
IKKα Induces Epithelial–Mesenchymal Changes in Mouse Skin Carcinoma Cells That Can Be Partially Reversed by Apigenin

 , , and
, , and
Abstract
:1. Introduction
2. Results
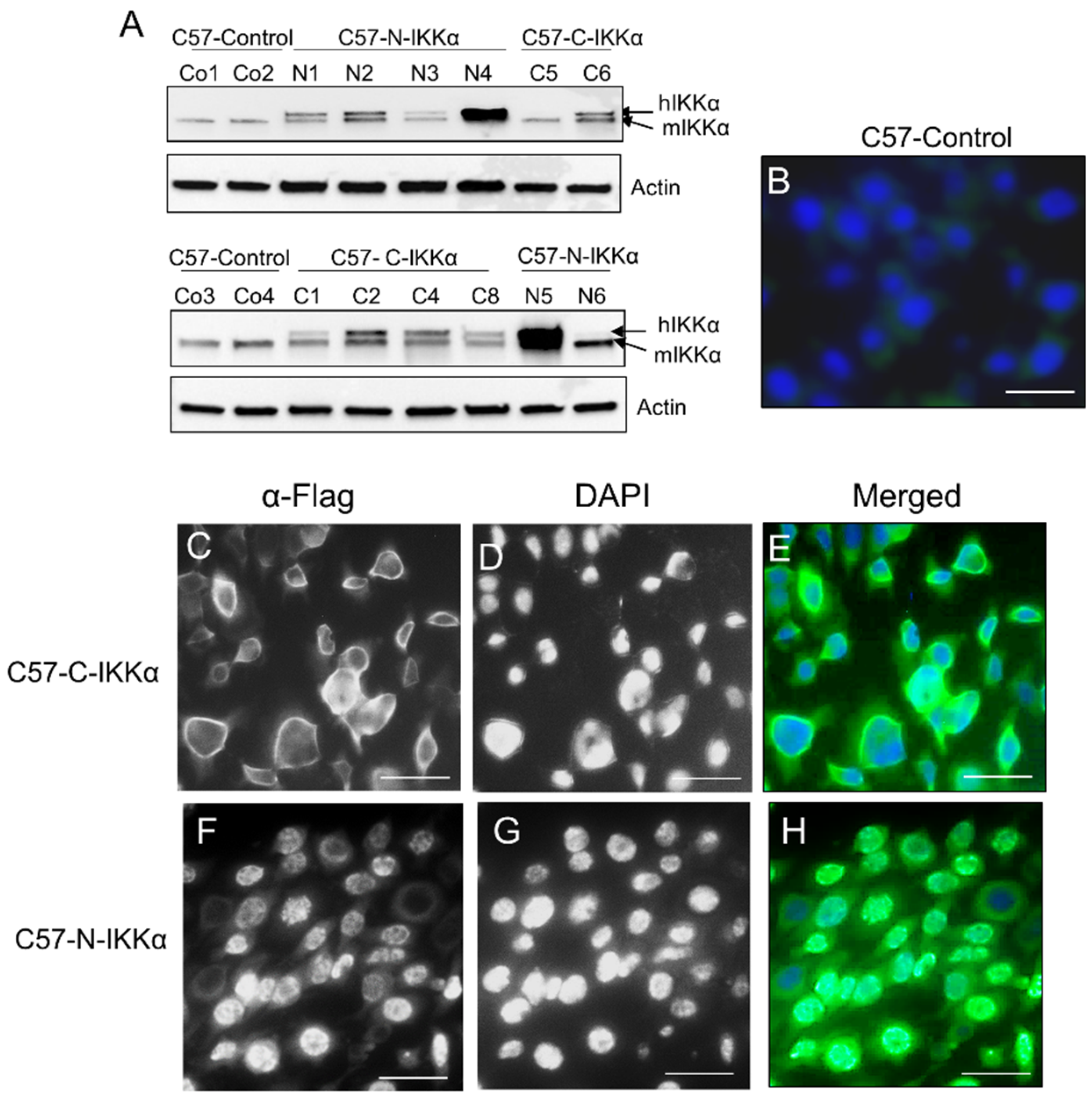
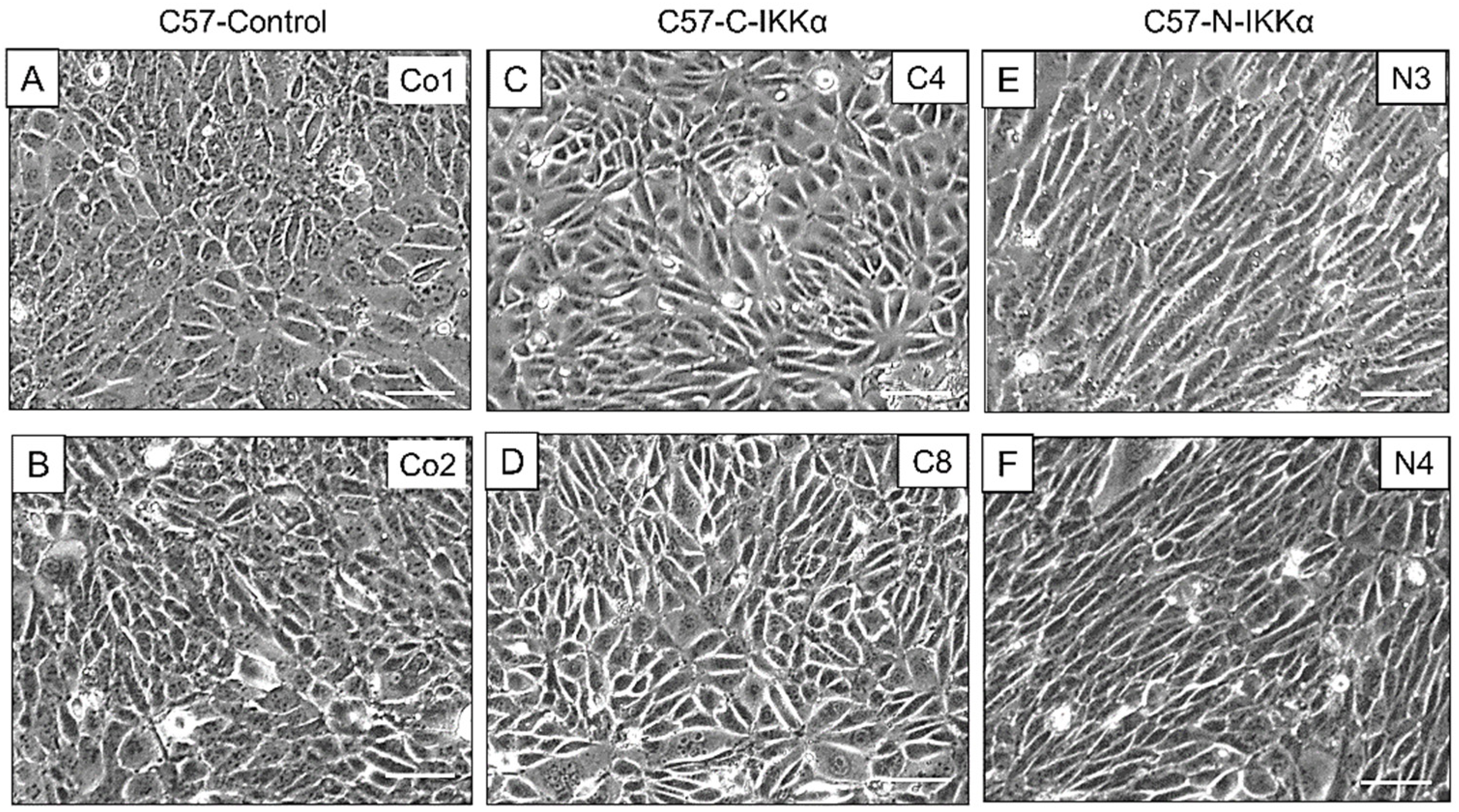
2.1. Exogenous Expression of IKKα in the Nucleus or Cytoplasm of PDVC57 Skin Carcinoma Cells
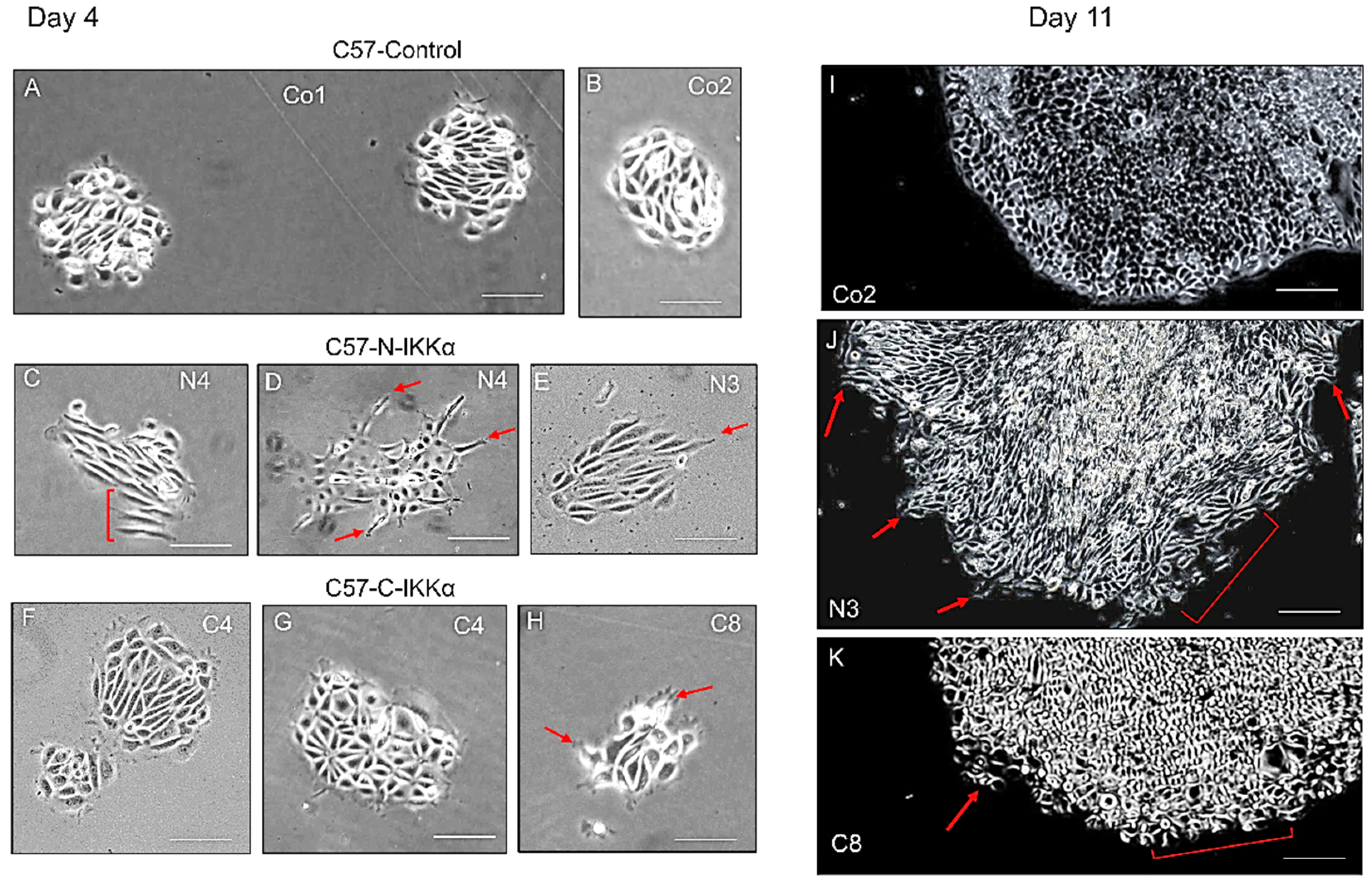
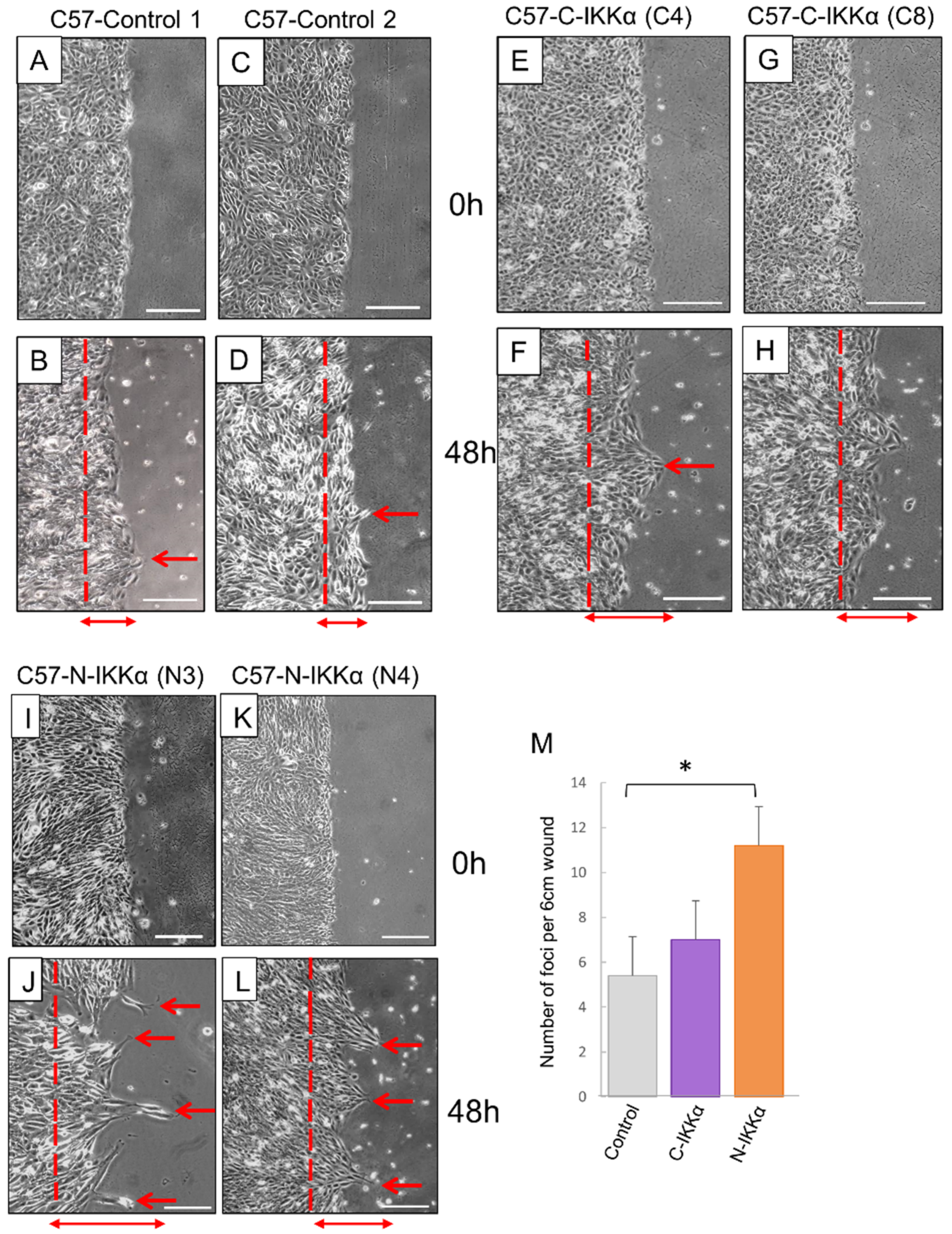
2.2. Increased Migratory Capacity of Both C57-C-IKKα and C57-N-IKKα Cells in Culture
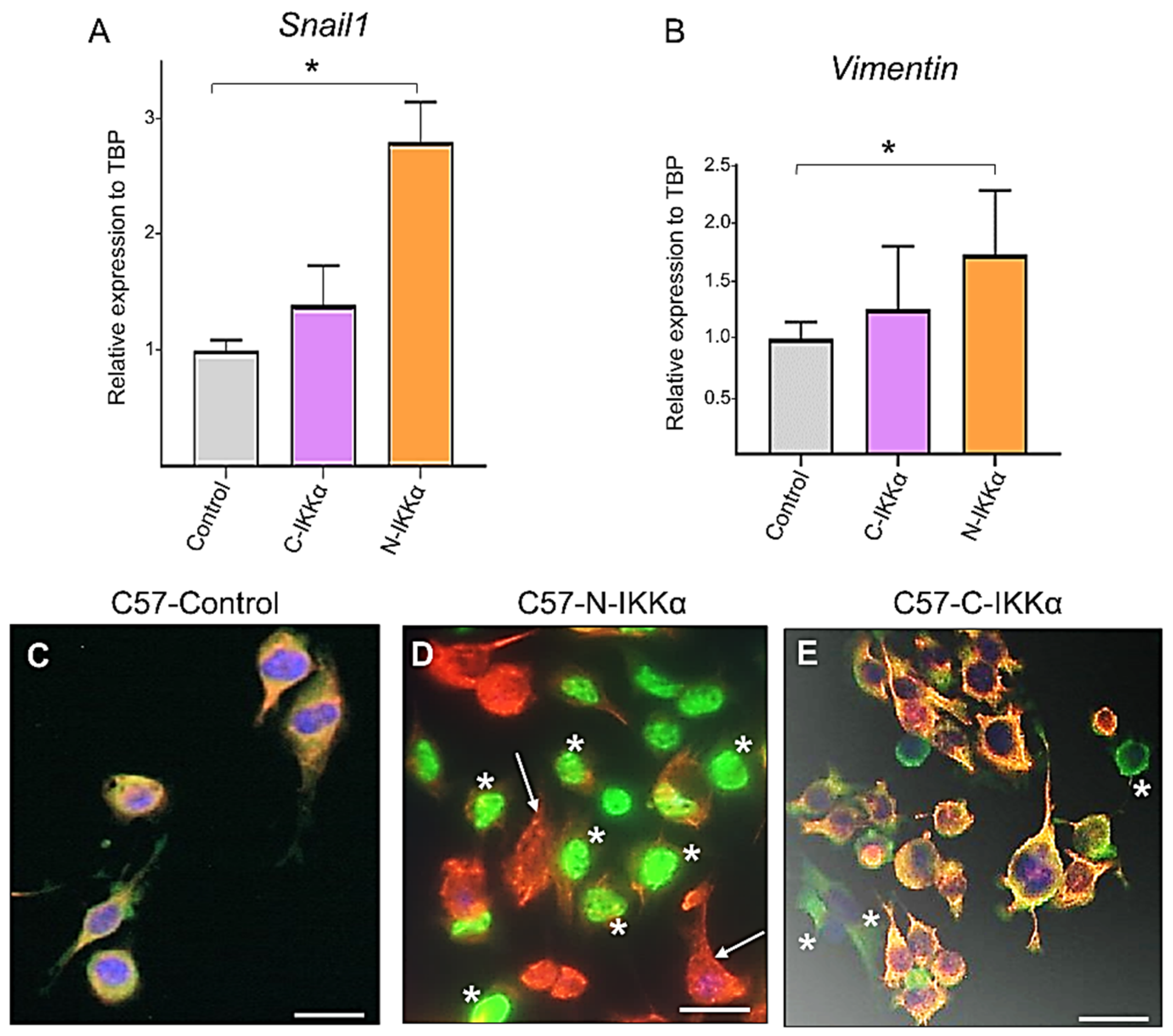
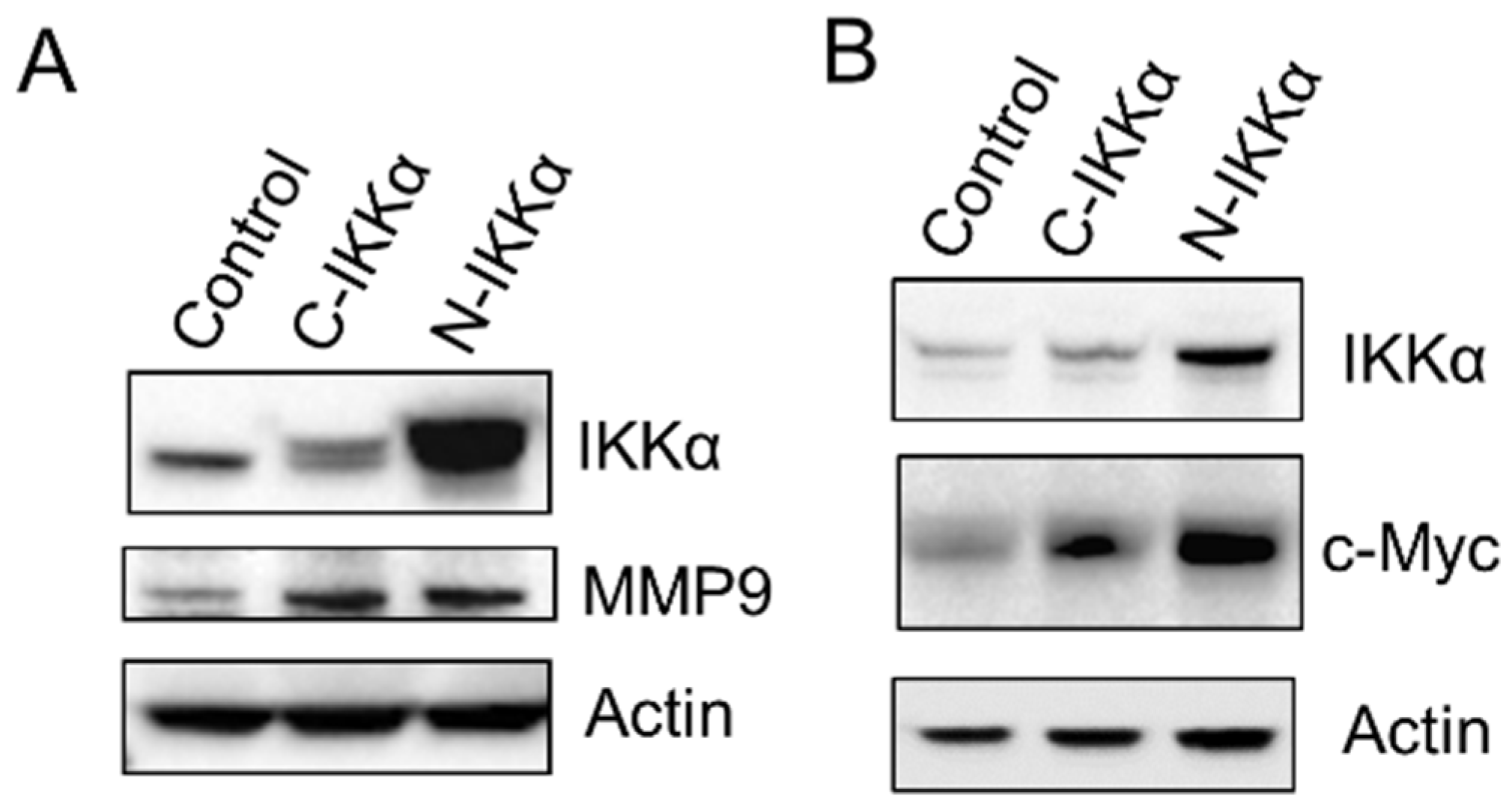
2.3. Induction of Snail and Vimentin Expression and Inhibition of K5 in C57-N-IKKα Cells
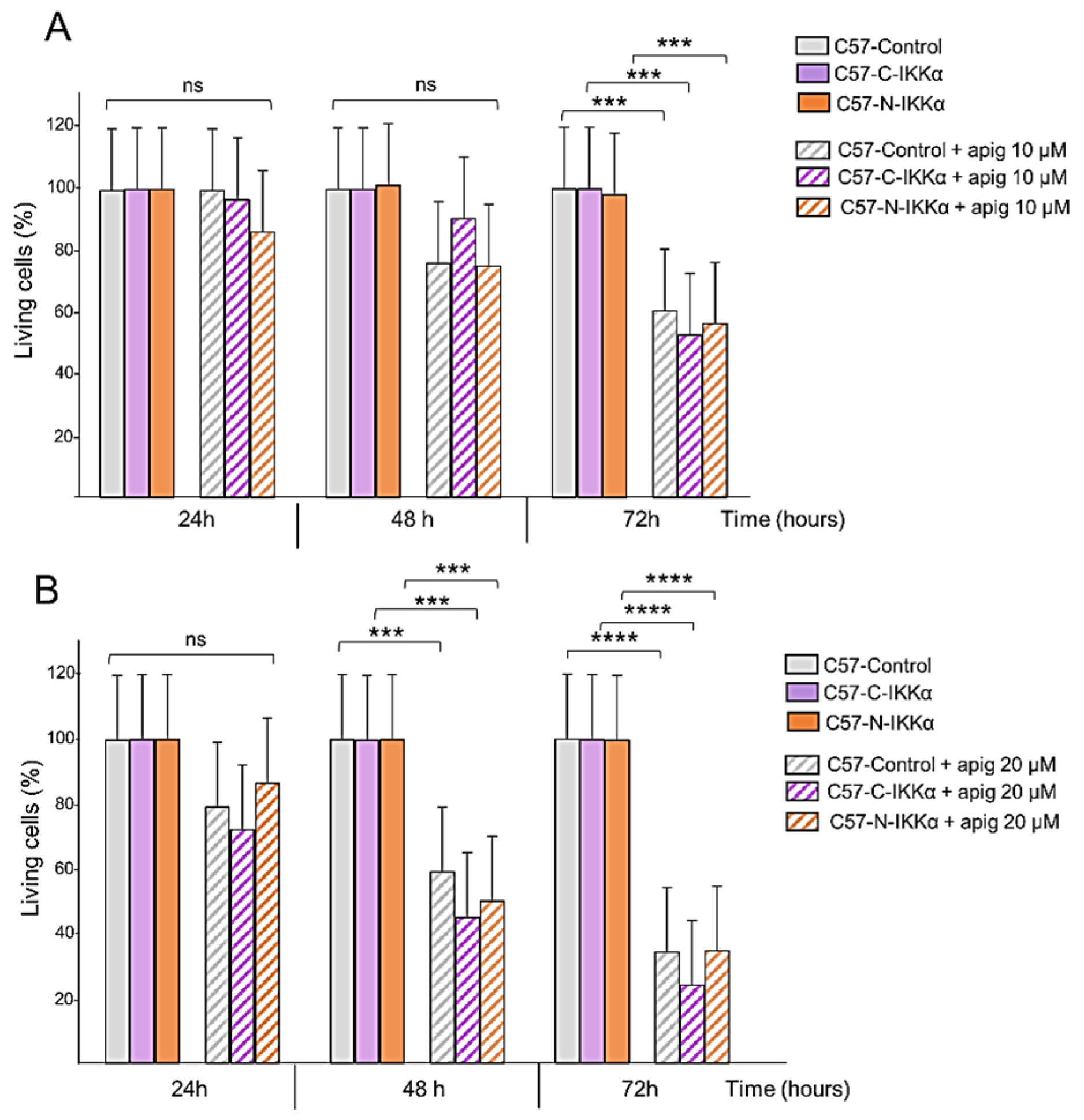
2.4. C57-N-IKKα Cells Have Increased Survival Capacity
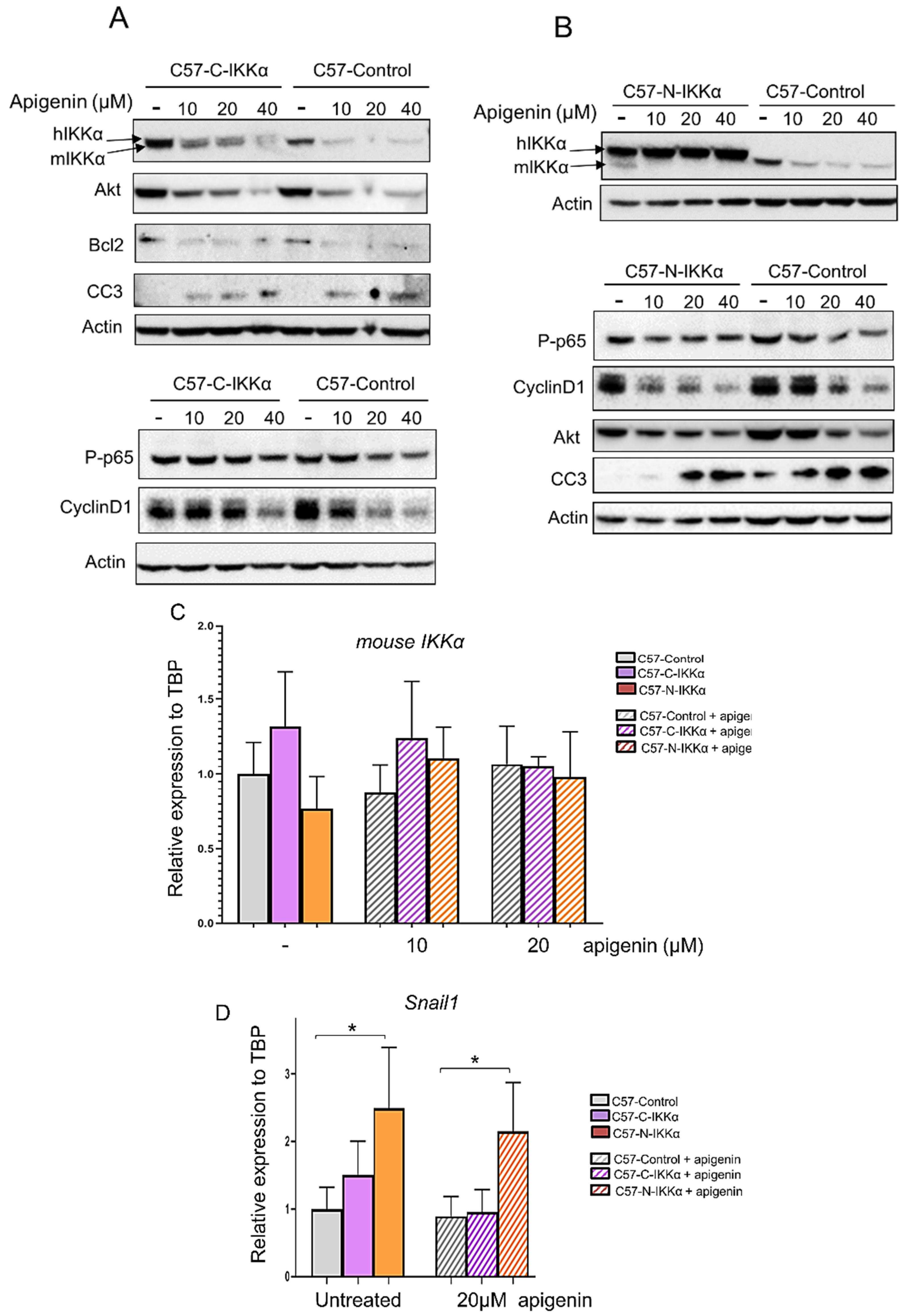
2.5. Apigenin Inhibits the Expression of the IKKα Protein in the Cytoplasm but Not in the Nucleus of C57 Cells
3. Discussion
4. Materials and Methods
4.1. DNA Constructs
4.2. Cells, Culture Conditions and Transfection Assays
4.3. Colony Forming Assays
4.4. Wound Healing Assays
4.5. Growth in Serum-Free Medium
4.6. Cell Viability Assay
4.7. Immunofluorescence Staining
4.8. Western Blot Analysis
4.9. RNA Isolation and Quantitative RT-qPCR Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BCCs | basal cell carcinomas |
| CC3 | cleaved-Caspase 3 |
| EMT | epithelial–mesenchymal transition |
| FBS | fetal bovine serum |
| NMSC | non melanoma skin cancer |
| NSCLC | non-small cell lung cancer |
| RDEB | recessive epidermolysis bullosa |
| SCCs | squamous cell carcinomas |
References
- Christenson, L.J.; Borrowman, T.A.; Vachon, C.M.; Tollefson, M.M.; Otley, C.C.; Weaver, A.L.; Roenigk, R.K. Incidence of basal cell and squamous cell carcinomas in a population younger than 40 years. JAMA 2005, 294, 681–690. [Google Scholar] [CrossRef] [Green Version]
- Karia, P.S.; Han, J.; Schmults, C.D. Cutaneous squamous cell carcinoma: Estimated incidence of disease, nodal metastasis, and deaths from disease in the United States, 2012. J. Am. Acad. Dermatol. 2013, 68, 957–966. [Google Scholar] [CrossRef]
- Condorelli, A.G.; Dellambra, E.; Logli, E.; Zambruno, G.; Castiglia, D. Epidermolysis Bullosa-associated squamous cell carcinoma: From pathogenesis to therapeutic perspectives. Int. J. Mol. Sci. 2019, 20, 5707. [Google Scholar] [CrossRef] [Green Version]
- Mittal, A.; Colegio, O.R. Skin cancers in organ transplant recipients. Am. J. Transplant. 2017, 17, 2509–2530. [Google Scholar] [CrossRef]
- Toll, A.; Margalef, P.; Masferrer, E.; Ferrándiz-Pulido, C.; Gimeno, J.; Pujol, R.M.; Bigas, A.; Espinosa, L. Active nuclear IKK correlates with metastatic risk in cutaneous squamous cell carcinoma. Arch. Dermatol. Res. 2015, 21, 21. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. Signaling to NF-kappaB. Genes Dev. 2004, 18, 2195–2224. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Lu, Q.; Hwang, J.W.; Büscher, D.; Lee, K.F.; Izpisua-Belmonte, J.C.; Verma, I.M. IKK1-deficient mice exhibit abnormal development of skin and skeleton. Genes Dev. 1999, 13, 1322–1328. [Google Scholar] [CrossRef]
- Takeda, K.; Takeuchi, O.; Tsujimura, T.; Itami, S.; Adachi, O.; Kawai, T.; Sanjo, H.; Yoshikawa, K.; Terada, N.; Akira, S. Limb and skin abnormalities in mice lacking IKKalpha. Science 1999, 284, 313–316. [Google Scholar] [CrossRef]
- Anest, V.; Hanson, J.L.; Cogswell, P.C.; Steinbrecher, K.A.; Strahl, B.D.; Baldwin, A.S. A nucleosomal function for IkappaB kinase-alpha in NF-kappaB-dependent gene expression. Nature 2003, 423, 659–663. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.L.; Tan, W.; Ricono, J.M.; Korchynskyi, O.; Zhang, M.; Gonias, S.L.; Cheresh, D.A.; Karin, M. Nuclear cytokine-activated IKKalpha controls prostate cancer metastasis by repressing Maspin. Nature 2007, 446, 690–694. [Google Scholar] [CrossRef]
- Fernandez-Majada, V.; Aguilera, C.; Villanueva, A.; Vilardell, F.; Robert-Moreno, A.; Aytés, A.; Real, F.X.; Capella, G.; Mayo, M.W.; Espinosa, L.; et al. Nuclear IKK activity leads to dysregulated notch-dependent gene expression in colorectal cancer. Proc. Natl. Acad. Sci. USA 2007, 104, 276–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Baud, V.; Delhase, M.; Zhang, P.; Deerinck, T.; Ellisman, M.; Johnson, R.; Karin, M. Abnormal morphogenesis but intact IKK activation in mice lacking the IKKalpha subunit of IkappaB kinase. Science 1999, 284, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Van Antwerp, D.; Mercurio, F.; Lee, K.F.; Verma, I.M. Severe liver degeneration in mice lacking the IkappaB kinase 2 gene. Science 1999, 284, 321–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sil, A.K.; Maeda, S.; Sano, Y.; Roop, D.R.; Karin, M. IkappaB kinase-alpha acts in the epidermis to control skeletal and craniofacial morphogenesis. Nature 2004, 428, 660–664. [Google Scholar] [CrossRef]
- Alameda, J.P.; Gaspar, M.; Ramírez, A.; Navarro, M.; Page, A.; Suárez-Cabrera, C.; Fernández, M.G.; Mérida, J.R.; Paramio, J.M.; García-Fernández, R.-A.; et al. Deciphering the role of nuclear and cytoplasmic IKKalpha in skin cancer. Oncotarget 2016, 7, 29531–29547. [Google Scholar] [CrossRef]
- Alameda, J.P.; Moreno-Maldonado, R.; Fernández-Aceñero, M.J.; Navarro, N.; Page, A.; Jorcano, J.L.; Bravo, A.; Ramírez, A.; Casanova, M.L. Increased IKKalpha expression in the Basal layer of the epidermis of transgenic mice enhances the malignant potential of skin tumors. PLoS ONE 2011, 6, e21984. [Google Scholar] [CrossRef]
- Moreno-Maldonado, R.; Ramírez, A.; Navarro, M.; Fernández-Aceñero, M.J.; Villanueva, C.; Page, A.; Jorcano, J.L.; Bravo, A.; Casanova, M.L. IKKalpha enhances human keratinocyte differentiation and determines the histological variant of epidermal squamous cell carcinomas. Cell Cycle 2008, 7, 2021–2029. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Park, E.; Zhu, F.; Bustos, T.; Liu, J.; Shen, J.; Fischer, S.M.; Hu, Y. A critical role for I kappaB kinase alpha in the development of human and mouse squamous cell carcinomas. Proc. Natl. Acad. Sci. USA 2006, 103, 17202–17207. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, M. Involvement of partial EMT in cancer progression. J. Biochem. 2018, 164, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Leggett, S.E.; Hruska, A.M.; Guo, M.; Wong, Y. The epithelial-mesenchymal transition and the cytoskeleton in bioengineered systems. Cell Commun. Signal. 2021, 19, 32. [Google Scholar] [CrossRef] [PubMed]
- Scheau, C.; Badarau, I.A.; Costache, R.; Caruntu, C.; Mihai, G.L.; Didilescu, A.C.; Constantin, C.; Neagu, M. The role of matrix metalloproteinases in the epithelial-mesenchymal transition of hepatocellular carcinoma. Anal. Cell Pathol. 2019, 2019, 9423907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, N.; Gheldof, A.; Tatari, M.; Christofori, G. EMT as the ultimate survival mechanism of cancer cells. Semin. Cancer Biol. 2012, 22, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Robson, E.J.; Khaled, W.T.; Abell, K.; Watson, C.J. Epithelial-to-mesenchymal transition confers resistance to apoptosis in three murine mammary epithelial cell lines. Differentiation 2006, 74, 254–264. [Google Scholar] [CrossRef]
- Kurrey, N.K.; Jalgaonkar, S.P.; Joglekar, A.V.; Ghanate, A.D.; Chaskar, P.D.; Doiphode, R.Y.; Bapat, S.A. Snail and slug mediate radioresistance and chemoresistance by antagonizing p53-mediated apoptosis and acquiring a stem-like phenotype in ovarian cancer cells. Stem Cells 2009, 27, 2059–2068. [Google Scholar] [CrossRef]
- Nieto, M.A. The snail superfamily of zinc-finger transcription factors. Nat. Rev. Mol. Cell Biol. 2002, 3, 155–166. [Google Scholar] [CrossRef]
- Smith, B.N.; Bhowmick, N.A. Role of EMT in metastasis and therapy resistance. J. Clin. Med. 2016, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Kalantari, M.; Mohammadinejad, R.; Javaheri, T.; Sethi, G. Association of the Epithelial-Mesenchymal Transition (EMT) with cisplatin resistance. Int. J. Mol. Sci. 2020, 21, 4002. [Google Scholar] [CrossRef]
- Saenz-Sardà, X.; Carrato, C.; Pérez-Roca, L.; Puig, L.; Ferrándiz, C.; Ariza, A.; Fernández-Figueras, M.T. Epithelial-to-mesenchymal transition contributes to invasion in squamous cell carcinomas originated from actinic keratosis through the differentiated pathway, whereas proliferation plays a more significant role in the classical pathway. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 581–586. [Google Scholar] [CrossRef]
- Barrette, K.; Van Kelst, S.; Wouters, J.; Marasigan, V.; Fieuws, S.; Agostinis, P.; van den Oord, J.; Garmyn, M. Epithelial-mesenchymal transition during invasion of cutaneous squamous cell carcinoma is paralleled by AKT activation. Br. J. Dermatol. 2014, 171, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Ashrafizadeh, M.; Bakhoda, M.R.; Bahmanpour, Z.; Ilkhani, K.; Zarrabi, A.; Makvandi, P.; Khan, H.; Mazaheri, S.; Darvish, M.; Mirzaei, H. Apigenin as tumor suppressor in cancers: Biotherapeutic activity, nanodelivery, and mechanisms with emphasis on pancreatic cancer. Front. Chem. 2020, 8, 829. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Kanwal, R.; Shankar, E.; Datt, M.; Chance, M.R.; Fu, P.; MacLennan, G.T.; Gupta, S. Apigenin blocks IKKalpha activation and suppresses prostate cancer progression. Oncotarget 2015, 6, 31216–31232. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Shankar, E.; Fu, P.; MacLennan, G.T.; Gupta, S. Suppression of NF-κB and NF-κB-regulated gene expression by apigenin through IκBα and IKK pathway in TRAMP mice. PLoS ONE 2015, 10, e0138710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiraly, A.J.; Soliman, E.; Jenkins, A.; Van Dross, R.T. Apigenin inhibits COX-2, PGE2, and EP1 and also initiates terminal differentiation in the epidermis of tumor bearing mice. Prostaglandins Leukot. Essent. Fatty Acids 2016, 104, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Mirzoeva, S.; Tong, X.; Bridgeman, B.B.; Plebanek, M.P.; Volpert, O.V. Apigenin inhibits UVB-induced skin carcinogenesis: The role of thrombospondin-1 as an anti-inflammatory factor. Neoplasia 2018, 20, 930–942. [Google Scholar] [CrossRef]
- Pierceall, W.E.; Goldberg, L.H.; Tainsky, M.A.; Mukhopadhyay, T.; Ananthaswamy, H.N. Ras gene mutation and amplification in human nonmelanoma skin cancers. Mol. Carcinog. 1991, 4, 196–202. [Google Scholar] [CrossRef]
- Spencer, J.M.; Kahn, S.M.; Jiang, W.; DeLeo, V.A.; Weinstein, I.B. Activated ras genes occur in human actinic keratoses, premalignant precursors to squamous cell carcinomas. Arch. Dermatol. 1995, 131, 796–800. [Google Scholar] [CrossRef]
- Caulin, C.; Bauluz, C.; Gandarillas, A.; Cano, A.; Quintanilla, M. Changes in keratin expression during malignant progression of transformed mouse epidermal keratinocytes. Exp. Cell Res. 1993, 204, 11–21. [Google Scholar] [CrossRef]
- Larcher, F.; Robles, A.I.; Duran, H.; Murillas, R.; Quintanilla, M.; Cano, A.; Conti, C.J.; Jorcano, J.L. Up-regulation of vascular endothelial growth factor/vascular permeability factor in mouse skin carcinogenesis correlates with malignant progression state and activated H-ras expression levels. Cancer Res. 1996, 56, 5391–5396. [Google Scholar]
- Eriksson, J.E.; Dechat, T.; Grin, B.; Helfand, B.; Mendez, M.; Pallari, H.M.; Goldman, R.D. Introducing intermediate filaments: From discovery to disease. J. Clin. Investig. 2009, 119, 1763–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shchors, K.; Nozawa, H.; Xu, J.; Rostker, F.; Swigart-Brown, L.; Evan, G.; Hanahan, D. Increased invasiveness of MMP-9-deficient tumors in two mouse models of neuroendocrine tumorigenesis. Oncogene 2013, 32, 502–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.B.; Cho, M.K.; Lee, W.Y.; Kang, K.W. Overexpression of c-myc induces epithelial mesenchymal transition in mammary epithelial cells. Cancer Lett. 2010, 293, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Page, A.; Ortega, A.; Alameda, J.P.; Navarro, M.; Paramio, J.M.; Saiz-Pardo, M.; Almeida, E.I.; Hernández, P.; Fernández-Aceñero, M.-J.; García-Fernández, R.A.; et al. IKKα promotes the progression and metastasis of non-small cell lung cancer independently of its subcellular localization. Comput. Struct. Biotechnol. J. 2019, 17, 251–262. [Google Scholar] [CrossRef]
- Yuspa, S.H.; Morgan, D.; Lichti, U.; Spangler, E.F.; Michael, D.; Kilkenny, A.; Hennings, H. Cultivation and characterization of cells derived from mouse skin papillomas induced by an initiation-promotion protocol. Carcinogenesis 1986, 7, 949–958. [Google Scholar] [CrossRef]
- Vivanco, I.; Chen, Z.C.; Tanos, B.; Oldrini, B.; Hsieh, W.Y.; Yannuzzi, N.; Campos, C.; Mellinghoff, I.K. A kinase-independent function of AKT promotes cancer cell survival. Elife 2014, 3, e03751. [Google Scholar] [CrossRef]
- Shukla, S.; Bhaskaran, N.; Babcook, M.A.; Fu, P.; Maclennan, G.T.; Gupta, S. Apigenin inhibits prostate cancer progression in TRAMP mice via targeting PI3K/Akt/FoxO pathway. Carcinogenesis 2014, 35, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Altomare, D.A.; Testa, J.R. Perturbations of the AKT signaling pathway in human cancer. Oncogene 2005, 24, 7455–7464. [Google Scholar] [CrossRef] [Green Version]
- Du, C.; Zhang, X.; Yao, M.; Lv, K.; Wang, J.; Chen, L.; Chen, Y.; Wang, S.; Fu, P. Bcl-2 promotes metastasis through the epithelial-to-mesenchymal transition in the BCap37 medullary breast cancer cell line. Oncol Lett. 2018, 15, 8991–8998. [Google Scholar] [CrossRef] [Green Version]
- Grille, S.J.; Bellacosa, A.; Upson, J.; Klein-Szanto, A.J.; van Roy, F.; Lee-Kwon, W.; Donowitz, M.; Tsichlis, F.N.; Larue, L. The protein kinase Akt induces epithelial mesenchymal transition and promotes enhanced motility and invasiveness of squamous cell carcinoma lines. Cancer Res. 2003, 63, 2172–2178. [Google Scholar]
- Cheong, H.T.; Xu, F.; Choy, C.T.; Hui, C.W.C.; Mok, T.S.K.; Wong, C.W. Upregulation of Bcl2 in NSCLC with acquired resistance to EGFR-TKI. Oncol Lett. 2018, 15, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, S.; Luo, T.; Yang, Y.; Li, Y.; Yang, J.; Xu, J.; Zheng, J.; Zeng, Y. Naa10p and IKKα interaction regulates EMT in oral squamous cell carcinoma via TGF-β1/Smad pathway. J. Cell. Mol. Med. 2021, 25, 6760–6772. [Google Scholar] [CrossRef] [PubMed]
- Brandl, M.; Seidler, B.; Haller, F.; Adamski, J.; Schmid, R.M.; Saur, D.; Schneider, G. IKK(α) controls canonical TGF(ß)-SMAD signaling to regulate genes expressing SNAIL and SLUG during EMT in panc1 cells. J. Cell Sci. 2010, 123 Pt 24, 4231–4239. [Google Scholar] [CrossRef] [Green Version]
- Alam, S.K.; Astone, M.; Liu, P.; Hall, S.R.; Coyle, A.M.; Dankert, E.N.; Hoffman, D.K.; Zhang, W.; Kuang, R.; Roden, A.C.; et al. DARPP-32 and t-DARPP promote non-small cell lung cancer growth through regulation of IKKalpha-dependent cell migration. Commun. Biol. 2018, 1, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufhold, S.; Bonavida, B. Central role of Snail1 in the regulation of EMT and resistance in cancer: A target for therapeutic intervention. J. Exp. Clin. Cancer Res. 2014, 33, 62. [Google Scholar] [CrossRef] [PubMed]
- Toll, A.; Salgado, R.; Yébenes, M.; Martín-Ezquerra, G.; Gilaberte, M.; Baró, T.; Solé, F.; Alameda, F.; Espinet, B.; Pujol, R.M. MYC gene numerical aberrations in actinic keratosis and cutaneous squamous cell carcinoma. Br. J. Dermatol. 2009, 161, 1112–1118. [Google Scholar] [CrossRef]
- Stoler, A.B.; Stenback, F.; Balmain, A. The conversion of mouse skin squamous cell carcinomas to spindle cell carcinomas is a recessive event. J. Cell Biol. 1993, 122, 1103–1117. [Google Scholar] [CrossRef]
- Vreka, M.; Lilis, I.; Papageorgopoulou, M.; Giotopoulou, G.A.; Lianou, M.; Giopanou, I.; Kanellakis, N.I.; Spella, M.; Agalioti, T.; Armenis, V.; et al. IkappaB Kinase alpha is required for development and progression of KRAS-mutant lung adenocarcinoma. Cancer Res. 2018, 78, 2939–2951. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Tye, L.; Bresnick, E.; Birt, D.F. Inhibitory effect of apigenin, a plant flavonoid, on epidermal ornithine decarboxylase and skin tumor promotion in mice. Cancer Res. 1990, 50, 499–502. [Google Scholar]
- Birt, D.F.; Mitchell, D.; Gold, B.; Pour, P.; Pinchet, H.C. Inhibition of ultraviolet light induced skin carcinogenesis in SKH-1 mice by apigenin, a plant flavonoid. Anticancer Res. 1997, 17, 85–91. [Google Scholar]
- Sharma, H.; Kanwal, R.; Bhaskaran, N.; Gupta, S. Plant flavone apigenin binds to nucleic acid bases and reduces oxidative DNA damage in prostate epithelial cells. PLoS ONE 2014, 9, e91588. [Google Scholar] [CrossRef] [Green Version]
- Arango, D.; Morohashi, K.; Yilmaz, A.; Kuramochi, K.; Parihar, A.; Brahimaj, B.; Grotewold, E.; Doseff, A.I. Molecular basis for the action of a dietary flavonoid revealed by the comprehensive identification of apigenin human targets. Proc. Natl. Acad. Sci. USA 2013, 110, E2153–E2162. [Google Scholar] [CrossRef] [Green Version]
- Khan-Malek, R.; Wang, Y. Statistical analysis of quantitative RT-PCR results. Methods Mol. Biol. 2017, 1641, 281–296. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotides | Sequence |
|---|---|
| Snail1 (forward) | CTTGTGTCTGCACGACCTGT |
| Snail1 (reverse) | GGAGCAGGAGAATGGCTTC |
| mIKKα (forward) | CCCTCCAGTATCAGCATGGC |
| mIKKα (reverse) | GTGCTAACGTCTCTCACACA |
| Vimentin (forward) | CCAACCTTTTCTTCCCTGAAC |
| Vimentin (reverse) | TTGAGTGGGTGTCAACCAGA |
| TBP (forward) | GGGAGAATCATGGACCAGAA |
| TBP (reverse) | GATGGGAATTCCAGGAGTCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-García, V.A.; Alameda, J.P.; Page, A.; Mérida-García, A.; Navarro, M.; Tejero, A.; Paramio, J.M.; García-Fernández, R.A.; Casanova, M.L. IKKα Induces Epithelial–Mesenchymal Changes in Mouse Skin Carcinoma Cells That Can Be Partially Reversed by Apigenin. Int. J. Mol. Sci. 2022, 23, 1375. https://doi.org/10.3390/ijms23031375
García-García VA, Alameda JP, Page A, Mérida-García A, Navarro M, Tejero A, Paramio JM, García-Fernández RA, Casanova ML. IKKα Induces Epithelial–Mesenchymal Changes in Mouse Skin Carcinoma Cells That Can Be Partially Reversed by Apigenin. International Journal of Molecular Sciences. 2022; 23(3):1375. https://doi.org/10.3390/ijms23031375
Chicago/Turabian StyleGarcía-García, Verónica A., Josefa P. Alameda, Angustias Page, Antonio Mérida-García, Manuel Navarro, Adrián Tejero, Jesús M. Paramio, Rosa A. García-Fernández, and M. Llanos Casanova. 2022. "IKKα Induces Epithelial–Mesenchymal Changes in Mouse Skin Carcinoma Cells That Can Be Partially Reversed by Apigenin" International Journal of Molecular Sciences 23, no. 3: 1375. https://doi.org/10.3390/ijms23031375