
Plastid Anionic Lipids Are Essential for the Development of Both Photosynthetic and Non-Photosynthetic Organs in Arabidopsis thaliana


Abstract
:1. Introduction
2. Results
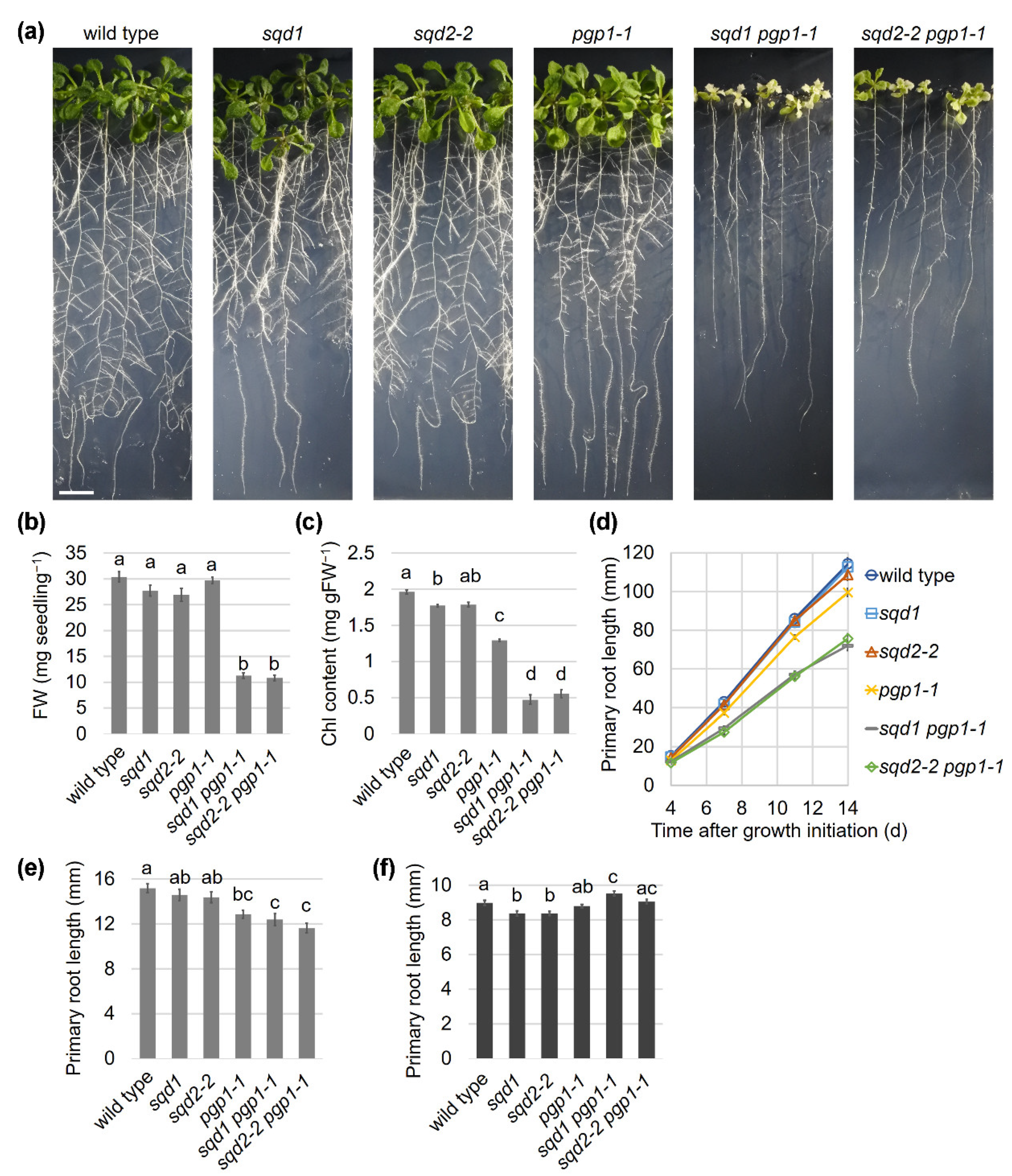
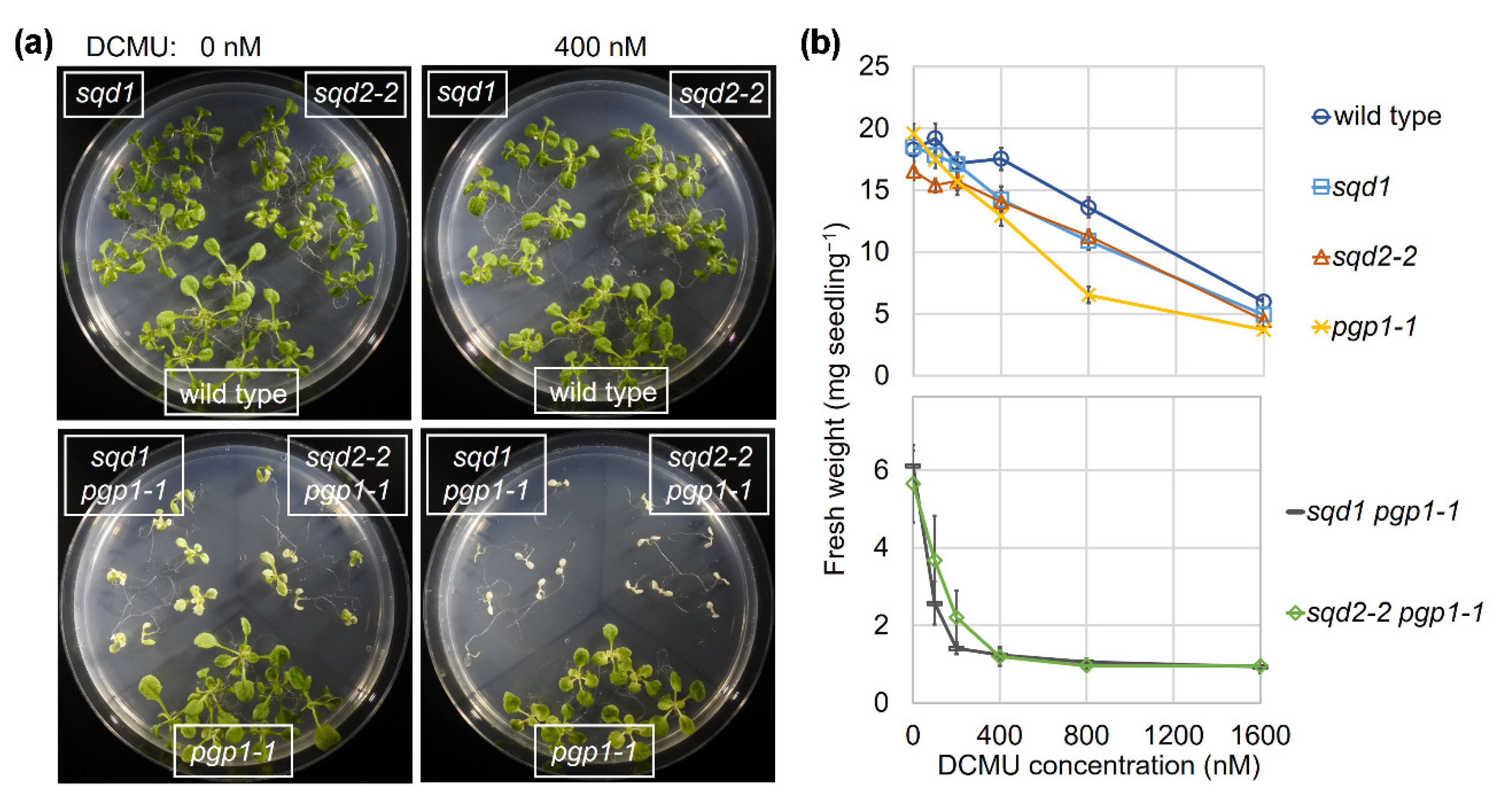
2.1. Loss of SQD1 and SQD2 in the pgp1-1 Background Causes the Same Growth Defects under P-sufficient Conditions
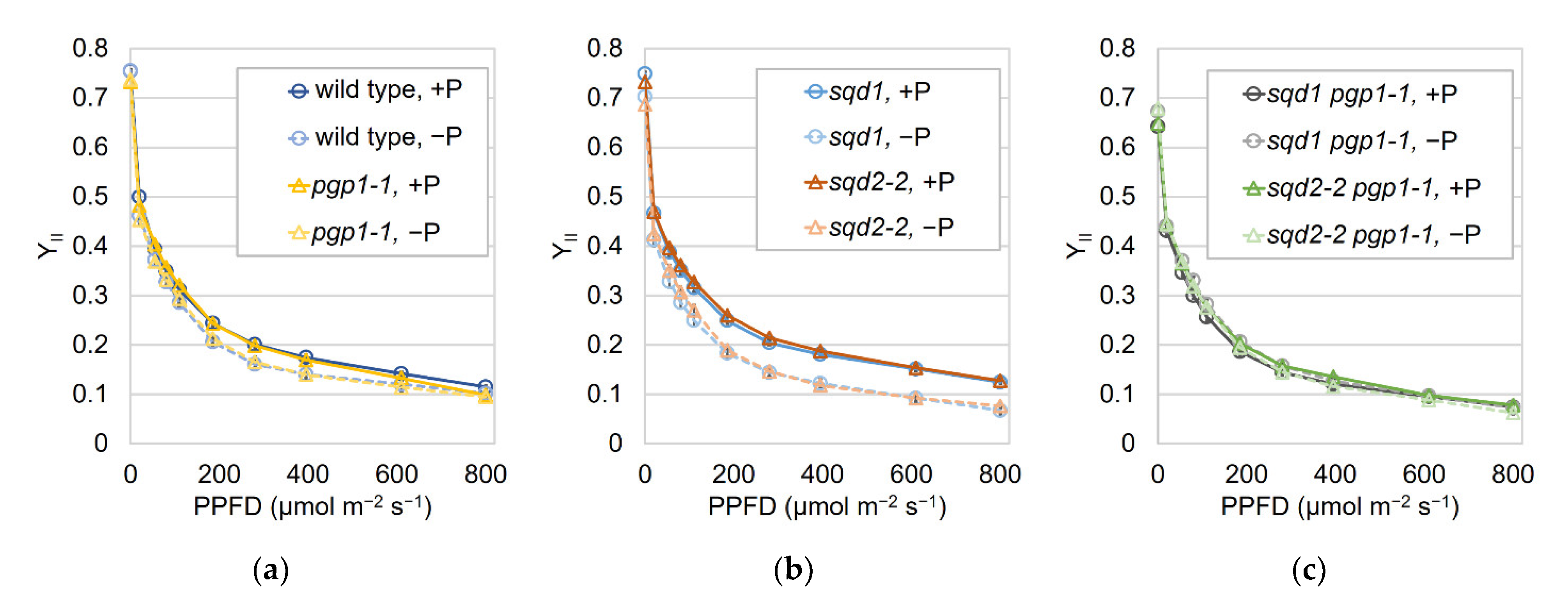
2.2. Loss of SQD1 and SQD2 Causes Similar Photosynthetic Defects
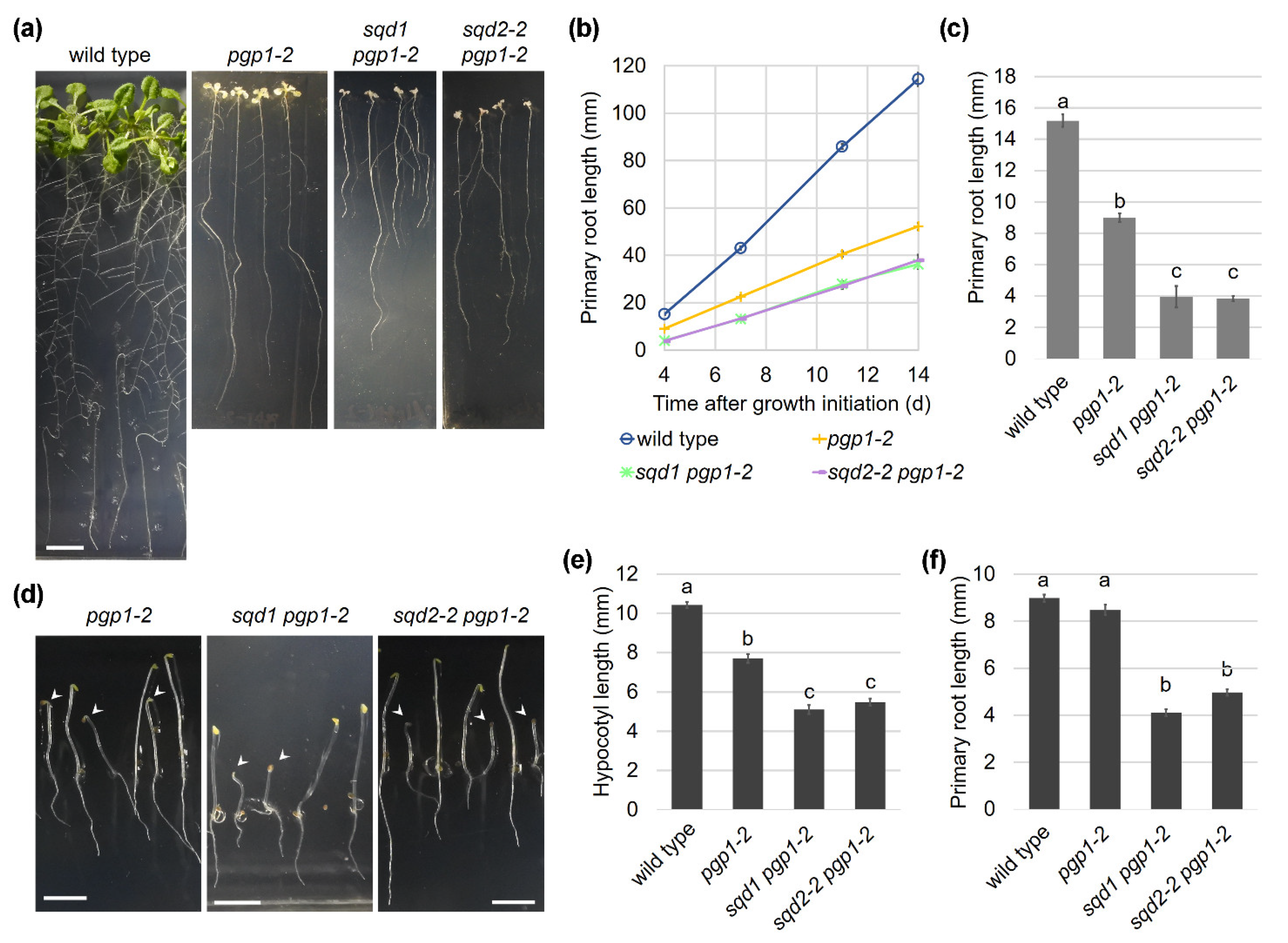
2.3. Complete Loss of Anionic Lipid Biosynthesis in Plastids Substantially Impairs Plant Development
2.4. The Photosystem II Function Is Completely Abolished by Loss of Anionic Lipid Biosynthesis in Plastids
2.5. Anionic Lipids Are Essential for Thylakoid Membrane Biogenesis
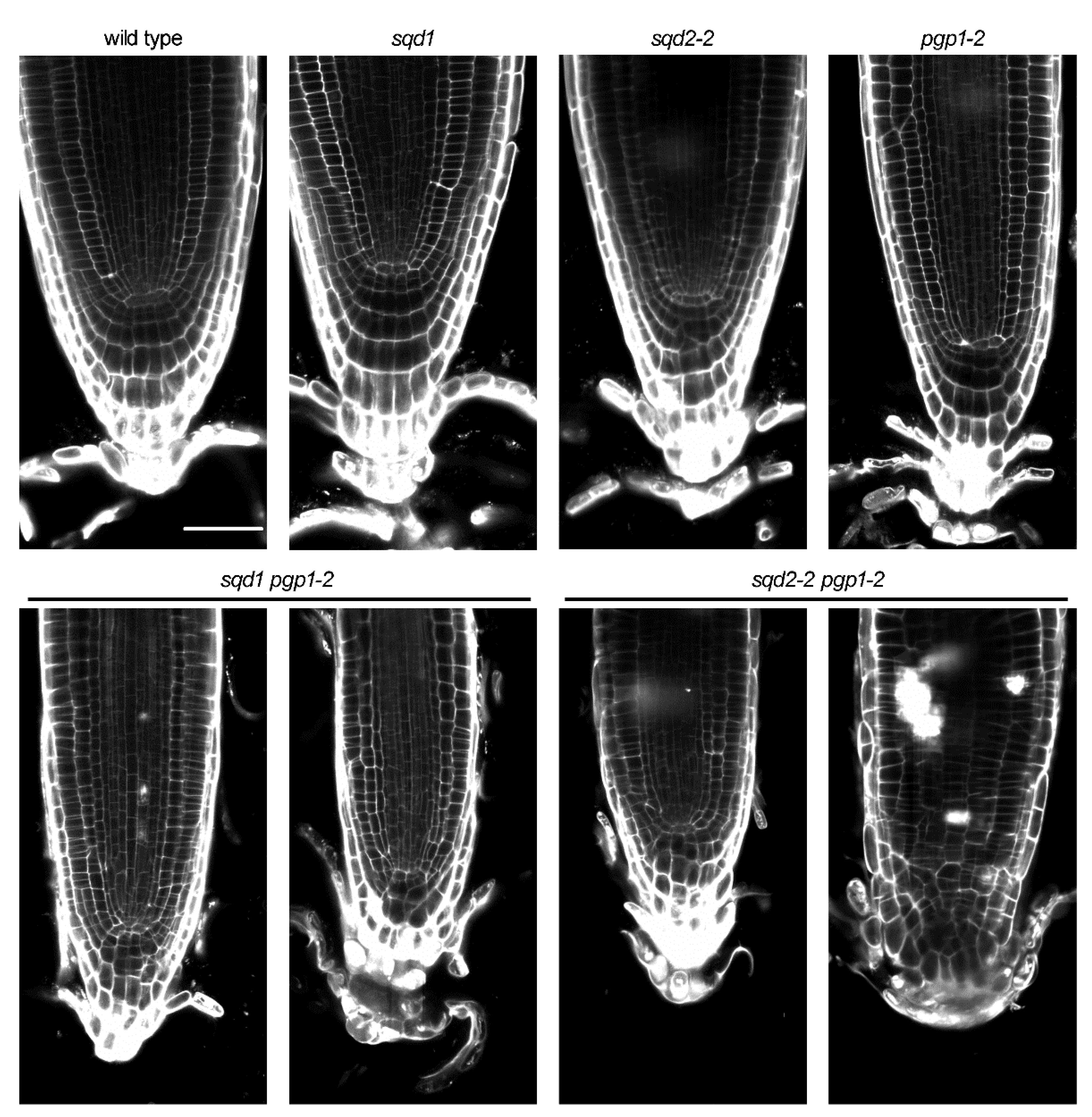
2.6. Root Growth Is Impaired in Anionic Lipid Mutants
2.7. Cellular Architecture of the Root Tip Was Disrupted by Loss of Anionic Lipids
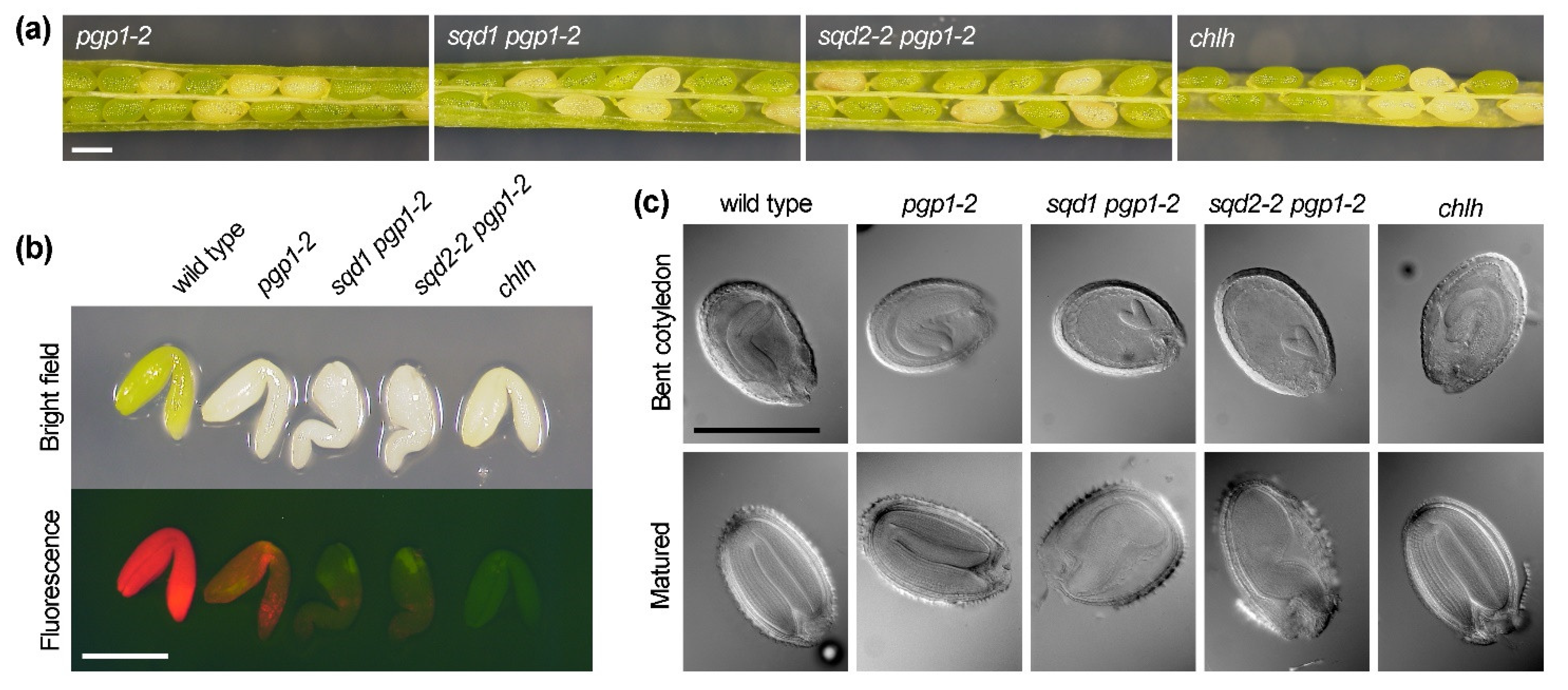
2.8. Loss of Plastid Anionic Lipids Impairs Embryo Development and Decreases Germination Capacity
3. Discussion
3.1. Role of SQDG in Complementing the Function of PG in Chloroplast Development and Photosynthesis
3.2. Role of Plastid Anionic Lipids in Root Growth and Embryo Development
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Measurement of FW and Chl Content
4.3. Analysis of Photosynthetic Efficiency
4.4. Transmission Electron Microscopic Analysis
4.5. Root Tip Observation
4.6. Embryo Observation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kobayashi, K. Role of membrane glycerolipids in photosynthesis, thylakoid biogenesis and chloroplast development. J. Plant Res. 2016, 129, 565–580. [Google Scholar] [CrossRef] [Green Version]
- Dorne, A.J.; Joyard, J.; Douce, R. Do thylakoids really contain phosphatidylcholine? Proc. Natl. Acad. Sci. USA 1990, 87, 71–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Endo, K.; Wada, H. Roles of lipids in photosynthesis. In Lipids in Plant and Algae Development. Subcellular Biochemistry; Nakamura, Y., Li-Beisson, Y., Eds.; Springer: New York, NY, USA, 2016; Volume 86, pp. 21–49. [Google Scholar]
- Heinz, S.; Rast, A.; Shao, L.; Gutu, A.; Gügel, I.L.; Heyno, E.; Labs, M.; Rengstl, B.; Viola, S.; Nowaczyk, M.M.; et al. Thylakoid membrane architecture in Synechocystis depends on CurT, a homolog of the granal CURVATURE THYLAKOID1 proteins. Plant Cell 2016, 28, 2238–2260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-C.C.; Kobayashi, K.; Wada, H.; Nakamura, Y. Phosphatidylglycerophosphate phosphatase is required for root growth in Arabidopsis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Yang, Z.; Kuang, T.; Physiology, E.M. Impaired photosynthesis in phosphatidylglycerol-deficient mutant of cyanobacterium Anabaena sp. PCC7120 with a disrupted gene encoding a putative phosphatidylglycerophosphatase. Plant Physiol. 2006, 141, 1274–1283. [Google Scholar] [CrossRef] [Green Version]
- Hagio, M.; Gombos, Z.; Várkonyi, Z.; Masamoto, K.; Sato, N.; Tsuzuki, M.; Wada, H. Direct evidence for requirement of phosphatidylglycerol in photosystem II of photosynthesis. Plant Physiol. 2000, 124, 795–804. [Google Scholar] [CrossRef] [Green Version]
- Bogos, B.; Ughy, B.; Domonkos, I.; Laczko-Dobos, H.; Komenda, J.; Abasova, L.; Cser, K.; Vass, I.; Sallai, A.; Wada, H.; et al. Phosphatidylglycerol depletion affects photosystem II activity in Synechococcus sp. PCC 7942 cells. Photosynth. Res. 2010, 103, 19–30. [Google Scholar] [CrossRef]
- Endo, K.; Kobayashi, K.; Wada, H. Sulfoquinovosyldiacylglycerol has an essential role in Thermosynechococcus elongatus BP-1 under phosphate-deficient conditions. Plant Cell Physiol. 2016, 57, pcw159. [Google Scholar] [CrossRef] [Green Version]
- Müller, F.; Frentzen, M. Phosphatidylglycerophosphate synthases from Arabidopsis thaliana. FEBS Lett. 2001, 509, 298–302. [Google Scholar] [CrossRef] [Green Version]
- Babiychuk, E.; Müller, F.; Eubel, H.; Braun, H.-P.; Frentzen, M.; Kushnir, S. Arabidopsis phosphatidylglycerophosphate synthase 1 is essential for chloroplast differentiation, but is dispensable for mitochondrial function. Plant J. 2003, 33, 899–909. [Google Scholar] [CrossRef]
- Tanoue, R.; Kobayashi, M.; Katayama, K.; Nagata, N.; Wada, H. Phosphatidylglycerol biosynthesis is required for the development of embryos and normal membrane structures of chloroplasts and mitochondria in Arabidopsis. FEBS Lett. 2014, 588, 1680–1685. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Härtel, H.; Wada, H.; Hagio, M.; Yu, B.; Eakin, C.; Benning, C. The pgp1 mutant locus of Arabidopsis encodes a phosphatidylglycerolphosphate synthase with impaired activity. Plant Physiol. 2002, 129, 594–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagio, M.; Sakurai, I.; Sato, S.; Kato, T.; Tabata, S.; Wada, H. Phosphatidylglycerol is essential for the development of thylakoid membranes in Arabidopsis thaliana. Plant Cell Physiol. 2002, 43, 1456–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Endo, K.; Wada, H. Multiple impacts of loss of plastidic phosphatidylglycerol biosynthesis on photosynthesis during seedling growth of Arabidopsis. Front. Plant Sci. 2016, 7, 336. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Fujii, S.; Sato, M.; Toyooka, K.; Wada, H. Specific role of phosphatidylglycerol and functional overlaps with other thylakoid lipids in Arabidopsis chloroplast biogenesis. Plant Cell Rep. 2015, 34, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Xu, C.; Benning, C. Arabidopsis disrupted in SQD2 encoding sulfolipid synthase is impaired in phosphate-limited growth. Proc. Natl. Acad. Sci. USA 2002, 99, 5732–5737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, Y.; Otsuki, H.; Narisawa, T.; Kobayashi, M.; Sawai, S.; Kamide, Y.; Kusano, M.; Aoki, T.; Hirai, M.Y.; Saito, K. A new class of plant lipid is essential for protection against phosphorus depletion. Nat. Commun. 2013, 4, 1510. [Google Scholar] [CrossRef] [Green Version]
- Aoki, M.; Sato, N.; Meguro, A.; Tsuzuki, M. Differing involvement of sulfoquinovosyl diacylglycerol in photosystem II in two species of unicellular cyanobacteria. Eur. J. Biochem. 2004, 271, 685–693. [Google Scholar] [CrossRef]
- Güler, S.; Seeliger, A.; Härtel, H.; Renger, G.; Benning, C. A null mutant of Synechococcus sp. PCC7942 deficient in the sulfolipid sulfoquinovosyl diacylglycerol. J. Biol. Chem. 1996, 271, 7501–7507. [Google Scholar] [CrossRef] [Green Version]
- Sato, N.; Tsuzuki, M.; Matsuda, Y.; Ehara, T.; Osafune, T.; Kawaguchi, A. Isolation and characterization of mutants affected in Lipid metabolism of Chlamydomonas reinhardtii. Eur. J. Biochem. 1995, 230, 987–993. [Google Scholar] [CrossRef]
- Riekhof, W.R.; Ruckle, M.E.; Lydic, T.A.; Sears, B.B.; Benning, C. The sulfolipids 2′-O-acyl-sulfoquinovosyldiacylglycerol and sulfoquinovosyldiacylglycerol are absent from a Chlamydomonas reinhardtii mutant deleted in SQD1. Plant Physiol. 2003, 133, 864–874. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Benning, C. Anionic lipids are required for chloroplast structure and function in Arabidopsis. Plant J. 2003, 36, 762–770. [Google Scholar] [CrossRef]
- Okazaki, Y.; Shimojima, M.; Sawada, Y.; Toyooka, K.; Narisawa, T.; Mochida, K.; Tanaka, H.; Matsuda, F.; Hirai, A.; Hirai, M.Y.; et al. A chloroplastic UDP-glucose pyrophosphorylase from Arabidopsis is the committed enzyme for the first step of sulfolipid biosynthesis. Plant Cell 2009, 21, 892–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Ohta, H. Possible requirement of galactolipids for embryogenesis. In Photosynthesis. Energy from the Sun; Allen, J.F., Gantt, E., Golbeck, J.H., Osmond, B., Eds.; Springer: Heidelberg, Germany, 2008; pp. 783–786. [Google Scholar]
- Huang, Y.-S.; Li, H.-M. Arabidopsis CHLI2 can substitute for CHLI1. Plant Physiol. 2009, 150, 636–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, K.; Mizusawa, N.; Shen, J.-R.; Yamada, M.; Tomo, T.; Komatsu, H.; Kobayashi, M.; Kobayashi, K.; Wada, H. Site-directed mutagenesis of amino acid residues of D1 protein interacting with phosphatidylglycerol affects the function of plastoquinone QB in photosystem II. Photosynth. Res. 2015, 126, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Endo, K.; Kobayashi, K.; Wang, H.T.; Chu, H.A.; Shen, J.R.; Wada, H. Site-directed mutagenesis of two amino acid residues in cytochrome b 559 α subunit that interact with a phosphatidylglycerol molecule (PG772) induces quinone-dependent inhibition of photosystem II activity. Photosynth. Res. 2019, 139, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Jordan, P.; Fromme, P.; Witt, H.T.; Klukas, O.; Saenger, W.; Krauss, N. Three-dimensional structure of cyanobacterial photosystem I at 2.5 Å resolution. Nature 2001, 411, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Mazor, Y.; Borovikova, A.; Caspy, I.; Nelson, N. Structure of the plant photosystem I supercomplex at 2.6 Å resolution. Nat. Plants 2017, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Umena, Y.; Kawakami, K.; Shen, J.-R.; Kamiya, N. Crystal structure of oxygen-evolving photosystem II at a resolution of 1.9 Å. Nature 2011, 473, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Su, X.; Cao, P.; Liu, X.; Chang, W.; Li, M.; Zhang, X.; Liu, Z. Structure of spinach photosystem II-LHCII supercomplex at 3.2 Å resolution. Nature 2016, 534, 69–74. [Google Scholar] [CrossRef]
- Nakajima, Y.; Umena, Y.; Nagao, R.; Endo, K.; Kobayashi, K.; Akita, F.; Suga, M.; Wada, H.; Noguchi, T.; Shen, J.R. Thylakoid membrane lipid sulfoquinovosyl-diacylglycerol (SQDG) is required for full functioning of photosystem II in Thermosynechococcus elongatus. J. Biol. Chem. 2018, 293, 14786–14797. [Google Scholar] [CrossRef] [Green Version]
- Sato, N.; Sonoike, K.; Tsuzuk, M.; Kawaguchi, A. Impaired photosystem II in a mutant of Chlamydomonas reinhardtii defective in sulfoquinovosyl diacylglycerol. Eur. J. Biochem. 1995, 234, 16–23. [Google Scholar] [CrossRef]
- Lin, Y.-C.C.; Kobayashi, K.; Hung, C.-H.H.; Wada, H.; Nakamura, Y. Arabidopsis phosphatidylglycerophosphate phosphatase 1 involved in phosphatidylglycerol biosynthesis and photosynthetic function. Plant J. 2016, 88, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Katayama, K.; Wada, H. T-DNA insertion in the CLS gene for cardiolipin synthase affects development of Arabidopsis thaliana. Cytologia (Tokyo) 2012, 77, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Jouhet, J.; Maréchal, E.; Baldan, B.; Bligny, R.; Joyard, J.; Block, M.A. Phosphate deprivation induces transfer of DGDG galactolipid from chloroplast to mitochondria. J. Cell Biol. 2004, 167, 863–874. [Google Scholar] [CrossRef]
- Kim, H.U.; Huang, A.H.C. Plastid lysophosphatidyl acyltransferase is essential for embryo development in Arabidopsis. Plant Physiol. 2004, 134, 1206–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazaz, S.; Barthole, G.; Domergue, F.; Ettaki, H.; To, A.; Vasselon, D.; de Vos, D.; Belcram, K.; Lepiniec, L.; Baud, S. Differential activation of partially redundant Δ9 stearoyl-ACP desaturase genes is critical for omega-9 monounsaturated fatty acid biosynthesis during seed development in arabidopsis. Plant Cell 2020, 32, 3613–3637. [Google Scholar] [CrossRef]
- Xu, C.; Fan, J.; Froehlich, J.E.; Awai, K.; Benning, C. Mutation of the TGD1 chloroplast envelope protein affects phosphatidate metabolism in Arabidopsis. Plant Cell 2005, 17, 3094–3110. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Kondo, M.; Fukuda, H.; Nishimura, M.; Ohta, H. Galactolipid synthesis in chloroplast inner envelope is essential for proper thylakoid biogenesis, photosynthesis, and embryogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 17216–17221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alban, C.; Joyard, J.; Douce, R. Preparation and characterization of envelope membranes from nongreen plastids. Plant Physiol. 1988, 88, 709–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Mutation | Lipidic Phenotype | Growth Phenotype | Reference |
|---|---|---|---|---|
| sqd1 | T-DNA insertion in the first exon of SQD1 | Complete lack of SQDG | Wild type-like phenotype under both P-sufficient and -starved conditions | [18] |
| sqd2-2 | T-DNA insertion in the first exon of SQD2 | Complete lack of SQDG and GlcADG | Wild type-like phenotype under a P-sufficient condition and growth defects under a P-starved condition | [17,18] |
| pgp1-1 | Single amino acid substitution (P170S) of PGP1 | 80% reduction of PGP1 activity and 30% reduction of PG content | Slight decrease in chlorophyll content and phtoosynthetic activity | [13] |
| sqd1 pgp1-1 | Double mutations of sqd1 and pgp1-1 | Complete lack of SQDG and decreased PGP1 activity | Stronger defects in growth, chlorophyll accumulation, and photosynthetic activity than pgp1-1 | This study (Figure 1, Figure 2 and Figure 3) |
| sqd2-2 pgp1-1 | Double mutations of sqd2-2 and pgp1-1 | Complete lack of SQDG and GlcADG and decreased PGP1 activity | Almost the same as sqd1 pgp1-1 in growth and photosynthetic characteristics | This study (Figure 1, Figure 2 and Figure 3) |
| pgp1-2 | T-DNA insertion in the first exon of PGP1 with a start codon deletion | 88% reduction of PG content by loss of PG biosynthesis in plastids | Seedling lethal phenotype with severe impairments of chlorophyll accumulation, thyalkoid formation, and photosynthesis | [14,15,16] |
| sqd1 pgp1-2 | Double mutations of sqd1 and pgp1-2 | Complete lack of SQDG and loss of PG biosynthesis in plastids | Stronger defects in growth, chlorophyll accumulation, thylakoid formation, and photosynthesis than pgp1-2 | This study (Figure 4, Figure 5, Figure 6, Figure 7, Figure 8 and Figure 9) |
| sqd2-2 pgp1-2 | Double mutations of sqd2-2 and pgp1-2 | Complete lack of SQDG and GlcADG and loss of PG biosynthesis in plastids | Almost the same as sqd1 pgp1-2 in growth and photosynthetic characteristics | This study (Figure 4, Figure 5, Figure 6, Figure 7, Figure 8 and Figure 9) |
| Phenotype | Wild Type | sqd1 | sqd2-2 | pgp1-2 | sqd1 pgp1-2 | sqd2-2 pgp1-2 |
|---|---|---|---|---|---|---|
| albino seed | 0.0% | 0.0% | 0.0% | 25.0% | 24.9% | 24.8% |
| (0/562) | (0/669) | (0/626) | (199/797) | (237/716) | (274/1105) | |
| ungerminated seed | 2.9% | 5.3% | 2.5% | 7.9% | 21.1% | 16.1% |
| (8/280) | (17/320) | (7/280) | (110/1370) | (304/1400) | (229/1391) | |
| albino seedling | 0.0% | 0.0% | 0.0% | 18.4% | 10.2% | 12.0% |
| (0/280) | (0/320) | (0/280) | (250/1370) | (133/1400) | (166/1391) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshihara, A.; Nagata, N.; Wada, H.; Kobayashi, K. Plastid Anionic Lipids Are Essential for the Development of Both Photosynthetic and Non-Photosynthetic Organs in Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 4860. https://doi.org/10.3390/ijms22094860
Yoshihara A, Nagata N, Wada H, Kobayashi K. Plastid Anionic Lipids Are Essential for the Development of Both Photosynthetic and Non-Photosynthetic Organs in Arabidopsis thaliana. International Journal of Molecular Sciences. 2021; 22(9):4860. https://doi.org/10.3390/ijms22094860
Chicago/Turabian StyleYoshihara, Akiko, Noriko Nagata, Hajime Wada, and Koichi Kobayashi. 2021. "Plastid Anionic Lipids Are Essential for the Development of Both Photosynthetic and Non-Photosynthetic Organs in Arabidopsis thaliana" International Journal of Molecular Sciences 22, no. 9: 4860. https://doi.org/10.3390/ijms22094860