
Novel Hydrogen Sulfide (H2S)-Releasing BW-HS-101 and Its Non-H2S Releasing Derivative in Modulation of Microscopic and Molecular Parameters of Gastric Mucosal Barrier

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
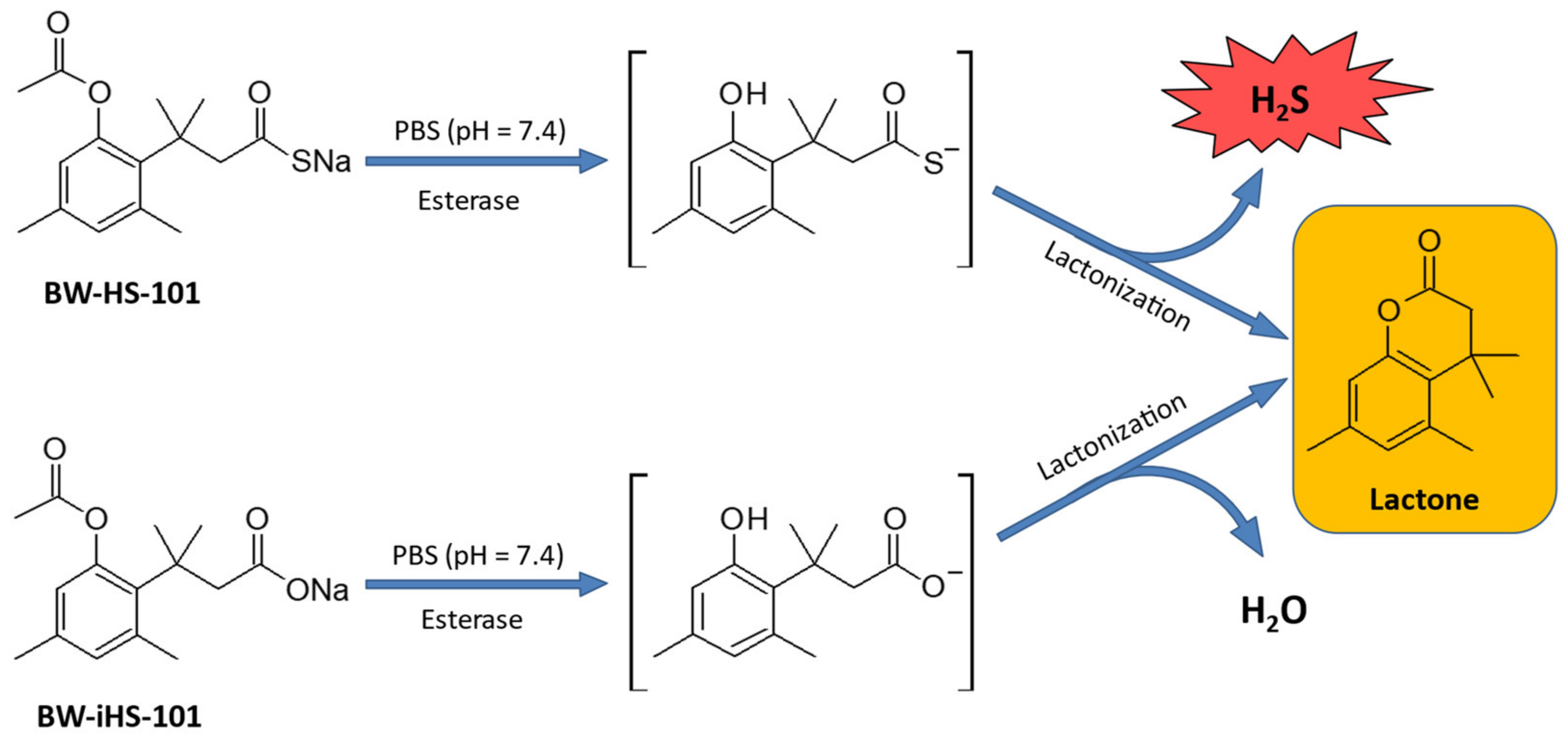
2.1. Chemical Conversion of BW-HS-101 and BW-iHS-101
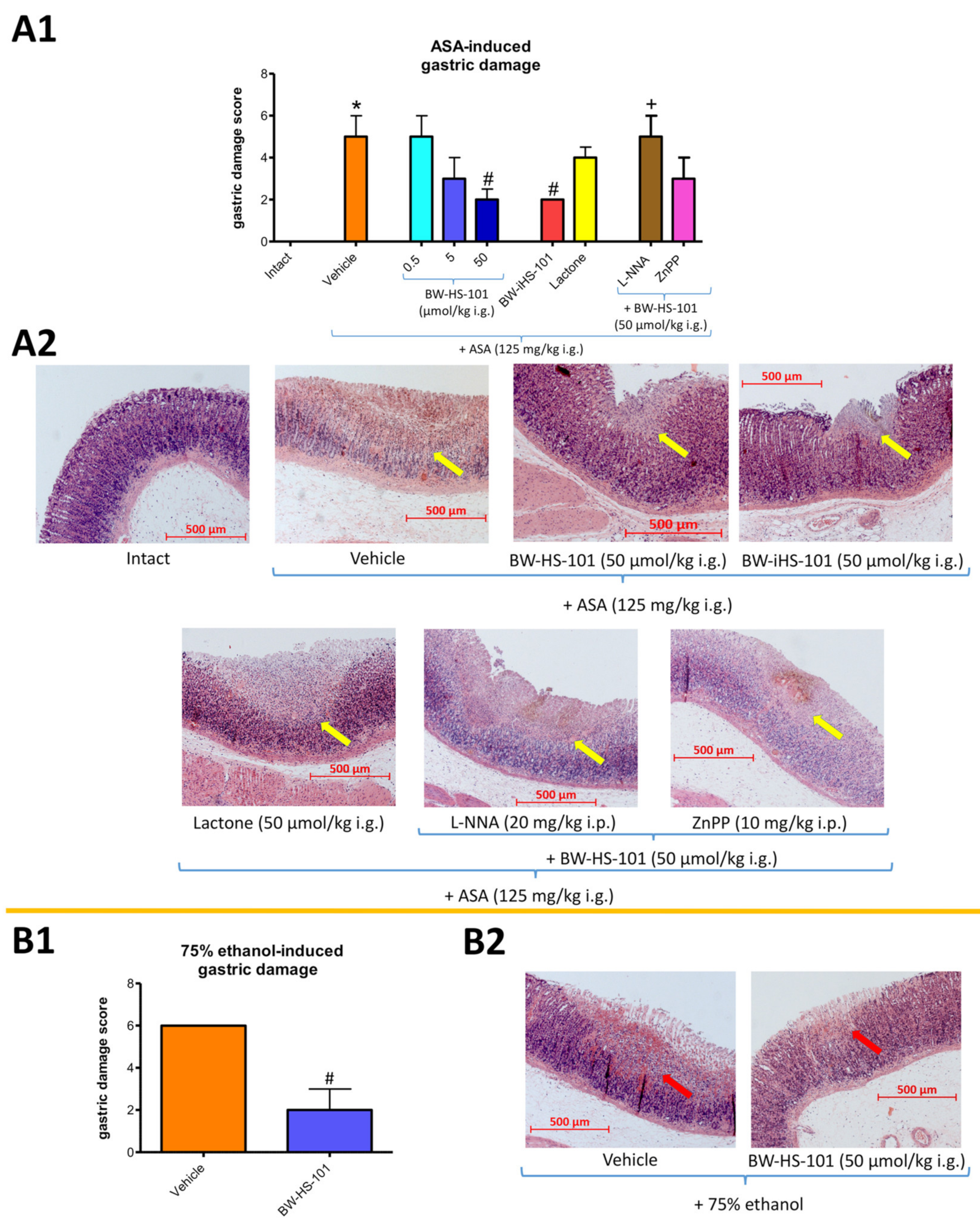
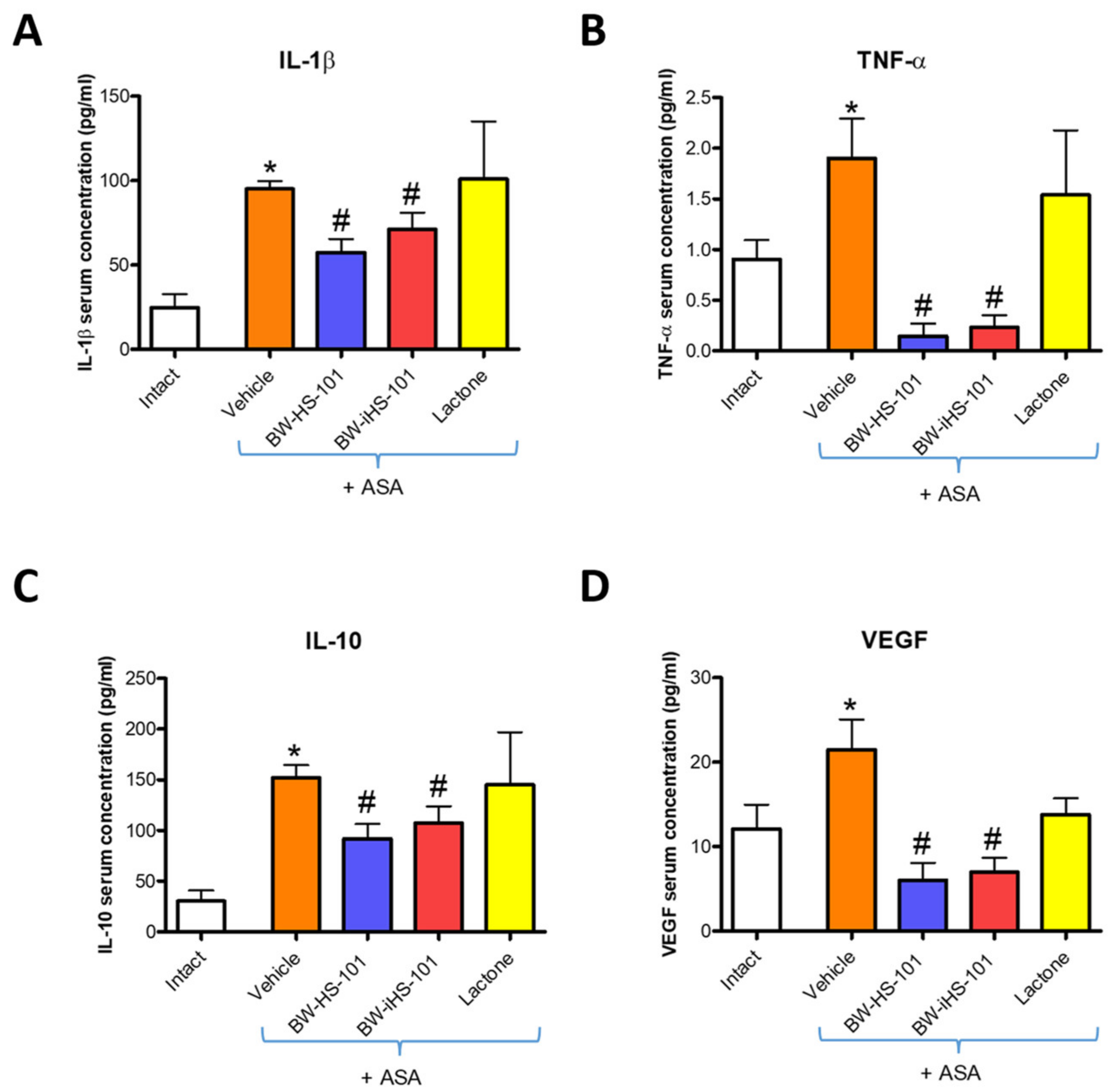
2.2. Possible Effects on the Gastric Damage Score, Gastric Blood Flow Alterations, and Systemic and Gastric Mucosal Inflammation. Pharmacological Evaluation of the Involvement of Endogenous NO and CO Biosynthesis Pathways
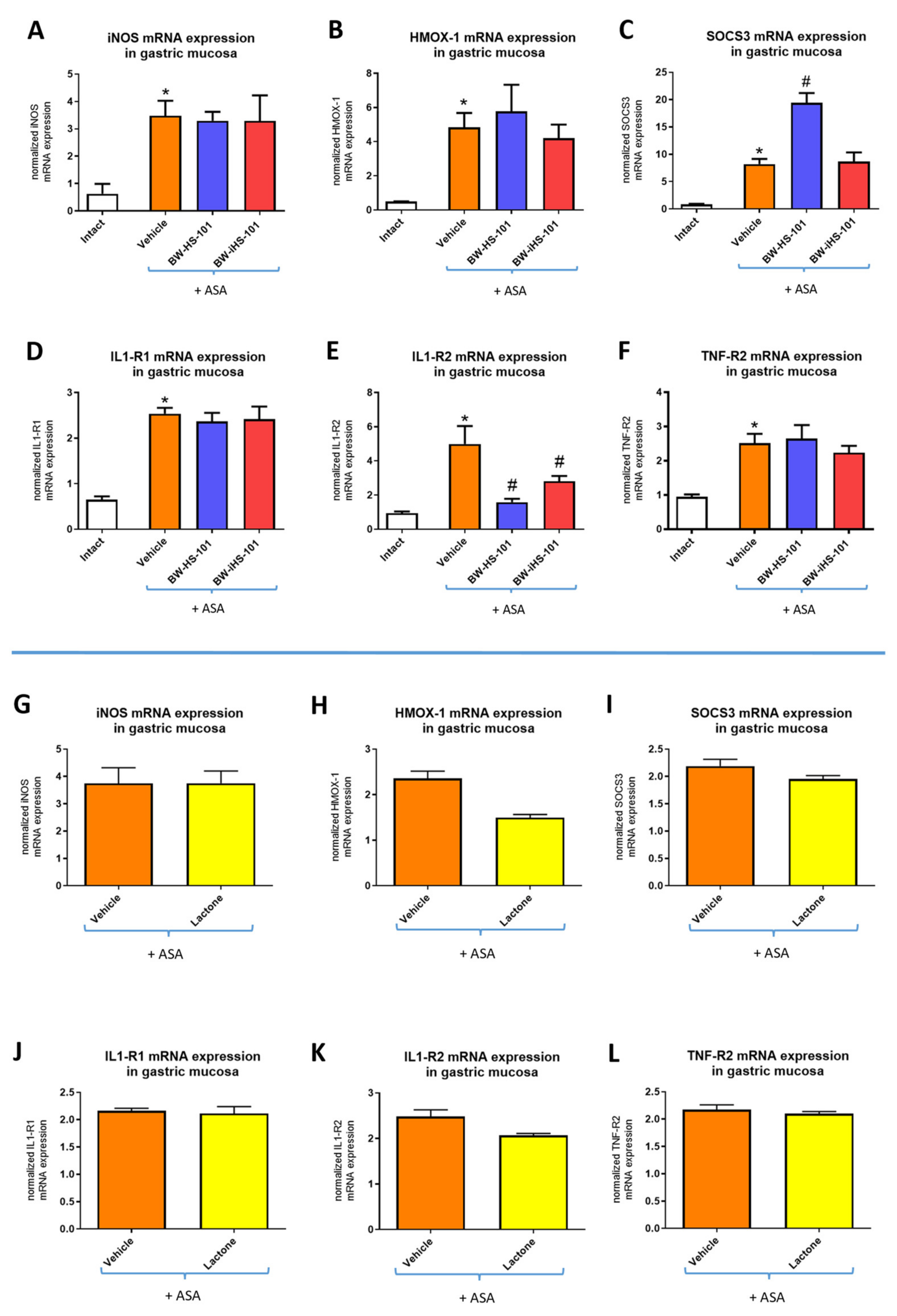
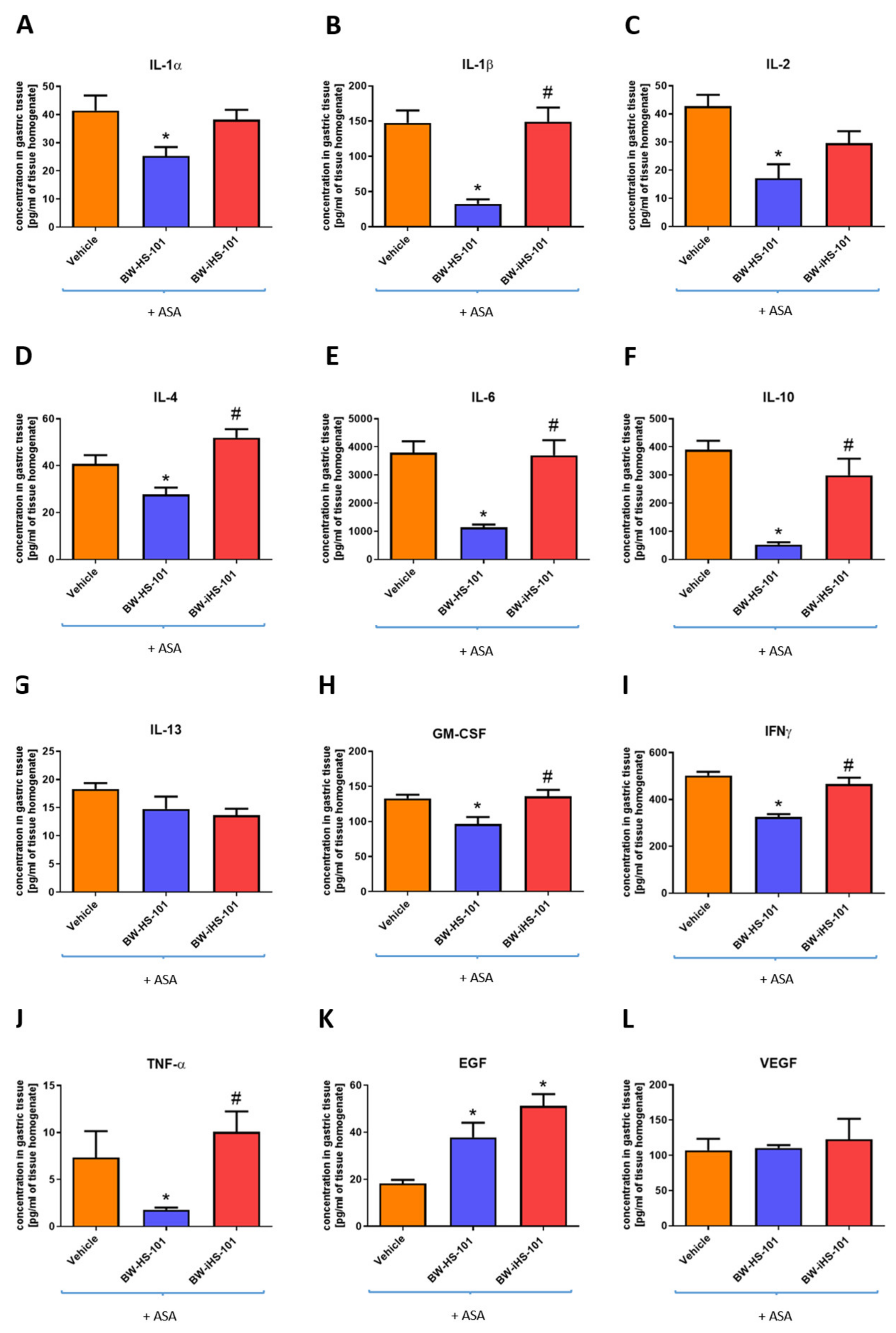
2.3. Molecular Pattern of Gastric Mucosal Proteins
2.4. Possible Alterations in the Gastric Mucosal Prostaglandins/Cyclooxygenase Pathway Activity
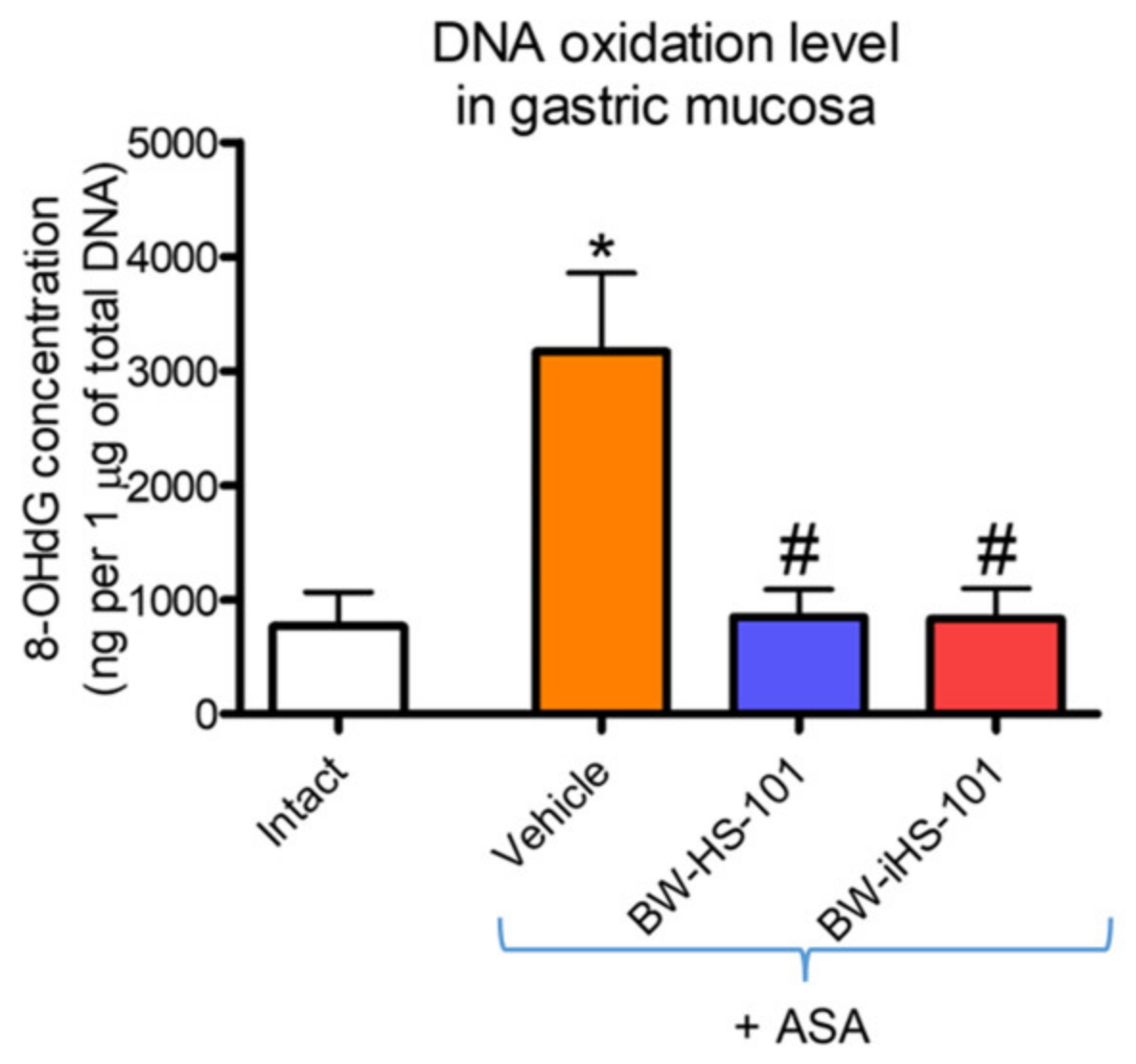
2.5. Oxidation of DNA in Gastric Mucosa
2.6. Bioinformatic Evaluation of Possible Molecular Targets
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Experimental Design
5.2. BW-HS-101 and BW-iHS-101 Synthesis and Chemical Conversion to Lactone
5.3. GBF Determination, Microscopic Gastric Damage Assessment and Biological Samples Collection
- 0
- no erosion/necrosis/inflammation
- 1
- length of injury <250 μm
- 2
- length of injury 251–500 μm
- 3
- length of injury 501–2000 μm
- 0
- no erosion/necrosis/inflammation
- 1
- depth of injury <500 μm per tissue section
- 2
- depth of injury >500 μm per slide
- 3
- depth of injury—erosion reaching submucosal layer
5.4. Determination of Gastric Mucosal mRNA Fold Changes by Real-Time PCR
5.5. Luminex Microbeads Fluorescent Assays
5.6. Determination of PGE2 and 8-OHdG Concentration in Gastric Mucosa
5.7. Bioinformatic Analysis of Possible Molecular Targets of BW-HS-101
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fang, J.; George, M.G.; Gindi, R.M.; Hong, Y.; Yang, Q.; Ayala, C.; Ward, B.W.; Loustalot, F. Use of low-dose aspirin as secondary prevention of atherosclerotic cardiovascular disease in US adults (from the National Health Interview Survey, 2012). Am. J. Cardiol. 2015, 115, 895–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sostres, C.; Lanas, A. Gastrointestinal effects of aspirin. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Laine, L. Nonsteroidal anti-inflammatory drug gastropathy. Gastrointest. Endosc. Clin. N. Am. 1996, 6, 489–504. [Google Scholar] [CrossRef]
- Matsui, H.; Shimokawa, O.; Kaneko, T.; Nagano, Y.; Rai, K.; Hyodo, I. The pathophysiology of non-steroidal anti-inflammatory drug (NSAID)-induced mucosal injuries in stomach and small intestine. J. Clin. Biochem. Nutr. 2011, 48, 107–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryer, B.; Mahaffey, K.W. Gastrointestinal ulcers, role of aspirin, and clinical outcomes: Pathobiology, diagnosis, and treatment. J. Multidiscip. Health 2014, 7, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.L.; Ferraz, J.G.; Muscara, M.N. Hydrogen Sulfide: An Endogenous Mediator of Resolution of Inflammation and Injury. Antioxid. Redox Signal. 2012, 17, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Kajimura, M.; Fukuda, R.; Bateman, R.M.; Yamamoto, T.; Suematsu, M. Interactions of Multiple Gas-Transducing Systems: Hallmarks and Uncertainties of CO, NO, and H2S Gas Biology. Antioxid. Redox Signal. 2010, 13, 157–192. [Google Scholar] [CrossRef] [Green Version]
- Shen, F.; Zhao, C.-S.; Shen, M.-F.; Wang, Z.; Chen, G. The role of hydrogen sulfide in gastric mucosal damage. Med. Gas. Res. 2019, 9, 88–92. [Google Scholar] [CrossRef]
- Flannigan, K.; Wallace, K.L.F.A.J.L. Hydrogen Sulfide-Based Anti-Inflammatory and Chemopreventive Therapies: An Experimental Approach. Curr. Pharm. Des. 2015, 21, 3012–3022. [Google Scholar] [CrossRef]
- Zanardo, R.C.O.; Brancaleone, V.; Distrutti, E.; Fiorucci, S.; Cirino, G.; Wallace, J.L. Hydrogen sulfide is an endogenous modulator of leukocyte-mediated inflammation. FASEB J. 2006, 20, 2118–2120. [Google Scholar] [CrossRef]
- Wang, R. The Gasotransmitter Role of Hydrogen Sulfide. Antioxid. Redox Signal. 2003, 5, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Two’s company, three’s a crowd: Can H2S be the third endogenous gaseous transmitter? FASEB J. 2002, 16, 1792–1798. [Google Scholar] [CrossRef] [Green Version]
- Magierowska, K.; Brzozowski, T.; Magierowski, M. Emerging role of carbon monoxide in regulation of cellular pathways and in the maintenance of gastric mucosal integrity. Pharmacol. Res. 2018, 129, 56–64. [Google Scholar] [CrossRef]
- Dilek, N.; Papapetropoulos, A.; Toliver-Kinsky, T.; Szabo, C. Hydrogen sulfide: An endogenous regulator of the immune system. Pharmacol. Res. 2020, 161, 105119. [Google Scholar] [CrossRef]
- Jasnos, K.; Magierowski, M.; Kwiecień, S.; Brzozowski, T. Carbon monoxide in human physiology—Its role in the gastrointestinal tract. Postępy Higieny i Medycyny Doświadczalnej 2014, 68, 101–109. [Google Scholar] [CrossRef]
- Stipanuk, M.H.; Beck, P.W. Characterization of the enzymic capacity for cysteine desulphhydration in liver and kidney of the rat. Biochem. J. 1982, 206, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Swaroop, M.; Bradley, K.; Ohura, T.; Tahara, T.; Roper, M.D.; Rosenberg, L.E.; Kraus, J.P. Rat cystathionine beta-synthase. Gene organization and alternative splicing. J. Biol. Chem. 1992, 267, 11455–11461. [Google Scholar] [CrossRef]
- Shibuya, N.; Mikami, Y.; Kimura, Y.; Nagahara, N.; Kimura, H. Vascular Endothelium Expresses 3-Mercaptopyruvate Sulfurtransferase and Produces Hydrogen Sulfide. J. Biochem. 2009, 146, 623–626. [Google Scholar] [CrossRef]
- Singh, S.B.; Lin, H.C. Hydrogen Sulfide in Physiology and Diseases of the Digestive Tract. Microorganisms 2015, 3, 866–889. [Google Scholar] [CrossRef] [Green Version]
- Su, Q.; Huang, Y.-S.; Xie, N.; Su, J.; Huang, C.; Liao, Q.-J. Diallyl disulfide inhibits the proliferation of HT-29 human colon cancer cells by inducing differentially expressed genes. Mol. Med. Rep. 2011, 4, 553–559. [Google Scholar] [CrossRef]
- Fiorucci, S.; Antonelli, E.; Distrutti, E.; Rizzo, G.; Mencarelli, A.; Orlandi, S.; Zanardo, R.; Renga, B.; Di Sante, M.; Morelli, A.; et al. Inhibition of Hydrogen Sulfide Generation Contributes to Gastric Injury Caused by Anti-Inflammatory Nonsteroidal Drugs. Gastroenterology 2005, 129, 1210–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magierowski, M.; Magierowska, K.; Hubalewska-Mazgaj, M.; Sliwowski, Z.; Pajdo, R.; Ginter, G.; Kwiecien, S.; Brzozowski, T. Exogenous and Endogenous Hydrogen Sulfide Protects Gastric Mucosa against the Formation and Time-Dependent Development of Ischemia/Reperfusion-Induced Acute Lesions Progressing into Deeper Ulcerations. Molecules 2017, 22, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, L.-X.; Geng, B.; Du, J.-B.; Tang, C.-S. Hydrogen Sulphide-Induced Hypothermia Attenuates Stress-Related Ulceration in Rats. Clin. Exp. Pharmacol. Physiol. 2007, 35, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Dicay, M.; McKnight, W.; Martin, G.R. Hydrogen sulfide enhances ulcer healing in rats. FASEB J. 2007, 21, 4070–4076. [Google Scholar] [CrossRef]
- Ise, F.; Takasuka, H.; Hayashi, S.; Takahashi, K.; Koyama, M.; Aihara, E.; Takeuchi, K. Stimulation of duodenal HCO3− secretion by hydrogen sulphide in rats: Relation to prostaglandins, nitric oxide and sensory neurones. Acta Physiol. 2010, 201, 117–126. [Google Scholar] [CrossRef]
- Wallace, J.L.; Nagy, P.; Feener, T.D.; Allain, T.; Ditrói, T.; Vaughan, D.J.; Muscara, M.N.; de Nucci, G.; Buret, A.G. A proof-of-concept, Phase 2 clinical trial of the gastrointestinal safety of a hydrogen sulfide-releasing anti-inflammatory drug. Br. J. Pharmacol. 2020, 177, 769–777. [Google Scholar] [CrossRef]
- Zheng, Y.; Yueqin, Z.; Ji, K.; Pan, Z.; Chittavong, V.; Wang, B. Esterase-Sensitive Prodrugs with Tunable Release Rates and Direct Generation of Hydrogen Sulfide. Angew. Chem. Int. Ed. 2016, 55, 4514–4518. [Google Scholar] [CrossRef]
- Bakalarz, D.; Surmiak, M.; Yang, X.; Wójcik, D.; Korbut, E.; Śliwowski, Z.; Ginter, G.; Buszewicz, G.; Brzozowski, T.; Cieszkowski, J.; et al. Organic carbon monoxide prodrug, BW-CO-111, in protection against chemically-induced gastric mucosal damage. Acta Pharm. Sin. B 2021, 11, 456–475. [Google Scholar] [CrossRef]
- Da Silva Hage-Melim, L.I.; Federico, L.B.; de Oliveira, N.K.S.; Francisco, V.C.C.; Correia, L.C.; de Lima, H.B.; Gomes, S.Q.; Barcelos, M.P.; Francischini, I.A.G.; de Paula de Silva, C.H.T. Virtual screening, ADME/Tox predictions and the drug repurposing concept for future use of old drugs against the COVID-19. Life Sci. 2020, 256, 117963. [Google Scholar] [CrossRef]
- Sun, H.; Yang, J.; Shi, Y.; Wang, Y.; Li, C.; Zhu, M. Hydrogen sulfide in the nucleus tractus solitarii regulates gastric acid secretion in rats. J. Physiol. Pharmacol. 2020, 71, 1–5. [Google Scholar]
- Cacanyiova, S.; Majzunova, M.; Golas, S.; Berenyiova, A. The role of perivascular adipose tissue and endogenous hydrogen sulfide in vasoactive responses of isolated mesenteric arteries in normotensive and spontaneously hypertensive rats. J. Physiol. Pharmacol. 2019, 70, 295–306. [Google Scholar]
- Lasheen, N.N.; Elayat, W.M.; Elrefai, M.F.M.; Zaki, W.S.; Ahmed, E.H.; El Sheikh, R.M.N.; Rayas, D.S.A.A.; Gad, F.R.S. Possible role of garlic oil in ameliorating renal injury after liver ischemia/reperfusion in rats. J. Physiol. Pharmacol. 2020, 70, 765–778. [Google Scholar]
- Gemici, B.; Wallace, J.L. Anti-inflammatory and Cytoprotective Properties of Hydrogen Sulfide. In Regulated Cell Death Part A: Apoptotic Mechanisms; Elsevier BV: Amsterdam, The Netherlands, 2015; Volume 555, pp. 169–193. [Google Scholar]
- Lazarević, M.; Mazzon, E.; Momčilović, M.; Basile, M.S.; Colletti, G.; Petralia, M.C.; Bramanti, P.; Nicoletti, F.; Miljković, Đ. The H2S Donor GYY4137 Stimulates Reactive Oxygen Species Generation in BV2 Cells While Suppressing the Secretion of TNF and Nitric Oxide. Molecules 2018, 23, 2966. [Google Scholar] [CrossRef] [Green Version]
- Magierowski, M.; Jasnos, K.; Kwiecień, S.; Brzozowski, T. Role of hydrogen sulfide in the physiology of gastrointestinal tract and in the mechanism of gastroprotection. Postępy Higieny i Medycyny Doświadczalnej 2013, 67, 150–156. [Google Scholar] [CrossRef]
- Yu, L.; Park, B.M.; Ahn, Y.J.; Lee, G.-J.; Kim, S.H. Hydrogen sulfide donor, NaHS, stimulates ANP secretion via the KATP channel and the NOS/sGC pathway in rat atria. Peptides 2019, 111, 89–97. [Google Scholar] [CrossRef]
- De Araújo, S.; Oliveira, A.P.; Sousa, F.B.; Souza, L.K.; Pacheco, G.; Filgueiras, M.C.; Nicolau, L.A.; Brito, G.A.C.; Cerqueira, G.S.; Silva, R.O.; et al. AMPK activation promotes gastroprotection through mutual interaction with the gaseous mediators H2S, NO, and CO. Nitric Oxide 2018, 78, 60–71. [Google Scholar] [CrossRef]
- Mard, S.A.; Neisi, N.; Solgi, G.; Hassanpour, M.; Darbor, M.; Maleki, M. Gastroprotective Effect of NaHS Against Mucosal Lesions Induced by Ischemia–Reperfusion Injury in Rat. Dig. Dis. Sci. 2012, 57, 1496–1503. [Google Scholar] [CrossRef]
- Medeiros, J.V.R.; Bezerra, V.H.; Gomes, A.S.; Barbosa, A.L.R.; Lima-Júnior, R.C.P.; Soares, P.M.G.; Brito, G.A.C.; Ribeiro, R.A.; Cunha, F.Q.; Souza, M.H.L.P. Hydrogen Sulfide Prevents Ethanol-Induced Gastric Damage in Mice: Role of ATP-Sensitive Potassium Channels and Capsaicin-Sensitive Primary Afferent Neurons. J. Pharmacol. Exp. Ther. 2009, 330, 764–770. [Google Scholar] [CrossRef]
- Sun, H.-Z.; Zheng, S.; Lu, K.; Hou, F.-T.; Bi, J.-X.; Liu, X.-L.; Wang, S.-S. Hydrogen sulfide attenuates gastric mucosal injury induced by restraint water-immersion stressviaactivation of KATPchannel and NF-κB dependent pathway. World J. Gastroenterol. 2017, 23, 87–92. [Google Scholar] [CrossRef]
- Jeddi, S.; Gheibi, S.; Kashfi, K.; Carlström, M.; Ghasemi, A. Dose-Dependent Effects of Long-Term Administration of Hydrogen Sulfide on Myocardial Ischemia–Reperfusion Injury in Male Wistar Rats: Modulation of RKIP, NF-κB, and Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 1415. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Ji, X.; Ji, K.; Wang, B. Hydrogen sulfide prodrugs—A review. Acta Pharm. Sin. B 2015, 5, 367–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Whiteman, M.; Guan, Y.Y.; Neo, K.L.; Cheng, Y.; Lee, S.W.; Zhao, Y.; Baskar, R.; Tan, C.-H.; Moore, P.K. Characterization of a Novel, Water-Soluble Hydrogen Sulfide–Releasing Molecule (GYY4137). Circulation 2008, 117, 2351–2360. [Google Scholar] [CrossRef] [Green Version]
- Gerő, D.; Torregrossa, R.; Perry, A.; Waters, A.; Le-Trionnaire, S.; Whatmore, J.L.; Wood, M.; Whiteman, M. The novel mitochondria-targeted hydrogen sulfide (H2S) donors AP123 and AP39 protect against hyperglycemic injury in microvascular endothelial cells in vitro. Pharmacol. Res. 2016, 113, 186–198. [Google Scholar] [CrossRef]
- Sakuma, S.; Minamino, S.; Takase, M.; Ishiyama, Y.; Hosokura, H.; Kohda, T.; Ikeda, Y.; Fujimoto, Y. Hydrogen sulfide donor GYY4137 suppresses proliferation of human colorectal cancer Caco-2 cells by inducing both cell cycle arrest and cell death. Heliyon 2019, 5, e02244. [Google Scholar] [CrossRef] [Green Version]
- Karwi, Q.G.; Bornbaum, J.; Boengler, K.; Torregrossa, R.; Whiteman, M.; Wood, M.E.; Schulz, R.; Baxter, G.F. AP39, a mitochondria-targeting hydrogen sulfide (H2S) donor, protects against myocardial reperfusion injury independently of salvage kinase signalling. Br. J. Pharmacol. 2017, 174, 287–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, S.K.; Muscara, M.N.; Allain, T.; Dallazen, J.; Gonzaga, L.; Buret, A.G.; Vaughan, D.J.; Fowler, C.J.; De Nucci, G.; Wallace, J.L. Enhanced Analgesic Effects and Gastrointestinal Safety of a Novel, Hydrogen Sulfide-Releasing Anti-Inflammatory Drug (ATB-352): A Role for Endogenous Cannabinoids. Antioxid. Redox Signal. 2020, 33, 1003–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, W.L. Distribution of Esterase in Gastric Mucosa. J. Gen. Physiol. 1954, 38, 141–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spenney, J.G. Acetylsalicylic acid hydrolase of gastric mucosa. Am. J. Physiol. Metab. 1978, 234, E606-10. [Google Scholar] [CrossRef]
- Magierowski, M.; Hubalewska-Mazgaj, M.; Magierowska, K.; Wojcik, D.; Sliwowski, Z.; Kwiecien, S.; Brzozowski, T. Nitric oxide, afferent sensory nerves, and antioxidative enzymes in the mechanism of protection mediated by tricarbonyldichlororuthenium(II) dimer and sodium hydrosulfide against aspirin-induced gastric damage. J. Gastroenterol. 2018, 53, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Magierowska, K.; Bakalarz, D.; Wójcik, D.; Korbut, E.; Danielak, A.; Głowacka, U.; Pajdo, R.; Buszewicz, G.; Ginter, G.; Surmiak, M.; et al. Evidence for Cytoprotective Effect of Carbon Monoxide Donor in the Development of Acute Esophagitis Leading to Acute Esophageal Epithelium Lesions. Cells 2020, 9, 1203. [Google Scholar] [CrossRef]
- Vrankova, S.; Zemancikova, A.; Torok, J.; Pechanova, O. Effect of low dose L-NAME pretreatment on nitric oxide/reactive oxygen species balance and vasoactivity in L-NAME/salt-induced hypertensive rats. J. Physiol. Pharmacol. 2019, 70, 535–544. [Google Scholar]
- An, J.M.; Kang, E.A.; Han, Y.M.; Kim, Y.S.; Hong, Y.G.; Hah, B.S.; Hong, S.P.; Hahm, K.B. Dietary threonine prevented stress-related mucosal diseases in rats. J. Physiol. Pharmacol. 2019, 70, 467–478. [Google Scholar]
- Zhang, C.-Y.; Li, X.-H.; Zhang, T.; Fu, J.; Cui, X.-D. Hydrogen sulfide upregulates heme oxygenase-1 expression in rats with volume overload-induced heart failure. Biomed. Rep. 2013, 1, 454–458. [Google Scholar] [CrossRef]
- Magierowski, M.; Magierowska, K.; Hubalewska-Mazgaj, M.; Sliwowski, Z.; Ginter, G.; Pajdo, R.; Chmura, A.; Kwiecien, S.; Brzozowski, T. Carbon monoxide released from its pharmacological donor, tricarbonyldichlororuthenium (II) dimer, accelerates the healing of pre-existing gastric ulcers. Br. J. Pharmacol. 2017, 174, 3654–3668. [Google Scholar] [CrossRef] [PubMed]
- Magierowska, K.; Korbut, E.; Hubalewska-Mazgaj, M.; Surmiak, M.; Chmura, A.; Bakalarz, D.; Buszewicz, G.; Wójcik, D.; Śliwowski, Z.; Ginter, G.; et al. Oxidative gastric mucosal damage induced by ischemia/reperfusion and the mechanisms of its prevention by carbon monoxide-releasing tricarbonyldichlororuthenium (II) dimer. Free Radic. Biol. Med. 2019, 145, 198–208. [Google Scholar] [CrossRef]
- Peinnequin, A.; Mouret, C.; Birot, O.; Alonso, A.; Mathieu, J.; Clarençon, D.; Agay, D.; Chancerelle, Y.; Multon, E. Rat pro-inflammatory cytokine and cytokine related mRNA quantification by real-time polymerase chain reaction using SYBR green. BMC Immunol. 2004, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | GBF [% of Control] |
|---|---|
| Intact | 99.99 ± 2.371 |
| Vehicle + ASA | 58.45 ± 2.795 * |
| BW-HS-101 + ASA | 81.16 ± 1.232 # |
| BW-iHS-101 + ASA | 70.05 ± 2.646 #^ |
| Lactone + ASA | 64.25 ± 1.963 |
| BW-HS-101 + L-NNA + ASA | 66.18 ± 3.206 ^ |
| BW-HS-101 + ZnPP + ASA | 84.54 ± 4.113 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakalarz, D.; Korbut, E.; Yuan, Z.; Yu, B.; Wójcik, D.; Danielak, A.; Magierowska, K.; Kwiecień, S.; Brzozowski, T.; Marcinkowska, M.; et al. Novel Hydrogen Sulfide (H2S)-Releasing BW-HS-101 and Its Non-H2S Releasing Derivative in Modulation of Microscopic and Molecular Parameters of Gastric Mucosal Barrier. Int. J. Mol. Sci. 2021, 22, 5211. https://doi.org/10.3390/ijms22105211
Bakalarz D, Korbut E, Yuan Z, Yu B, Wójcik D, Danielak A, Magierowska K, Kwiecień S, Brzozowski T, Marcinkowska M, et al. Novel Hydrogen Sulfide (H2S)-Releasing BW-HS-101 and Its Non-H2S Releasing Derivative in Modulation of Microscopic and Molecular Parameters of Gastric Mucosal Barrier. International Journal of Molecular Sciences. 2021; 22(10):5211. https://doi.org/10.3390/ijms22105211
Chicago/Turabian StyleBakalarz, Dominik, Edyta Korbut, Zhengnan Yuan, Bingchen Yu, Dagmara Wójcik, Aleksandra Danielak, Katarzyna Magierowska, Slawomir Kwiecień, Tomasz Brzozowski, Monika Marcinkowska, and et al. 2021. "Novel Hydrogen Sulfide (H2S)-Releasing BW-HS-101 and Its Non-H2S Releasing Derivative in Modulation of Microscopic and Molecular Parameters of Gastric Mucosal Barrier" International Journal of Molecular Sciences 22, no. 10: 5211. https://doi.org/10.3390/ijms22105211