
Quaternary Structure and Hetero-Oligomerization of Recombinant Human Small Heat Shock Protein HspB7 (cvHsp)


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
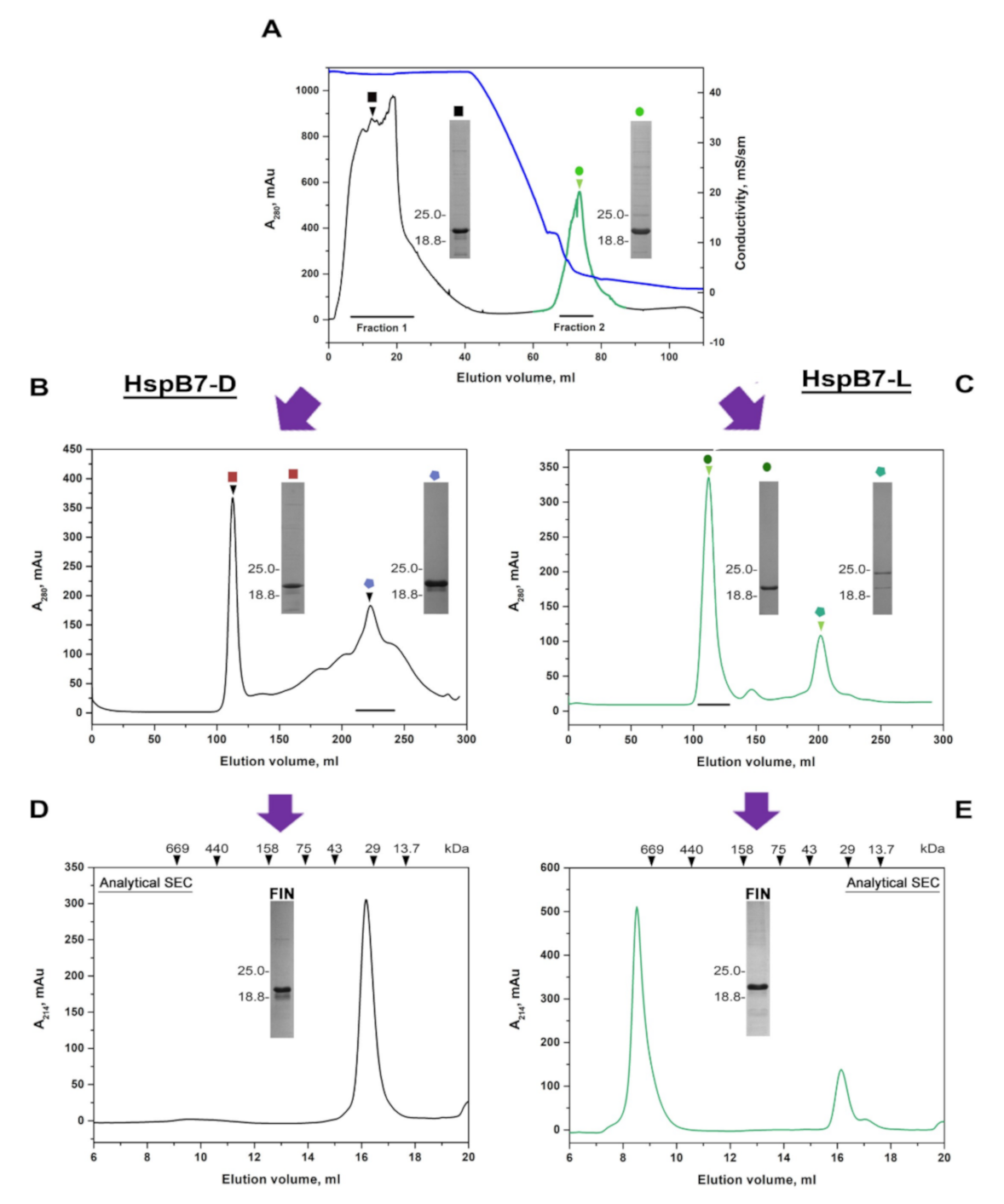
2.1. Isolation of Untagged Recombinant HspB7
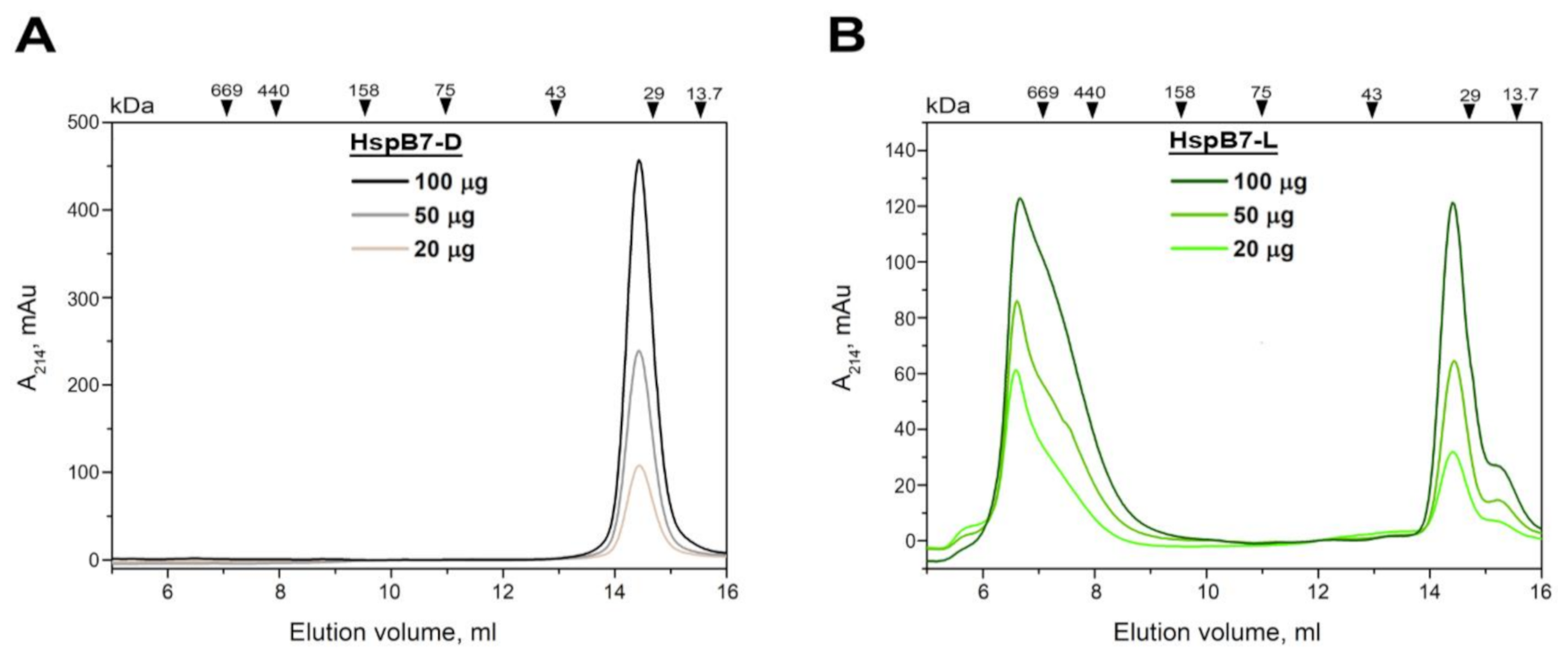
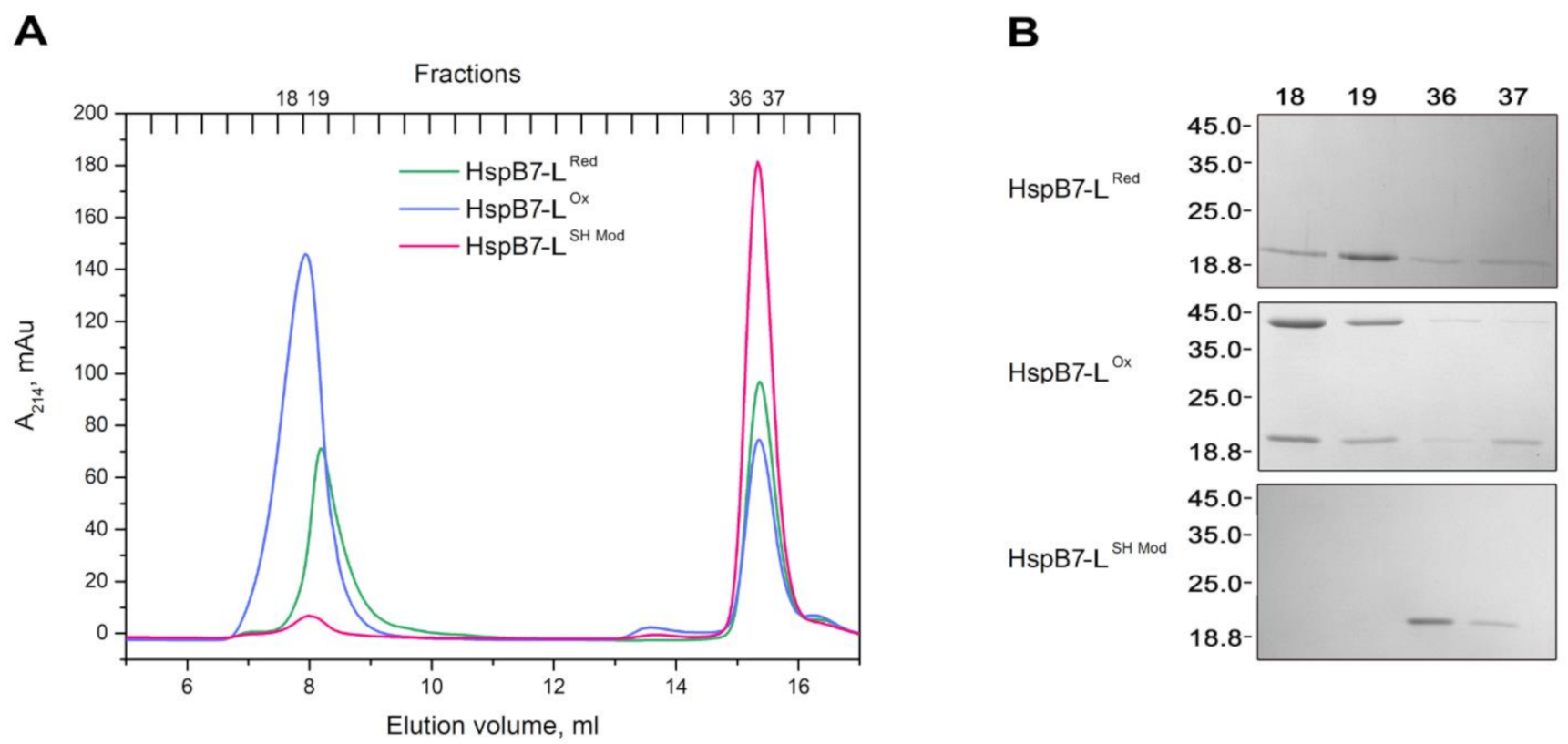
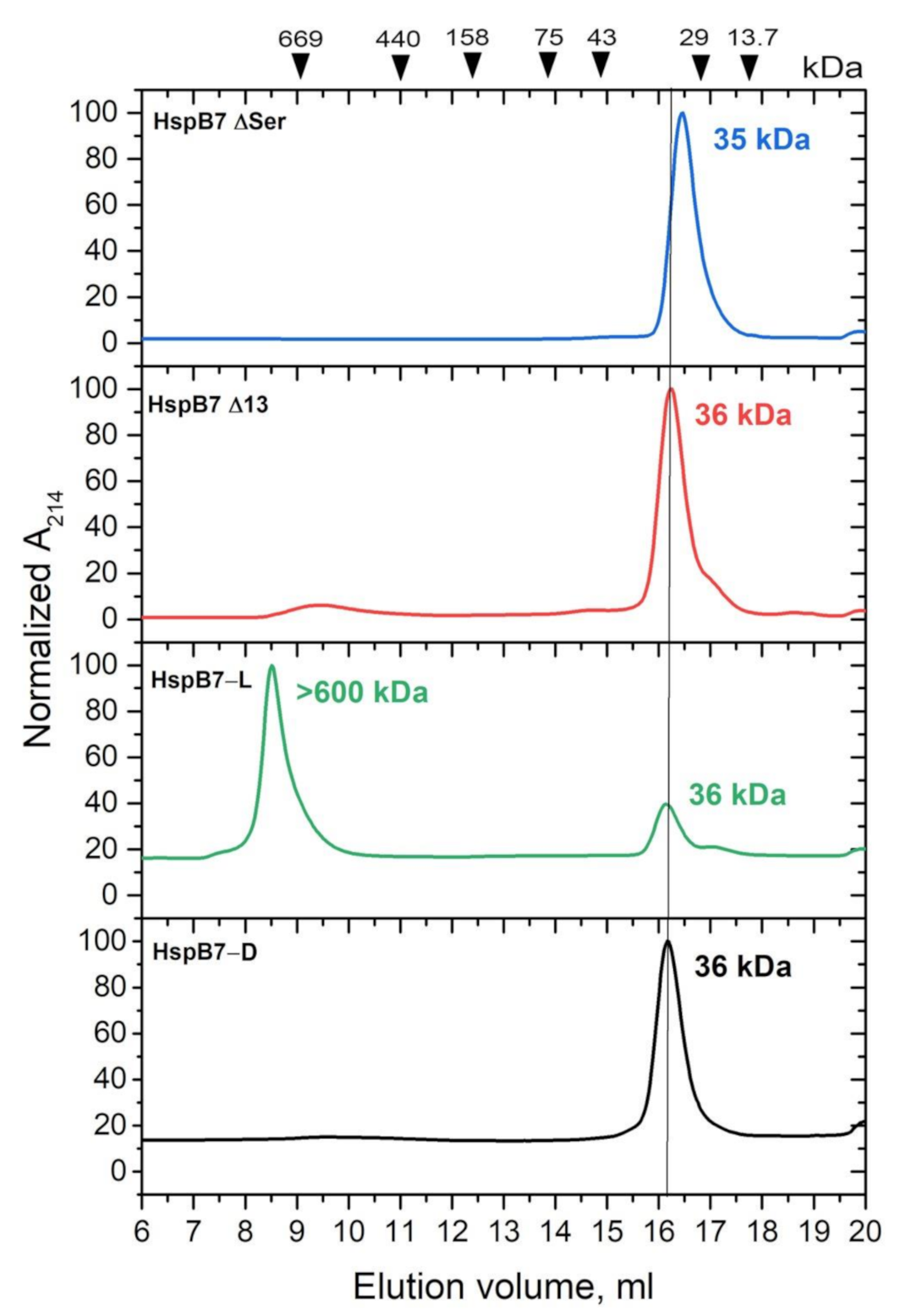
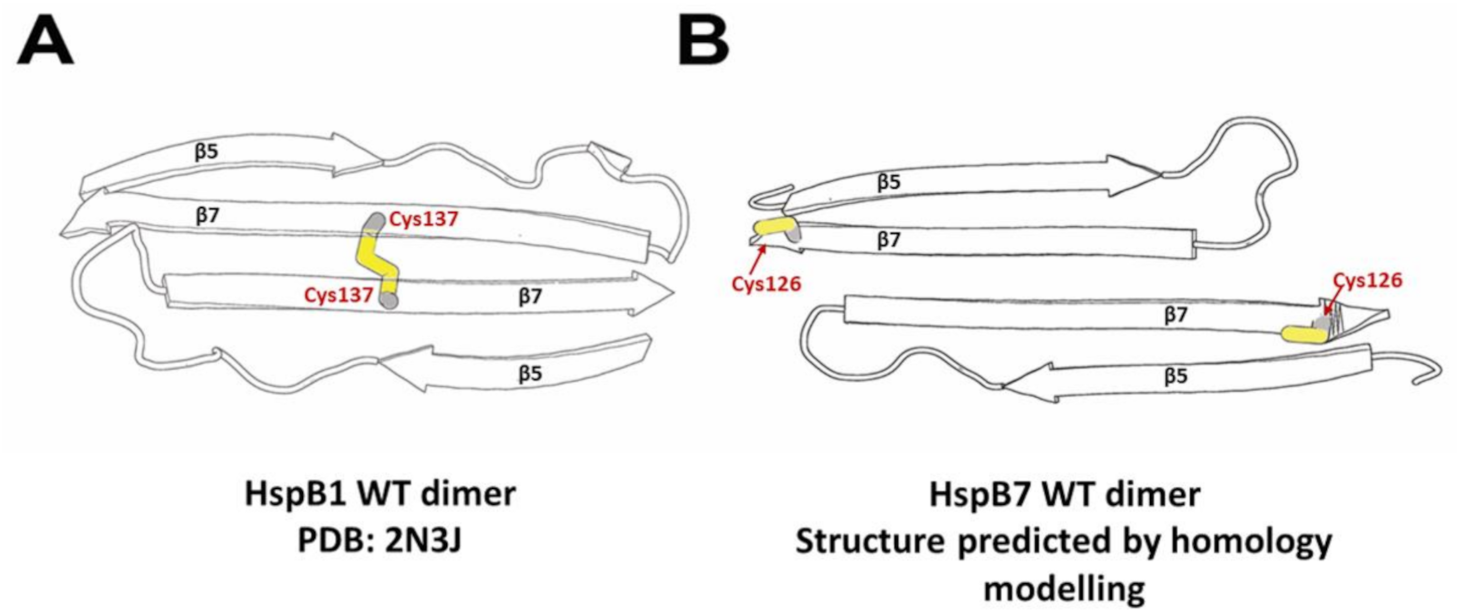
2.2. Oligomeric State of HspB7 and Its Dependence on the N-Terminal Domain and Oxidation
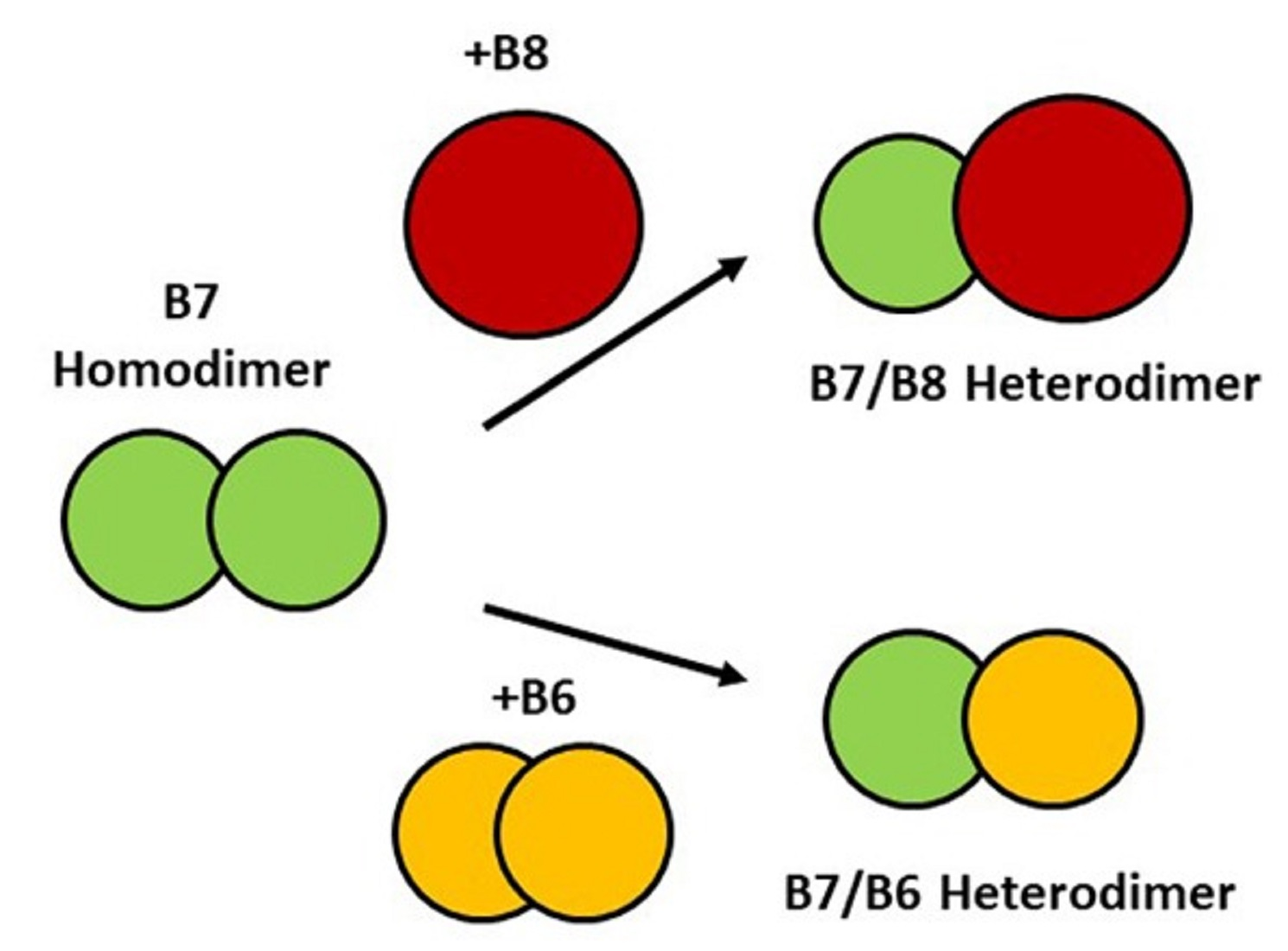
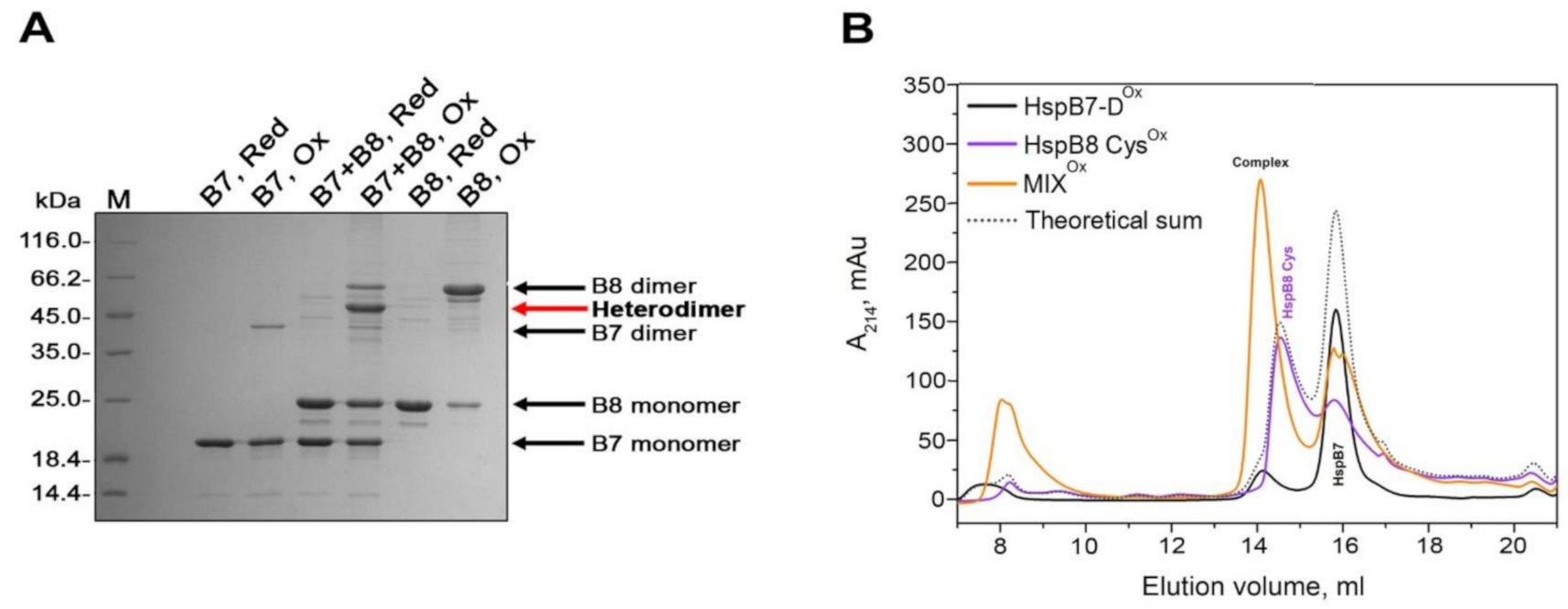
2.3. Interaction of HspB7 with HspB6 and HspB8
3. Discussion
4. Materials and Methods
4.1. Cloning of the Wild-Type Proteins and Their Mutants
4.2. Expression and Purification of Recombinant Proteins
4.3. Size-Exclusion Chromatography
4.4. Modification of SH-Group of HspB7
4.5. Interaction of HspB7 with Hydrophobic Probe Bis-ANS
4.6. Crosslinking of HspB7 and Cys-Mutants of HspB6 and HspB8
4.7. ELISA Experiments
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACD | α-crystallin domain |
| CTD | C-terminal domain |
| DTT | dithiotreitol |
| IPTG | isopropyl β-D-1-thiogalactopyranoside |
| ME | β-mercaptoethanol |
| NTD | N-terminal domain |
| PMSF | phenylmethanesulfonyl fluoride |
| SDS | sodium dodecyl sulfate |
| SEC | size-exclusion chromatography |
| sHsp | small heat shock proteins |
References
- Kriehuber, T.; Rattei, T.; Weinmaier, T.; Bepperling, A.; Haslbeck, M.; Buchner, J. Independent evolution of the core domain and its flanking sequences in small heat shock proteins. FASEB J. 2010, 24, 3633–3642. [Google Scholar] [CrossRef]
- Maaroufi, H.; Tanguay, R.M. Analysis and phylogeny of small heat shock proteins from marine viruses and their cyanobacteria host. PLoS ONE 2013, 8, e81207. [Google Scholar] [CrossRef] [PubMed]
- Mogk, A.; Ruger-Herreros, C.; Bukau, B. Cellular functions and mechanisms of action of small heat shock proteins. Annu. Rev. Microbiol. 2019, 73, 89–110. [Google Scholar] [CrossRef]
- Waters, E.R.; Vierling, E. Plant small heat shock proteins—Evolutionary and functional diversity. New Phytol. 2020, 227, 24–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haslbeck, M.; Vierling, E. A first line of stress defense: Small heat shock proteins and their function in protein homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar] [CrossRef] [Green Version]
- Kappe, G.; Boelens, W.C.; de Jong, W.W. Why proteins without an alpha-crystallin domain should not be included in the human small heat shock protein family hspb. Cell Stress Chaperones 2010, 15, 457–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mymrikov, E.V.; Seit-Nebi, A.S.; Gusev, N.B. Large potentials of small heat shock proteins. Physiol. Rev. 2011, 91, 1123–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janowska, M.K.; Baughman, H.E.R.; Woods, C.N.; Klevit, R.E. Mechanisms of small heat shock proteins. Cold Spring Harb. Perspect. Biol. 2019, 11, a034025. [Google Scholar] [CrossRef]
- Bagneris, C.; Bateman, O.A.; Naylor, C.E.; Cronin, N.; Boelens, W.C.; Keep, N.H.; Slingsby, C. Crystal structures of alpha-crystallin domain dimers of alphab-crystallin and hsp20. J. Mol. Biol. 2009, 392, 1242–1252. [Google Scholar] [CrossRef] [PubMed]
- Delbecq, S.P.; Jehle, S.; Klevit, R. Binding determinants of the small heat shock protein, alphab-crystallin: Recognition of the ‘ixi’ motif. EMBO J. 2012, 31, 4587–4594. [Google Scholar] [CrossRef] [Green Version]
- Hilton, G.R.; Hochberg, G.K.; Laganowsky, A.; McGinnigle, S.I.; Baldwin, A.J.; Benesch, J.L. C-terminal interactions mediate the quaternary dynamics of alphab-crystallin. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2013, 368, 20110405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delbecq, S.P.; Rosenbaum, J.C.; Klevit, R.E. A mechanism of subunit recruitment in human small heat shock protein oligomers. Biochemistry 2015, 54, 4276–4284. [Google Scholar] [CrossRef] [Green Version]
- Kappe, G.; Franck, E.; Verschuure, P.; Boelens, W.C.; Leunissen, J.A.; de Jong, W.W. The human genome encodes 10 alpha-crystallin-related small heat shock proteins: Hspb1-10. Cell Stress Chaperones 2003, 8, 53–61. [Google Scholar] [CrossRef]
- Fontaine, J.M.; Rest, J.S.; Welsh, M.J.; Benndorf, R. The sperm outer dense fiber protein is the 10th member of the superfamily of mammalian small stress proteins. Cell Stress Chaperones 2003, 8, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Vos, M.J.; Kanon, B.; Kampinga, H.H. Hspb7 is a sc35 speckle resident small heat shock protein. Biochim. Biophys. Acta 2009, 1793, 1343–1353. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.P.; Benjamin, I.J. Small heat shock proteins: A new classification scheme in mammals. J. Mol. Cell. Cardiol. 2005, 38, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Mainz, A.; Peschek, J.; Stavropoulou, M.; Back, K.C.; Bardiaux, B.; Asami, S.; Prade, E.; Peters, C.; Weinkauf, S.; Buchner, J.; et al. The chaperone alphab-crystallin uses different interfaces to capture an amorphous and an amyloid client. Nat. Struct. Mol. Biol. 2015, 22, 898–905. [Google Scholar] [CrossRef]
- Jovcevski, B.; Kelly, M.A.; Rote, A.P.; Berg, T.; Gastall, H.Y.; Benesch, J.L.; Aquilina, J.A.; Ecroyd, H. Phosphomimics destabilize hsp27 oligomeric assemblies and enhance chaperone activity. Chem. Biol. 2015, 22, 186–195. [Google Scholar] [CrossRef]
- Shemetov, A.A.; Seit-Nebi, A.S.; Gusev, N.B. Structure, properties, and functions of the human small heat-shock protein hsp22 (hspb8, h11, e2ig1): A critical review. J. Neurosci. Res. 2008, 86, 264–269. [Google Scholar] [CrossRef]
- Cristofani, R.; Piccolella, M.; Crippa, V.; Tedesco, B.; Montagnani Marelli, M.; Poletti, A.; Moretti, R.M. The role of hspb8, a component of the chaperone-assisted selective autophagy machinery, in cancer. Cells 2021, 10, 335. [Google Scholar] [CrossRef]
- Krief, S.; Faivre, J.F.; Robert, P.; Le Douarin, B.; Brument-Larignon, N.; Lefrere, I.; Bouzyk, M.M.; Anderson, K.M.; Greller, L.D.; Tobin, F.L.; et al. Identification and characterization of cvhsp. A novel human small stress protein selectively expressed in cardiovascular and insulin-sensitive tissues. J. Biol. Chem. 1999, 274, 36592–36600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vos, M.J.; Zijlstra, M.P.; Kanon, B.; van Waarde-Verhagen, M.A.; Brunt, E.R.; Oosterveld-Hut, H.M.; Carra, S.; Sibon, O.C.; Kampinga, H.H. Hspb7 is the most potent polyq aggregation suppressor within the hspb family of molecular chaperones. Hum. Mol. Genet. 2010, 19, 4677–4693. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Vonk, J.J.; Salles, F.; Vonk, D.; Haslbeck, M.; Melki, R.; Bergink, S.; Kampinga, H.H. The n terminus of the small heat shock protein hspb7 drives its polyq aggregation-suppressing activity. J. Biol. Chem. 2019, 294, 9985–9994. [Google Scholar] [CrossRef] [Green Version]
- Surya, S.L.; Long, M.J.C.; Urul, D.A.; Zhao, Y.; Mercer, E.J.; IM, E.I.; Evans, T.; Aye, Y. Cardiovascular small heat shock protein hspb7 is a kinetically privileged reactive electrophilic species (res) sensor. ACS Chem. Biol. 2018, 13, 1824–1831. [Google Scholar] [CrossRef]
- Mercer, E.J.; Lin, Y.F.; Cohen-Gould, L.; Evans, T. Hspb7 is a cardioprotective chaperone facilitating sarcomeric proteostasis. Dev. Biol. 2018, 435, 41–55. [Google Scholar] [CrossRef]
- Juo, L.Y.; Liao, W.C.; Shih, Y.L.; Yang, B.Y.; Liu, A.B.; Yan, Y.T. Hspb7 interacts with dimerized flnc and its absence results in progressive myopathy in skeletal muscles. J. Cell Sci. 2016, 129, 1661–1670. [Google Scholar] [CrossRef] [Green Version]
- Asthana, A.; Raman, B.; Ramakrishna, T.; Rao Ch, M. Structural aspects and chaperone activity of human hspb3: Role of the “c-terminal extension”. Cell Biochem. Biophys. 2012, 64, 61–72. [Google Scholar] [CrossRef]
- Mymrikov, E.V.; Daake, M.; Richter, B.; Haslbeck, M.; Buchner, J. The chaperone activity and substrate spectrum of human small heat shock proteins. J. Biol. Chem. 2017, 292, 672–684. [Google Scholar] [CrossRef] [Green Version]
- Mymrikov, E.V.; Bukach, O.V.; Seit-Nebi, A.S.; Gusev, N.B. The pivotal role of the beta 7 strand in the intersubunit contacts of different human small heat shock proteins. Cell Stress Chaperones 2010, 15, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mymrikov, E.V.; Seit-Nebi, A.S.; Gusev, N.B. Heterooligomeric complexes of human small heat shock proteins. Cell Stress Chaperones 2011, 17, 157–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrigo, A.P. Human small heat shock proteins: Protein interactomes of homo- and hetero-oligomeric complexes: An update. FEBS Lett. 2013, 587, 1959–1969. [Google Scholar] [CrossRef] [Green Version]
- Alderson, T.R.; Roche, J.; Gastall, H.Y.; Dias, D.M.; Pritisanac, I.; Ying, J.; Bax, A.; Benesch, J.L.P.; Baldwin, A.J. Local unfolding of the hsp27 monomer regulates chaperone activity. Nat. Commun. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochberg, G.K.; Ecroyd, H.; Liu, C.; Cox, D.; Cascio, D.; Sawaya, M.R.; Collier, M.P.; Stroud, J.; Carver, J.A.; Baldwin, A.J.; et al. The structured core domain of alphab-crystallin can prevent amyloid fibrillation and associated toxicity. Proc. Natl. Acad. Sci. USA 2014, 111, E1562–E1570. [Google Scholar] [CrossRef] [Green Version]
- Fan, G.C.; Kranias, E.G. Small heat shock protein 20 (hspb6) in cardiac hypertrophy and failure. J. Mol. Cell. Cardiol. 2011, 51, 574–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labbe, J.P.; Boyer, M.; Mejean, C.; Roustan, C.; Benyamin, Y. Localization of two myosin-subfragment-1 binding contacts in the 96-132 region of actin subdomain-1. Eur. J. Biochem. 1993, 215, 17–24. [Google Scholar] [CrossRef]
- Mejean, C.; Lebart, M.C.; Boyer, M.; Roustan, C.; Benyamin, Y. Localization and identification of actin structures involved in the filamin-actin interaction. Eur. J. Biochem. 1992, 209, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, Y.; Lu, Y.; Zhao, X. Molecular characterization of rat cvhsp/hspb7 in vitro and its dynamic molecular architecture. Mol. Med. Rep. 2011, 4, 105–111. [Google Scholar]
- Wu, T.; Mu, Y.; Bogomolovas, J.; Fang, X.; Veevers, J.; Nowak, R.B.; Pappas, C.T.; Gregorio, C.C.; Evans, S.M.; Fowler, V.M.; et al. Hspb7 is indispensable for heart development by modulating actin filament assembly. Proc. Natl. Acad. Sci. USA 2017, 114, 11956–11961. [Google Scholar] [CrossRef] [Green Version]
- Laganowsky, A.; Benesch, J.L.; Landau, M.; Ding, L.; Sawaya, M.R.; Cascio, D.; Huang, Q.; Robinson, C.V.; Horwitz, J.; Eisenberg, D. Crystal structures of truncated alphaa and alphab crystallins reveal structural mechanisms of polydispersity important for eye lens function. Protein Sci. 2010, 19, 1031–1043. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, C.J.O.; Peters, C.; Schmid, P.W.N.; Stavropoulou, M.; Zou, J.; Dahiya, V.; Mymrikov, E.V.; Rockel, B.; Asami, S.; Haslbeck, M.; et al. The structure and oxidation of the eye lens chaperone alphaa-crystallin. Nat. Struct. Mol. Biol. 2019, 26, 1141–1150. [Google Scholar] [CrossRef]
- Clouser, A.F.; Baughman, H.E.; Basanta, B.; Guttman, M.; Nath, A.; Klevit, R.E. Interplay of disordered and ordered regions of a human small heat shock protein yields an ensemble of ‘quasi-ordered’ states. eLife 2019, 8, e50259. [Google Scholar] [CrossRef]
- Bukach, O.V.; Glukhova, A.E.; Seit-Nebi, A.S.; Gusev, N.B. Heterooligomeric complexes formed by human small heat shock proteins hspb1 (hsp27) and hspb6 (hsp20). Biochim. Biophys. Acta 2009, 1794, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Fontaine, J.M.; Rest, J.S.; Shelden, E.A.; Welsh, M.J.; Benndorf, R. Interaction of human hsp22 (hspb8) with other small heat shock proteins. J. Biol. Chem. 2004, 279, 2394–2402. [Google Scholar] [CrossRef] [Green Version]
- Golenhofen, N.; Perng, M.D.; Quinlan, R.A.; Drenckhahn, D. Comparison of the small heat shock proteins alphab-crystallin, mkbp, hsp25, hsp20, and cvhsp in heart and skeletal muscle. Histochem. Cell Biol. 2004, 122, 415–425. [Google Scholar] [CrossRef]
- Bukach, O.V.; Seit-Nebi, A.S.; Marston, S.B.; Gusev, N.B. Some properties of human small heat shock protein hsp20 (hspb6). Eur. J. Biochem. 2004, 271, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Kasakov, A.S.; Bukach, O.V.; Seit-Nebi, A.S.; Marston, S.B.; Gusev, N.B. Effect of mutations in the beta5-beta7 loop on the structure and properties of human small heat shock protein hsp22 (hspb8, h11). FEBS J. 2007, 274, 5628–5642. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage t4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Reisfeld, R.A.; Lewis, U.J.; Williams, D.E. Disk electrophoresis of basic proteins and peptides on polyacrylamide gels. Nature 1962, 195, 281–283. [Google Scholar] [CrossRef]
- Davis, B.J. Disc electrophoresis. Ii. Method and application to human serum proteins. Ann. N. Y. Acad. Sci. 1964, 121, 404–427. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muranova, L.K.; Shatov, V.M.; Slushchev, A.V.; Gusev, N.B. Quaternary Structure and Hetero-Oligomerization of Recombinant Human Small Heat Shock Protein HspB7 (cvHsp). Int. J. Mol. Sci. 2021, 22, 7777. https://doi.org/10.3390/ijms22157777
Muranova LK, Shatov VM, Slushchev AV, Gusev NB. Quaternary Structure and Hetero-Oligomerization of Recombinant Human Small Heat Shock Protein HspB7 (cvHsp). International Journal of Molecular Sciences. 2021; 22(15):7777. https://doi.org/10.3390/ijms22157777
Chicago/Turabian StyleMuranova, Lydia K., Vladislav M. Shatov, Andrey V. Slushchev, and Nikolai B. Gusev. 2021. "Quaternary Structure and Hetero-Oligomerization of Recombinant Human Small Heat Shock Protein HspB7 (cvHsp)" International Journal of Molecular Sciences 22, no. 15: 7777. https://doi.org/10.3390/ijms22157777