
Maternal Neutrophil Depletion Fails to Avert Systemic Lipopolysaccharide-Induced Early Pregnancy Defects in Mice


Abstract
:1. Introduction
2. Results
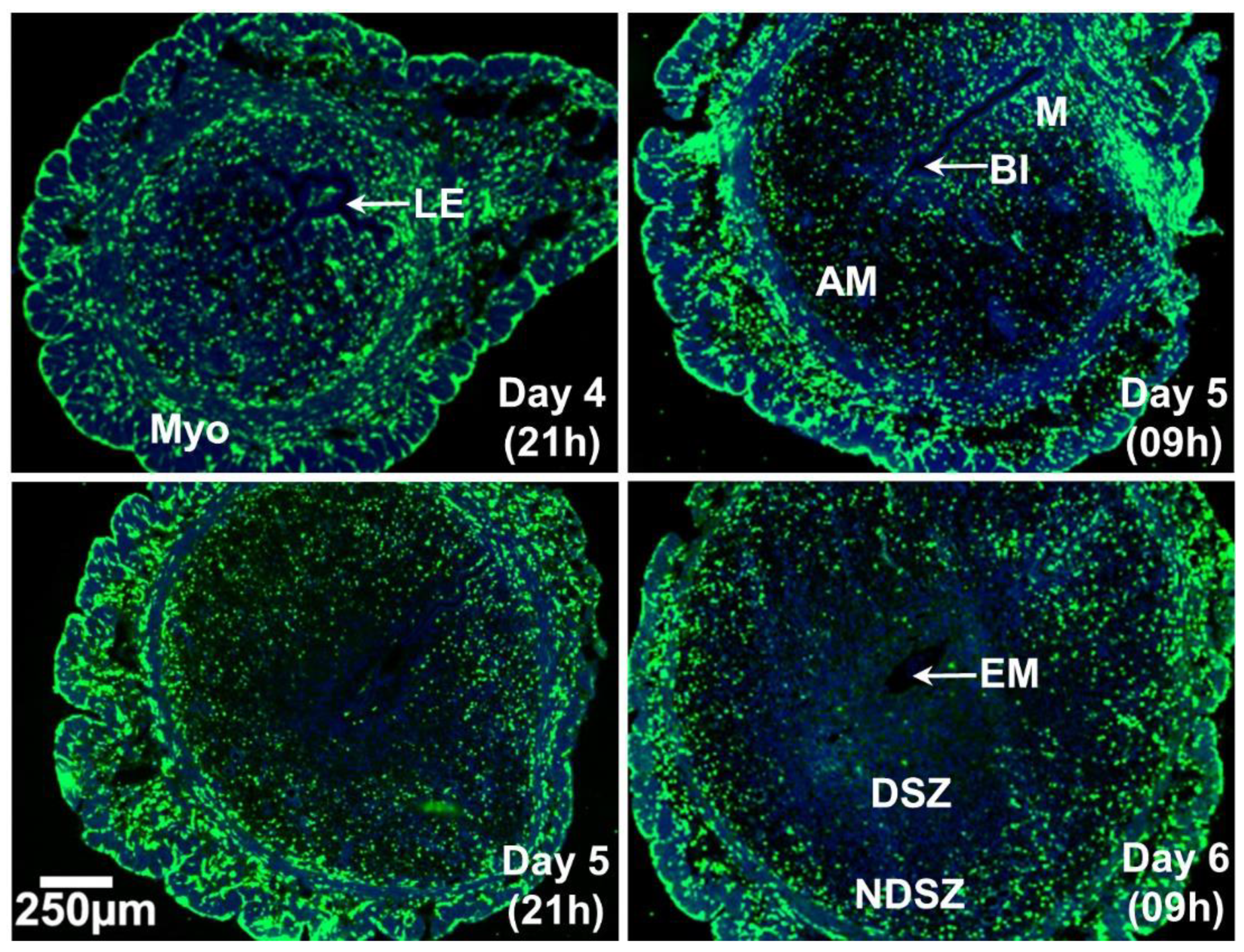
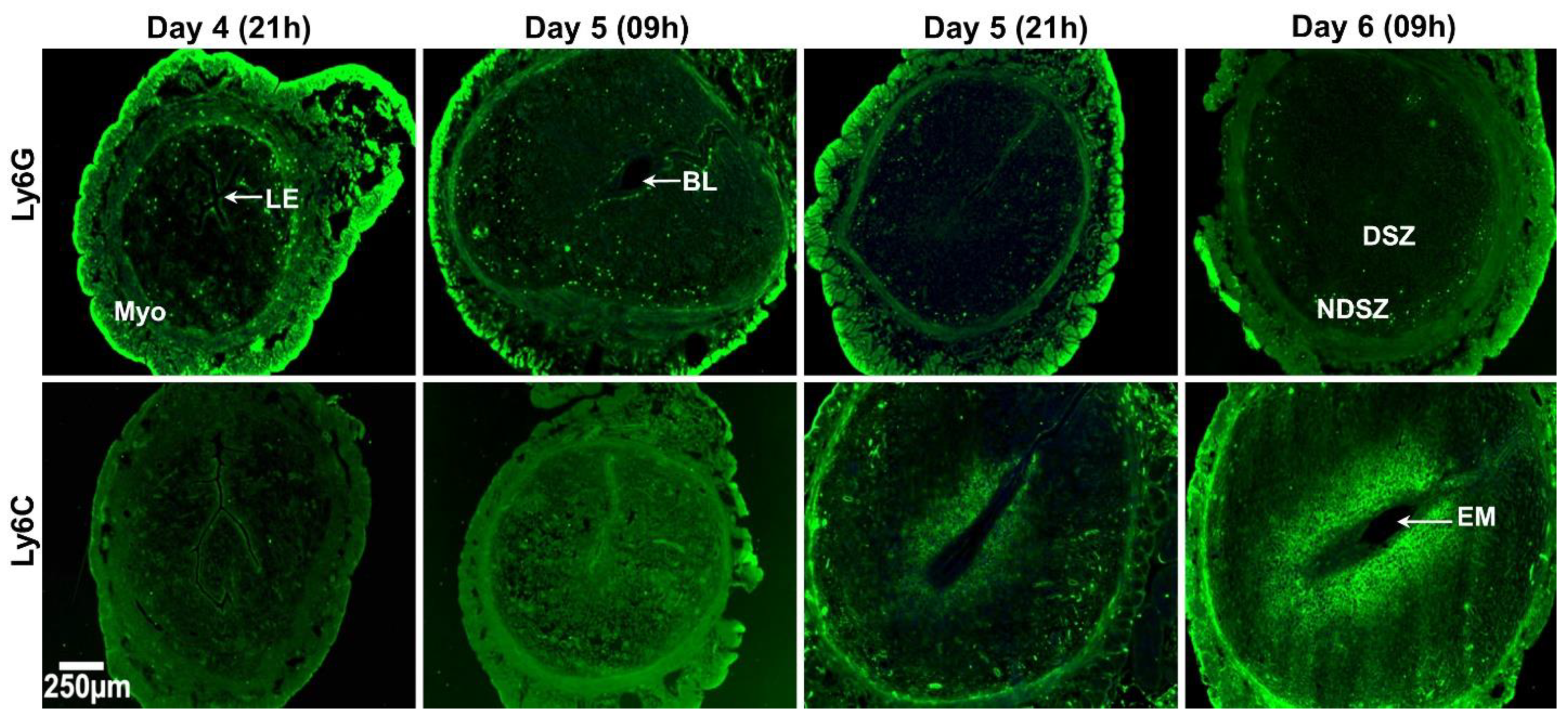
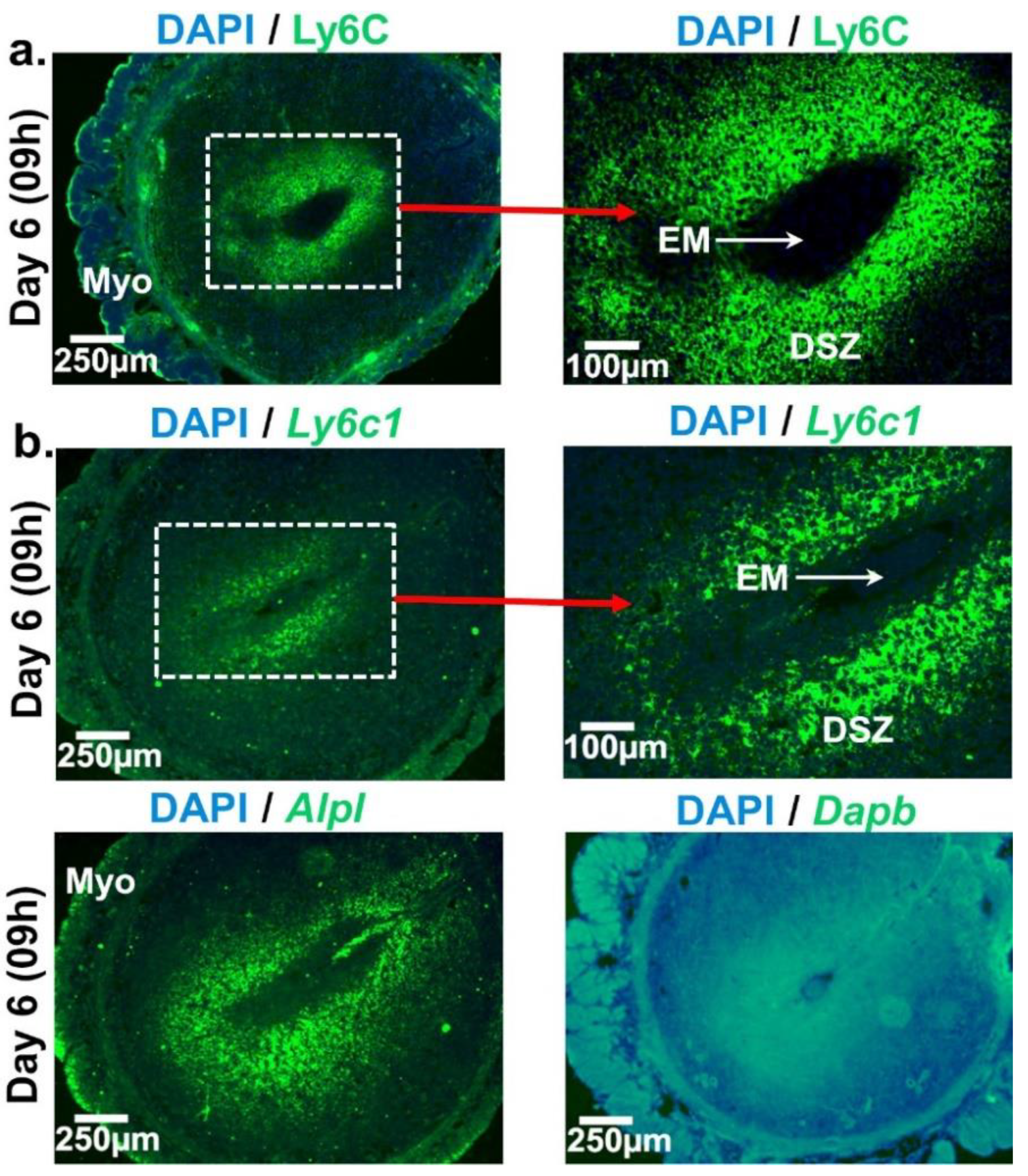
2.1. Spatio-Temporal Distribution of Leukocytes in the Day 4 Receptive Uterus and Days 5 and 6 EBISs
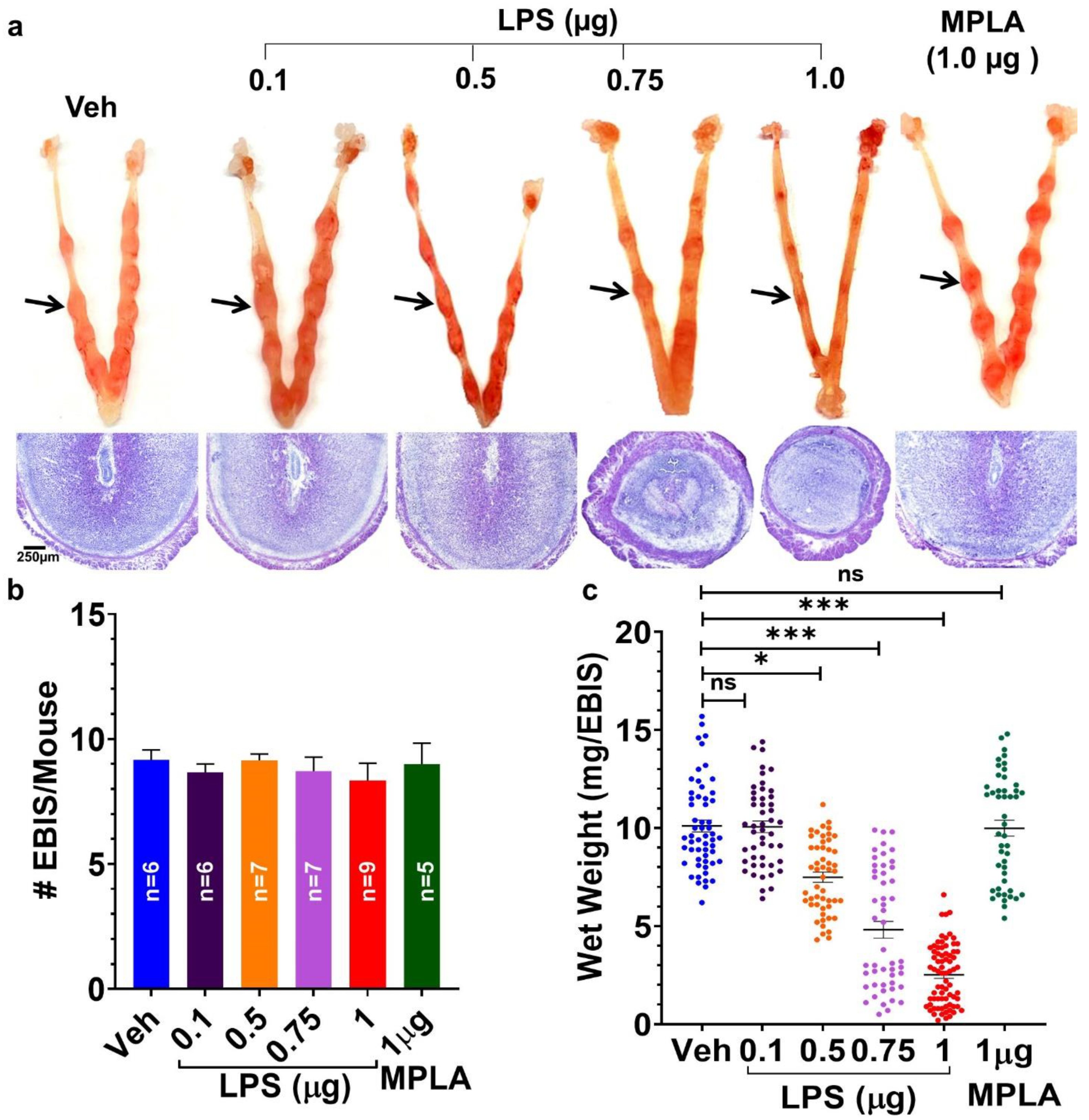
2.2. Maternal LPS-Exposure Dose-Dependently Causes Defects/Loss of EBISs
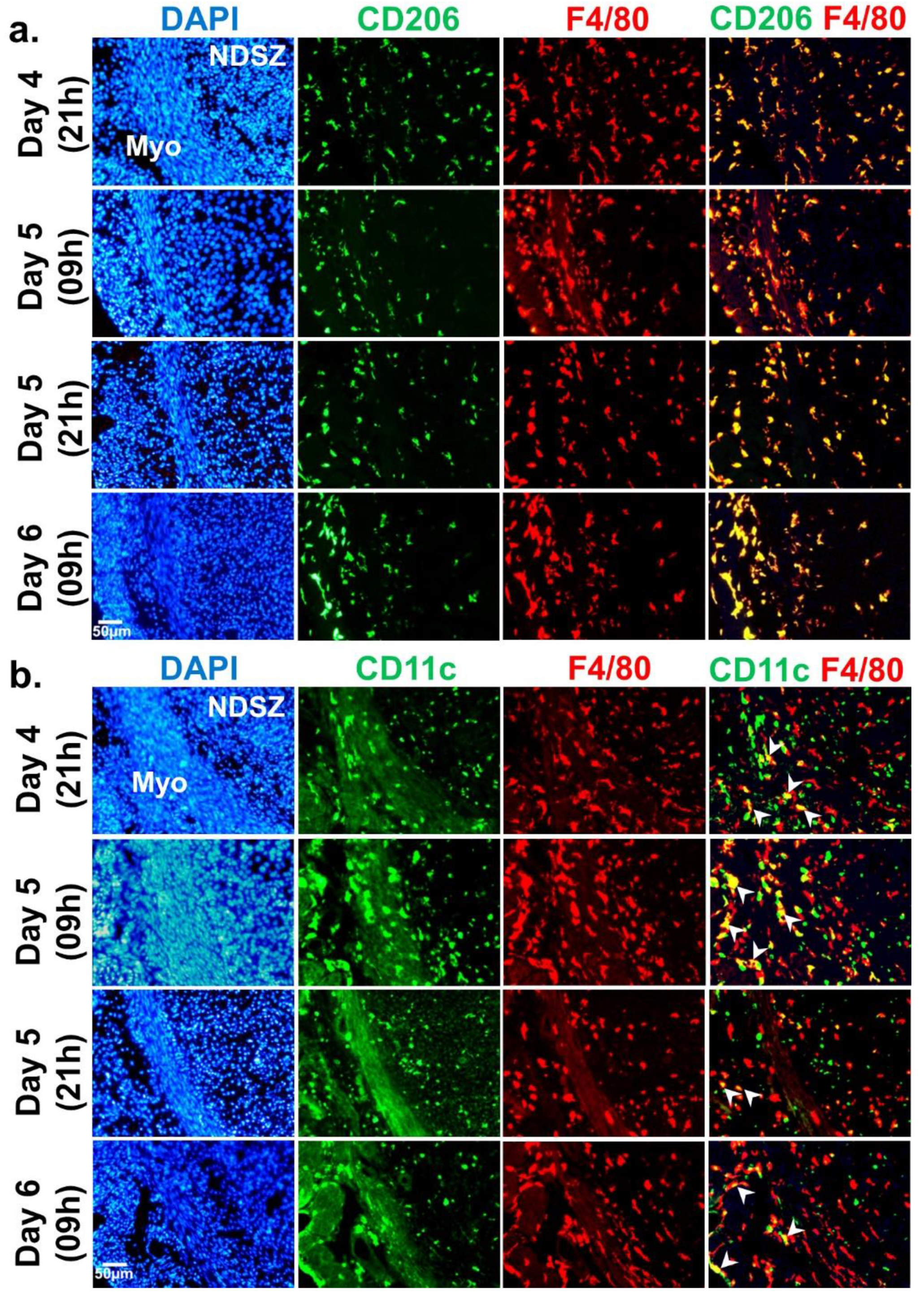
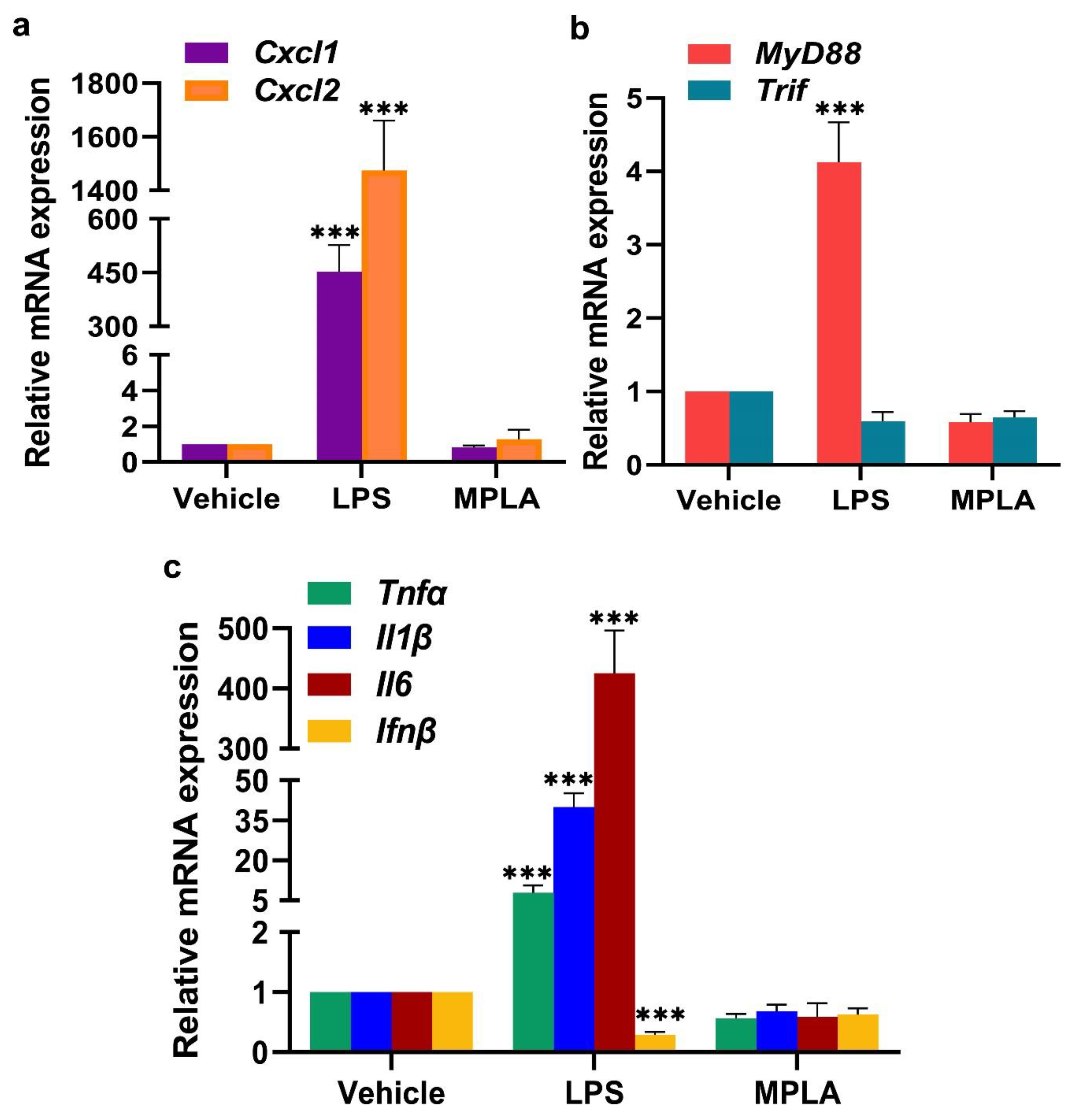
2.2.1. LPS-Induced Inflammatory Response at the Day 5 EBIS
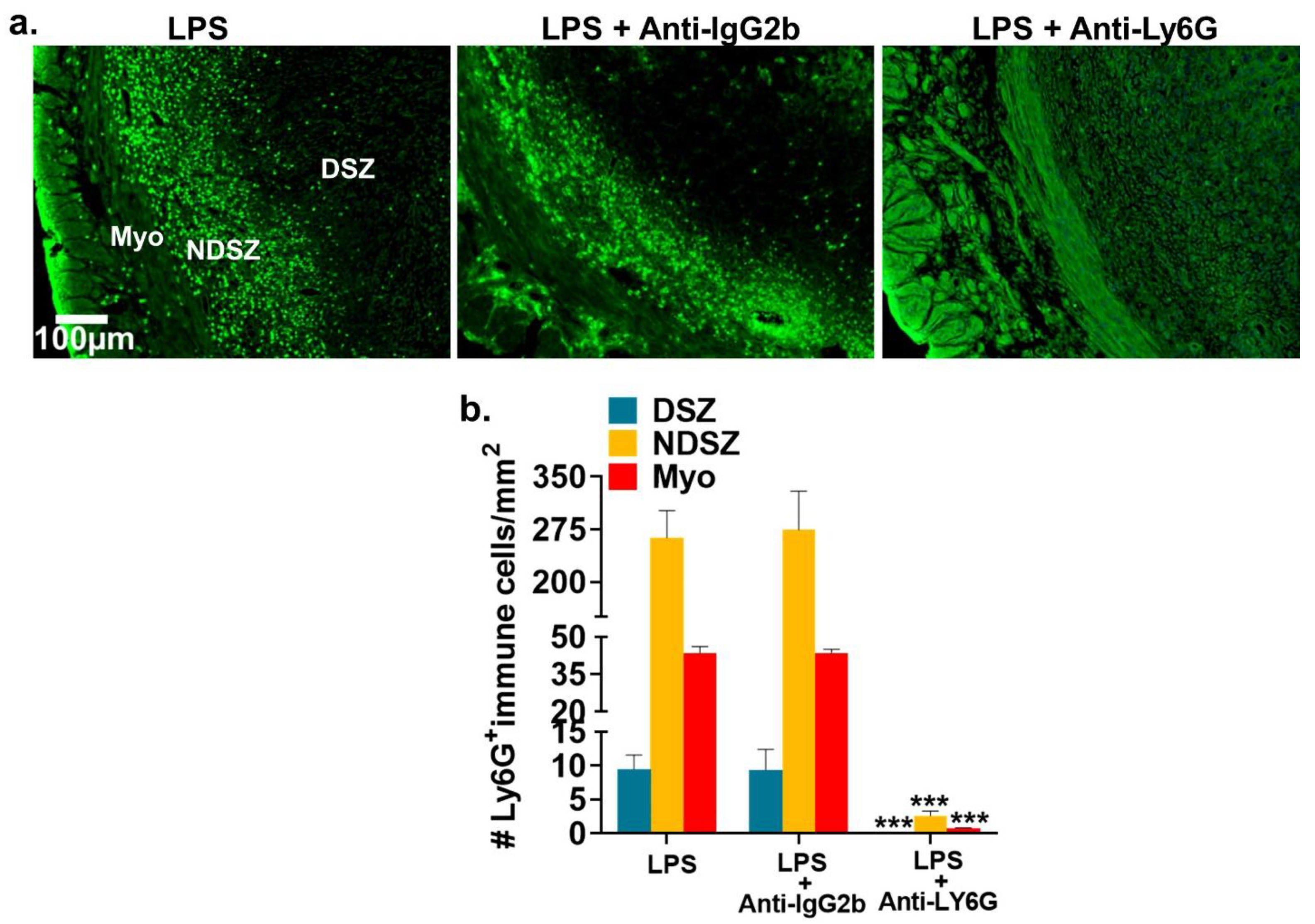
2.2.2. LPS-Mediated Neutrophil Accumulation at the EBIS
2.2.3. Administration of anti-Ly6G antibody Prevented LPS-Induced NP Accumulation at the EBIS
2.2.4. NP Depletion Prior to LPS Treatment Fails to Prevent LPS-induced EBIS Defects
3. Discussion
4. Materials and Methods
4.1. Animal and Tissue Preparation
4.2. LPS-Induced Inflammation at the EBIS and Early Pregnancy Defect
4.3. Depletion of NPs Prior to LPS Injection in the Pregnant Mice
4.4. Immunofluorescence
4.5. Quantitative RT-PCR
4.6. In Situ Localization of Ly6C mRNA
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Alpl | Tissue non-specific alkaline phosphatase |
| AM | Anti-mesometrial side |
| EBIS | Early blastocyst implantation site |
| BLs | Blastocyst |
| Cxcl1 | C-X-C motif chemokine ligand 1 |
| Cxcl2 | C-X-C motif chemokine ligand 2 |
| Dapb | Dihydrodipicolinate reductase |
| DSZs | Embryo |
| IFNβ | Interferon β |
| Il1β | Interleukin 1β |
| Il6 | Interleukin 6 |
| LE | Luminal epithelium |
| LPS | Lipopolysaccharide |
| Ly6C | Lymphocyte antigen 6 complex locus C |
| Ly6G | Lymphocyte antigen 6 complex locus G6D |
| M | Mesometrial side |
| MMP8 | Matrix metalloproteinase 8 |
| MPLAs | Monophosphoryl lipid A |
| MyD88 | Myeloid differentiation primary response 88 |
| Myos | Myometrium |
| MΦs | Macrophages |
| NDSZ | Non-decidual stromal zone |
| NKs | Natural killer cells |
| NPs | Neutrophils |
| PBS | Phosphate-buffered saline |
| ROS | Reactive oxygen species |
| SDZ | Stromal decidual zone |
| TNFα | Tumor necrosis factor-alpha |
| Trif | TIR-domain-containing adapter-inducing interferon-β |
| uNk | Uterine natural killer cells |
| Veh | Vehicle |
References
- Ford, H.B.; Schust, D.J. Recurrent Pregnancy Loss: Etiology, Diagnosis, and Therapy. Rev. Obstet. Gynecol. 2009, 2, 76–83. [Google Scholar]
- Minassian, C.; Thomas, S.L.; Williams, D.J.; Campbell, O.; Smeeth, L. Acute Maternal Infection and Risk of Pre-Eclampsia: A Population-Based Case-Control Study. PLoS ONE 2013, 8, e73047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbella, S.; Taschieri, S.; Francetti, L.; De Siena, F.; Del Fabbro, M. Periodontal disease as a risk factor for adverse pregnancy outcomes: A systematic review and meta-analysis of case–control studies. Odontology 2011, 100, 232–240. [Google Scholar] [CrossRef]
- Harris, J.; Sheiner, E. Does an Upper Respiratory Tract Infection During Pregnancy Affect Perinatal Outcomes? A Literature Review. Curr. Infect. Dis. Rep. 2013, 15, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Ramos, B.D.A.; Kanninen, T.T.; Sisti, G.; Witkin, S.S. Microorganisms in the Female Genital Tract during Pregnancy: Tolerance versus Pathogenesis. Am. J. Reprod. Immunol. 2014, 73, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Denker, H.-W. Implantation: A cell biological paradox. J. Exp. Zoöl. 1993, 266, 541–558. [Google Scholar] [CrossRef] [PubMed]
- Chavan, A.R.; Griffith, O.; Wagner, G.P. The inflammation paradox in the evolution of mammalian pregnancy: Turning a foe into a friend. Curr. Opin. Genet. Dev. 2017, 47, 24–32. [Google Scholar] [CrossRef]
- Griffith, O.W.; Chavan, A.R.; Protopapas, S.; Maziarz, J.; Romero, R.; Wagner, G.P. Embryo implantation evolved from an ancestral inflammatory attachment reaction. Proc. Natl. Acad. Sci. USA 2017, 114, E6566–E6575. [Google Scholar] [CrossRef] [Green Version]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [Green Version]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef]
- Dale, D.C.; Boxer, L.; Liles, W.C. The phagocytes: Neutrophils and monocytes. Blood 2008, 112, 935–945. [Google Scholar] [CrossRef]
- Metzemaekers, M.; Gouwy, M.; Proost, P. Neutrophil chemoattractant receptors in health and disease: Double-edged swords. Cell. Mol. Immunol. 2020, 17, 433–450. [Google Scholar] [CrossRef]
- He, B.; Zhang, H.; Wang, J.; Liu, M.; Sun, Y.; Guo, C.; Lu, J.; Wang, H.; Kong, S. Blastocyst activation engenders transcriptome reprogram affecting X-chromosome reactivation and inflammatory trigger of implantation. Proc. Natl. Acad. Sci. USA 2019, 116, 16621–16630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, J.S. Immunologically Relevant Cells in the Uterus. Biol. Reprod. 1994, 50, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Mor, G.; Cardenas, I.; Abrahams, V.; Guller, S. Inflammation and pregnancy: The role of the immune system at the implantation site. Ann. N. Y. Acad. Sci. 2011, 1221, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Collins, M.K.; Tay, C.-S.; Erlebacher, A. Dendritic cell entrapment within the pregnant uterus inhibits immune surveillance of the maternal/fetal interface in mice. J. Clin. Investig. 2009, 119, 2062–2073. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, R.; Pulendran, B. Restraining order for dendritic cells: All quiet on the fetal front. J. Clin. Investig. 2009, 119, 1854–1857. [Google Scholar] [CrossRef] [Green Version]
- Hunt, J.S.; Petroff, M.G.; Burnett, T.G. Uterine leukocytes: Key players in pregnancy. Semin. Cell Dev. Biol. 2000, 11, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Zenclussen, A.C.; Hämmerling, G.J. Cellular Regulation of the Uterine Microenvironment That Enables Embryo Implantation. Front. Immunol. 2015, 6, 321. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Lee, M.; Lee, S.K. Role of endometrial immune cells in implantation. Clin. Exp. Reprod. Med. 2011, 38, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Lima, P.D.A.; Zhang, J.; Dunk, C.; Lye, S.J.; Croy, B.A. Leukocyte driven-decidual angiogenesis in early pregnancy. Cell. Mol. Immunol. 2014, 11, 522–537. [Google Scholar] [CrossRef] [Green Version]
- Robertson, S.A.; Moldenhauer, L.M. Immunological determinants of implantation success. Int. J. Dev. Biol. 2014, 58, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, A.; Sharkey, D.J.; Robertson, S.A.; Zenclussen, A.C. Immune Cells at the Fetomaternal Interface: How the Microenvironment Modulates Immune Cells To Foster Fetal Development. J. Immunol. 2018, 201, 325–334. [Google Scholar] [CrossRef]
- Tirado-Gonzalez, I.; Barrientos, G.; Freitag, N.; Otto, T.; Thijssen, V.L.; Moschansky, P.; von Kwiatkowski, P.; Klapp, B.F.; Winterhager, E.; Bauersachs, S.; et al. Uterine NK cells are critical in shaping DC immunogenic functions compatible with pregnancy progression. PLoS ONE 2012, 7, e46755. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: Evidence for TLR4 as the Lps gene product. J. Immunol. 1999, 162, 3749–3752. [Google Scholar] [PubMed]
- Yamamoto, M.; Sato, S.; Hemmi, H.; Hoshino, K.; Kaisho, T.; Sanjo, H.; Takeuchi, O.; Sugiyama, M.; Okabe, M.; Takeda, K.; et al. Role of Adaptor TRIF in the MyD88-Independent Toll-Like Receptor Signaling Pathway. Science 2003, 301, 640–643. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol. 2005, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lei, W.; Ni, H.; Herington, J.; Reese, J.; Paria, B.C. Alkaline Phosphatase Protects Lipopolysaccharide-Induced Early Pregnancy Defects in Mice. PLoS ONE 2015, 10, e0123243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.; Wang, X.; Paria, B.; Damm, D.; Abraham, J.; Klagsbrun, M.; Andrews, G.; Dey, S. Heparin-binding EGF-like growth factor gene is induced in the mouse uterus temporally by the blastocyst solely at the site of its apposition: A possible ligand for interaction with blastocyst EGF-receptor in implantation. Development 1994, 120, 1071–1083. [Google Scholar] [CrossRef] [PubMed]
- Paria, B.C.; Huet-Hudson, Y.M.; Dey, S.K. Blastocyst’s state of activity determines the “window” of implantation in the receptive mouse uterus. Proc. Natl. Acad. Sci. USA 1993, 90, 10159–10162. [Google Scholar] [CrossRef] [Green Version]
- Nakano, A.; Harada, T.; Morikawa, S.; Kato, Y. Expression of Leukocyte Common Antigen (CD45) on Various Human Leukemia/Lymphoma Cell Lines. Acta Pathol. Jpn. 1990, 40, 107–115. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, C.J.; Kim, D.-J.; Kang, J.-H. Immune Cells in the Female Reproductive Tract. Immune Netw. 2015, 15, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Croy, B.A.; Zhang, J.; Tayade, C.; Colucci, F.; Yadi, H.; Yamada, A. Analysis of Uterine Natural Killer Cells in Mice. Methods Mol. Biol. 2010, 612, 465–503. [Google Scholar] [CrossRef] [PubMed]
- Daley, J.M.; Thomay, A.A.; Connolly, M.D.; Reichner, J.; Albina, J.E. Use of Ly6G-specific monoclonal antibody to deplete neutrophils in mice. J. Leukoc. Biol. 2007, 83, 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polletti, S.; Natoli, G. Understanding Spontaneous Conversion: The Case of the Ly6C− Monocyte. Immunity 2017, 46, 764–766. [Google Scholar] [CrossRef] [Green Version]
- Austyn, J.M.; Gordon, S. F4/80, a monoclonal antibody directed specifically against the mouse macrophage. Eur. J. Immunol. 1981, 11, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.-J.; Gu, Y.; Wang, C.-Z.; Jin, Y.; Wen, X.-M.; Ma, J.-C.; Tang, L.-J.; Mao, Z.-W.; Qian, J.; Lin, J. The M2 macrophage marker CD206: A novel prognostic indicator for acute myeloid leukemia. OncoImmunology 2020, 9, 1683347. [Google Scholar] [CrossRef] [Green Version]
- Singh-Jasuja, H.; Thiolat, A.; Ribon, M.; Boissier, M.-C.; Bessis, N.; Rammensee, H.-G.; Decker, P. The mouse dendritic cell marker CD11c is down-regulated upon cell activation through Toll-like receptor triggering. Immunobiology 2013, 218, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, L.; Lu, Q.; Gao, Y.; Cai, Y.; Sui, A.; Su, T.; Shen, X.; Xie, B. Identification of different macrophage subpopulations with distinct activities in a mouse model of oxygen-induced retinopathy. Int. J. Mol. Med. 2017, 40, 281–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentala, H.; Verweij, W.R.; Der Vlag, A.H.-V.; van Loenen-Weemaes, A.M.; Meijer, D.K.F.; Poelstra, K. Removal of Phosphate from Lipid A as a Strategy to Detoxify Lipopolysaccharide. Shock 2002, 18, 561–566. [Google Scholar] [CrossRef]
- Bruhn, K.W.; Dekitani, K.; Nielsen, T.B.; Pantapalangkoor, P.; Spellberg, B. Ly6G-mediated depletion of neutrophils is dependent on macrophages. Results Immunol. 2016, 6, 5–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandon, J.M. Leucocyte distribution in the uterus during the preimplantation period of pregnancy and phagocyte recruitment to sites of blastocyst attachment in mice. J. Reprod. Fertil. 1993, 98, 567–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.Y.; Chi, J.-T.; Dudoit, S.; Bondre, C.; van de Rijn, M.; Botstein, D.; Brown, P.O. Diversity, topographic differentiation, and positional memory in human fibroblasts. Proc. Natl. Acad. Sci. USA 2002, 99, 12877–12882. [Google Scholar] [CrossRef] [Green Version]
- Filer, A.; Parsonage, G.; Smith, E.; Osborne, C.; Thomas, A.M.C.; Curnow, J.; Rainger, G.E.; Raza, K.; Nash, G.; Lord, J.; et al. Differential survival of leukocyte subsets mediated by synovial, bone marrow, and skin fibroblasts: Site-specific versus activation-dependent survival of T cells and neutrophils. Arthritis Rheum. 2006, 54, 2096–2108. [Google Scholar] [CrossRef]
- Paria, B.C.; Zhao, X.; Das, S.K.; Dey, S.K.; Yoshinaga, K. Zonula Occludens-1 and E-cadherin Are Coordinately Expressed in the Mouse Uterus with the Initiation of Implantation and Decidualization. Dev. Biol. 1999, 208, 488–501. [Google Scholar] [CrossRef] [Green Version]
- Giese, M.A.; Hind, L.E.; Huttenlocher, A. Neutrophil plasticity in the tumor microenvironment. Blood 2019, 133, 2159–2167. [Google Scholar] [CrossRef] [PubMed]
- Ingersoll, M.A.; Spanbroek, R.; Lottaz, C.; Gautier, E.; Frankenberger, M.; Hoffmann, R.; Lang, R.; Haniffa, M.; Collin, M.; Tacke, F.; et al. Comparison of gene expression profiles between human and mouse monocyte subsets. Blood 2010, 115, e10–e19. [Google Scholar] [CrossRef]
- Ramachandran, P.; Pellicoro, A.; Vernon, M.A.; Boulter, L.; Aucott, R.L.; Ali, A.; Hartland, S.N.; Snowdon, V.K.; Cappon, A.; Gordon-Walker, T.T.; et al. Differential Ly-6C expression identifies the recruited macrophage phenotype, which orchestrates the regression of murine liver fibrosis. Proc. Natl. Acad. Sci. USA 2012, 109, E3186–E3195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Italiani, P.; Boraschi, D. From Monocytes to M1/M2 Macrophages: Phenotypical vs. Functional Differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef] [Green Version]
- Laskarin, G.; Kämmerer, U.; Rukavina, D.; Thomson, A.W.; Fernandez, N.; Blois, S.M. Antigen-Presenting Cells and Materno-Fetal Tolerance: An Emerging Role for Dendritic Cells. Am. J. Reprod. Immunol. 2007, 58, 255–267. [Google Scholar] [CrossRef]
- Plaks, V.; Birnberg, T.; Berkutzki, T.; Sela, S.; BenYashar, A.; Kalchenko, V.; Mor, G.; Keshet, E.; Dekel, N.; Neeman, M.; et al. Uterine DCs are crucial for decidua formation during embryo implantation in mice. J. Clin. Investig. 2008, 118, 3954–3965. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Beers, D.R.; Thonhoff, J.R.; Thome, A.D.; Faridar, A.; Wang, J.; Wen, S.; Ornelas, L.; Sareen, D.; Goodridge, H.S.; et al. Immunosuppressive Functions of M2 Macrophages Derived from iPSCs of Patients with ALS and Healthy Controls. iScience 2020, 23, 101192. [Google Scholar] [CrossRef]
- Flanagan, K.; Modrusan, Z.; Cornelius, J.; Chavali, A.; Kasman, I.; Komuves, L.; Mo, L.; Diehl, L. Intestinal epithelial cell up-regulation of LY6 molecules during colitis results in enhanced chemokine secretion. J. Immunol. 2008, 180, 3874–3881. [Google Scholar] [CrossRef] [Green Version]
- Royer, D.J.; Elliott, M.H.; Le, Y.Z.; Carr, D.J. Corneal Epithelial Cells Exhibit Myeloid Characteristics and Present Antigen via MHC Class II. Investig. Ophthalmol. Vis. Sci. 2018, 59, 1512–1522. [Google Scholar] [CrossRef] [Green Version]
- Yue, L.; Daikoku, T.; Hou, X.; Li, M.; Wang, H.; Nojima, H.; Dey, S.K.; Das, S.K. Cyclin G1 and Cyclin G2 Are Expressed in the Periimplantation Mouse Uterus in a Cell-Specific and Progesterone-Dependent Manner: Evidence for Aberrant Regulation with Hoxa-10 Deficiency. Endocrinology 2005, 146, 2424–2433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Paria, B.C. Importance of uterine cell death, renewal, and their hormonal regulation in hamsters that show progesterone-dependent implantation. Endocrinology 2006, 147, 2215–2227. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Kim, J.; Yi, J.; Kim, D.; Kim, H.-O.; Han, D.; Sprent, J.; Lee, Y.J.; Surh, C.D.; Cho, J.-H. Phenotypic and Functional Changes of Peripheral Ly6C+ T Regulatory Cells Driven by Conventional Effector T Cells. Front. Immunol. 2018, 9, 437. [Google Scholar] [CrossRef] [PubMed]
- Alkhani, A.; Levy, C.S.; Tsui, M.; Rosenberg, K.; Polovina, K.; Mattis, A.N.; Mack, M.; Van Dyken, S.; Wang, B.M.; Maher, J.J.; et al. Ly6cLo non-classical monocytes promote resolution of rhesus rotavirus-mediated perinatal hepatic inflammation. Sci. Rep. 2020, 10, 7165. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.A.; Davies, M.L.; Reider, I.E.; Heipertz, E.L.; Epler, M.R.; Sei, J.J.; Ingersoll, M.A.; Van Rooijen, N.; Randolph, G.J.; Norbury, C.C. CD11b+, Ly6G+ Cells Produce Type I Interferon and Exhibit Tissue Protective Properties Following Peripheral Virus Infection. PLoS Pathog. 2011, 7, e1002374. [Google Scholar] [CrossRef] [Green Version]
- Dagvadorj, J.; Shimada, K.; Chen, S.; Jones, H.D.; Tumurkhuu, G.; Zhang, W.; Wawrowsky, K.A.; Crother, T.; Arditi, M. Lipopolysaccharide Induces Alveolar Macrophage Necrosis via CD14 and the P2X7 Receptor Leading to Interleukin-1alpha Release. Immunity 2015, 42, 640–653. [Google Scholar] [CrossRef] [Green Version]
- Tung, H.N.; Parr, M.B.; Parr, E.L. The permeability of the primary decidual zone in the rat uterus: An ultrastructural tracer and freeze-fracture study. Biol. Reprod. 1986, 35, 1045–1058. [Google Scholar] [CrossRef]
- Jose, S.; Madan, R. Neutrophil-mediated inflammation in the pathogenesis of Clostridium difficile infections. Anaerobe 2016, 41, 85–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadden, K.A.; Walsh, S.W. Neutrophils, but Not Lymphocytes or Monocytes, Infiltrate Maternal Systemic Vasculature in Women with Preeclampsia. Hypertens. Pregnancy 2008, 27, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Czuprynski, C.J.; Brown, J.F.; Maroushek, N.; Wagner, R.D.; Steinberg, H. Administration of anti-granulocyte mAb RB6-8C5 impairs the resistance of mice to Listeria monocytogenes infection. J. Immunol. 1994, 152, 1836–1846. [Google Scholar] [PubMed]
- Rinaldi, S.F.; Catalano, R.D.; Wade, J.; Rossi, A.G.; Norman, J. Decidual Neutrophil Infiltration Is Not Required for Preterm Birth in a Mouse Model of Infection-Induced Preterm Labor. J. Immunol. 2014, 192, 2315–2325. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhao, Y.; Zhong, X. Protective Effects of Baicalin on Decidua Cells of LPS-Induced Mice Abortion. J. Immunol. Res. 2014, 2014, 859812. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, A.R.; Zhong, S.; Dang, H.; Fordham, J.B.; Nares, S.; Khan, A. Expression Profiling of LPS Responsive miRNA in Primary Human Macrophages. J. Microb. Biochem. Technol. 2016, 8, 136–143. [Google Scholar] [CrossRef]
- Liu, W.; Niu, Z.; Li, Q.; Pang, R.T.; Chiu, C.N.; Yeung, W.S.-B. MicroRNA and Embryo Implantation. Am. J. Reprod. Immunol. 2015, 75, 263–271. [Google Scholar] [CrossRef]
- Brown, A.G.; Maubert, M.E.; Anton, L.; Heiser, L.M.; Elovitz, M. The tracking of lipopolysaccharide through the feto-maternal compartment and the involvement of maternal TLR4 in inflammation-induced fetal brain injury. Am. J. Reprod. Immunol. 2019, 82, e13189. [Google Scholar] [CrossRef] [Green Version]
- Kohmura, Y.; Kirikae, T.; Kirikae, F.; Nakano, M.; Sato, I. Lipopolysaccharide (LPS)-induced intra-uterine fetal death (IUFD) in mice is principally due to maternal cause but not fetal sensitivity to LPS. Microbiol. Immunol. 2000, 44, 897–904. [Google Scholar] [CrossRef] [Green Version]
- Hudalla, H.; Karenberg, K.; Kuon, R.-J.; Pöschl, J.; Tschada, R.; Frommhold, D. LPS-induced maternal inflammation promotes fetal leukocyte recruitment and prenatal organ infiltration in mice. Pediatr. Res. 2018, 84, 757–764. [Google Scholar] [CrossRef] [PubMed]
- de Vos, A.F.; Pater, J.M.; van den Pangaart, P.S.; de Kruif, M.D.; van’t Veer, C.; van der Poll, T. In vivo lipopolysaccharide exposure of human blood leukocytes induces cross-tolerance to multiple TLR ligands. J. Immunol. 2009, 183, 533–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, B. Biological basis of the behavior of sick animals. Neurosci. Biobehav. Rev. 1988, 12, 123–137. [Google Scholar] [CrossRef]
- Vitoratos, N.; Papadias, C.; Economou, E.; Makrakis, E.; Panoulis, C.; Creatsas, G. Elevated Circulating IL-1beta and TNF-Alpha, and Unaltered IL-6 in First-Trimester Pregnancies Complicated by Threatened Abortion With an Adverse Outcome. Mediat. Inflamm. 2006, 2006, 030485. [Google Scholar] [CrossRef]
- Romero, R.; Gotsch, F.; Pineles, B.; Kusanovic, J.P. Inflammation in Pregnancy: Its Roles in Reproductive Physiology, Obstetrical Complications, and Fetal Injury. Nutr. Rev. 2007, 65, 194–202. [Google Scholar] [CrossRef]
- Guzeloglu-Kayisli, O.; Kayisli, U.A.; Taylor, H.S. The Role of Growth Factors and Cytokines during Implantation: Endocrine and Paracrine Interactions. Semin. Reprod. Med. 2009, 27, 062–079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar]
- Benjamin, J.T.; Plosa, E.J.; Sucre, J.M.; Van Der Meer, R.; Dave, S.; Gutor, S.S.; Nichols, D.S.; Gulleman, P.M.; Jetter, C.S.; Han, W.; et al. Neutrophilic inflammation during lung development disrupts elastin assembly and predisposes adult mice to COPD. J. Clin. Investig. 2021, 131, e139481. [Google Scholar] [CrossRef]
- Coffelt, S.B.; Kersten, K.; Doornebal, C.W.; Weiden, J.; Vrijland, K.; Hau, C.S.; Verstegen, N.J.M.; Ciampricotti, M.; Hawinkels, L.J.A.C.; Jonkers, J.; et al. IL-17-producing gammadelta T cells and neutrophils conspire to promote breast cancer metastasis. Nature 2015, 522, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Finisguerra, V.; Di Conza, G.; Di Matteo, M.; Serneels, J.; Costa, S.; Thompson, R.; Wauters, E.; Walmsley, S.; Prenen, H.; Granot, Z.; et al. MET is required for the recruitment of anti-tumoural neutrophils. Nature 2015, 522, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Friggeri, A.; Yang, Y.; Park, Y.-J.; Tsuruta, Y.; Abraham, E. miR-147, a microRNA that is induced upon Toll-like receptor stimulation, regulates murine macrophage inflammatory responses. Proc. Natl. Acad. Sci. USA 2009, 106, 15819–15824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, W.; Nguyen, H.; Brown, N.; Ni, H.; Kiffer-Moreira, T.; Reese, J.; Millán, J.L.; Paria, B.C. Alkaline phosphatases contribute to uterine receptivity, implantation, decidualization, and defense against bacterial endotoxin in hamsters. Reproduction 2013, 146, 419–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Treatment | Mice (#) | EBIS/Mouse (#) | Wet Weight/EBIS (mg) |
|---|---|---|---|---|
| 1 | Saline (vehicle) | 11 | 6.7 | 14.73 ± 0.18 |
| 2 | Anti-Ly6G Ab | 5 | 6.4 | 14.64 ± 0.30 |
| 3 | LPS | 13 | 6.6 | 3.43 ± 0.24 *** |
| 4 | Anti-IgG2b + LPS | 5 | 6.2 | 3.69 ± 0.28 *** |
| 5 | Anti-Ly6G Ab + LPS | 5 | 6.2 | 3.92 ± 0.29 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panja, S.; Benjamin, J.T.; Paria, B.C. Maternal Neutrophil Depletion Fails to Avert Systemic Lipopolysaccharide-Induced Early Pregnancy Defects in Mice. Int. J. Mol. Sci. 2021, 22, 7932. https://doi.org/10.3390/ijms22157932
Panja S, Benjamin JT, Paria BC. Maternal Neutrophil Depletion Fails to Avert Systemic Lipopolysaccharide-Induced Early Pregnancy Defects in Mice. International Journal of Molecular Sciences. 2021; 22(15):7932. https://doi.org/10.3390/ijms22157932
Chicago/Turabian StylePanja, Sourav, John T. Benjamin, and Bibhash C. Paria. 2021. "Maternal Neutrophil Depletion Fails to Avert Systemic Lipopolysaccharide-Induced Early Pregnancy Defects in Mice" International Journal of Molecular Sciences 22, no. 15: 7932. https://doi.org/10.3390/ijms22157932