglial Connexin 43 Hemichannels Modulate Olfactory Bulb Slow Oscillations. J. Neurosci. 2015, 35, 15339–15352. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Wang, Y.-Q.; Xu, C.-F.; Li, Y.-N.; Guo, R.; Li, L. Involvement of connexin43 in the acute hyperosmotic stimulus-induced synthesis and release of vasopressin in the supraoptic nucleus of rats. Mol. Med. Rep. 2014, 10, 2165–2171. [Google Scholar] [CrossRef] [Green Version]
- Dospinescu, V.-M.; Nijjar, S.; Spanos, F.; Cook, J.; De Wolf, E.; Biscotti, M.A.; Gerdol, M.; Dale, N. Structural determinants of CO2-sensitivity in the β connexin family suggested by evolutionary analysis. Commun. Biol. 2019, 2, 331. [Google Scholar] [CrossRef] [Green Version]
- Contreras, J.E.; Sáez, J.C.; Bukauskas, F.F.; Bennett, M.V.L. Gating and regulation of connexin 43 (Cx43) hemichannels. Proc. Natl. Acad. Sci. USA 2003, 100, 11388–11393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Retamal, M.A.; Reyes, E.P.; García, I.E.; Pinto, B.; Martínez, A.D.; González, C. Diseases associated with leaky hemichannels. Front. Cell. Neurosci. 2015, 9, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Retamal, M.A.; Fernandez-Olivares, A.; Stehberg, J. Over-activated hemichannels: A possible therapeutic target for human diseases. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2021, 1867, 166232. [Google Scholar] [CrossRef] [PubMed]
- Retamal, M.A.; Cortés, C.J.; Reuss, L.; Bennett, M.V.L.; Sáez, J.C. S-nitrosylation and permeation through connexin 43 hemichannels in astrocytes: Induction by oxidant stress and reversal by reducing agents. Proc. Natl. Acad. Sci. USA 2006, 103, 4475–4480. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, L.; Zhang, L.; Chen, B.; Yang, L.; Li, X.; Li, Y.; Yu, H. Inhibition of Connexin 43 Hemichannels Alleviates Cerebral Ischemia/Reperfusion Injury via the TLR4 Signaling Pathway. Front. Cell. Neurosci. 2018, 12, 372. [Google Scholar] [CrossRef] [Green Version]
- Stong, B.C.; Chang, Q.; Ahmad, S.; Lin, X. A Novel Mechanism for Connexin 26 Mutation Linked Deafness: Cell Death Caused by Leaky Gap Junction Hemichannels. Laryngoscope 2006, 116, 2205–2210. [Google Scholar] [CrossRef]
- Liang, G.S.L.; De Miguel, M.; Glass, J.D.; Scherer, S.S.; Mintz, M.; Barrio, L.C.; Fischbeck, K.H.; Gómez-Hernández, J.M. Severe neuropathy with leaky connexin32 hemichannels. Ann. Neurol. 2005, 57, 749–754. [Google Scholar] [CrossRef]
- Sánchez, H.A.; Meşe, G.; Srinivas, M.; White, T.W.; Verselis, V.K. Differentially altered Ca2+ regulation and Ca2+ permeability in Cx26 hemichannels formed by the A40V and G45E mutations that cause keratitis ichthyosis deafness syndrome. J. Gen. Physiol. 2010, 136, 47–62. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hao, H. Conserved glycine at position 45 of major cochlear connexins constitutes a vital component of the Ca2+ sensor for gating of gap junction hemichannels. Biochem. Biophys. Res. Commun. 2013, 436, 424–429. [Google Scholar] [CrossRef]
- Press, E.R.; Shao, Q.; Kelly, J.J.; Chin, K.; Alaga, A.; Laird, D.W. Induction of cell death and gain-of-function properties of connexin26 mutants predict severity of skin disorders and hearing loss. J. Biol. Chem. 2017, 292, 9721–9732. [Google Scholar] [CrossRef] [Green Version]
- Chi, J.; Li, L.; Liu, M.; Tan, J.; Tang, C.; Pan, Q.; Wang, D.; Zhang, Z. Pathogenic Connexin-31 Forms Constitutively Active Hemichannels to Promote Necrotic Cell Death. PLoS ONE 2012, 7, e32531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasciani, I.; Temperán, A.; Pérez-Atencio, L.F.; Escudero, A.; Martínez-Montero, P.; Molano, J.; Gómez-Hernández, J.M.; Paino, C.; Nieto, D.G.; Barrio, L.C. Regulation of connexin hemichannel activity by membrane potential and the extracellular calcium in health and disease. Neuropharmacology 2013, 75, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Ripps, H.; Qian, H.; Zakevicius, J. Properties of Connexin26 Hemichannels Expressed in Xenopus Oocytes. Cell. Mol. Neurobiol. 2004, 24, 647–665. [Google Scholar] [CrossRef] [PubMed]
- González, D.; Gómez-Hernández, J.M.; Barrio, L.C. Species specificity of mammalian connexin-26 to form open voltage-gated hemichannels. FASEB J. 2006, 20, 2329–2338. [Google Scholar] [CrossRef]
- Sánchez, H.A.; Orellana, J.A.; Verselis, V.K.; Sáez, J.C. Metabolic inhibition increases activity of connexin-32 hemichannels permeable to Ca2+ in transfected HeLa cells. Am. J. Physiol. Physiol. 2009, 297, C665–C678. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Hernández, J.M.; de Miguel, M.; Larrosa, B.; González, D.; Barrio, L.C. Molecular basis of calcium regulation in connexin-32 hemichannels. Proc. Natl. Acad. Sci. USA 2003, 100, 16030–16035. [Google Scholar] [CrossRef] [Green Version]
- Retamal, M.A.; Schalper, K.A.; Shoji, K.F.; Bennett, M.V.L.; Sáez, J.C. Opening of connexin 43 hemichannels is increased by lowering intracellular redox potential. Proc. Natl. Acad. Sci. USA 2007, 104, 8322–8327. [Google Scholar] [CrossRef] [Green Version]
- Valiunas, V. Biophysical Properties of Connexin-45 Gap Junction Hemichannels Studied in Vertebrate Cells. J. Gen. Physiol. 2002, 119, 147–164. [Google Scholar] [CrossRef] [Green Version]
- Retamal, M.A.; Evangelista-Martínez, F.; León-Paravic, C.G.; Altenberg, G.A.; Reuss, L. Biphasic effect of linoleic acid on connexin 46 hemichannels. Pflügers Arch.-Eur. J. Physiol. 2011, 461, 635–643. [Google Scholar] [CrossRef] [Green Version]
- Ebihara, L.; Korzyukov, Y.; Kothari, S.; Tong, J.-J. Cx46 hemichannels contribute to the sodium leak conductance in lens fiber cells. Am. J. Physiol. Physiol. 2014, 306, C506–C513. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, T.F.; Lazrak, A.; Peracchia, C.; Goldberg, G.S.; Lampe, P.D.; Johnson, R.G. Properties and regulation of gap junctional hemichannels in the plasma membranes of cultured cells. J. Cell Biol. 1996, 134, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Pfahnl, A.; Dahl, G. Gating of cx46 gap junction hemichannels by calcium and voltage. Pflügers Arch. 1999, 437, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, L.; Liu, X.; Pal, J.D. Effect of External Magnesium and Calcium on Human Connexin46 Hemichannels. Biophys. J. 2003, 84, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Pogoda, K.; Kameritsch, P.; Retamal, M.A.; Vega, J.L. Regulation of gap junction channels and hemichannels by phosphorylation and redox changes: A revision. BMC Cell Biol. 2016, 17 (Suppl. 1), 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alstrøm, J.S.; Hansen, D.B.; Nielsen, M.S.; MacAulay, N. Isoform-specific phosphorylation-dependent regulation of connexin hemichannels. J. Neurophysiol. 2015, 114, 3014–3022. [Google Scholar] [CrossRef] [Green Version]
- Fiori, M.C.; Reuss, L.; Cuello, L.G.; Altenberg, G.A. Functional analysis and regulation of purified connexin hemichannels. Front. Physiol. 2014, 5, 71. [Google Scholar] [CrossRef] [Green Version]
- Retamal, M.A. Connexin and Pannexin hemichannels are regulated by redox potential. Front. Physiol. 2014, 5, 80. [Google Scholar] [CrossRef] [Green Version]
- García, I.E.; Sánchez, H.A.; Martínez, A.D.; Retamal, M.A. Redox-mediated regulation of connexin proteins; focus on nitric oxide. Biochim. Biophys. Acta (BBA)-Biomembr. 2018, 1860, 91–95. [Google Scholar] [CrossRef]
- León-Paravic, C.G.; Figueroa, V.A.; Guzmán, D.J.; Valderrama, C.F.; Vallejos, A.A.; Fiori, M.C.; Altenberg, G.; Reuss, L.; Retamal, M.A. Carbon Monoxide (CO) Is a Novel Inhibitor of Connexin Hemichannels. J. Biol. Chem. 2014, 289, 36150–36157. [Google Scholar] [CrossRef] [Green Version]
- Retamal, M.A.; Orellana, V.P.; Arévalo, N.J.; Rojas, C.G.; Arjona, R.J.; Alcaíno, C.A.; González, W.; Canan, J.G.; Moraga-Amaro, R.; Stehberg, J.; et al. Cx46 hemichannel modulation by nitric oxide: Role of the fourth transmembrane helix cysteine and its possible involvement in cataract formation. Nitric Oxide 2019, 86, 54–62. [Google Scholar] [CrossRef]
- Straub, A.C.; Billaud, M.; Johnstone, S.R.; Best, A.K.; Yemen, S.; Dwyer, S.T.; Looft-Wilson, R.; Lysiak, J.J.; Gaston, B.; Palmer, L.; et al. Compartmentalized Connexin 43 S-Nitrosylation/Denitrosylation Regulates Heterocellular Communication in the Vessel Wall. Arter. Thromb. Vasc. Biol. 2011, 31, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lillo, M.; Himelman, E.; Shirokova, N.; Xie, L.-H.; Fraidenraich, D.; Contreras, J.E. S-nitrosylation of connexin43 hemichannels elicits cardiac stress–induced arrhythmias in Duchenne muscular dystrophy mice. JCI Insight 2019, 4, e130091. [Google Scholar] [CrossRef] [PubMed]
- Retamal, M.A.; García, I.E.; Pinto, B.I.; Pupo, A.; Báez, D.; Stehberg, J.; Del Rio, R.; González, C. Extracellular Cysteine in Connexins: Role as Redox Sensors. Front. Physiol. 2016, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Retamal, M.A.; Fiori, M.C.; Fernandez-Olivares, A.; Linsambarth, S.; Peña, F.; Quintana, D.; Stehberg, J.; Altenberg, G.A. 4-Hydroxynonenal induces Cx46 hemichannel inhibition through its carbonylation. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2020, 1865, 158705. [Google Scholar] [CrossRef]
- Retamal, M.A.; Yin, S.; Altenberg, G.A.; Reuss, L. Voltage-dependent facilitation of Cx46 hemichannels. Am. J. Physiol. Physiol. 2010, 298, C132–C139. [Google Scholar] [CrossRef]
- Bennett, B.C.; Purdy, M.D.; Baker, K.A.; Acharya, C.; McIntire, W.E.; Stevens, R.C.; Zhang, Q.; Harris, A.L.; Abagyan, R.; Yeager, M. An electrostatic mechanism for Ca2+-mediated regulation of gap junction channels. Nat. Commun. 2016, 7, 8770. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, B.S.; Zonta, F.; Farkas, T.; Litman, T.; Nielsen, M.S.; MacAulay, N. Structural determinants underlying permeant discrimination of the Cx43 hemichannel. J. Biol. Chem. 2019, 294, 16789–16803. [Google Scholar] [CrossRef]
- Flores, J.A.; Haddad, B.G.; Dolan, K.A.; Myers, J.B.; Yoshioka, C.C.; Copperman, J.; Zuckerman, D.M.; Reichow, S.L. Connexin-46/50 in a dynamic lipid environment resolved by CryoEM at 1.9 Å. Nat. Commun. 2020, 11, 4331. [Google Scholar] [CrossRef]
- Bao, X.; Chen, Y.; Reuss, L.; Altenberg, G. Functional Expression in Xenopus Oocytes of Gap-junctional Hemichannels Formed by a Cysteine-less Connexin 43. J. Biol. Chem. 2004, 279, 9689–9692. [Google Scholar] [CrossRef] [Green Version]
- Koval, M.; Harley, J.E.; Hick, E.; Steinberg, T.H. Connexin46 Is Retained as Monomers in a trans-Golgi Compartment of Osteoblastic Cells. J. Cell Biol. 1997, 137, 847–857. [Google Scholar] [CrossRef] [Green Version]
- Dahl, G.; Levine, E.; Rabadan-Diehl, C.; Werner, R. Cell/cell channel formation involves disulfide exchange. JBIC J. Biol. Inorg. Chem. 1991, 197, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Foote, C.I.; Zhou, L.; Zhu, X.; Nicholson, B. The Pattern of Disulfide Linkages in the Extracellular Loop Regions of Connexin 32 Suggests a Model for the Docking Interface of Gap Junctions. J. Cell Biol. 1998, 140, 1187–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Héja, L.; Simon, Á.; Szabó, Z.; Kardos, J. Connexons Coupling to Gap Junction Channel: Potential Role for Extracellular Protein Stabilization Centers. Biomolecules 2021, 12, 49. [Google Scholar] [CrossRef]
- Iovine, M.K.; Gumpert, A.M.; Falk, M.M.; Mendelson, T.C. Cx23, a connexin with only four extracellular-loop cysteines, forms functional gap junction channels and hemichannels. FEBS Lett. 2008, 582, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Good, M.E.; Ek-Vitorín, J.F.; Burt, J.M. Extracellular loop cysteine mutant of cx37 fails to suppress proliferation of rat insulinoma cells. J. Membr. Biol. 2012, 245, 369–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | 5′ to 3′ Sequence |
|---|---|
| Cx46-C54A Forward | cagtcagacttcaccgccaacacccagcagcc |
| Cx46-C54A Reverse | ggctgctgggtgttggcggtgaagtctgactg |
| Cx46-C61A Forward | ccagcagccgggcgccgagaacgtctgc |
| Cx46-C61A Reverse | gcagacgttctcggcgcccggctgctgg |
| Cx46-C65A Forward | gctgcgagaacgtcgcctacgacagggcct |
| Cx46-C65A Reverse | aggccctgtcgtaggcgacgttctcgcagc |
| Cx46-C181A Forward | ccgctctaccgcgccgaccgctggcc |
| Cx46-C181A Reverse | ggccagcggtcggcgcggtagagcgg |
| Cx46-C186A Forward | cgaccgctggcccgcccccaacacggtg |
| Cx46-C186A Reverse | caccgtgttgggggcgggccagcggtcg |
| Cx46-C192A Forward | cccaacacggtggacgccttcatctccaggcc |
| Cx46-C192A Reverse | ggcctggagatgaaggcgtccaccgtgttggg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Olivares, A.; Durán-Jara, E.; Verdugo, D.A.; Fiori, M.C.; Altenberg, G.A.; Stehberg, J.; Alfaro, I.; Calderón, J.F.; Retamal, M.A. Extracellular Cysteines Are Critical to Form Functional Cx46 Hemichannels. Int. J. Mol. Sci. 2022, 23, 7252. https://doi.org/10.3390/ijms23137252
Fernández-Olivares A, Durán-Jara E, Verdugo DA, Fiori MC, Altenberg GA, Stehberg J, Alfaro I, Calderón JF, Retamal MA. Extracellular Cysteines Are Critical to Form Functional Cx46 Hemichannels. International Journal of Molecular Sciences. 2022; 23(13):7252. https://doi.org/10.3390/ijms23137252
Chicago/Turabian StyleFernández-Olivares, Ainoa, Eduardo Durán-Jara, Daniel A. Verdugo, Mariana C. Fiori, Guillermo A. Altenberg, Jimmy Stehberg, Iván Alfaro, Juan Francisco Calderón, and Mauricio A. Retamal. 2022. "Extracellular Cysteines Are Critical to Form Functional Cx46 Hemichannels" International Journal of Molecular Sciences 23, no. 13: 7252. https://doi.org/10.3390/ijms23137252