1. Introduction
The G protein-coupled receptor kinases (GRKs) were discovered as cytosolic, membrane-associated serine/threonine kinases. Through phosphorylation of ligand-activated G protein-coupled receptors (GPCRs), GRKs enable the binding of arrestins and induce desensitization as well as internalization of the receptor [
1]. The human genome encodes seven GRKs (GRK1-7) that are grouped into three subfamilies: the visual GRK subfamily (GRK1 and 7), the GRK2 subfamily (GRK2 and 3), and the GRK4 subfamily (GRK4, 5, and 6). Four of those GRKs, namely GRK2, 3, 5, and 6 are reported to be ubiquitously expressed [
2].
Today, GRKs are not only known to phosphorylate GPCRs, but have also been demonstrated to act on other substrates, such as receptor tyrosine kinases, cytoplasmic kinases (e.g., src-family kinases), and even nuclear proteins [
3,
4,
5]. Moreover, GRKs were reported to have phosphorylation-independent scaffolding properties [
6,
7,
8]. All these activities together explain why alterations of GRK expression levels are found to be important in many pathological conditions, such as cancer, malaria, Parkinson’s-, cardiovascular-, and metabolic disease [
9,
10,
11,
12].
Taking a closer look at the distribution of the so-called ubiquitously expressed GRKs in specific tissues, it becomes apparent that there are actually striking differences in the expression levels when comparing mRNA data [
13]. However, the relationship between mRNA abundance and actual expressed protein level is not always linear and depends on many different factors, such as availability of components for biosynthesis or proteasomal degradation [
14,
15]. In standard laboratory procedures, Western blot is a commonly used technique to investigate the actual protein level expressed in cells or tissues. Various companies offer antibodies that are advertised to specifically detect certain proteins. Unfortunately, in addition to the intended protein, many of these commercially available antibodies cause unspecific background bands, which leads to difficult interpretation; in the worst case, some antibodies fail to detect their target protein at all [
16,
17,
18,
19,
20,
21]. Here, we investigated the ability of eight different anti-GRK antibodies to detect the targeted GRK isoform and possible cross-reactivity against other GRK family members. We have created expression constructs for all four ubiquitously expressed human GRKs (GRK2, 3, 5, and 6) in various isoforms (
Table 1), including versions with point mutations rendering them catalytically inactive (“kinase dead”: GRK2-K220R, GRK3-1-K220R, GRK5-K215R, and GRK6-1-K215R). We utilized these expression plasmids to overexpress the GRKs in HEK293 cells and determined the ability of selected commercially available antibodies to detect the proteins.
The use of validated isoform-specific antibodies to detect protein levels in cell lines or primary tissues allows the comparison of protein levels of one specific protein, but the direct comparison of expression levels of proteins or isoforms using different antibodies is not possible: antibody concentration, specificity, and clonality might have unpredictable influences on the result, e.g., a higher concentration of one antibody would result in more signal, although the protein content was identical. To overcome this problem, we established the cost-effective simple tag-guided analysis of relative protein abundance (STARPA). This method allows the relative quantification of different protein isoforms via Western blotting with validated antibodies.
2. Results
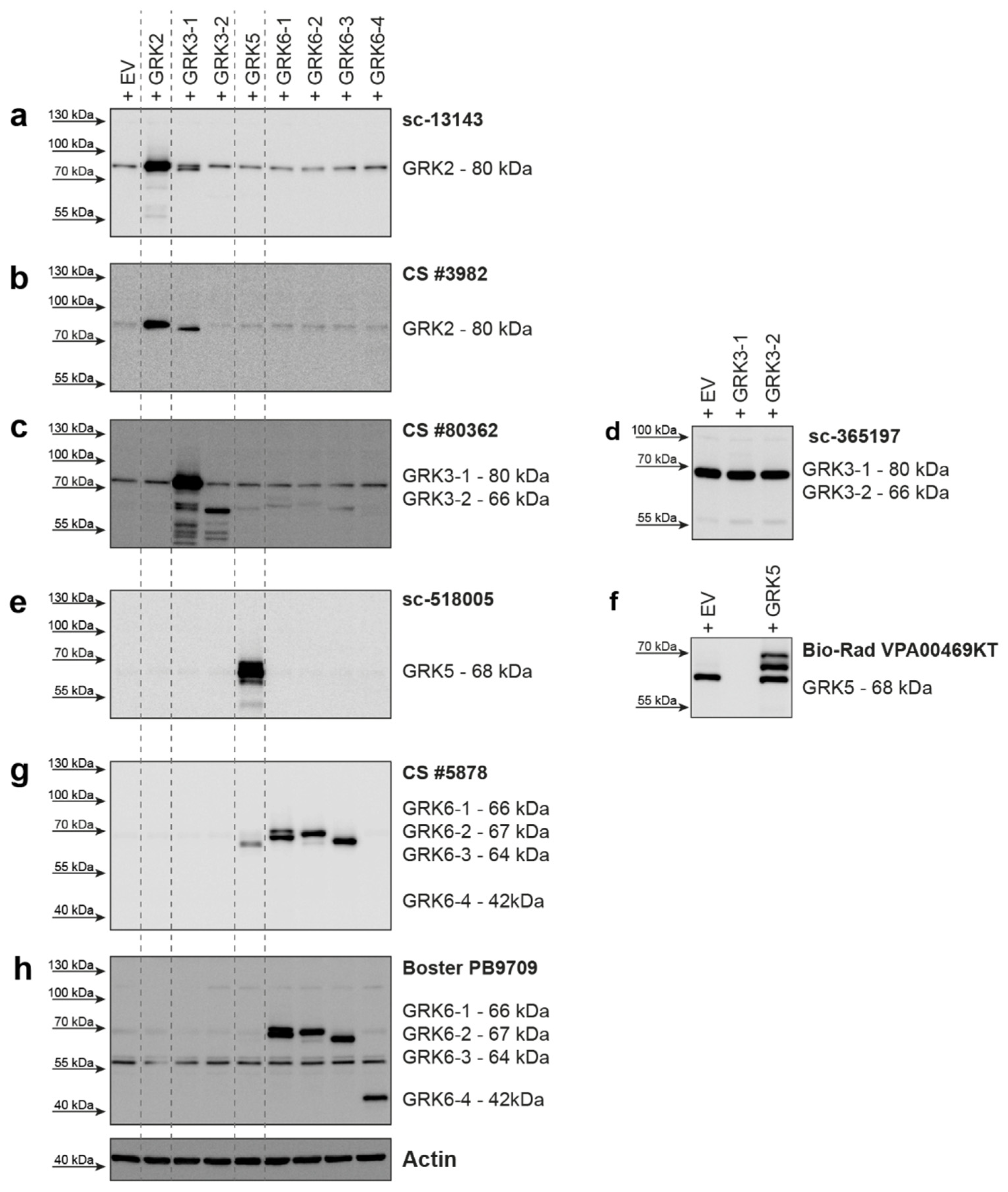
Lysates of HEK293 cells with overexpression of GRK2, GRK3-1, GRK3-2, GRK5, GRK6-1, GRK6-2, GRK6-3, and GRK6-4 were prepared and analyzed using eight different commercially available GRK antibodies as listed in
Table 1.
For GRK2, two different antibodies were tested (
Figure 1a,b). The Santa Cruz Biotechnology antibody raised against GRK2 (
Figure 1a) detected overexpressed GRK2 and endogenous levels of GRK2, with some detection of GRK3-1. The Cell Signaling Technology antibody #3982 (
Figure 1b) was also able to detect overexpressed GRK2, but the signal for endogenously expressed GRK2 was weak. This antibody detected overexpressed GRK3-1 with a stronger signal than the Santa Cruz antibody. The differences in the cross-reactivity of these two different GRK2 antibodies with the GRK3-1 protein are very surprising, as the alignment of the GRK2 epitope regions compared to the GRK3-1 sequence showed 78% (Santa Cruz antibody,
Supplementary Figure S1a) or 77% (Cell Signaling Technology antibody,
Supplementary Figure S1b) identical amino acids. While the similarity between these two GRK isoforms can explain the occurrence of cross-reactivity, the relative difference in the extent of cross-reactivity cannot be explained by this alignment.
The overexpression of GRK3-1 and GRK3-2 was detected by the Cell Signaling Technology antibody #80362 (
Figure 1c). This antibody also mildly detected overexpressed GRK5, GRK6-1, 6-2, and 6-3 isoforms, although only 34–35% of the amino acids in the epitope region match (
Supplementary Figure S1c). The Santa Cruz Biotechnology antibody sc-365197 was not able to detect the overexpressed GRK3 isoforms, but it gave a strong background band in all HEK293 lysates slightly below the 70 kDa marker (
Figure 1d).
Overexpressed GRK5 protein was strongly detected by Santa Cruz Biotechnology sc-518005 antibody with no visible cross-reactivity to overexpressed GRK6 isoforms (
Figure 1e). A second tested antibody from Bio-Rad (VPA00469KT) was also able to detect overexpressed GRK5, but additionally displayed a strong background band slightly below the specific protein band (
Figure 1f). Our findings were reported to the supplier and the antibody was discontinued.
We created expression plasmids for four different GRK6 isoforms. GRK6-1, GRK6-2, and GRK6-3 only differ in the 30 amino acids on the C-terminus, whereas GRK6-4 is a 210 amino acid N-terminally truncated version of GRK6-1. We tested two antibodies for this GRK. The antibody from Cell Signaling Technologies (#5878) raised against the N-terminus of GRK6-1, which is identical in GRK6-2 and 6-3, was able to detect overexpressed proteins of these isoforms, but was unable to detect GRK6-4 (
Figure 1g), as this isoform does not include the epitope of this antibody. We observed a slight cross-reactivity with overexpressed GRK5, which has 59% similarity in the amino acid sequence surrounding glutamate 89 of GRK6, which, according to the supplier, was used as the antigen for antibody generation (
Supplementary Figure S1d). The second antibody we tested from Boster Biological Technology (PB9709) was raised against amino acids 382-417 of GRK6. This region is also present in GRK6-4. As expected, this antibody also detected GRK6-4 and the isoforms GRK6-1, 6-2, and 6-3. However, it gave a strong unspecific band around 55 kDa (
Figure 1h).
The Western blots of overexpressed GRK5 and GRK6-1 (
Figure 1e–h) revealed double bands of the proteins. Notably, GRKs are known to be (auto)phosphorylated [
4,
22,
23]. To clarify whether the identified size shift was caused by phosphorylation of the GRKs, we transfected HEK293 cells with either GRK2, 3, 5, and 6 isoforms or with the catalytically inactive variants GRK2-K220R, GRK3-1-K220R, GRK5-K215R, and GRK6-1-K215R. These cells were then lysed in the presence of phosphatase inhibitors (+) or in the absence of EDTA and phosphatase inhibitors (−) to allow endogenous phosphatases to dephosphorylate the proteins while preparing the lysates. In samples overexpressing GRK2 (
Supplementary Figure S2a) or GRK3 (
Supplementary Figure S2b), we could not detect an influence of the endogenous phosphatases or the expression of the kinase-dead mutants on the Western blot pattern. Since the upper band is strongly diminished in the lysates with the active phosphatases (
Supplementary Figure S2c), the Western blot analysis of lysates from GRK5-overexpressing cells revealed that the upper band visible in the lysates of wild-type GRK5 treated with phosphatase inhibitors is due to phosphorylation of the kinase. The kinase-dead mutant did not show any change with or without active phosphatases. Similar findings were obtained in the case of GRK6-1 with both tested antibodies (
Supplementary Figure S2d,e). These findings indicate a strong phosphorylation, which led to a size shift of GRK5 and 6 in our experimental setup.
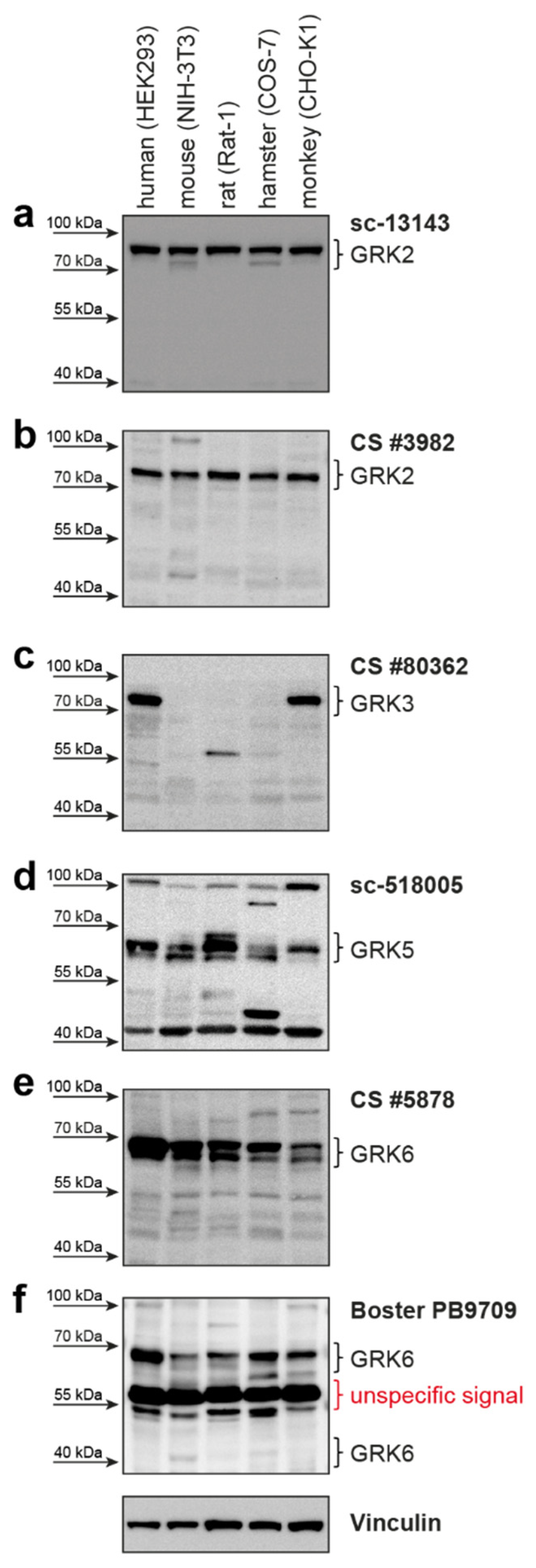
The manufacturer claims the antibodies can detect the respective GRK not only in humans, but also in other species, such as mice, hamsters, rats, and monkeys (
Table 1). Therefore, we tested the ability of the GRK antibodies to detect endogenously expressed GRKs in HEK293 (human), NIH-3T3 (mouse), COS-7 (hamster), Rat-1 (rat), and CHO-K1 (monkey) cell lines. All these species express different GRK isoforms homologous to the human GRKs (
Supplementary Table S1). Usage of both GRK2 antibodies resulted in a clear signal corresponding to the size of human GRK2 in all analyzed cell lines (
Figure 2a,b). The amino acid sequence alignment in the respective regions of antibody recognition shows 96% to 100% identity to the human GRK2 sequence (
Supplementary Figure S3a,b).
The GRK3 antibody only detected the endogenously expressed GRK3 in the human and monkey cell lines (
Figure 2c). These two species have the identical amino acid sequence in the antibody detection region. The amino acid sequences of mouse, rat, and hamster GRK3, however, are more variable, with only 89%, 88%, and 90% identity, respectively (
Supplementary Figure S4). These differences might explain the absence of a specific GRK3 band. However, GRK3 might be absent in these analyzed cell lines under our experimental conditions and is therefore not detectable.
Utilization of the GRK5 antibody leads to detectable signals in all tested cell lines from the different species (
Figure 2d) at the expected protein size. The similarity to human GRK5 is 87–95% (
Supplementary Figure S5).
A signal corresponding to the size of human GRK6 can be detected in all analyzed cell lines with both tested GRK6 antibodies (
Figure 2e,f). Notably, there is no clearly visible band reflecting the GRK6-4 isoform (
Figure 2f). The GRK6 sequences of the different species are very similar to human GRK6-1—ranging from 92% to 96% similarity for the epitope region of the Cell Signaling Technology antibody (
Supplementary Figure S6a) and 88–100% similarity for the Boster Biological Technology antibody (
Supplementary Figure S6b).
Next, using our quadruple GRK knockout HEK293 cells (ΔQ-GRK, lacking endogenous expression of GRK2, 3, 5, and 6 [
24]), we established the cost-effective simple tag-guided analysis of relative protein abundance (STARPA). This method allows the relative quantification of GRK protein amount in Western blot samples with unknown GRK isoform expression levels.
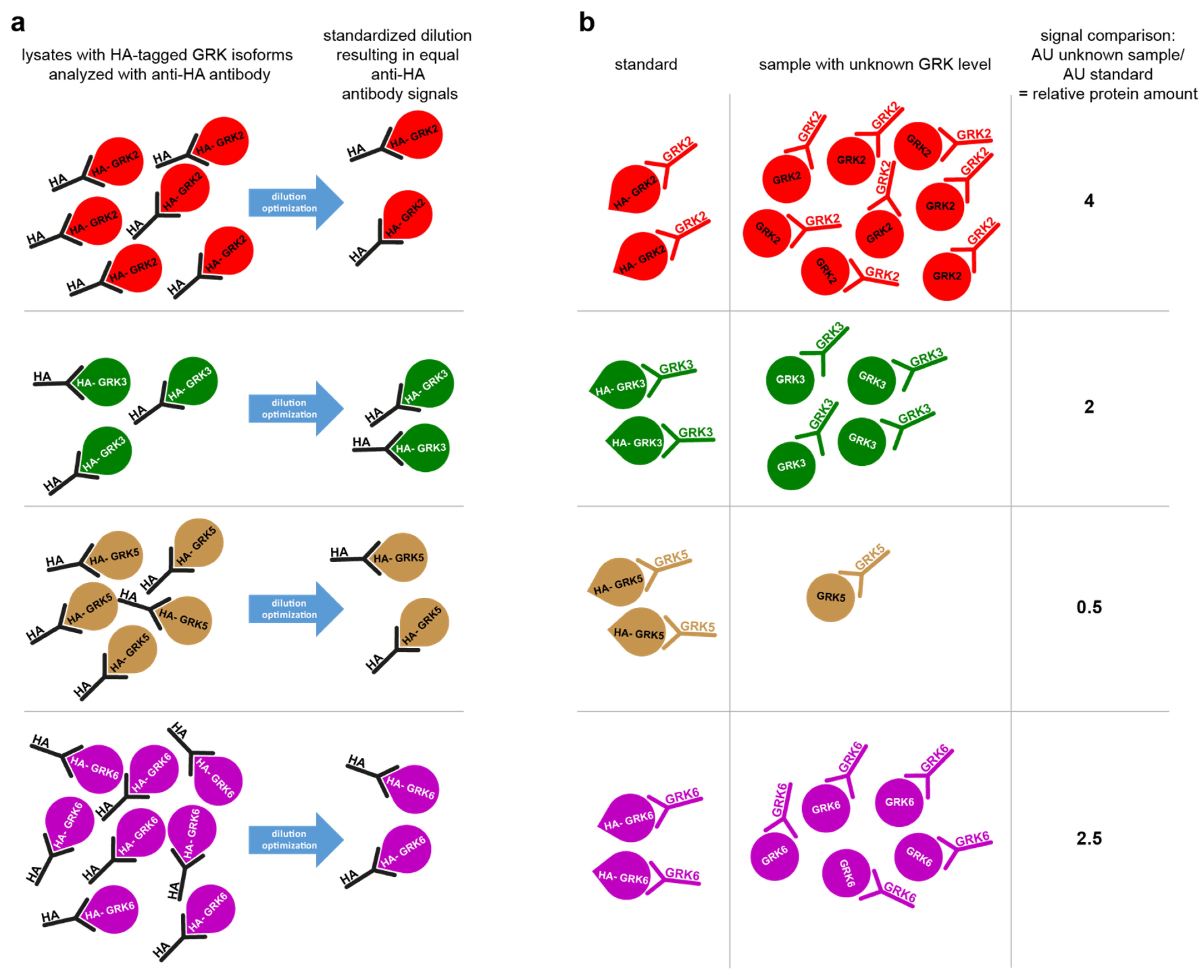
Figure 3a,b schematically depicts the principle STARPA concept: after transfection of the hemagglutinin-tagged GRK (HA-GRK) isoforms, cell lysates are analyzed for their anti-HA-antibody signal, and the optimal dilution factor to result in equal signals for all HA-GRK isoforms is experimentally determined by sample dilution (
Figure 3a). These standardized samples are loaded onto the gel as references alongside the lysates with unknown GRK expression. Utilizing the GRK-specific antibodies, the normalized signal will reflect the relative amount of this specific isoform (
Figure 3b), Equation (1):
where
is the relative amount of protein
in the unknown sample
, and
is the densitometric signal acquired with antibody
in the STARPA standard of protein
(
, all measured on the same blot. For example:
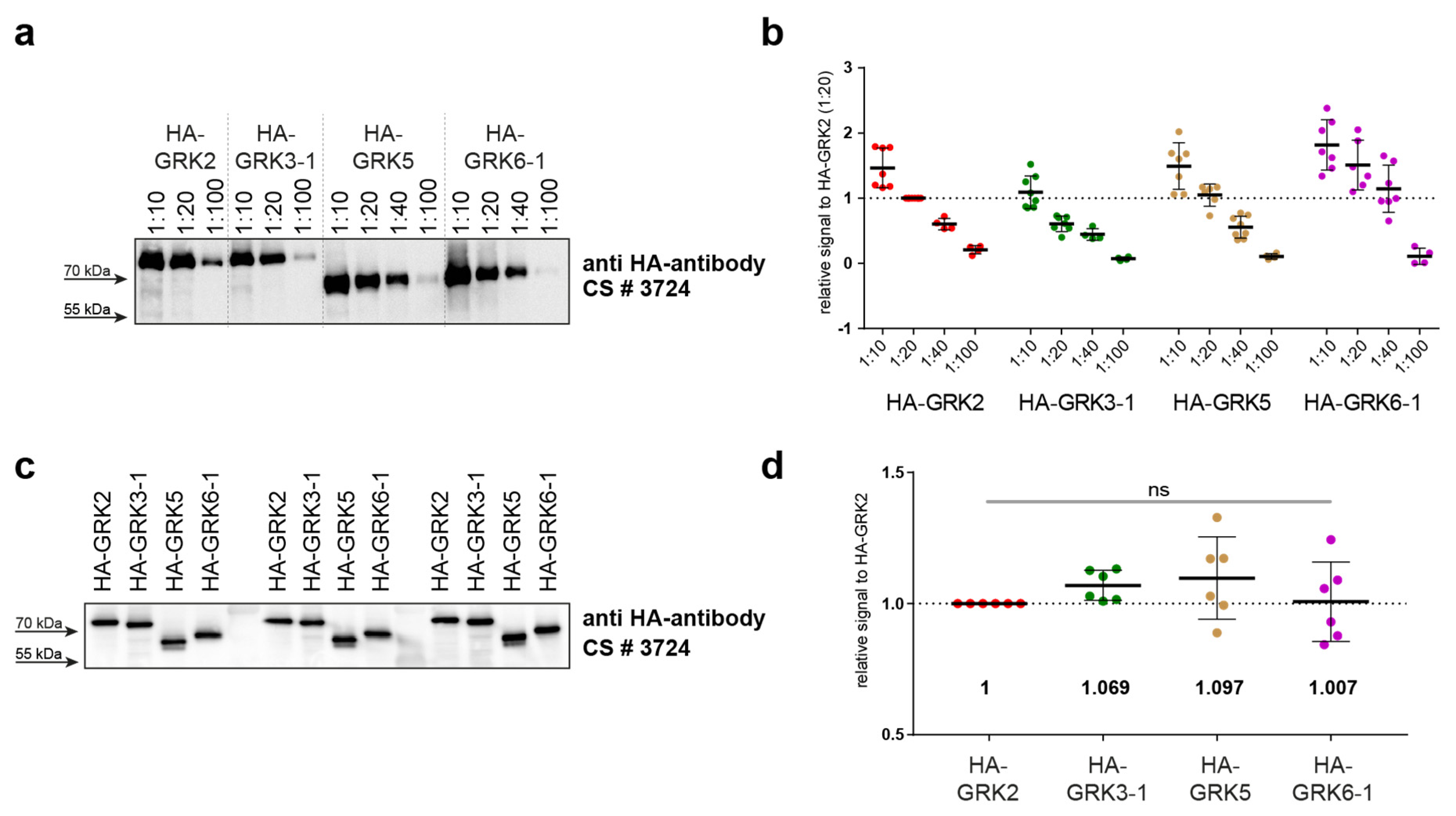
First, we prepared lysates of overexpressed HA-tagged GRK isoforms in ΔQ-GRK cells, as these are devoid of endogenous GRK expression. Therefore, any Western blot signal of GRKs would be caused by the overexpressed HA-GRK versions, and no purification steps were necessary. We analyzed a dilution series ranging from 1:10 to 1:100 by Western blotting using an anti-HA antibody (
Figure 4a). Quantification of multiple analyses of several dilutions allowed the estimation of optimal standard dilutions (
Figure 4b). In our setup, we chose the HA-GRK2 1:20 dilution as reference. In order to minimize differences in developing times between different blots, all signals obtained for the individual blots were calculated relative to this HA-GRK2 signal. We subsequently identified the dilutions of the other GRK isoforms that displayed the smallest differences in HA signals compared to our GRK2 reference dilution (namely GRK3 1:10, GRK5 1:20, and GRK6 1:40). Next, new dilution series for each GRK were prepared in order to more finely optimize results; these were again analyzed by immunoblotting (data not shown). The best-matching dilutions were then extensively validated (
Figure 4c,d).
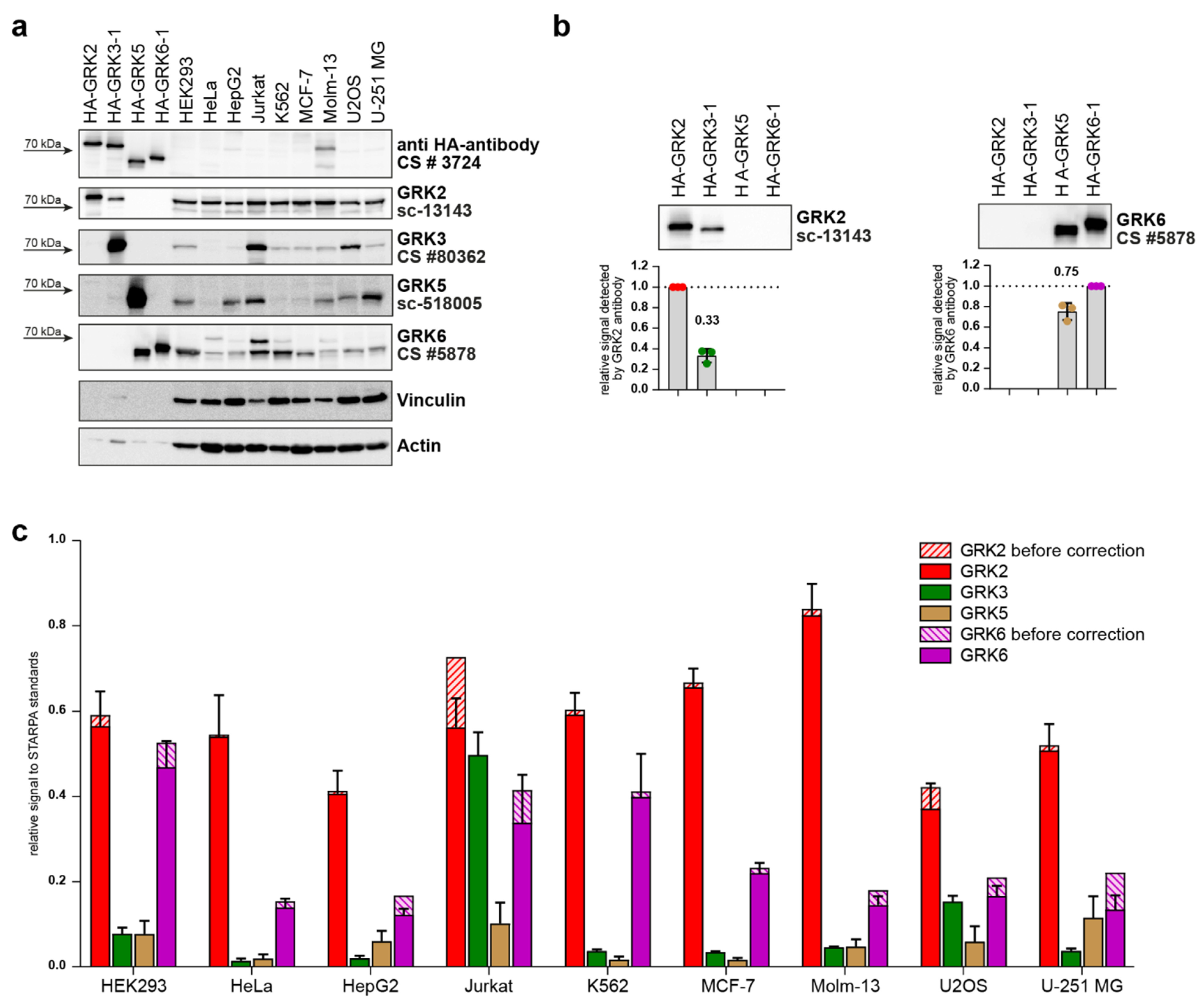
These validated standards were then used to determine relative GRK protein levels in nine different commonly used cell lines. Thus, standards were loaded onto Western blot gels in parallel with 30 µg of total protein from HEK293, HeLa, HepG2, Jurkat, K562, MCF-7, Molm-13, U2OS, and U-251 MG cell lysates with unknown GRK content (
Figure 5a). The samples were analyzed with GRK-specific antibodies as indicated, and additionally with anti-HA antibody. The previously described cross-reactivity of the utilized GRK2 (sc-13143) and GRK6 (CS #5878) antibodies against GRK3 and GRK5, respectively, while weakly observed in the non-standardized lysates (
Figure 1a,g;
Table 1), were clearly visible in the standards with an equal amount of HA-GRK (
Figure 5a,b). Quantification of GRK3 signal detected by the GRK2 antibody revealed cross-reactivity resulting in a 33% signal, and for GRK5 detection by GRK6 antibody a 75% signal (
Figure 5b). These levels of cross-reactivity need to be considered when calculating relative protein levels using STARPA. Therefore, the cross-reactivity was quantified as a cross-reactivity coefficient for antibody
using the STARPA standards, Equation (2):
where the densitometric signal of STARPA standard
detected by antibody
(
) divided by the densitometric signal of STARPA standard
detected by antibody
(
) results in the cross-reactivity coefficient for antibody
towards protein
(
). For example, calculating the cross-reactivity coefficient for the anti-GRK2 antibody (sc-13143) exhibiting cross-reactivity towards GRK3 (
):
Consequently, when determining the relative protein content of a specific protein using antibody
, which cross-reacts with protein
, the real protein content of
can only be calculated if the amount of
is subtracted accordingly. Thus, Equation (1) is modified to include the cross-reactivity coefficient, Equation (3):
where
is the relative amount of protein
in sample
corrected for the cross-reactivity of antibody
towards protein
. Continuing the example above:
After quantification of the immunoblots, the cell line signals obtained from incubation with GRK-specific antibodies were divided by the signal with the GRK-specific antibody of the respective standard (Equation (1)), resulting in the relative GRK expression pattern before correction of the analyzed cell lines (
Figure 5c). Subsequently, the obtained relative expression of GRK2 and GRK6 were corrected for cross-reactivity signals as described above (Equations (2) and (3),
Figure 5c). Notably, the impact of this correction is reduced if GRK3 or GRK5 protein levels are low.
GRK2 was found to be the most expressed isoform in all nine tested cell lines. Although GRK6 was the second most abundant isoform in all cell lines except for Jurkat cells, its expression compared to the respective GRK2 level varied between the cell lines. While GRK6 protein levels reach up to 83% of GRK2 in HEK293 cells, our analysis showed that GRK6 expression is much lower in HeLa, HepG2, MCF-7, Molm-13, and U2OS cells, reaching between 18% (Molm-13) and 34% (MCF-7) of the respective GRK2 level.
In five out of nine cell lines, the measured GRK3 and GRK5 protein levels were less than a third of the respective GRK2 or GRK6 expression. In HepG2, Jurkat, U2OS, and U-251 MG cells, however, GRK3 and GRK5 levels differed. GRK5 reached 49% and 86% of the recorded GRK6 expression in HepG2 and U-251 MG cells, respectively. U2OS cells expressed similar protein levels of GRK3 and GRK6 (92% of GRK6 expression). In Jurkat cells, GRK3 was the second most abundant isoform, reaching 89% of the GRK2 protein level, in contrast to the remaining analyzed cell lines.
Taken together, the employment of STARPA demonstrates differential GRK isoform expression in these commonly utilized cell lines.
3. Discussion
Most of the tested antibodies (GRK2: sc-13143 and CS #3982; GRK3: CS #80362; GRK5: sc-518005 and VPA00469KT; GRK6: CS #5878 and PB9709) are able to detect the targeted protein, but some also strongly label background bands with similar protein size, leading to difficult interpretation of the expression levels, especially at endogenous levels (GRK5: VPA00469KT; GRK6: PB9709). One antibody (sc-365197, GRK3) did not detect its target GRK at all; it detected only a very strong background band (
Figure 1d). In our test setup, we also observed cross-reactivity with the other overexpressed GRK isoforms. This cross-reactivity is also the reason why the four standards used for STARPA could not be mixed into one standard containing all GRK isoforms. We could detect GRK3 while using GRK2 antibodies and GRK5 when using GRK6 antibody (
Figure 1a,b,g;
Figure 5a,b). This observed cross-reactivity should be taken into consideration, as the influence on the obtained result depends on the expression ratio of GRK3 to GRK2 and GRK5 to GRK6.
Apart from detecting more than one GRK isoform, a given antibody could possibly detect the same GRK in other species, as GRKs are conserved among mammals [
25]. To test whether the antibodies would recognize the proteins of mice, rats, hamsters, or monkeys, we analyzed cell lines of these species. For GRK2, GRK5, and GRK6, we detected bands in the size comparable to the human GRKs, but in the case of GRK3, we did not observe a signal in mouse, rat, or hamster cells, although the supplier states the recognition of mouse GRK3. The absence of detectable protein in the tested cell lines could be either due to the lack of antibody affinity to the species-typical GRK, or simply due to the absence of that GRK in the tested cell line. To clearly validate the recognition of GRKs from different species, it would be mandatory to test the antibodies with overexpressed isoforms of the species of interest, and/or in protein lysates of knockout (KO) animals or cell lines.
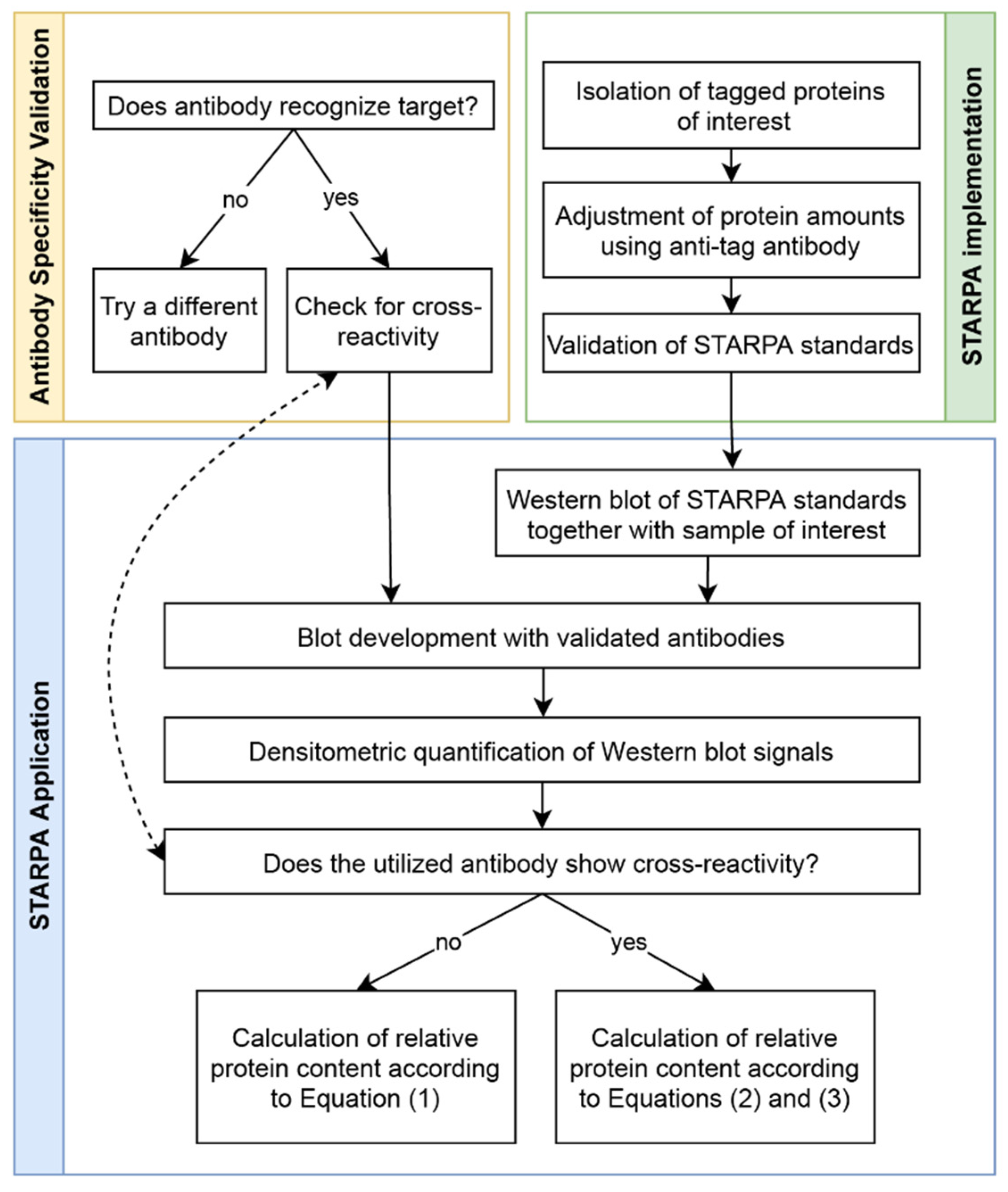
Utilizing the validated anti-GRK antibodies, we established STARPA as an economical method for relative quantification of GRK isoform levels. The sequence of events for STARPA are summarized in
Figure 6. Due to the cross-reactivity of GRK2 and GRK6 antibodies as mentioned above, recorded levels of these isoforms must be corrected. In order to calculate the real amount of GRK2 or 6, the quantification of GRK3 or 5 respectively needs to be carried out as well, using an antibody without detectable cross-reactivity against the other isoforms. For protein samples where GRK2 or GRK6 levels are much higher than GRK3 or GRK5 levels, respectively, the correction could be neglected (
Figure 5c). The protein standards are perfectly suited to determine the cross-reactivity of the utilized antibody, since the level of cross-reactivity could be over- or underestimated depending on the expression level of the different proteins under endogenous conditions.
Our analysis of nine commonly used cell lines demonstrates the ability of STARPA to reveal the differential protein expression of the GRK isoforms in cells derived from various tissues. Notably, the here-reported differences and expression patterns reflect a snapshot of the GRK levels in our growth conditions. GRK levels can change during progression of the cell cycle [
26], or the expression pattern might also be influenced by the level of available nutrients, cell density, plasticware, cell passage, and other factors [
27]. Taking all these factors into account, the expression level of a given cell line might vary between different laboratories and must be individually analyzed.
Differences in GRK expression levels in a given cellular context potentially lead to changes in downstream signaling and trafficking of GPCRs. The knockdown of GRK2/3 or GRK5/6 differentially influenced downstream extracellular-signal regulated kinases (ERK) signaling of angiotensin II type 1 receptor (AT1R) or vasopressin 2 receptor (V2R) [
28,
29]. Additionally, some GPCRs can only be internalized upon ligand stimulation if specific GRKs are expressed. For internalization of the muscarinic acetylcholine receptor 5 (M5R) and µ-opioid receptor (MOP) the presence of either GRK2 or 3 is mandatory [
24]. GPCRs are involved in numerous diseases, and GRK expression alterations have been reported for several pathophysiological conditions. In heart failure for example, the upregulation of GRK2 and GRK5 leads to the downregulation and desensitization of the beta adrenergic receptors [
30]. Hence, investigation of the impact of GRK levels on GPCR signaling becomes essential [
13]. While the expression of each isoform can be compared based on the mRNA level using standardized real-time quantitative PCR, the here-described STARPA method now allows this comparison at the protein level to obtain specific GRK expression patterns.
In this study we describe STARPA as a method that can be conducted using only standard laboratory procedures and equipment to determine relative GRK protein levels, given that antibodies recognizing the protein are available and appropriately validated. As availability of antibodies targeting a protein of interest might be limited, biologicals—as many of them are clinically used antibodies [
31]—or the newly developed nanobodies [
32] could be additionally tested.
By introducing standards, the variability of antibody concentration and affinity can be normalized and thereby allow direct comparison. Of note, all samples and the standards need to be loaded onto one Western blot gel, and the blots need to be conducted multiple times from the identical lysate to obtain reliable results. This limits the sample number that can be processed in one experimental setup.
STARPA enables the relative comparison of different expression levels of any protein that can be expressed with the same protein tag. We anticipate that STARPA is not limited to cytosolic proteins but could be applied to any protein class. Notably, GPCRs are also differentially expressed in different tissues or cancer [
33,
34]. Studies of GPCR protein levels in relation to other GPCRs, GRKs, β-arrestins, G proteins, or any other protein could be conducted using STARPA.
We used the ΔQ-GRK cell line with a KO of GRK2, 3, 5, and 6 [
24] for expression of HA-GRK isoforms. In that case, no purification of the HA-tagged GRKs was needed, as there is no endogenous GRK present. If no KO cells are available to overexpress the desired protein, an immunoprecipitation of the tagged proteins should be carried out before the creation of the dilution series.
Taken together, we strongly recommend testing any antibody with exogenously expressed proteins to clearly confirm identity of the obtained Western blot results. Furthermore, we propose STARPA as a cost-effective method using standard laboratory equipment to compare relative levels of different proteins.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}