
The Structural and the Functional Aspects of Intercellular Communication in iPSC-Cardiomyocytes


{kind=link}
Abstract
:1. Introduction
2. Why iPSC-Cardiomyocytes?
3. iPSC-Cardiomyocytes for Cell Replacement Therapy in the Diseased Post-MI Heart
4. Gap Junctions and Electrical Signal Propagation
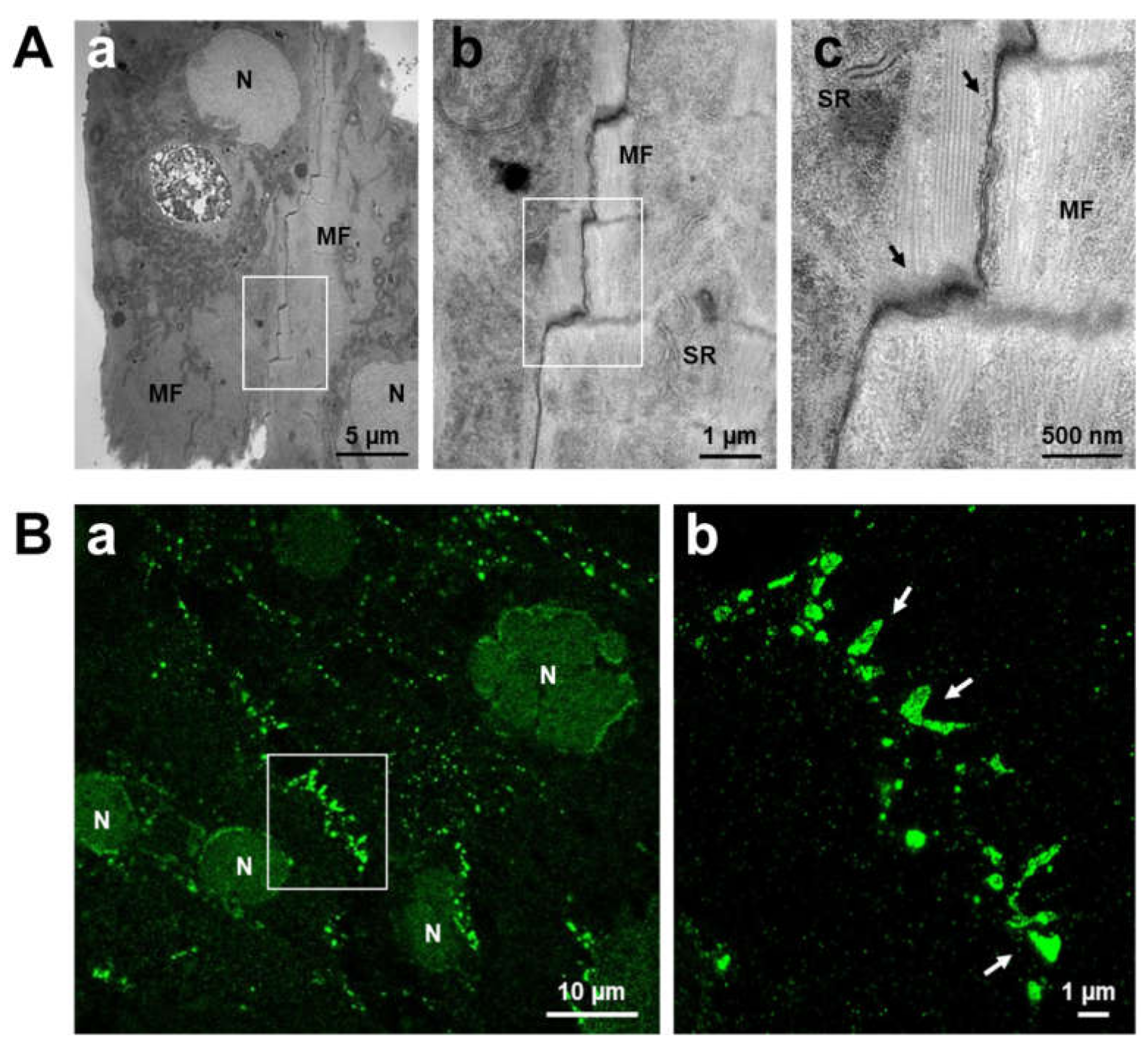
5. Intercalated Discs
6. Gap Junctions in iPSC-Cardiomyocytes
7. Improvement of Gap Junction Formation and Intercellular Coupling in iPSC-CMs
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carpenter, L.; Carr, C.; Yang, C.T.; Stuckey, D.J.; Clarke, K.; Watt, S.M. Efficient Differentiation of Human Induced Pluripotent Stem Cells Generates Cardiac Cells That Provide Protection Following Myocardial Infarction in the Rat. Stem Cells Dev. 2012, 21, 977–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fishman, G.I. Connexins and the Heart. Trends Cardiovasc. Med. 1992, 2, 50–55. [Google Scholar] [CrossRef]
- Kléber, A.G.; Jin, Q. Coupling between Cardiac Cells—An Important Determinant of Electrical Impulse Propagation and Arrhythmogenesis. Biophys. Rev. 2021, 2, 031301. [Google Scholar] [CrossRef] [PubMed]
- Jalife, J.; Morley, G.E.; Vaidya, D. Connexins and Impulse Propagation in the Mouse Heart. J. Cardiovasc. Electrophysiol. 1999, 10, 1649–1663. [Google Scholar] [CrossRef]
- Rohr, S. Role of Gap Junctions in the Propagation of the Cardiac Action Potential. Cardiovasc. Res. 2004, 62, 309–322. [Google Scholar] [CrossRef]
- Oxford, E.M.; Musa, H.; Maass, K.; Coombs, W.; Taffet, S.M.; Delmar, M. Connexin43 Remodeling Caused by Inhibition of Plakophilin-2 Expression in Cardiac Cells. Circ. Res. 2007, 101, 703–711. [Google Scholar] [CrossRef] [Green Version]
- Patel, D.M.; Dubash, A.D.; Kreitzer, G.; Green, K.J. Disease Mutations in Desmoplakin Inhibit Cx43 Membrane Targeting Mediated by Desmoplakin-EB1 Interactions. J. Cell Biol. 2014, 206, 779–797. [Google Scholar] [CrossRef] [Green Version]
- Vreeker, A.; Van Stuijvenberg, L.; Hund, T.J.; Mohler, P.J.; Nikkels, P.G.J.; Van Veen, T.A.B. Assembly of the Cardiac Intercalated Disk during Preand Postnatal Development of the Human Heart. PLoS ONE 2014, 9, e94722. [Google Scholar] [CrossRef]
- McKay, R. Stem Cells—Hype and Hope. Nature 2000, 406, 361–364. [Google Scholar] [CrossRef]
- Mummery, C.; Ward, D.; Van Den Brink, C.E.; Bird, S.D.; Doevendans, P.A.; Opthof, T.; Brutel De La Riviere, A.; Tertoolen, L.; Van Der Heyden, M.; Pera, M. Cardiomyocyte Differentiation of Mouse and Human Embryonic Stem Cells. J. Anat. 2002, 200, 233–242. [Google Scholar] [CrossRef]
- Beltrami, A.P.; Barlucchi, L.; Torella, D.; Baker, M.; Limana, F.; Chimenti, S.; Kasahara, H.; Rota, M.; Musso, E.; Urbanek, K.; et al. Adult Cardiac Stem Cells Are Multipotent and Support Myocardial Regeneration. Cell 2003, 114, 763–776. [Google Scholar] [CrossRef] [Green Version]
- Kehat, I.; Kenyagin-Karsenti, D.; Snir, M.; Segev, H.; Amit, M.; Gepstein, A.; Livne, E.; Binah, O.; Itskovitz-Eldor, J.; Gepstein, L. Human Embryonic Stem Cells Can Differentiate into Myocytes with Structural and Functional Properties of Cardiomyocytes. J. Clin. Investig. 2001, 108, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Katarzyna, R. Adult Stem Cell Therapy for Cardiac Repair in Patients After Acute Myocardial Infarction Leading to Ischemic Heart Failure: An Overview of Evidence from the Recent Clinical Trials. Curr. Cardiol. Rev. 2017, 13, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Habib, M.; Caspi, O.; Gepstein, L. Human Embryonic Stem Cells for Cardiomyogenesis. J. Mol. Cell. Cardiol. 2008, 45, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.J.; Kaufman, M.H. Establishment in Culture of Pluripotential Cells from Mouse Embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Rong, Z.; Wang, M.; Hu, Z.; Stradner, M.; Zhu, S.; Kong, H.; Yi, H.; Goldrath, A.; Yang, Y.G.; Xu, Y.; et al. An Effective Approach to Prevent Immune Rejection of Human ESC-Derived Allografts. Cell Stem Cell 2014, 14, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Orlic, D.; Kajstura, J.; Chimenti, S.; Jakoniuk, I.; Anderson, S.M.; Li, B.; Pickel, J.; McKay, R.; Nadal-Ginard, B.; Bodine, D.M.; et al. Bone Marrow Cells Regenerate Infarcted Myocardium. Nature 2001, 410, 701–705. [Google Scholar] [CrossRef]
- Krishna, K.; Krishna, K.; Berrocal, R.; Rao, K.; Rao, K.R.S. Myocardial Infarction and Stem Cells. J. Pharm. Bioallied Sci. 2011, 3, 182–188. [Google Scholar]
- Zomer, H.D.; Vidane, A.S.; Gonçalves, N.N.; Ambrósio, C.E. Mesenchymal and Induced Pluripotent Stem Cells: General Insights and Clinical Perspectives. Stem Cells Cloning Adv. Appl. 2015, 8, 125–134. [Google Scholar] [CrossRef]
- Messina, E.; De Angelis, L.; Frati, G.; Morrone, S.; Chimenti, S.; Fiordaliso, F.; Salio, M.; Battaglia, M.; Latronico, M.V.G.; Coletta, M.; et al. Isolation and Expansion of Adult Cardiac Stem Cells from Human and Murine Heart. Circ. Res. 2004, 95, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Malliaras, K.; Kreke, M.; Marbán, E. The Stuttering Progress of Cell Therapy for Heart Disease. Clin. Pharmacol. Ther. 2011, 90, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mummery, C.L.; Zhang, J.; Ng, E.S.; Elliott, D.A.; Elefanty, A.G.; Kamp, T.J. Differentiation of Human Embryonic Stem Cells and Induced Pluripotent Stem Cells to Cardiomyocytes: A Methods Overview. Circ. Res. 2012, 111, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Talkhabi, M.; Aghdami, N.; Baharvand, H. Human Cardiomyocyte Generation from Pluripotent Stem Cells: A State-of-Art. Life Sci. 2016, 145, 98–113. [Google Scholar] [CrossRef]
- Lian, X.; Hsiao, C.; Wilson, G.; Zhu, K.; Hazeltine, L.B.; Azarin, S.M.; Raval, K.K.; Zhang, J.; Kamp, T.J.; Palecek, S.P. Robust Cardiomyocyte Differentiation from Human Pluripotent Stem Cells via Temporal Modulation of Canonical Wnt Signaling. Proc. Natl. Acad. Sci. USA 2012, 109, E1848–E1857. [Google Scholar] [CrossRef] [Green Version]
- Kattman, S.J.; Koonce, C.H.; Swanson, B.J.; Anson, B.D. Stem Cells and Their Derivatives: A Renaissance in Cardiovascular Translational Research. J. Cardiovasc. Transl. Res. 2011, 4, 66–72. [Google Scholar] [CrossRef]
- Karakikes, I.; Ameen, M.; Termglinchan, V.; Wu, J.C. Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes: Insights into Molecular, Cellular, and Functional Phenotypes. Circ. Res. 2015, 117, 80–88. [Google Scholar] [CrossRef] [Green Version]
- Freund, C.; Mummery, C.L. Prospects for Pluripotent Stem Cell-Derived Cardiomyocytes in Cardiac Cell Therapy and as Disease Models. J. Cell. Biochem. 2009, 107, 592–599. [Google Scholar] [CrossRef]
- Okano, H.; Nakamura, M.; Yoshida, K.; Okada, Y.; Tsuji, O.; Nori, S.; Ikeda, E.; Yamanaka, S.; Miura, K. Steps toward Safe Cell Therapy Using Induced Pluripotent Stem Cells. Circ. Res. 2013, 112, 523–533. [Google Scholar] [CrossRef] [Green Version]
- Yoshihara, M.; Hayashizaki, Y.; Murakawa, Y. Genomic Instability of IPSCs: Challenges Towards Their Clinical Applications. Stem Cell Rev. Rep. 2017, 13, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Garreta, E.; Sanchez, S.; Lajara, J.; Montserrat, N.; Belmonte, J.C.I. Roadblocks in the Path of IPSC to the Clinic. Curr. Transplant. Rep. 2018, 5, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Zhang, Z.N.; Rong, Z.; Xu, Y. Immunogenicity of Induced Pluripotent Stem Cells. Nature 2011, 474, 212–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki, R.; Uda, M.; Hoki, Y.; Sunayama, M.; Nakamura, M.; Ando, S.; Sugiura, M.; Ideno, H.; Shimada, A.; Nifuji, A.; et al. Negligible Immunogenicity of Terminally Differentiated Cells Derived from Induced Pluripotent or Embryonic Stem Cells. Nature 2013, 494, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Morizane, A.; Kikuchi, T.; Hayashi, T.; Mizuma, H.; Takara, S.; Doi, H.; Mawatari, A.; Glasser, M.F.; Shiina, T.; Ishigaki, H.; et al. MHC Matching Improves Engraftment of IPSC-Derived Neurons in Non-Human Primates. Nat. Commun. 2017, 8, 385. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, R.; Wada, H.; Murata, T.; Seino, K.I. Immune Reaction and Regulation in Transplantation Based on Pluripotent Stem Cell Technology. Inflamm. Regen. 2020, 40, 12. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, J.J.; Sanchez, P.; Repetti, G.G.; Juneman, E.; Pandey, A.C.; Chinyere, I.R.; Moukabary, T.; LaHood, N.; Daugherty, S.L.; Goldman, S. Human Induced Pluripotent Stem Cell-Derived Cardiomyocyte Patch in Rats With Heart Failure. Ann. Thorac. Surg. 2019, 108, 1169–1177. [Google Scholar] [CrossRef] [PubMed]
- Citro, L.; Naidu, S.; Hassan, F.; Kuppusamy, M.L.; Kuppusamy, P.; Angelos, M.G.; Khan, M. Comparison of Human Induced Pluripotent Stem-Cell Derived Cardiomyocytes with Human Mesenchymal Stem Cells Following Acute Myocardial Infarction. PLoS ONE 2014, 9, e116281. [Google Scholar] [CrossRef] [Green Version]
- Hao, M.; Wang, R.; Wang, W. Cell Therapies in Cardiomyopathy: Current Status of Clinical Trials. Anal. Cell. Pathol. 2017, 2017, 9404057. [Google Scholar] [CrossRef] [Green Version]
- Swirski, F.K.; Nahrendorf, M. Leukocyte Behavior in Atherosclerosis, Myocardial Infarction, and Heart Failure. Science 2013, 339, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, Y.; Aguilar, E.G.; Troncoso, M.; Ilatovskaya, D.V.; DeLeon-Pennell, K.Y. Immune Regulation of Cardiac Fibrosis Post Myocardial Infarction. Cell. Signal. 2021, 77, 109837. [Google Scholar] [CrossRef]
- Hitscherich, P.; Lee, E.J. Crosstalk between Cardiac Cells and Macrophages Postmyocardial Infarction: Insights from in Vitro Studies. Tissue Eng.-Part B Rev. 2021, 27, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tu, H.; Wang, C.; Cao, L.; Muelleman, R.L.; Wadman, M.C.; Li, Y.L. Correlation of Ventricular Arrhythmogenesis with Neuronal Remodeling of Cardiac Postganglionic Parasympathetic Neurons in the Late Stage of Heart Failure after Myocardial Infarction. Front. Neurosci. 2017, 11, 252. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.S.; Chen, L.S.; Cao, J.M.; Sharifi, B.; Karagueuzian, H.S.; Fishbein, M.C. Sympathetic Nerve Sprouting, Electrical Remodeling and the Mechanisms of Sudden Cardiac Death. Cardiovasc. Res. 2001, 50, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Vaseghi, M. The Autonomic Nervous System and Ventricular Arrhythmias in Myocardial Infarction and Heart Failure. PACE—Pacing Clin. Electrophysiol. 2020, 43, 172–180. [Google Scholar] [CrossRef]
- McMurray, J.; Pfeffer, M.A. New Therapeutic Options in Congestive Heart Failure: Part I. Circulation 2002, 105, 2099–2106. [Google Scholar] [CrossRef] [Green Version]
- McMurray, J.; Pfeffer, M.A. New Therapeutic Options in Congestive Heart Failure: Part II. Circulation 2002, 105, 2223–2228. [Google Scholar] [CrossRef]
- Lalit, P.A.; Hei, D.J.; Raval, A.N.; Kamp, T.J. Induced Pluripotent Stem Cells for Post-Myocardial Infarction Repair: Remarkable Opportunities and Challenges. Circ. Res. 2014, 114, 1328–1345. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, H.; Xiao, D.; Yang, H.; Itzhaki, I.; Qin, X.; Chour, T.; Aguirre, A.; Lehmann, K.; Kim, Y.; et al. Comparison of Non-Human Primate versus Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes for Treatment of Myocardial Infarction. Stem Cell Rep. 2018, 10, 422–435. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Xu, W.; Zhang, H.; Wang, Q.; Yu, J.; Zhang, R.; Chen, Y.; Xia, Y.; Wang, J.; Wang, D. Transplantation of Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes Improves Myocardial Function and Reverses Ventricular Remodeling in Infarcted Rat Hearts. Stem Cell Res. Ther. 2020, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Ishida, M.; Miyagawa, S.; Saito, A.; Fukushima, S.; Harada, A.; Ito, E.; Ohashi, F.; Watabe, T.; Hatazawa, J.; Matsuura, K.; et al. Transplantation of Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes Is Superior to Somatic Stem Cell Therapy for Restoring Cardiac Function and Oxygen Consumption in a Porcine Model of Myocardial Infarction. Transplantation 2019, 103, 291–298. [Google Scholar] [CrossRef]
- Shiba, Y.; Gomibuchi, T.; Seto, T.; Wada, Y.; Ichimura, H.; Tanaka, Y.; Ogasawara, T.; Okada, K.; Shiba, N.; Sakamoto, K.; et al. Allogeneic Transplantation of IPS Cell-Derived Cardiomyocytes Regenerates Primate Hearts. Nature 2016, 538, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.J.H.; Yang, X.; Don, C.W.; Minami, E.; Liu, Y.W.; Weyers, J.J.; Mahoney, W.M.; Van Biber, B.; Cook, S.M.; Palpant, N.J.; et al. Human Embryonic-Stem-Cell-Derived Cardiomyocytes Regenerate Non-Human Primate Hearts. Nature 2014, 510, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Chen, B.; Yang, X.; Fugate, J.A.; Kalucki, F.A.; Futakuchi-Tsuchida, A.; Couture, L.; Vogel, K.W.; Astley, C.A.; Baldessari, A.; et al. Human Embryonic Stem Cell-Derived Cardiomyocytes Restore Function in Infarcted Hearts of Non-Human Primates. Nat. Biotechnol. 2018, 36, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Kashiyama, N.; Miyagawa, S.; Fukushima, S.; Kawamura, T.; Kawamura, A.; Yoshida, S.; Eiraku, S.; Harada, A.; Matsunaga, K.; Watabe, T.; et al. MHC-Mismatched Allotransplantation of Induced Pluripotent Stem Cell-Derived Cardiomyocyte Sheets to Improve Cardiac Function in a Primate Ischemic Cardiomyopathy Model. Transplantation 2019, 103, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.; Cunningham, N.J.; Shenoy, S.; Wu, J.C. Human-Induced Pluripotent Stem Cells in Cardiovascular Research: Current Approaches in Cardiac Differentiation, Maturation Strategies, and Scalable Production. Cardiovasc. Res. 2022, 118, 20–36. [Google Scholar] [CrossRef] [PubMed]
- Takada, T.; Sasaki, D.; Matsuura, K.; Miura, K.; Sakamoto, S.; Goto, H.; Ohya, T.; Iida, T.; Homma, J.; Shimizu, T.; et al. Aligned Human Induced Pluripotent Stem Cell-Derived Cardiac Tissue Improves Contractile Properties through Promoting Unidirectional and Synchronous Cardiomyocyte Contraction. Biomaterials 2022, 281, 121351. [Google Scholar] [CrossRef] [PubMed]
- Marchianò, S.; Bertero, A.; Murry, C.E. Learn from Your Elders: Developmental Biology Lessons to Guide Maturation of Stem Cell-Derived Cardiomyocytes. Pediatr. Cardiol. 2019, 40, 1367–1387. [Google Scholar] [CrossRef]
- Soma, Y.; Morita, Y.; Kishino, Y.; Kanazawa, H.; Fukuda, K.; Tohyama, S. The Present State and Future Perspectives of Cardiac Regenerative Therapy Using Human Pluripotent Stem Cells. Front. Cardiovasc. Med. 2021, 8, 774389. [Google Scholar] [CrossRef]
- Marcu, I.C.; Illaste, A.; Heuking, P.; Jaconi, M.E.; Ullrich, N.D. Functional Characterization and Comparison of Intercellular Communication in Stem Cell-Derived Cardiomyocytes. Stem Cells 2015, 33, 2208–2218. [Google Scholar] [CrossRef] [Green Version]
- Sottas, V.; Wahl, C.M.; Trache, M.C.; Bartolf-Kopp, M.; Cambridge, S.; Hecker, M.; Ullrich, N.D. Improving Electrical Properties of IPSC-Cardiomyocytes by Enhancing Cx43 Expression. J. Mol. Cell. Cardiol. 2018, 120, 31–41. [Google Scholar] [CrossRef]
- Vermij, S.H.; Abriel, H.; Van Veen, T.A.B. Refining the Molecular Organization of the Cardiac Intercalated Disc. Cardiovasc. Res. 2017, 113, 259–275. [Google Scholar] [CrossRef]
- Harris, A.L. Connexin Channel Permeability to Cytoplasmic Molecules. Prog. Biophys. Mol. Biol. 2007, 94, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Severs, N.J. Pathophysiology of Gap Junctions in Heart Disease. J. Cardiovasc. Electrophysiol. 1994, 5, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Pieperhoff, S.; Franke, W.W. The Area Composita of Adhering Junctions Connecting Heart Muscle Cells of Vertebrates—IV: Coalescence and Amalgamation of Desmosomal and Adhaerens Junction Components—Late Processes in Mammalian Heart Development. Eur. J. Cell Biol. 2007, 86, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Lemcke, H.; Steinhoff, G.; David, R. Gap Junctional Shuttling of MiRNA--A Novel Pathway of Intercellular Gene Regulation and Its Prospects in Clinical Application. Cell. Signal. 2015, 27, 2506–2514. [Google Scholar] [CrossRef]
- Söhl, G.; Willecke, K. Gap Junctions and the Connexin Protein Family. Cardiovasc. Res. 2004, 62, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solan, J.L.; Lampe, P.D. Connexin Phosphorylation as a Regulatory Event Linked to Gap Junction Channel Assembly. Biochim. Biophys. Acta 2005, 1711, 154–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yancey, S.B.; John, S.A.; Lal, R.; Austin, B.J.; Revel, J.P. The 43-KD Polypeptide of Heart Gap Junctions: Immunolocalization, Tolopogy, and Functional Domains. J. Cell Biol. 1989, 108, 2241–2254. [Google Scholar] [CrossRef] [Green Version]
- Brissette, J.L.; Kumar, N.M.; Gilula, N.B.; Hall, J.E.; Dotto, G.P. Switch in Gap Junction Protein Expression Is Associated with Selective Changes in Junctional Permeability during Keratinocyte Differentiation. Proc. Natl. Acad. Sci. USA 1994, 91, 6453–6457. [Google Scholar] [CrossRef] [Green Version]
- Lo, C.W. Role of Gap Junctions in Cardiac Conduction and Development: Insights from the Connexin Knockout Mice. Circ. Res. 2000, 87, 346–348. [Google Scholar] [CrossRef] [Green Version]
- Vozzi, C.; Dupont, E.; Coppen, S.R.; Hung-I, Y.; Severs, N.J. Chamber-Related Differences in Connexin Expression in the Human Heart. J. Mol. Cell. Cardiol. 1999, 31, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Van Kempen, M.J.A.; Fromaget, C.; Gros, D.; Moorman, A.F.M.; Lamers, W.H. Spatial Distribution of Connexin43, the Major Cardiac Gap Junction Protein, in the Developing and Adult Rat Heart. Circ. Res. 1991, 68, 1638–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Kempen, M.J.A.; Velde, I.T.; Wessels, A.; Oosthoek, P.W.; Gros, D.; Jongsma, H.J.; Moorman, A.F.M.; Lamers, W.H. Differential Connexin Distribution Accommodates Cardiac Function in Different Species. Microsc. Res. Tech. 1995, 31, 420–436. [Google Scholar] [CrossRef] [PubMed]
- Jordan, K.; Chodock, R.; Hand, A.R.; Laird, D.W. The Origin of Annular Junctions: A Mechanism of Gap Junction Internalization. J. Cell Sci. 2001, 114, 763–773. [Google Scholar] [CrossRef]
- Gaietta, G.; Deerinck, T.J.; Adams, S.R.; Bouwer, J.; Tour, O.; Laird, D.W.; Sosinsky, G.E.; Tsien, R.Y.; Ellisman, M.H. Multicolor and Electron Microscopic Imaging of Connexin Trafficking. Science 2002, 296, 503–507. [Google Scholar] [CrossRef] [Green Version]
- Beardslee, M.A.; Laing, J.G.; Beyer, E.C.; Saffitz, J.E. Rapid Turnover of Connexin43 in the Adult Rat Heart. Circ. Res. 1998, 83, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Fallon, R.F.; Goodenough, D.A. Five-Hour Half-Life of Mouse Liver Gap-Junction Protein. J. Cell Biol. 1981, 90, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Laird, D.W.; Puranam, K.L.; Revel, J.P. Turnover and Phosphorylation Dynamics of Connexin43 Gap Junction Protein in Cultured Cardiac Myocytes. Biochem. J. 1991, 273 Pt 1, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Musil, L.S.; Goodenough, D.A. Multisubunit Assembly of an Integral Plasma Membrane Channel Protein, Gap Junction Connexin43, Occurs after Exit from the ER. Cell 1993, 74, 1065–1077. [Google Scholar] [CrossRef]
- Das Sarma, J.; Wang, F.; Koval, M. Targeted Gap Junction Protein Constructs Reveal Connexin-Specific Differences in Oligomerization. J. Biol. Chem. 2002, 277, 20911–20918. [Google Scholar] [CrossRef] [Green Version]
- Laird, D.W. Life Cycle of Connexins in Health and Disease. Biochem. J. 2006, 394, 527–543. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.S.; Shaw, R.M. Trafficking Highways to the Intercalated Disc: New Insights Unlocking the Specificity of Connexin 43 Localization. Cell Commun. Adhes. 2014, 21, 43–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorgen, P.L.; Trease, A.J.; Spagnol, G.; Delmar, M.; Nielsen, M.S. Protein−Protein Interactions with Connexin 43: Regulation and Function. Int. J. Mol. Sci. 2018, 19, 1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, A.L. Emerging Issues of Connexin Channels: Biophysics Fills the Gap. Q. Rev. Biophys. 2001, 34, 325–472. [Google Scholar] [CrossRef] [Green Version]
- Revel, J.P.; Karnovsky, M.J. Hexagonal Array of Subunits in Intercellular Junctions of the Mouse Heart and Liver. J. Cell Biol. 1967, 33, C7–C12. [Google Scholar] [CrossRef]
- Forbes, M.S.; Sperelakis, N. Intercalated Discs of Mammalian Heart: A Review of Structure and Function. Tissue Cell 1985, 17, 605–648. [Google Scholar] [CrossRef]
- Hirschy, A.; Schatzmann, F.; Ehler, E.; Perriard, J.C. Establishment of Cardiac Cytoarchitecture in the Developing Mouse Heart. Dev. Biol. 2006, 289, 430–441. [Google Scholar] [CrossRef] [Green Version]
- Noorman, M.; Hakim, S.; Kessler, E.; Groeneweg, J.A.; Cox, M.G.P.J.; Asimaki, A.; Van Rijen, H.V.M.; Van Stuijvenberg, L.; Chkourko, H.; Van Der Heyden, M.A.G.; et al. Remodeling of the Cardiac Sodium Channel, Connexin43, and Plakoglobin at the Intercalated Disk in Patients with Arrhythmogenic Cardiomyopathy. Hear. Rhythm 2013, 10, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Agullo-Pascual, E.; Cerrone, M.; Delmar, M. Arrhythmogenic Cardiomyopathy and Brugada Syndrome: Diseases of the Connexome. FEBS Lett. 2014, 588, 1322–1330. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.; Cerrone, M.; Saguner, A.M.; Brunckhorst, C.; Delmar, M.; Duru, F. Arrhythmogenic Cardiomyopathy: An in-Depth Look at Molecular Mechanisms and Clinical Correlates. Trends Cardiovasc. Med. 2021, 31, 395–402. [Google Scholar] [CrossRef]
- Rampazzo, A.; Calore, M.; Van Hengel, J.; Van Roy, F. Intercalated Discs and Arrhythmogenic Cardiomyopathy. Circ. Cardiovasc. Genet. 2014, 7, 930–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gherghiceanu, M.; Barad, L.; Novak, A.; Reiter, I.; Itskovitz-Eldor, J.; Binah, O.; Popescu, L.M. Cardiomyocytes Derived from Human Embryonic and Induced Pluripotent Stem Cells: Comparative Ultrastructure. J. Cell. Mol. Med. 2011, 15, 2539–2551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuppinger, C.; Gibbons, G.; Dutta-Passecker, P.; Segiser, A.; Most, H.; Suter, T.M. Characterization of Cytoskeleton Features and Maturation Status of Cultured Human IPSC-Derived Cardiomyocytes. Eur. J. Histochem. 2017, 61, 145–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silbernagel, N.; Körner, A.; Balitzki, J.; Jaggy, M.; Bertels, S.; Richter, B.; Hippler, M.; Hellwig, A.; Hecker, M.; Bastmeyer, M.; et al. Shaping the Heart: Structural and Functional Maturation of IPSC-Cardiomyocytes in 3D-Micro-Scaffolds. Biomaterials 2020, 227, 119551. [Google Scholar] [CrossRef]
- Kucera, J.P.; Prudat, Y.; Marcu, I.C.; Azzarito, M.; Ullrich, N.D. Slow Conduction in Mixed Cultured Strands of Primary Ventricular Cells and Stem Cell-Derived Cardiomyocytes. Front. Cell Dev. Biol. 2015, 3, 58. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.Y.; Tse, H.F.; Chan, Y.C.; Mei-Chu Yip, P.; Zhang, Y.; Liu, Y.; Li, R.A. Overexpression of Kir2.1 Channel in Embryonic Stem Cell-Derived Cardiomyocytes Attenuates Posttransplantation Proarrhythmic Risk in Myocardial Infarction. Hear. Rhythm 2013, 10, 273–282. [Google Scholar] [CrossRef]
- Liao, S.Y.; Liu, Y.; Siu, C.W.; Zhang, Y.; Lai, W.H.; Au, K.W.; Lee, Y.K.; Chan, Y.C.; Yip, P.M.C.; Wu, E.X.; et al. Proarrhythmic Risk of Embryonic Stem Cell-Derived Cardiomyocyte Transplantation in Infarcted Myocardium. Hear. Rhythm 2010, 7, 1852–1859. [Google Scholar] [CrossRef]
- Chong, J.J.H.; Murry, C.E. Cardiac Regeneration Using Pluripotent Stem Cells—Progression to Large Animal Models. Stem Cell Res. 2014, 13, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Roell, W.; Lewalter, T.; Sasse, P.; Tallini, Y.N.; Choi, B.R.; Breitbach, M.; Doran, R.; Becher, U.M.; Hwang, S.M.; Bostani, T.; et al. Engraftment of Connexin 43-Expressing Cells Prevents Post-Infarct Arrhythmia. Nature 2007, 450, 819–824. [Google Scholar] [CrossRef]
- Hirt, M.N.; Boeddinghaus, J.; Mitchell, A.; Schaaf, S.; Börnchen, C.; Müller, C.; Schulz, H.; Hubner, N.; Stenzig, J.; Stoehr, A.; et al. Functional Improvement and Maturation of Rat and Human Engineered Heart Tissue by Chronic Electrical Stimulation. J. Mol. Cell. Cardiol. 2014, 74, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Nunes, S.S.; Miklas, J.W.; Liu, J.; Aschar-Sobbi, R.; Xiao, Y.; Zhang, B.; Jiang, J.; Massé, S.; Gagliardi, M.; Hsieh, A.; et al. Biowire: A Platform for Maturation of Human Pluripotent Stem Cell-Derived Cardiomyocytes. Nat. Methods 2013, 10, 781–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Nunes, S.S. Maturation of Human Stem Cell-Derived Cardiomyocytes in Biowires Using Electrical Stimulation. J. Vis. Exp. 2017, 2017, e55373. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Feric, N.; Pallotta, I.; Singh, R.; Sobbi, R.; Vargas, H.M. Inotropic Assessment in Engineered 3D Cardiac Tissues Using Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes in the Biowire TM II Platform. J. Pharmacol. Toxicol. Methods 2020, 105, 106886. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, T.; Oyamada, M.; Fujimoto, K.; Yasuda, Y.; Masuda, S.; Wada, Y.; Takahiro, O.; Takamatsu, T. Remodeling of Cell-Cell and Cell-Extracellular Matrix Interactions at the Border Zone of Rat Myocardial Infarcts. Circ. Res. 1999, 85, 1046–1055. [Google Scholar] [CrossRef] [Green Version]
- Severs, N.J.; Bruce, A.F.; Dupont, E.; Rothery, S. Remodelling of Gap Junctions and Connexin Expression in Diseased Myocardium. Cardiovasc. Res. 2008, 80, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Agullo-Pascual, E.; Lin, X.; Leo-Macias, A.; Zhang, M.; Liang, F.X.; Li, Z.; Pfenniger, A.; Lübkemeier, I.; Keegan, S.; Fenyo, D.; et al. Super-Resolution Imaging Reveals That Loss of the C-Terminus of Connexin43 Limits Microtubule plus-End Capture and NaV1.5 Localization at the Intercalated Disc. Cardiovasc. Res. 2014, 104, 371–381. [Google Scholar] [CrossRef]
- Jansen, J.A.; Noorman, M.; Musa, H.; Stein, M.; De Jong, S.; Van Der Nagel, R.; Hund, T.J.; Mohler, P.J.; Vos, M.A.; Van Veen, T.A.; et al. Reduced Heterogeneous Expression of Cx43 Results in Decreased Nav1.5 Expression and Reduced Sodium Current That Accounts for Arrhythmia Vulnerability in Conditional Cx43 Knockout Mice. Hear. Rhythm 2012, 9, 600–607. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Li, H.; Cannon, A.; Trease, A.J.; Spagnol, G.; Zheng, H.; Radio, S.; Patel, K.; Batra, S.; Sorgen, P.L. Phosphorylation of Cx43 Residue Y313 by Src Contributes to Blocking the Interaction with Drebrin and Disassembling Gap Junctions. J. Mol. Cell. Cardiol. 2019, 126, 36–49. [Google Scholar] [CrossRef]
- Dhein, S. Gap Junction Channels in the Cardiovascular System: Pharmacological and Physiological Modulation. Trends Pharmacol. Sci. 1998, 19, 229–241. [Google Scholar] [CrossRef]
- Koutsoulidou, A.; Mastroyiannopoulos, N.P.; Furling, D.; Uney, J.B.; Phylactou, L.A. Expression of MiR-1, MiR-133a, MiR-133b and MiR-206 Increases during Development of Human Skeletal Muscle. BMC Dev. Biol. 2011, 11, 34. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhou, H.; Tang, Q. MiR-133: A Suppressor of Cardiac Remodeling? Front. Pharmacol. 2018, 9, 903. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiss, E.; Fischer, C.; Sauter, J.-M.; Sun, J.; Ullrich, N.D. The Structural and the Functional Aspects of Intercellular Communication in iPSC-Cardiomyocytes. Int. J. Mol. Sci. 2022, 23, 4460. https://doi.org/10.3390/ijms23084460
Kiss E, Fischer C, Sauter J-M, Sun J, Ullrich ND. The Structural and the Functional Aspects of Intercellular Communication in iPSC-Cardiomyocytes. International Journal of Molecular Sciences. 2022; 23(8):4460. https://doi.org/10.3390/ijms23084460
Chicago/Turabian StyleKiss, Eva, Carolin Fischer, Jan-Mischa Sauter, Jinmeng Sun, and Nina D. Ullrich. 2022. "The Structural and the Functional Aspects of Intercellular Communication in iPSC-Cardiomyocytes" International Journal of Molecular Sciences 23, no. 8: 4460. https://doi.org/10.3390/ijms23084460