
The Expression Characteristics of NPF Genes and Their Response to Vernalization and Nitrogen Deficiency in Rapeseed

Abstract
:1. Introduction
2. Results
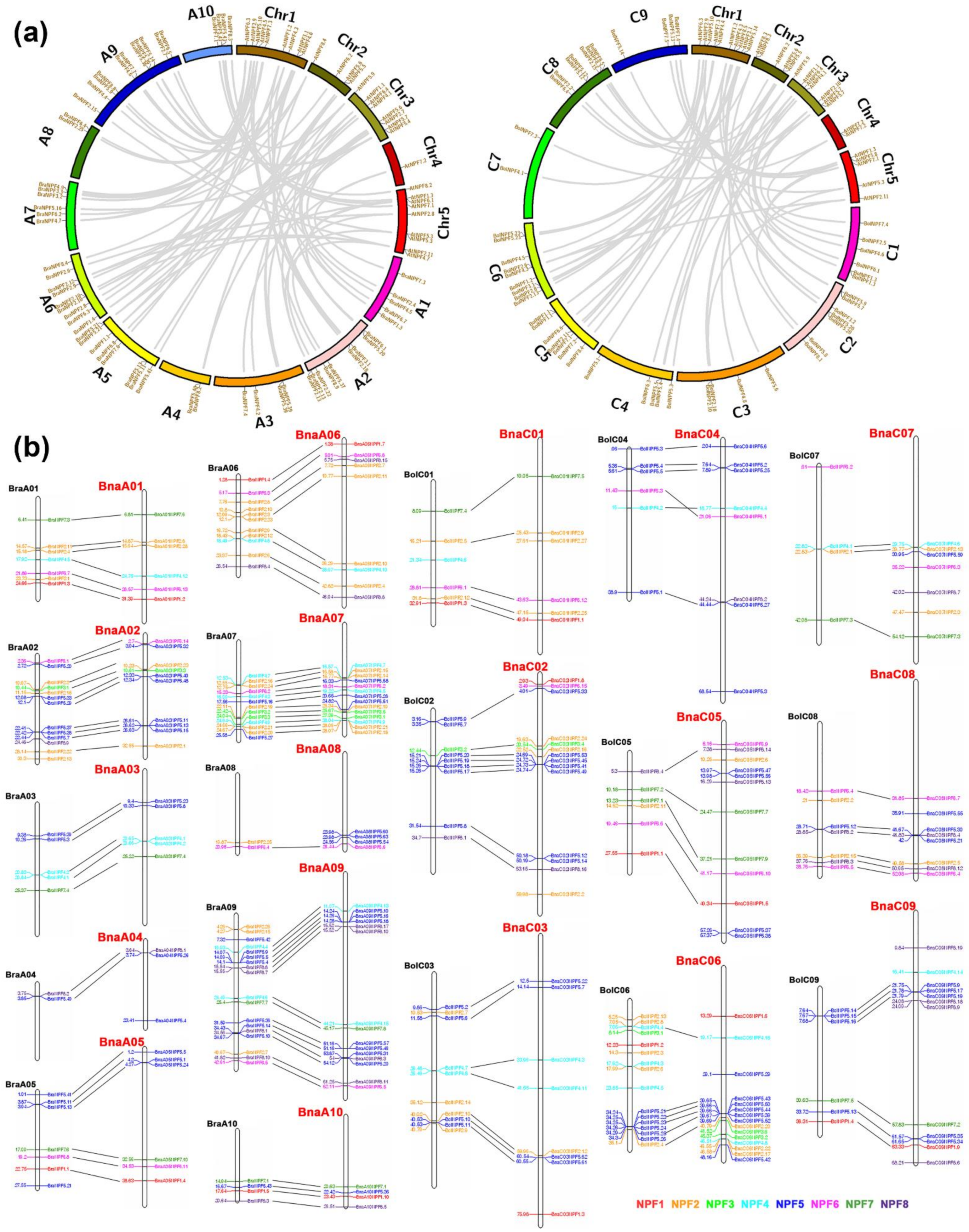
2.1. Distribution and Synteny Analysis of NPF Genes in Four Brassica Species
2.2. B. napus Genome Possessed the Most NPF Genes
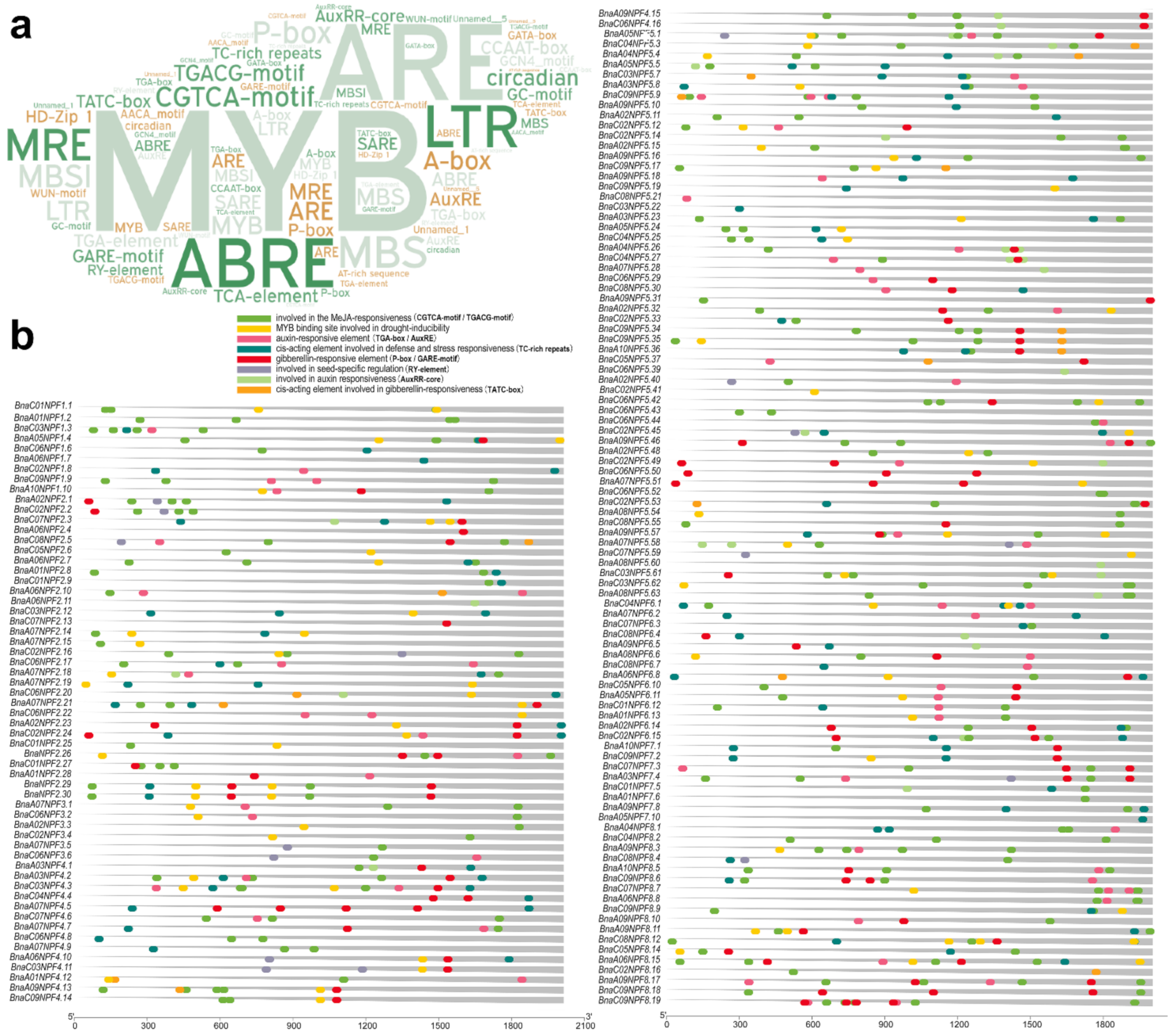
2.3. BnaNPF Gene Owning the PTR2 Functional Domain Might Be Regulated by Multiple Phytohormones
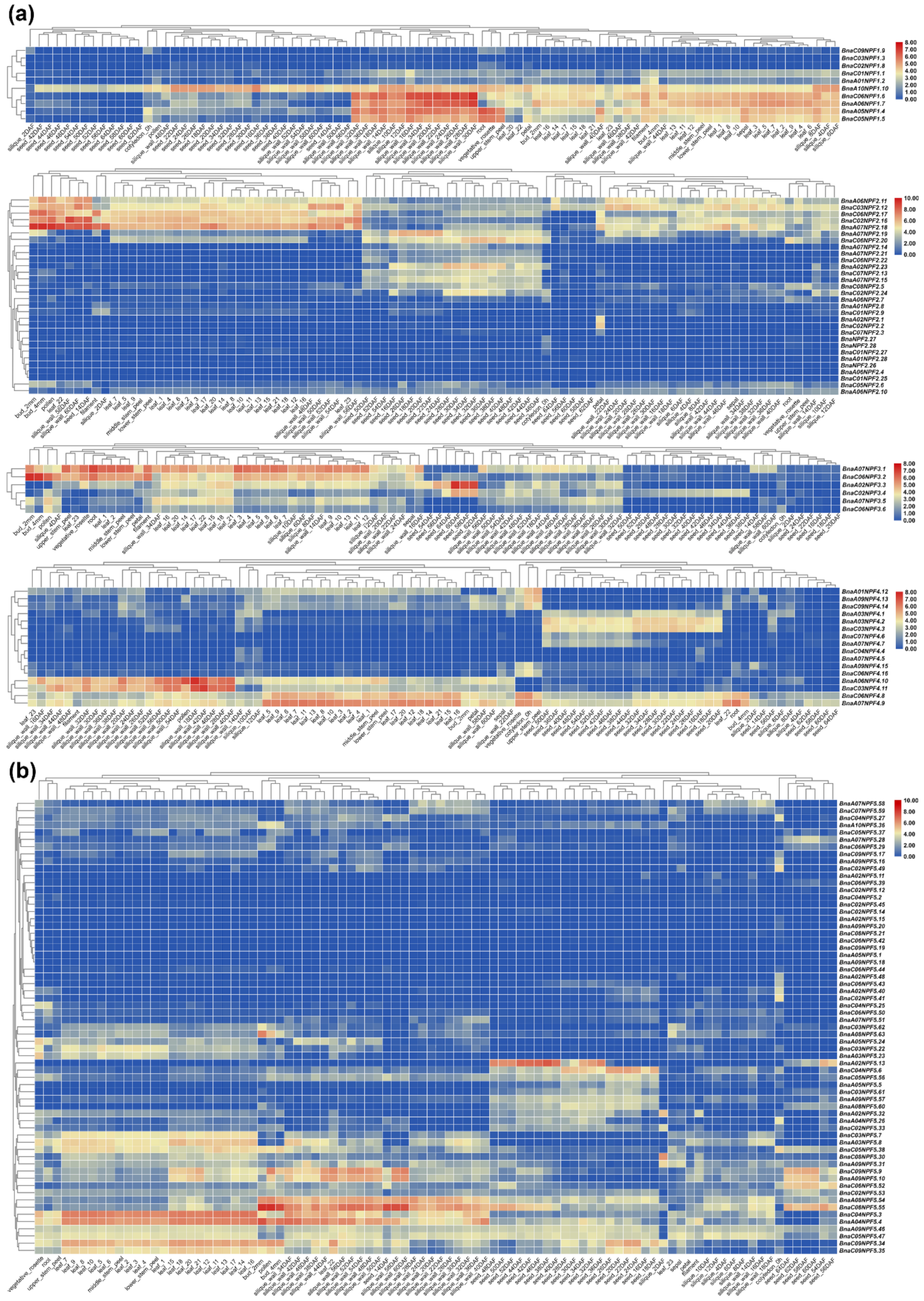
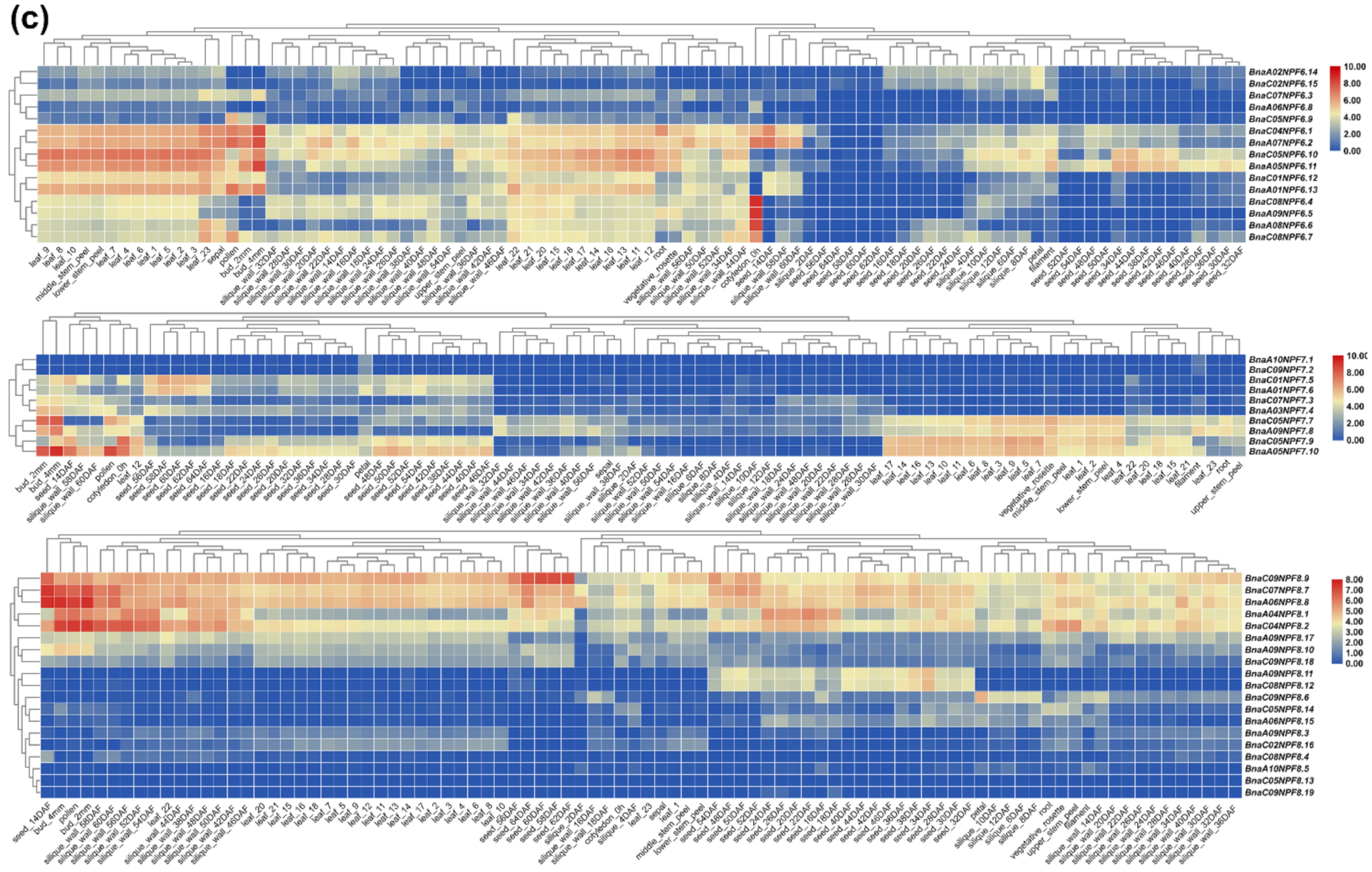
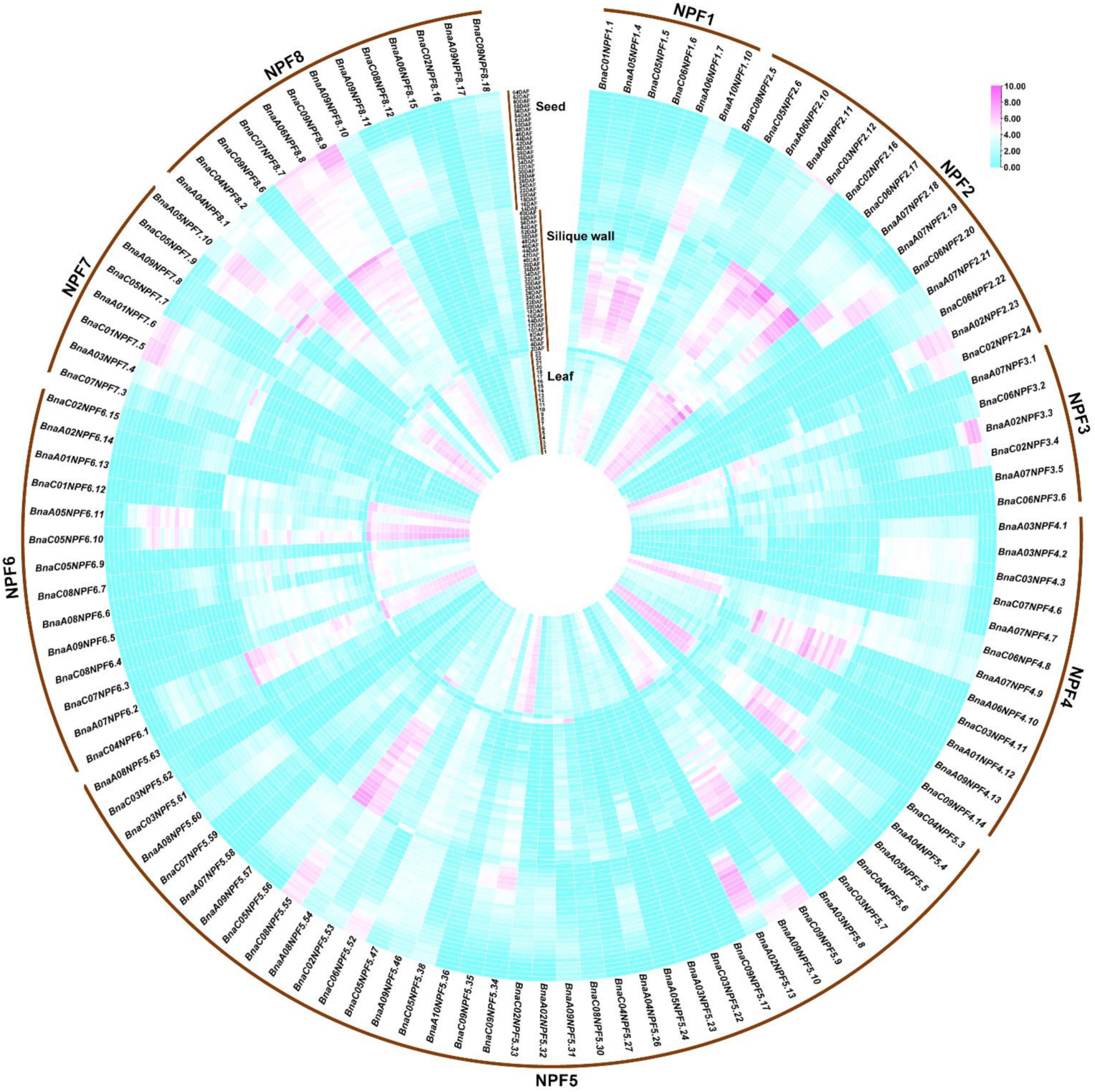
2.4. Gene Expression Pattern Analysis of NPF Genes in Diverse Tissues of B. napus
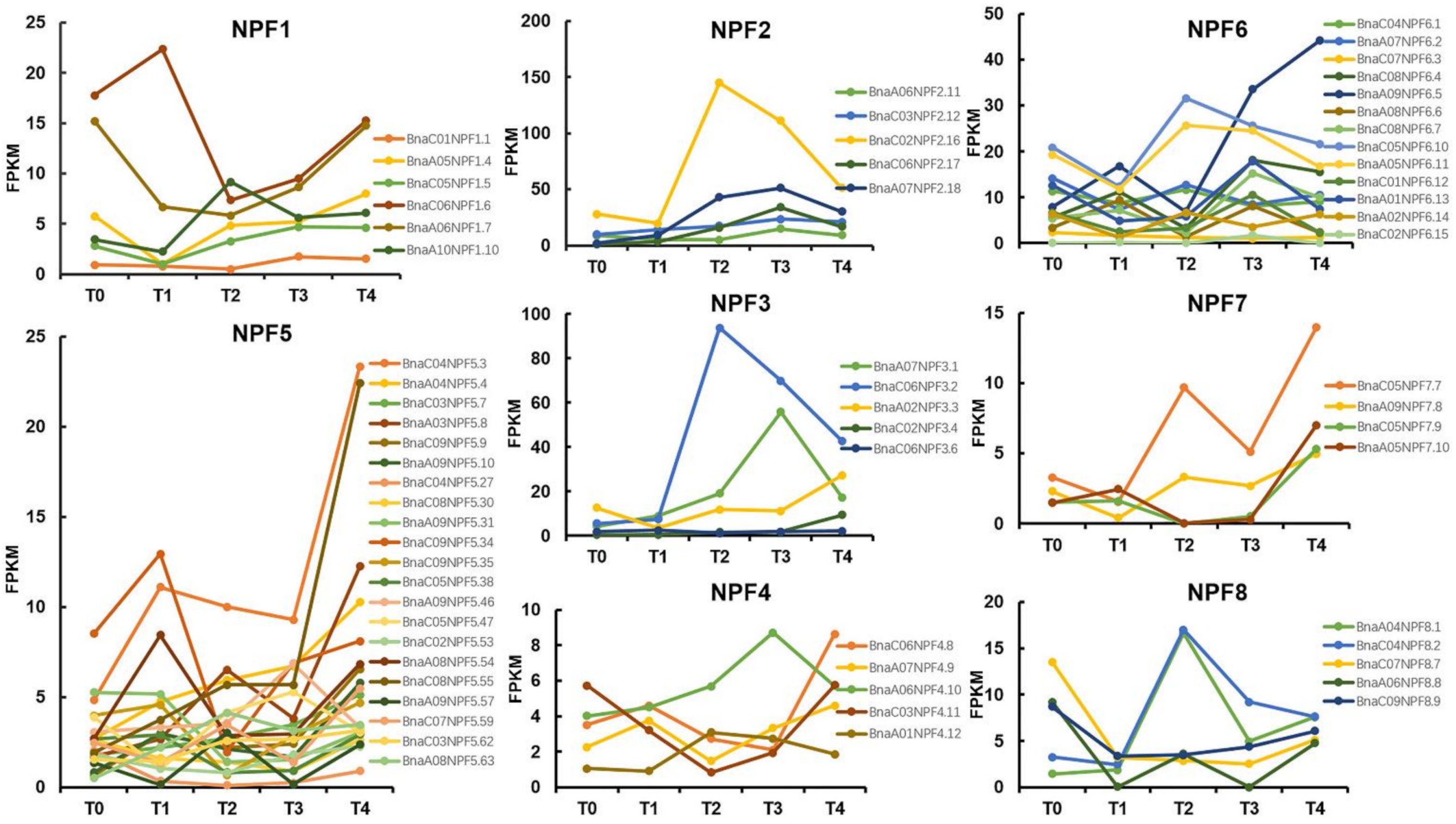
2.5. Expression Dynamic of NPF Genes during the Growth of B. napus under Vernalization
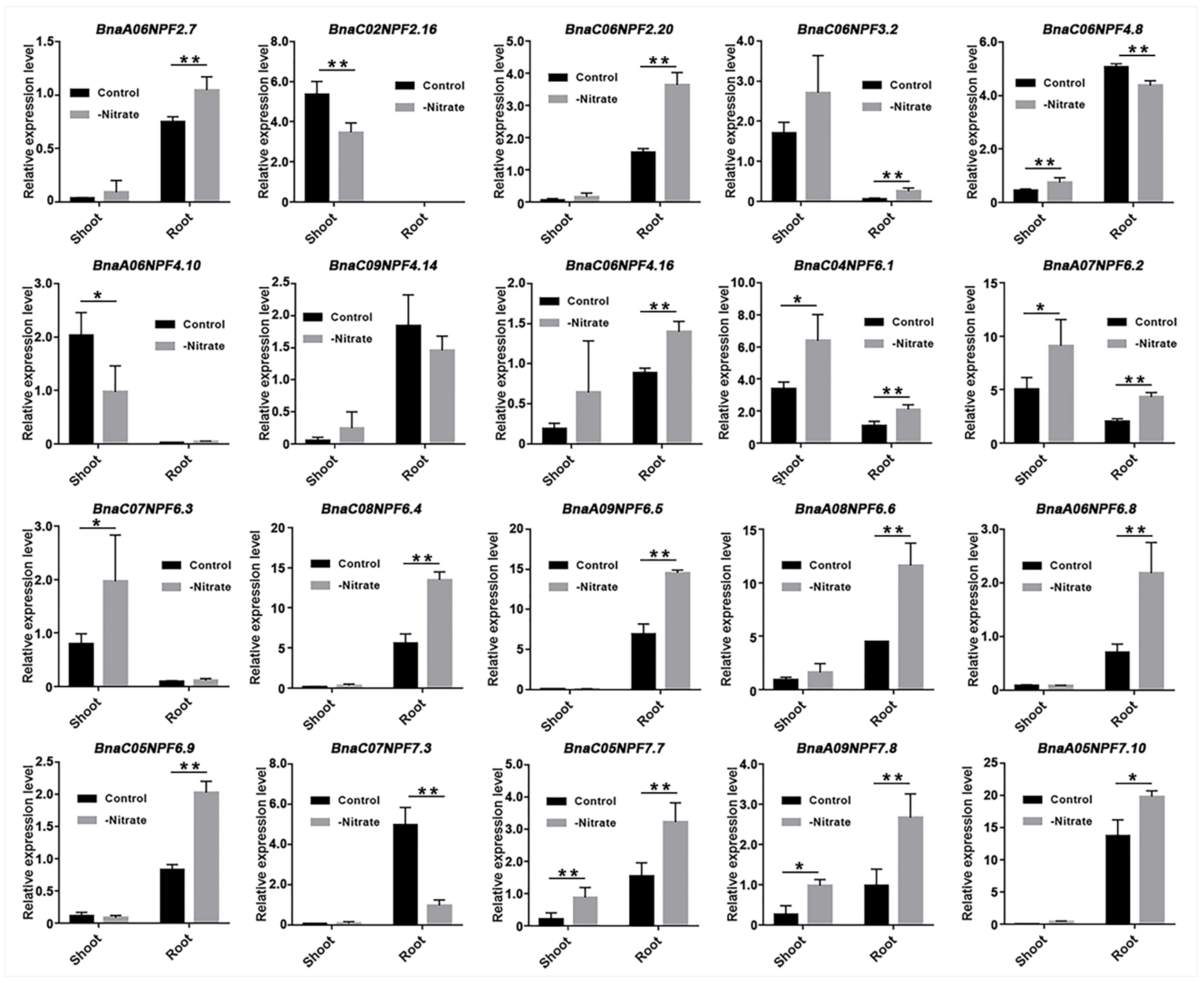
2.6. Transcriptional Analysis of BnaNPF Genes under Nitrate Deficiency
3. Discussion
4. Materials and Methods
4.1. Data Resource Related to NPF Gene Acquisition
4.2. Multiple Sequence Alignment and BnaNPF Genes Nomenclature
4.3. Chromosomal Location and Syntenic Analysis
4.4. Functional Domain Validation and Cis-Acting Regulatory Elements (CREs) Prediction
4.5. Identification of the Expression Pattern of BnaNPF Genes in B. napus
4.6. Expression Analysis of BnaNPF Genes under Low Nitrate Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leran, S.; Varala, K.; Boyer, J.C.; Chiurazzi, M.; Crawford, N.; Daniel-Vedele, F.; David, L.; Dickstein, R.; Fernandez, E.; Forde, B.; et al. A Unified Nomenclature of Nitrate Transporter 1/Peptide Transporter Family Members in Plants. Trends. Plant. Sci. 2014, 19, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Daniel, H.; Spanier, B.; Kottra, G.; Weitz, D. From Bacteria to Man: Archaic Proton-Dependent Peptide Transporters at Work. Physiology (Bethesda) 2006, 21, 93–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meredith, D. Review. The Mammalian Proton-Coupled Peptide Cotransporter PEPT1: Sitting on the Transporter-Channel Fence? Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2009, 364, 203–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsay, Y.F.; Schroeder, J.I.; Feldmann, K.A.; Crawford, N.M. The Herbicide Sensitivity Gene Chl1 of Arabidopsis Encodes a Nitrate-Inducible Nitrate Transporter. Cell 1993, 72, 705–713. [Google Scholar] [CrossRef]
- Tsay, Y.F. Plant Science How to Switch Affinity. Nature 2014, 507, 44–45. [Google Scholar] [CrossRef]
- Sun, J.; Zheng, N. Molecular Mechanism Underlying the Plant NRT1.1 Dual-Affinity Nitrate Transporter. Front. Physiol. 2015, 6, 386. [Google Scholar] [CrossRef]
- Tegeder, M.; Rentsch, D. Uptake and Partitioning of Amino Acids and Peptides. Mol. Plant 2010, 3, 997–1011. [Google Scholar] [CrossRef]
- Corratge-Faillie, C.; Lacombe, B. Substrate (Un)Specificity of Arabidopsis NRT1/PTR Family (NPF) Proteins. J. Exp. Bot. 2017, 68, 3107–3113. [Google Scholar] [CrossRef]
- Krouk, G.; Lacombe, B.; Bielach, A.; Perrine-Walker, F.; Malinska, K.; Mounier, E.; Hoyerova, K.; Tillard, P.; Leon, S.; Ljung, K.; et al. Nitrate-Regulated Auxin Transport by NRT1.1 Defines a Mechanism for Nutrient Sensing in Plants. Dev. Cell. 2010, 18, 927–937. [Google Scholar] [CrossRef]
- Kanno, Y.; Hanada, A.; Chiba, Y.; Ichikawa, T.; Nakazawa, M.; Matsui, M.; Koshiba, T.; Kamiya, Y.; Seo, M. Identification of an Abscisic Acid Transporter by Functional Screening Using the Receptor Complex as a Sensor. Proc. Natl. Acad. Sci. USA 2012, 109, 9653–9658. [Google Scholar] [CrossRef] [Green Version]
- Nour-Eldin, H.H.; Andersen, T.G.; Burow, M.; Madsen, S.R.; Jorgensen, M.E.; Olsen, C.E.; Dreyer, I.; Hedrich, R.; Geiger, D.; Halkier, B.A. NRT/PTR Transporters Are Essential for Translocation of Glucosinolate Defence Compounds to Seeds. Nature 2012, 488, 531–534. [Google Scholar] [CrossRef]
- Boursiac, Y.; Leran, S.; Corratge-Faillie, C.; Gojon, A.; Krouk, G.; Lacombe, B. ABA Transport and Transporters. Trends Plant Sci. 2013, 18, 325–333. [Google Scholar] [CrossRef]
- Wang, H.; Wan, Y.; Buchner, P.; King, R.; Ma, H.; Hawkesford, M.J. Phylogeny and Gene Expression of the Complete Nitrate Transporter 1/Peptide Transporter Family in Triticum aestivum. J. Exp. Bot. 2020, 71, 4531–4546. [Google Scholar] [CrossRef]
- Chiba, Y.; Shimizu, T.; Miyakawa, S.; Kanno, Y.; Koshiba, T.; Kamiya, Y.; Seo, M. Identification of Arabidopsis thaliana NRT1/PTR Family (NPF) Proteins Capable of Transporting Plant Hormones. J. Plant. Res. 2015, 128, 679–686. [Google Scholar] [CrossRef]
- Krouk, G.; Crawford, N.M.; Coruzzi, G.M.; Tsay, Y.F. Nitrate Signaling: Adaptation to Fluctuating Environments. Curr. Opin. Plant. Biol. 2010, 13, 266–273. [Google Scholar] [CrossRef]
- Bouguyon, E.; Brun, F.; Meynard, D.; Kubes, M.; Pervent, M.; Leran, S.; Lacombe, B.; Krouk, G.; Guiderdoni, E.; Zazimalova, E.; et al. Multiple Mechanisms of Nitrate Sensing by Arabidopsis Nitrate Transceptor NRT1.1. Nat. Plants 2015, 1, 15015. [Google Scholar] [CrossRef]
- Chiang, C.S.; Stacey, G.; Tsay, Y.F. Mechanisms and Functional Properties of Two Peptide Transporters, AtPTR2 and FPTR2. J. Biol. Chem. 2004, 279, 30150–30157. [Google Scholar] [CrossRef] [Green Version]
- Hammes, U.Z.; Meier, S.; Dietrich, D.; Ward, J.M.; Rentsch, D. Functional Properties of the Arabidopsis Peptide Transporters AtPTR1 and AtPTR5. J. Biol. Chem. 2010, 285, 39710–39717. [Google Scholar] [CrossRef] [Green Version]
- Yendrek, C.R.; Lee, Y.C.; Morris, V.; Liang, Y.; Pislariu, C.I.; Burkart, G.; Meckfessel, M.H.; Salehin, M.; Kessler, H.; Wessler, H.; et al. A Putative Transporter Is Essential for Integrating Nutrient and Hormone Signaling with Lateral Root Growth and Nodule Development in Medicago truncatula. Plant. J. 2010, 62, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Bagchi, R.; Salehin, M.; Adeyemo, O.S.; Salazar, C.; Shulaev, V.; Sherrier, D.J.; Dickstein, R. Functional Assessment of the Medicago Truncatula NIP/LATD Protein Demonstrates That It Is a High-Affinity Nitrate Transporter. Plant. Physiol. 2012, 160, 906–916. [Google Scholar] [CrossRef] [Green Version]
- Salehin, M.; Huang, Y.S.; Bagchi, R.; Sherrier, D.J.; Dickstein, R. Allelic Differences in Medicago Truncatula Nip/Latd Mutants Correlate with Their Encoded Proteins’ Transport Activities in Planta. Plant. Signal. Behav. 2013, 8, e22813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, P.K.; Tsay, Y.F. Two Phloem Nitrate Transporters, NRT1.11 and NRT1.12, Are Important for Redistributing Xylem-Borne Nitrate to Enhance Plant Growth. Plant. Physiol. 2013, 163, 844–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taochy, C.; Gaillard, I.; Ipotesi, E.; Oomen, R.; Leonhardt, N.; Zimmermann, S.; Peltier, J.B.; Szponarski, W.; Simonneau, T.; Sentenac, H.; et al. The Arabidopsis Root Stele Transporter NPF2.3 Contributes to Nitrate Translocation to Shoots under Salt Stress. Plant. J. 2015, 83, 466–479. [Google Scholar] [CrossRef] [PubMed]
- Almagro, A.; Lin, S.H.; Tsay, Y.F. Characterization of the Arabidopsis Nitrate Transporter NRT1.6 Reveals a Role of Nitrate in Early Embryo Development. Plant. Cell 2008, 20, 3289–3299. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, M.; Georgescu, M.N.; Takahashi, M. A Nitrite Transporter Associated with Nitrite Uptake by Higher Plant Chloroplasts. Plant. Cell Physiol. 2007, 48, 1022–1035. [Google Scholar] [CrossRef] [Green Version]
- Karim, S.; Holmstrom, K.O.; Mandal, A.; Dahl, P.; Hohmann, S.; Brader, G.; Palva, E.T.; Pirhonen, M. AtPTR3, a Wound-Induced Peptide Transporter Needed for Defence against Virulent Bacterial Pathogens in Arabidopsis. Planta 2007, 225, 1431–1445. [Google Scholar] [CrossRef]
- Leran, S.; Edel, K.H.; Pervent, M.; Hashimoto, K.; Corratge-Faillie, C.; Offenborn, J.N.; Tillard, P.; Gojon, A.; Kudla, J.; Lacombe, B. Nitrate Sensing and Uptake in Arabidopsis Are Enhanced by Abi2, a Phosphatase Inactivated by the Stress Hormone Abscisic Acid. Sci Signal. 2015, 8, ra43. [Google Scholar] [CrossRef]
- Okamoto, M.; Vidmar, J.J.; Glass, A.D. Regulation of NRT1 and NRT2 Gene Families of Arabidopsis thaliana: Responses to Nitrate Provision. Plant. Cell Physiol. 2003, 44, 304–317. [Google Scholar] [CrossRef]
- Chiu, C.C.; Lin, C.S.; Hsia, A.P.; Su, R.C.; Lin, H.L.; Tsay, Y.F. Mutation of a Nitrate Transporter, AtNRT1:4, Results in a Reduced Petiole Nitrate Content and Altered Leaf Development. Plant. Cell Physiol. 2004, 45, 1139–1148. [Google Scholar] [CrossRef]
- Morere-Le Paven, M.C.; Viau, L.; Hamon, A.; Vandecasteele, C.; Pellizzaro, A.; Bourdin, C.; Laffont, C.; Lapied, B.; Lepetit, M.; Frugier, F.; et al. Characterization of a Dual-Affinity Nitrate Transporter MtNRT1.3 in the Model Legume Medicago truncatula. J. Exp. Bot. 2011, 62, 5595–5605. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Y.; Fu, Y.L.; Pike, S.M.; Bao, J.; Tian, W.; Zhang, Y.; Chen, C.Z.; Zhang, Y.; Li, H.M.; Huang, J.; et al. The Arabidopsis Nitrate Transporter NRT1.8 Functions in Nitrate Removal from the Xylem Sap and Mediates Cadmium Tolerance. Plant. Cell 2010, 22, 1633–1646. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Z.; Lv, X.F.; Li, J.Y.; Yi, H.Y.; Gong, J.M. Arabidopsis NRT1.5 Is Another Essential Component in the Regulation of Nitrate Reallocation and Stress Tolerance. Plant. Physiol. 2012, 159, 1582–1590. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, J.; Cai, Z.Y.; Xia, K.F.; Wang, Y.Q.; Duan, J.; Zhang, M.Y. Identification and Analysis of Eight Peptide Transporter Homologs in Rice. Plant. Sci. 2010, 179, 374–382. [Google Scholar] [CrossRef]
- Komarova, N.Y.; Thor, K.; Gubler, A.; Meier, S.; Dietrich, D.; Weichert, A.; Suter Grotemeyer, M.; Tegeder, M.; Rentsch, D. AtPTR1 and AtPTR5 Transport Dipeptides in Planta. Plant. Physiol. 2008, 148, 856–869. [Google Scholar] [CrossRef]
- Choi, M.G.; Kim, E.J.; Song, J.Y.; Choi, S.B.; Cho, S.W.; Park, C.S.; Kang, C.S.; Park, Y.I. Peptide Transporter2 (PTR2) Enhances Water Uptake During Early Seed Germination in Arabidopsis thaliana. Plant. Mol. Biol. 2020, 102, 615–624. [Google Scholar] [CrossRef] [Green Version]
- Pellizzaro, A.; Alibert, B.; Planchet, E.; Limami, A.M.; Morere-Le Paven, M.C. Nitrate Transporters: An Overview in Legumes. Planta 2017, 246, 585–595. [Google Scholar] [CrossRef] [Green Version]
- Babst, B.A.; Gao, F.; Acosta-Gamboa, L.M.; Karve, A.; Schueller, M.J.; Lorence, A. Three NPF Genes in Arabidopsis Are Necessary for Normal Nitrogen Cycling under Low Nitrogen Stress. Plant. Physiol. Biochem. 2019, 143, 1–10. [Google Scholar] [CrossRef]
- Bai, H.; Euring, D.; Volmer, K.; Janz, D.; Polle, A. The Nitrate Transporter (Nrt) Gene Family in Poplar. PLoS ONE 2013, 8, e72126. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, C.; Dong, Q.; Huang, D.; Li, C.; Li, P.; Ma, F. Genome-Wide Identification and Analysis of Apple Nitrate Transporter 1/Peptide Transporter Family (NPF) Genes Reveals MdNPF6.5 Confers High Capacity for Nitrogen Uptake under Low-Nitrogen Conditions. Int. J. Mol. Sci. 2018, 19, 2761. [Google Scholar] [CrossRef] [Green Version]
- Lysak, M.A.; Koch, M.A.; Pecinka, A.; Schubert, I. Chromosome Triplication Found across the Tribe Brassiceae. Genome Res. 2005, 15, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The Genome of the Mesopolyploid Crop Species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Plant Genetics. Early Allopolyploid Evolution in the Post-Neolithic Brassica napus Oilseed Genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea Genome Reveals the Asymmetrical Evolution of Polyploid Genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Fan, G.; Hu, Q.; Zhou, Y.; Guan, M.; Tong, C.; Li, J.; Du, D.; Qi, C.; Jiang, L.; et al. The High-Quality Genome of Brassica napus Cultivar ‘Zs11’ Reveals the Introgression History in Semi-Winter Morphotype. Plant. J. 2017, 92, 452–468. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Li, S.; Shi, M.; Wang, S.; Shi, L.; Xu, F.; Ding, G. Genome-Wide Systematic Characterization of the NPF Family Genes and Their Transcriptional Responses to Multiple Nutrient Stresses in Allotetraploid Rapeseed. Int. J. Mol. Sci. 2020, 21, 5947. [Google Scholar] [CrossRef]
- Wen, J.; Li, P.F.; Ran, F.; Guo, P.C.; Zhu, J.T.; Yang, J.; Zhang, L.L.; Chen, P.; Li, J.N.; Du, H. Genome-Wide Characterization, Expression Analyses, and Functional Prediction of the NPF Family in Brassica napus. BMC Genomics 2020, 21, 871. [Google Scholar] [CrossRef]
- Wittkopp, P.J.; Kalay, G. Cis-regulatory Elements: Molecular Mechanisms and Evolutionary Processes Underlying Divergence. Nat. Rev. Genet. 2011, 6, 59–69. [Google Scholar] [CrossRef]
- Town, C.D.; Cheung, F.; Maiti, R.; Crabtree, J.; Haas, B.J.; Wortman, J.R.; Hine, E.E.; Althoff, R.; Arbogast, T.S.; Tallon, L.J.; et al. Comparative Genomics of Brassica Oleracea and Arabidopsis thaliana Reveal Gene Loss, Fragmentation, and Dispersal after Polyploidy. Plant. Cell 2006, 18, 1348–1359. [Google Scholar] [CrossRef] [Green Version]
- De Bodt, S.; Maere, S.; Van de Peer, Y. Genome Duplication and the Origin of Angiosperms. Trends Ecol. Evol. 2005, 20, 591–597. [Google Scholar] [CrossRef]
- Mun, J.H.; Kwon, S.J.; Yang, T.J.; Seol, Y.J.; Jin, M.; Kim, J.A.; Lim, M.H.; Kim, J.S.; Baek, S.; Choi, B.S.; et al. Genome-Wide Comparative Analysis of the Brassica rapa Gene Space Reveals Genome Shrinkage and Differential Loss of Duplicated Genes after Whole Genome Triplication. Genome Biol. 2009, 10, R111. [Google Scholar] [CrossRef] [Green Version]
- Jeffares, D.C.; Penkett, C.J.; Bahler, J. Rapidly Regulated Genes Are Intron Poor. Trends Genet. 2008, 24, 375–378. [Google Scholar] [CrossRef]
- Fan, S.C.; Lin, C.S.; Hsu, P.K.; Lin, S.H.; Tsay, Y.F. The Arabidopsis Nitrate Transporter NRT1.7, Expressed in Phloem, Is Responsible for Source-to-Sink Remobilization of Nitrate. Plant. Cell 2009, 21, 2750–2761. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.L.; Tsay, Y.F. Influence of Differing Nitrate and Nitrogen Availability on Flowering Control in Arabidopsis. J. Exp. Bot. 2017, 68, 2603–2609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Liu, B. Genetic Regulation of Maize Flower Development and Sex Determination. Planta 2017, 245, 1–14. [Google Scholar] [CrossRef]
- Fredes, I.; Moreno, S.; Diaz, F.P.; Gutierrez, R.A. Nitrate Signaling and the Control of Arabidopsis Growth and Development. Curr. Opin. Plant. Biol. 2019, 47, 112–118. [Google Scholar] [CrossRef]
- Andersen, T.G.; Nour-Eldin, H.H.; Fuller, V.L.; Olsen, C.E.; Burow, M.; Halkier, B.A. Integration of Biosynthesis and Long-Distance Transport Establish Organ-Specific Glucosinolate Profiles in Vegetative Arabidopsis. Plant. Cell 2013, 25, 3133–3145. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Memelink, J. Jasmonate-Responsive Transcription Factors Regulating Plant Secondary Metabolism. Biotechnol. Adv. 2016, 34, 441–449. [Google Scholar] [CrossRef]
- Li, B.; Zhao, W.; Li, D.; Chao, H.; Zhao, X.; Ta, N.; Li, Y.; Guan, Z.; Guo, L.; Zhang, L.; et al. Genetic Dissection of the Mechanism of Flowering Time Based on an Environmentally Stable and Specific Qtl in Brassica napus. Plant. Sci. 2018, 277, 296–310. [Google Scholar] [CrossRef]
- Li, H.; Fan, Y.; Yu, J.; Chai, L.; Zhang, J.; Jiang, J.; Cui, C.; Zheng, B.; Jiang, L.; Lu, K. Genome-Wide Identification of Flowering-Time Genes in Brassica Species and Reveals a Correlation between Selective Pressure and Expression Patterns of Vernalization-Pathway Genes in Brassica napus. Int. J. Mol. Sci. 2018, 19, 3632. [Google Scholar] [CrossRef] [Green Version]
- Bernier, G.; Perilleux, C. A Physiological Overview of the Genetics of Flowering Time Control. Plant. Biotechnol. J. 2005, 3, 3–16. [Google Scholar] [CrossRef]
- Vega, A.; O’Brien, J.A.; Gutierrez, R.A. Nitrate and Hormonal Signaling Crosstalk for Plant Growth and Development. Curr. Opin. Plant. Biol. 2019, 52, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.Y.; Tyerman, S.D.; Dechorgnat, J.; Ovchinnikova, E.; Dhugga, K.S.; Kaiser, B.N. Maize NPF6 Proteins Are Homologs of Arabidopsis Chl1 That Are Selective for Both Nitrate and Chloride. Plant. Cell 2017, 29, 2581–2596. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Hu, B.; Yuan, D.; Liu, Y.; Che, R.; Hu, Y.; Ou, S.; Liu, Y.; Zhang, Z.; Wang, H.; et al. Expression of the Nitrate Transporter Gene sNRT1.1A/OsNPF6.3 Confers High Yield and Early Maturation in Rice. Plant. Cell 2018, 30, 638–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Y.; Tsay, Y.F. Arabidopsis Nitrate Transporter NRT1.9 Is Important in Phloem Nitrate Transport. Plant. Cell 2011, 23, 1945–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.J.; Ye, J.; Yao, X.M.; Zhao, P.Z.; Xuan, W.; Tian, Y.L.; Zhang, Y.Y.; Xu, S.; An, H.Z.; Chen, G.M.; et al. Genome-Wide Associated Study Identifies Nac42-Activated Nitrate Transporter Conferring High Nitrogen Use Efficiency in Rice. Nat. Commun. 2019, 10, 5279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pike, S.; Gao, F.; Kim, M.J.; Kim, S.H.; Schachtman, D.P.; Gassmann, W. Members of the NPF3 Transporter Subfamily Encode Pathogen-Inducible Nitrate/Nitrite Transporters in Grapevine and Arabidopsis. Plant. Cell Physiol. 2014, 55, 162–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, L.C.; Berquin, P.; Kanno, Y.; Seo, M.; Daniel-Vedele, F.; Ferrario-Mery, S. N Availability Modulates the Role of NPF3.1, a Gibberellin Transporter, in GA-Mediated Phenotypes in Arabidopsis. Planta 2016, 244, 1315–1328. [Google Scholar] [CrossRef]
- Tal, I.; Zhang, Y.; Jorgensen, M.E.; Pisanty, O.; Barbosa, I.C.R.; Zourelidou, M.; Regnault, T.; Crocoll, C.; Olsen, C.E.; Weinstain, R.; et al. The Arabidopsis NPF3 Protein Is a GA Transporter. Nat. Com. 2016, 7. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Holub, E.B. The Arms Race Is Ancient History in Arabidopsis, the Wildflower. Nat. Rev. Genet. 2001, 2, 516–527. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, J.; Zhao, X.Q.; Wang, J.; Wong, G.K.; Yu, J. Kaks_Calculator: Calculating Ka and Ks through Model Selection and Model Averaging. Genom. Proteom. Bioinform. 2006, 4, 259–263. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. Tbtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Cui, J.Q.; Hua, Y.P.; Zhou, T.; Liu, Y.; Huang, J.Y.; Yue, C.P. Global Landscapes of the Na(+)/H(+) Antiporter (Nhx) Family Members Uncover Their Potential Roles in Regulating the Rapeseed Resistance to Salt Stress. Int. J. Mol. Sci. 2020, 21, 3429. [Google Scholar]
- Lu, K.; Li, T.; He, J.; Chang, W.; Zhang, R.; Liu, M.; Yu, M.; Fan, Y.; Ma, J.; Sun, W.; et al. Qprimerdb: A Thermodynamics-Based Gene-Specific qPCR Primer Database for 147 Organisms. Nucleic Acids Res. 2018, 46, 1229–1236. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ratio a | 0:1 | 1:0 | 1:1 | 1:2 | 1:3 | 1:5 |
|---|---|---|---|---|---|---|
| B. rapa | 2 | 9 | 55 | 16 | 1 | 1 |
| B. oleracea | 4 | 9 | 54 | 7 |
| Organism Name | NPF1 | NPF2 | NPF3 | NPF4 | NPF5 | NPF6 | NPF7 | NPF8 | Total | Genome Size (Mb) |
|---|---|---|---|---|---|---|---|---|---|---|
| Arabidopsis lyrata (D) | 3 | 14 | 1 | 9 | 17 | 4 | 3 | 5 | 56 | 202.97 |
| Arabidopsis thaliana (D) | 3 | 14 | 1 | 7 | 16 | 4 | 3 | 5 | 53 | 127.42 |
| Aquilaria agallochum (D) | 6 | 7 | 3 | 12 | 13 | 5 | 3 | 6 | 55 | 726.71 |
| Brachypodium distachyon (M) | 2 | 6 | 4 | 13 | 21 | 8 | 11 | 17 | 82 | 271.3 |
| Brassica rapa (D) | 4 | 23 | 3 | 9 | 23 | 7 | 5 | 8 | 82 | 401.93 |
| Brassica oleracea (D) | 4 | 15 | 2 | 8 | 26 | 6 | 5 | 4 | 70 | 554.98 |
| Brassica napus (D) | 10 | 30 | 6 | 16 | 63 | 15 | 10 | 19 | 169 | 976.19 |
| Carica papaya (D) | 4 | 14 | 3 | 8 | 12 | 8 | 6 | 4 | 59 | 370.42 |
| Capsella rubella (D) | 3 | 12 | 1 | 6 | 17 | 4 | 3 | 5 | 51 | 133.06 |
| Citrus clementina (D) | 9 | 7 | 3 | 9 | 17 | 6 | 4 | 4 | 59 | 301.37 |
| Citrus sinensis (D) | 8 | 7 | 3 | 10 | 17 | 6 | 4 | 4 | 59 | 319.23 |
| Cuscuta campestris (D) | 4 | 9 | 3 | 8 | 19 | 6 | 5 | 5 | 59 | 476.79 |
| Eucalyptus grandis (D) | 6 | 12 | 4 | 11 | 19 | 6 | 4 | 6 | 68 | 691.43 |
| Fragaria vesca (D) | 0 | 13 | 2 | 8 | 23 | 3 | 5 | 6 | 60 | 214.37 |
| Glycine max (D) | 13 | 14 | 6 | 22 | 41 | 11 | 14 | 13 | 134 | 927.71 |
| Gossypium raimondii (D) | 7 | 10 | 4 | 14 | 14 | 11 | 7 | 8 | 75 | 773.77 |
| Linum usitatissimum (D) | 12 | 7 | 4 | 14 | 25 | 9 | 11 | 10 | 92 | 316.17 |
| Malus domestica (D) | 2 | 34 | 4 | 21 | 44 | 17 | 8 | 9 | 139 | 1874.77 |
| Manihot esculenta (D) | 7 | 12 | 6 | 10 | 23 | 7 | 5 | 5 | 75 | 292.1 |
| Medicago truncatula (D) | 8 | 12 | 3 | 14 | 25 | 8 | 9 | 1 | 80 | 412.92 |
| Oryza sativa (M) | 3 | 6 | 5 | 12 | 29 | 6 | 11 | 21 | 93 | 389.75 |
| Phaseolus vulgaris (D) | 8 | 11 | 3 | 12 | 22 | 5 | 7 | 6 | 74 | 521.08 |
| Populus trichocarpa (D) | 15 | 9 | 5 | 12 | 26 | 6 | 5 | 7 | 85 | 434.29 |
| Prunus persica (D) | 2 | 15 | 1 | 8 | 16 | 5 | 5 | 5 | 57 | 214.22 |
| Ricinus communis (D) | 5 | 20 | 3 | 7 | 13 | 5 | 4 | 3 | 60 | 350.62 |
| Setaria italica (M) | 4 | 11 | 8 | 16 | 19 | 7 | 12 | 21 | 98 | 405.87 |
| Solanum tuberosum (D) | 17 | 10 | 2 | 15 | 8 | 9 | 4 | 8 | 73 | 772.25 |
| Solanum lycopersicum (D) | 19 | 16 | 2 | 12 | 11 | 12 | 7 | 11 | 90 | 760.07 |
| Sorghum bicolor (M) | 4 | 8 | 7 | 16 | 22 | 6 | 9 | 19 | 91 | 709.35 |
| Theobroma cacao (D) | 4 | 14 | 3 | 10 | 19 | 7 | 4 | 5 | 66 | 345.99 |
| Vitis vinifera (D) | 4 | 7 | 2 | 6 | 21 | 5 | 4 | 3 | 52 | 486.2 |
| Zea mays (M) | 4 | 4 | 6 | 12 | 17 | 8 | 12 | 16 | 79 | 2271.03 |
| Amborella trichopoda (D) | 1 | 5 | 2 | 7 | 15 | 4 | 3 | 7 | 45 | 706.60 |
| Physcomitrella patens (L) | 0 | 0 | 1 | 1 | 8 | 6 | 3 | 4 | 23 | 472.081 |
| Selaginella moellendorffii (L) | 0 | 0 | 4 | 4 | 11 | 6 | 5 | 16 | 46 | 212.315 |
| Selaginella moellendorffii (L) | 0 | 0 | 4 | 4 | 11 | 6 | 5 | 16 | 46 | 212.315 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chao, H.; He, J.; Cai, Q.; Zhao, W.; Fu, H.; Hua, Y.; Li, M.; Huang, J. The Expression Characteristics of NPF Genes and Their Response to Vernalization and Nitrogen Deficiency in Rapeseed. Int. J. Mol. Sci. 2021, 22, 4944. https://doi.org/10.3390/ijms22094944
Chao H, He J, Cai Q, Zhao W, Fu H, Hua Y, Li M, Huang J. The Expression Characteristics of NPF Genes and Their Response to Vernalization and Nitrogen Deficiency in Rapeseed. International Journal of Molecular Sciences. 2021; 22(9):4944. https://doi.org/10.3390/ijms22094944
Chicago/Turabian StyleChao, Hongbo, Jianjie He, Qianqian Cai, Weiguo Zhao, Hong Fu, Yingpeng Hua, Maoteng Li, and Jinyong Huang. 2021. "The Expression Characteristics of NPF Genes and Their Response to Vernalization and Nitrogen Deficiency in Rapeseed" International Journal of Molecular Sciences 22, no. 9: 4944. https://doi.org/10.3390/ijms22094944