
SNX-PXA-RGS-PXC Subfamily of SNXs in the Regulation of Receptor-Mediated Signaling and Membrane Trafficking

 , ,
, ,
Abstract
:1. Introduction
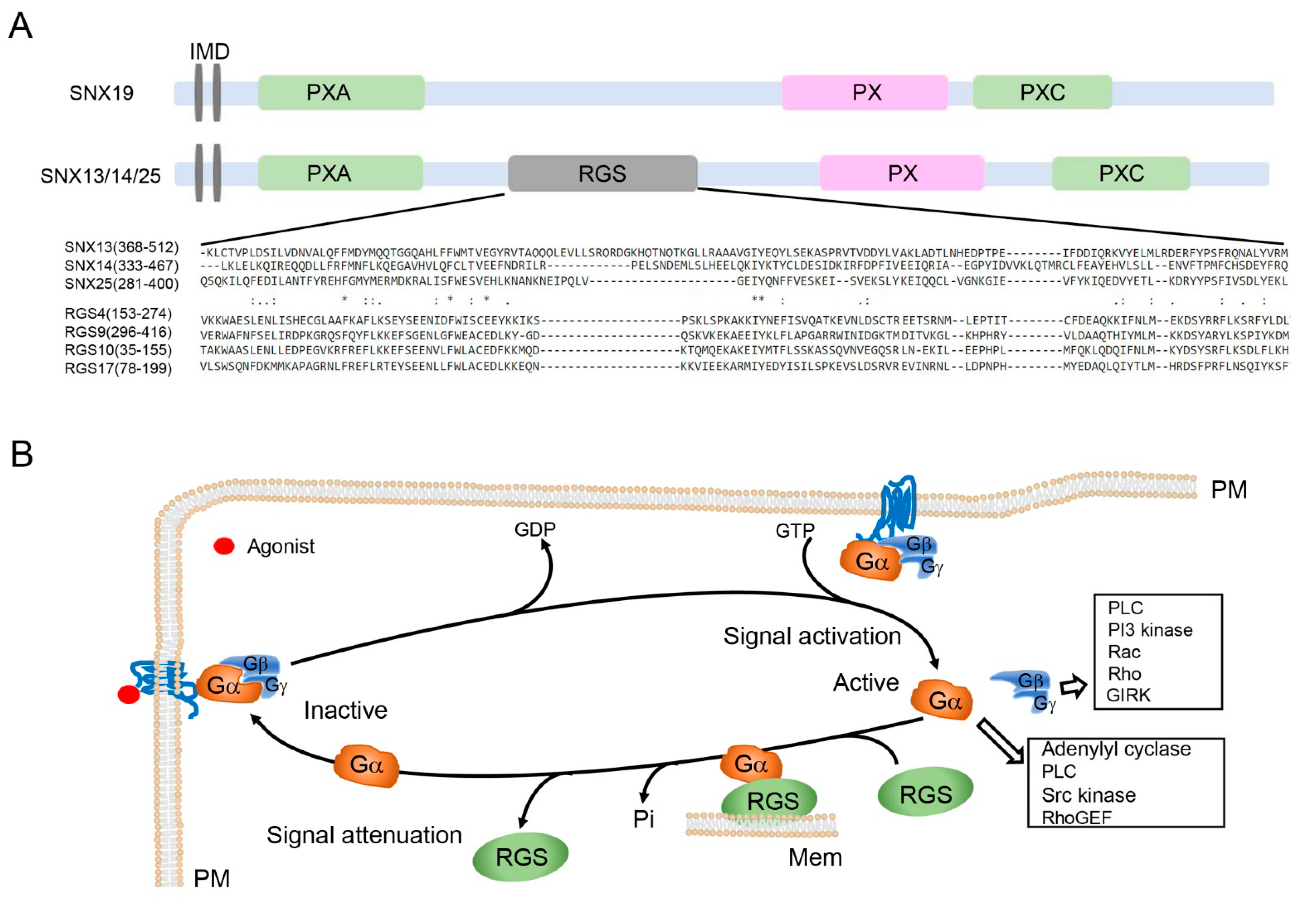
2. SNX-PXA-RGS-PXC Subfamily Domain Structure and Biochemical Properties
3. SNX-PXA-RGS-PXC Subfamily in Receptor Signaling
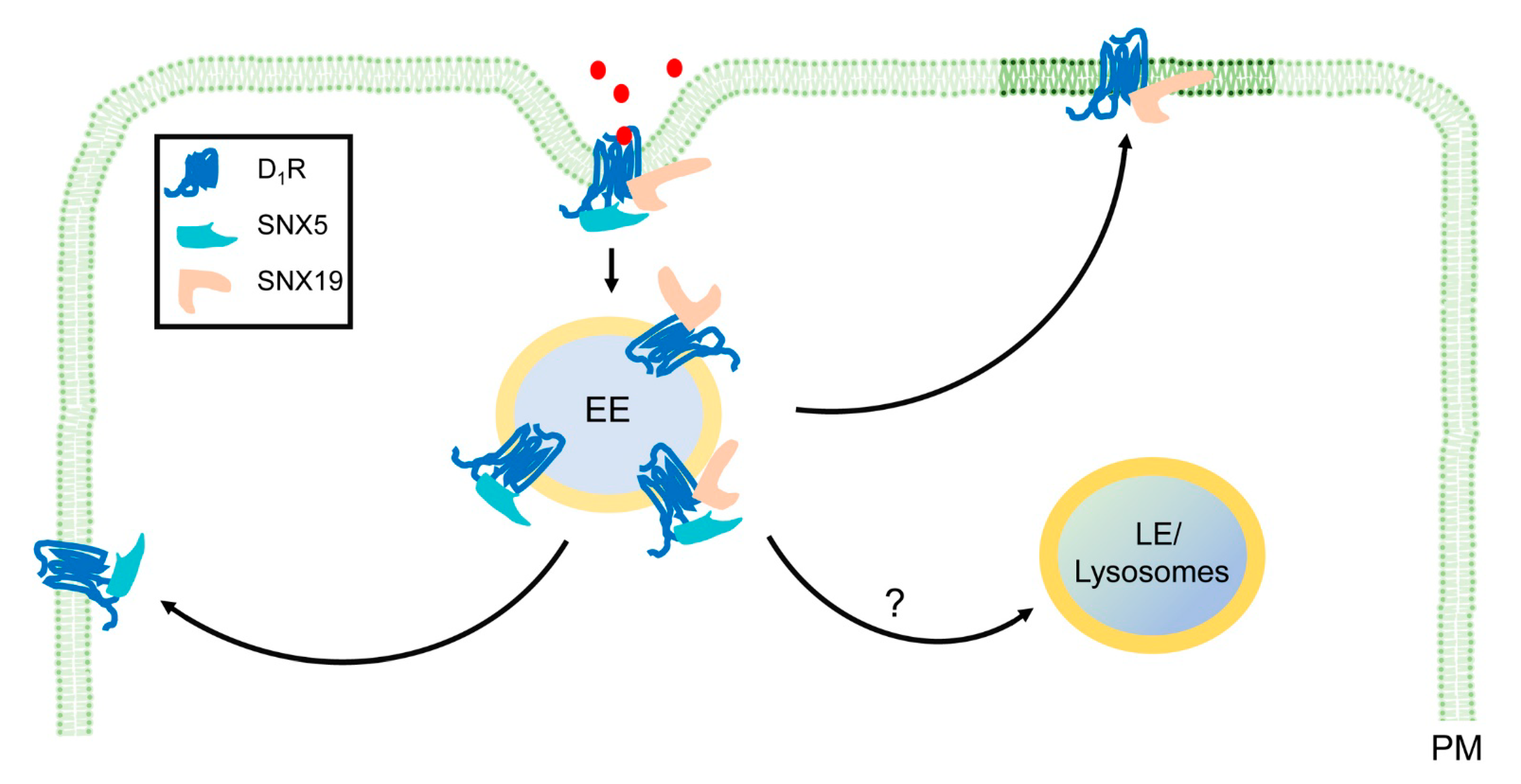
4. SNX-PXA-RGS-PXC Subfamily in Membrane Trafficking
4.1. SNX-PXA-RGS-PXC Subfamily in Lysosomal-Mediated Degradation
4.2. SNX-PXA-RGS-PXC Subfamily in Membrane Recycling
5. Comparison of SNX-PXA-RGS-PXC Subfamily with SNX-BAR Subfamily in Receptor Signaling and Membrane Trafficking
6. Comparison of SNX-PXA-RGS-PXC Subfamily with Other SNX Subfamilies
7. SNX-PXA-RGS-PXC Subfamily in Physiology and Pathophysiology
8. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Akt/PKB | protein kinase B |
| AR | adrenergic receptor |
| ARC | apoptosis receptor with caspase recruitment domain |
| BACE 1 | β-site amyloid precursor protein-cleaving enzyme 1 |
| BAR | Bin-Amphiphysin-Rvs |
| cAMP/PKA | cyclic adenosine monophosphate/protein kinase A |
| CED-1 | cell death abnormality protein-1 |
| CI-MPR | cation-independent mannose 6-phosphate receptor |
| CNS | central nervous system |
| DCV | dense core vesicles |
| dHMN | distal hereditary motor neuropathies |
| DMT1-II | divalent metal transporter 1-II |
| D1R | dopamine receptor 1 |
| D2R | dopamine receptor 2 |
| EGFR | epithelial growth factor receptor |
| EOAD | early-onset Alzheimer’s Disease |
| ER | endoplasmic reticulum |
| ESCPE-1 | endosomal SNX-BAR sorting complex for promoting exit-1 |
| ESCRT | endosomal sorting complex required for transport proteins |
| FERM | protein 4.1/ezrin/radixin/ moesin |
| GAP | GTPase-activating proteins |
| Gαs | G protein alpha stimulatory subunit |
| GPCR | G protein-coupled receptor |
| Hrs | hepatocyte growth factor-regulated tyrosine kinase substrate |
| 5HT6R | serotonin receptor 6 |
| IMD | integrated transmembrane domain |
| LC3 | microtubule-associated proteins 1A/1B light chain 3B |
| LD | lipid droplet |
| LOAD | late-onset Alzheimer’s Disease |
| MCS | membrane contact sites |
| Mdm1 | mitochondrial distribution and morphology 1 |
| NHE3 | sodium hydrogen exchanger 3 |
| PI | phosphoinositide |
| PI(3)P | phosphatidylinositol 3-phosphate |
| PLC | phospholipase C |
| PM | plasma membrane |
| PX | Phox-homology domain |
| PXA | PX-associated domain |
| PXC | C-terminal PX-associated domain |
| RGS | regulators of G protein signaling |
| GEF | guanine nucleotide exchange factor |
| SCAR20 | autosomal recessive spinocerebellar ataxia 20 |
| SXN | sorting nexin |
| TGF | transforming growth factor |
| TGN | trans-Golgi network |
| TrkB | tropomyosin receptor kinase B |
| U2OS | human bone osteosarcoma epithelial cells |
| WASH | Wiskott Aldrich Syndrome protein and scar homologue |
References
- Lobingier, B.T.; von Zastrow, M. When trafficking and signaling mix: How subcellular location shapes G protein-coupled receptor activation of heterotrimeric G proteins. Traffic 2019, 20, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Fedoseienko, A.; Chen, B.; Burstein, E.; Jia, D.; Billadeau, D.D. Endosomal receptor trafficking: Retromer and beyond. Traffic 2018, 19, 578–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naslavsky, N.; Caplan, S. The enigmatic endosome-sorting the ins and outs of endocytic trafficking. J. Cell Sci. 2018, 131, jcs216499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, M.; Collins, B.M. The phox homology (PX) domain. Adv. Exp. Med. Biol. 2019, 1111, 1–17. [Google Scholar] [PubMed]
- Cullen, P.J.; Korswagen, H.C. Sorting nexins provide diversity for retromer-dependent trafficking events. Nat. Cell Biol. 2012, 14, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Teasdale, R.D.; Collins, B.M. Insights into the PX (phox-homology) domain and SNX (sorting nexin) protein families: Structures, functions and roles in disease. Biochem. J. 2012, 441, 39–59. [Google Scholar] [CrossRef]
- Gallon, M.; Cullen, P.J. Retromer and sorting nexins in endosomal sorting. Biochem. Soc. Trans. 2015, 43, 33–47. [Google Scholar] [CrossRef]
- Yang, J.; Villar, V.A.M.; Rozyyev, S.; Jose, P.A.; Zeng, C. The emerging role of sorting nexins in cardiovascular diseases. Clin. Sci. 2019, 133, 723–737. [Google Scholar] [CrossRef] [Green Version]
- Falasca, M.; Maffucci, T. Rethinking phosphatidylinositol 3-monophosphate. Biochim. Biophys. Acta 2009, 1793, 1795–1803. [Google Scholar] [CrossRef] [Green Version]
- Mas, C.; Norwood, S.J.; Bugarcic, A.; Kinna, G.; Leneva, N.; Kovtun, O.; Ghai, R.; Ona Yanez, L.E.; Davis, J.L.; Teasdale, R.D.; et al. Structural basis for different phosphoinositide specificities of the PX domains of sorting nexins regulating G-protein signaling. J. Biol. Chem. 2014, 289, 28554–28568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, M.; Chin, Y.K.; Mas, C.; Feathers, J.R.; Paul, B.; Datta, S.; Chen, K.E.; Jia, X.; Yang, Z.; Norwood, S.J.; et al. Classification of the human phox homology (PX) domains based on their phosphoinositide binding specificities. Nat. Commun. 2019, 10, 1528. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Zhang, D.; Shi, D.; Qi, M.; Feng, J.; Yuan, T.; Xu, X.; Liang, D.; Xu, L.; et al. SNX13 reduction mediates heart failure through degradative sorting of apoptosis repressor with caspase recruitment domain. Nat. Commun. 2014, 5, 5177. [Google Scholar] [CrossRef] [Green Version]
- Ha, C.M.; Park, D.; Kim, Y.; Na, M.; Panda, S.; Won, S.; Kim, H.; Ryu, H.; Park, Z.Y.; Rasenick, M.M.; et al. SNX14 is a bifunctional negative regulator for neuronal 5-HT6 receptor signaling. J. Cell Sci. 2015, 128, 1848–1861. [Google Scholar] [CrossRef] [Green Version]
- Bryant, D.; Liu, Y.; Datta, S.; Hariri, H.; Seda, M.; Anderson, G.; Peskett, E.; Demetriou, C.; Sousa, S.; Jenkins, D.; et al. SNX14 mutations affect endoplasmic reticulum-associated neutral lipid metabolism in autosomal recessive spinocerebellar ataxia 20. Hum. Mol. Genet. 2018, 27, 1927–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, S.; Liu, Y.; Hariri, H.; Bowerman, J.; Henne, W.M. Cerebellar ataxia disease-associated Snx14 promotes lipid droplet growth at ER-droplet contacts. J. Cell Biol. 2019, 218, 1335–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akizu, N.; Cantagrel, V.; Zaki, M.S.; Al-Gazali, L.; Wang, X.; Rosti, R.O.; Dikoglu, E.; Gelot, A.B.; Rosti, B.; Vaux, K.K.; et al. Biallelic mutations in SNX14 cause a syndromic form of cerebellar atrophy and lysosome-autophagosome dysfunction. Nat. Genet. 2015, 47, 528–534. [Google Scholar] [CrossRef]
- Thomas, A.C.; Williams, H.; Setó-Salvia, N.; Bacchelli, C.; Jenkins, D.; O’Sullivan, M.; Mengrelis, K.; Ishida, M.; Ocaka, L.; Chanudet, E.; et al. Mutations in SNX14 cause a distinctive autosomal-recessive cerebellar ataxia and intellectual disability syndrome. Am. J. Hum. Genet. 2014, 95, 611–621. [Google Scholar] [CrossRef] [Green Version]
- Bare, L.A.; Morrison, A.C.; Rowland, C.M.; Shiffman, D.; Luke, M.M.; Iakoubova, O.A.; Kane, J.P.; Malloy, M.J.; Ellis, S.G.; Pankow, J.S.; et al. Five common gene variants identify elevated genetic risk for coronary heart disease. Genet. Med. 2007, 9, 682–689. [Google Scholar] [CrossRef] [Green Version]
- Fullard, J.F.; Giambartolomei, C.; Hauberg, M.E.; Xu, K.; Voloudakis, G.; Shao, Z.; Bare, C.; Dudley, J.T.; Mattheisen, M.; Robakis, N.K.; et al. Open chromatin profiling of human postmortem brain infers functional roles for non-coding schizophrenia loci. Hum. Mol. Genet. 2017, 26, 1942–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiu, A.C.; Yang, J.; Asico, L.D.; Konkalmatt, P.; Zheng, X.; Cuevas, S.; Wang, X.; Lee, H.; Mazhar, M.; Felder, R.A.; et al. Lipid rafts are required for effective renal D1 dopamine receptor function. FASEB J. 2020, 34, 6999–7017. [Google Scholar] [CrossRef] [Green Version]
- Kan, A.; Ikeda, T.; Saito, T.; Yano, F.; Fukai, A.; Hojo, H.; Ogasawara, T.; Ogata, N.; Nakamura, K.; Chung, U.I.; et al. Screening of chondrogenic factors with a real-time fluorescence-monitoring cell line ATDC5-C2ER: Identification of sorting nexin 19 as a novel factor. Arthritis Rheum. 2009, 60, 3314–3323. [Google Scholar] [CrossRef] [PubMed]
- Harashima, S.I.; Harashima, C.; Nishimura, T.; Hu, Y.; Notkins, A.L. Overexpression of the autoantigen IA-2 puts beta cells into a pre-apoptotic state: Autoantigen-induced, but non-autoimmune-mediated, tissue destruction. Clin. Exp. Immunol. 2007, 150, 49–60. [Google Scholar] [CrossRef]
- Hao, X.; Wang, Y.; Ren, F.; Zhu, S.; Ren, Y.; Jia, B.; Li, Y.P.; Shi, Y.; Chang, Z. SNX25 regulates TGF-β signaling by enhancing the receptor degradation. Cell Signal. 2011, 23, 935–946. [Google Scholar] [CrossRef]
- Takemura, S.; Nagano, M.; Isonishi, A.; Tanaka, T.; Tatsumi, K.; Yamano, M.; Minami, Y.; Shigeyoshi, Y.; Wanaka, A. Circadian rhythms of sorting nexin 25 in the mouse suprachiasmatic nucleus. Neurosci. Lett. 2020, 727, 134897. [Google Scholar] [CrossRef]
- Du, Y.; Zou, Y.; Yu, W.; Shi, R.; Zhang, M.; Yang, W.; Duan, J.; Deng, Y.; Wang, X.; Lü, Y. Expression pattern of sorting Nexin 25 in temporal lobe epilepsy: A study on patients and pilocarpine-induced rats. Brain Res. 2013, 1509, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Cheng, R.; Vardarajan, B.; Lantigua, R.; Reyes-Dumeyer, D.; Ortmann, W.; Graham, R.R.; Bhangale, T.; Behrens, T.W.; Medrano, M.; et al. Genetic modifiers of age at onset in carriers of the G206A mutation in PSEN1 with Familial Alzheimer Disease among Caribbean Hispanics. JAMA Neurol. 2015, 72, 1043–1051. [Google Scholar] [CrossRef] [Green Version]
- Free, R.B.; Namkung, Y.; Hazelwood, L.A.; Sibley, D.R. Sorting nexin-25 interacts with D1 and D2 dopamine receptors to regulate receptor expression and signaling. FASEB J. 2010, 24, 771–778. [Google Scholar]
- Tesmer, J.J. Structure and function of regulator of G protein signaling homology domains. Prog. Mol. Biol. Transl. Sci. 2009, 86, 75–113. [Google Scholar]
- Squires, K.E.; Montañez-Miranda, C.; Pandya, R.R.; Torres, M.P.; Hepler, J.R. Genetic analysis of rare human variants of regulators of G protein signaling proteins and their role in human physiology and disease. Pharmacol. Rev. 2018, 70, 446–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willars, G.B. Mammalian RGS proteins: Multifunctional regulators of cellular signalling. Semin. Cell Dev. Biol. 2006, 17, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.; Fisher, R.A. Introduction: G protein-coupled receptors and RGS proteins. Prog. Mol. Biol. Transl. Sci. 2015, 133, 1–11. [Google Scholar] [PubMed]
- Worby, C.A.; Dixon, J.E. Sorting out the cellular functions of sorting nexins. Nat. Rev. Mol. Cell Biol. 2002, 3, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Ma, Y.C.; Ostrom, R.S.; Lavoie, C.; Gill, G.N.; Insel, P.A.; Huang, X.Y.; Farquhar, M.G. RGS-PX1, a GAP for Gαs and sorting nexin in vesicular trafficking. Science 2001, 294, 1939–1942. [Google Scholar] [CrossRef]
- Ross, E.M.; Wilkie, T.M. GTPase-activating proteins for heterotrimeric G proteins: Regulators of G protein signaling (RGS) and RGS-like proteins. Annu. Rev. Biochem. 2000, 69, 795–827. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Lavoie, C.; Tang, T.D.; Ma, P.; Meerloo, T.; Beas, A.; Farquhar, M.G. Regulation of epidermal growth factor receptor degradation by heterotrimeric Gαs protein. Mol. Biol. Cell 2004, 15, 5538–5550. [Google Scholar] [CrossRef] [Green Version]
- Su, K.; Xu, T.; Yu, Z.; Zhu, J.; Zhang, Y.; Wu, M.; Xiong, Y.; Liu, J.; Xu, J. Structure of the PX domain of SNX25 reveals a novel phospholipid recognition model by dimerization in the PX domain. FEBS Lett. 2017, 591, 2011–2018. [Google Scholar] [CrossRef] [Green Version]
- Rao, F.; Wessel, J.; Wen, G.; Zhang, L.; Rana, B.K.; Kennedy, B.P.; Greenwood, T.A.; Salem, R.M.; Chen, Y.; Khandrika, S.; et al. Renal albumin excretion: Twin studies identify influences of heredity, environment, and adrenergic pathway polymorphism. Hypertension 2007, 49, 1015–1031. [Google Scholar] [CrossRef] [PubMed]
- Bein, K.; Ganguly, K.; Martin, T.M.; Concel, V.J.; Brant, K.A.; Di, Y.P.; Upadhyay, S.; Fabisiak, J.P.; Vuga, L.J.; Kaminski, N.; et al. Genetic determinants of ammonia-induced acute lung injury in mice. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Irannejad, R.; Tsvetanova, N.G.; Lobingier, B.T.; von Zastrow, M. Effects of endocytosis on receptor-mediated signaling. Curr. Opin. Cell Biol. 2015, 35, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Ballabio, A.; Bonifacino, J.S. Lysosomes as dynamic regulators of cell and organismal homeostasis. Nat. Rev. Mol. Cell Biol. 2020, 21, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Ebner, M.; Koch, P.A.; Haucke, V. Phosphoinositides in the control of lysosome function and homeostasis. Biochem. Soc. Trans. 2019, 47, 1173–1185. [Google Scholar] [CrossRef] [PubMed]
- Henne, W.M.; Zhu, L.; Balogi, Z.; Stefan, C.; Pleiss, J.A.; Emr, S.D. Mdm1/Snx13 is a novel ER-endolysosomal interorganelle tethering protein. J. Cell Biol. 2015, 210, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Bache, K.G.; Brech, A.; Mehlum, A.; Stenmark, H. Hrs regulates multivesicular body formation via ESCRT recruitment to endosomes. J. Cell Biol. 2003, 162, 435–442. [Google Scholar] [CrossRef] [Green Version]
- Klumperman, J.; Raposo, G. The complex ultrastructure of the endolysosomal system. Cold Spring Harb. Perspect Biol. 2014, 6, a016857. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Tang, T.; Tang, N.; Kudlicka, K.; Ohtsubo, K.; Ma, P.; Marth, J.D.; Farquhar, M.G.; Lehtonen, E. Essential role of RGS-PX1/sorting nexin 13 in mouse development and regulation of endocytosis dynamics. Proc. Natl. Acad. Sci. USA 2006, 103, 16776–16781. [Google Scholar] [CrossRef] [Green Version]
- Hariri, H.; Speer, N.; Bowerman, J.; Rogers, S.; Fu, G.; Reetz, E.; Datta, S.; Feathers, J.R.; Ugrankar, R.; Nicastro, D.; et al. Mdm1 maintains endoplasmic reticulum homeostasis by spatially regulating lipid droplet biogenesis. J. Cell Biol. 2019, 218, 1319–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harashima, S.; Horiuchi, T.; Wang, Y.; Notkins, A.L.; Seino, Y.; Inagaki, N. Sorting nexin 19 regulates the number of dense core vesicles in pancreatic β-cells. J. Diabetes Investig. 2012, 3, 52–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemura, S.; Isonishi, A.; Tanaka, T.; Okuda, H.; Tatsumi, K.; Yamano, M.; Wanaka, A. Neural expression of sorting nexin 25 and its regulation of tyrosine receptor kinase B trafficking. Brain Struct. Funct. 2020, 225, 2615–2642. [Google Scholar] [CrossRef]
- Weeratunga, S.; Paul, B.; Collins, B.M. Recognising the signals for endosomal trafficking. Curr. Opin. Cell Biol. 2020, 65, 17–27. [Google Scholar] [CrossRef]
- Villar, V.A.; Jones, J.E.; Armando, I.; Asico, L.D.; Escano, C.S., Jr.; Lee, H.; Wang, X.; Yang, Y.; Pascua-Crusan, A.M.; Palmes-Saloma, C.P.; et al. Sorting nexin 1 loss results in D5 dopamine receptor dysfunction in human renal proximal tubule cells and hypertension in mice. J. Biol. Chem. 2013, 288, 152–163. [Google Scholar] [CrossRef] [Green Version]
- Villar, V.A.; Armando, I.; Sanada, H.; Frazer, L.C.; Russo, C.M.; Notario, P.M.; Lee, H.; Comisky, L.; Russell, H.A.; Yang, Y.; et al. Novel role of sorting nexin 5 in renal D(1) dopamine receptor trafficking and function: Implications for hypertension. FASEB J. 2013, 27, 1808–1819. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Yang, J.; Jones, J.E.; Villar, V.A.; Yu, P.; Armando, I.; Felder, R.A.; Jose, P.A. Sorting nexin 5 and dopamine D1 receptor regulate the expression of the insulin receptor in human renal proximal tubule cells. Endocrinology 2015, 156, 2211–2221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Yang, J.; Villar, V.A.M.; Asico, L.D.; Ma, X.; Armando, I.; Sanada, H.; Yoneda, M.; Felder, R.A.; Jose, P.A.; et al. Loss of renal SNX5 results in impaired IDE activity and insulin resistance in mice. Diabetologia 2018, 61, 727–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, R.; Kametaka, S.; Haft, C.R.; Bonifacino, J.S. Interchangeable but essential functions of SNX1 and SNX2 in the association of retromer with endosomes and the trafficking of mannose 6-phosphate receptors. Mol. Cell Biol. 2007, 27, 1112–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellado, M.; Cuartero, Y.; Brugada, R.; Verges, M. Subcellular localisation of retromer in post-endocytic pathways of polarised Madin-Darby canine kidney cells. Biol. Cell 2014, 106, 377–393. [Google Scholar] [CrossRef]
- Chen, D.; Xiao, H.; Zhang, K.; Wang, B.; Gao, Z.; Jian, Y.; Qi, X.; Sun, J.; Miao, L.; Yang, C. Retromer is required for apoptotic cell clearance by phagocytic receptor recycling. Science 2010, 327, 1261–1264. [Google Scholar] [CrossRef]
- Lieu, Z.Z.; Gleeson, P.A. Identification of different itineraries and retromer components for endosome-to-Golgi transport of TGN38 and Shiga toxin. Eur. J. Cell Biol. 2010, 89, 379–393. [Google Scholar] [CrossRef]
- Robinson, D.G.; Neuhaus, J.M. Receptor-mediated sorting of soluble vacuolar proteins: Myths, facts, and a new model. J. Exp. Bot. 2016, 67, 4435–4449. [Google Scholar] [CrossRef] [Green Version]
- Okada, H.; Zhang, W.; Peterhoff, C.; Hwang, J.C.; Nixon, R.A.; Ryu, S.H.; Kim, T.W. Proteomic identification of sorting nexin 6 as a negative regulator of BACE1-mediated APP processing. FASEB J. 2010, 24, 2783–2794. [Google Scholar] [CrossRef] [Green Version]
- Jaillais, Y.; Santambrogio, M.; Rozier, F.; Fobis-Loisy, I.; Miege, C.; Gaude, T. The retromer protein VPS29 links cell polarity and organ initiation in plants. Cell 2007, 130, 1057–1070. [Google Scholar] [CrossRef] [Green Version]
- Traer, C.J.; Rutherford, A.C.; Palmer, K.J.; Wassmer, T.; Oakley, J.; Attar, N.; Carlton, J.G.; Kremerskothen, J.; Stephens, D.J.; Cullen, P.J. SNX4 coordinates endosomal sorting of TfnR with dynein-mediated transport into the endocytic recycling compartment. Nat. Cell Biol. 2007, 9, 1370–1380. [Google Scholar] [CrossRef]
- Kim, N.Y.; Cho, M.H.; Won, S.H.; Kang, H.J.; Yoon, S.Y.; Kim, D.H. Sorting nexin-4 regulates β-amyloid production by modulating β-site-activating cleavage enzyme-1. Alzheimers Res. Ther. 2017, 9, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solis, G.P.; Hulsbusch, N.; Radon, Y.; Katanaev, V.L.; Plattner, H.; Stuermer, C.A. Reggies/flotillins interact with Rab11a and SNX4 at the tubulovesicular recycling compartment and function in transferrin receptor and E-cadherin trafficking. Mol. Biol. Cell 2013, 24, 2689–2702. [Google Scholar] [CrossRef] [PubMed]
- Knaevelsrud, H.; Carlsson, S.R.; Simonsen, A. SNX18 tubulates recycling endosomes for autophagosome biogenesis. Autophagy 2013, 9, 1639–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Weering, J.R.; Verkade, P.; Cullen, P.J. SNX-BAR proteins in phosphoinositide-mediated, tubular-based endosomal sorting. Semin. Cell Dev. Biol. 2010, 21, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, H.F.; Felder, R.A.; Periasamy, A.; Jose, P.A. Rab4 and Rab11 coordinately regulate the recycling of angiotensin II type I receptor as demonstrated by fluorescence resonance energy transfer microscopy. J. Biomed. Opt. 2008, 13, 031206. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yu, P.; Sun, Y.; Felder, R.A.; Periasamy, A.; Jose, P.A. Actin cytoskeleton-dependent Rab GTPase-regulated angiotensin type I receptor lysosomal degradation studied by fluorescence lifetime imaging microscopy. J. Biomed. Opt. 2010, 15, 056003. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Armando, I.; Yu, P.; Escano, C.; Mueller, S.C.; Asico, L.; Pascua, A.; Lu, Q.; Wang, X.; Villar, V.A.; et al. Dopamine 5 receptor mediates Ang II type 1 receptor degradation via a ubiquitin-proteasome pathway in mice and human cells. J. Clin. Invest. 2008, 118, 2180–2189. [Google Scholar] [CrossRef] [Green Version]
- McNally, K.E.; Cullen, P.J. Endosomal retrieval of cargo: Retromer is not alone. Trends Cell Biol. 2018, 28, 807–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonetti, B.; Danson, C.M.; Heesom, K.J.; Cullen, P.J. Sequence-dependent cargo recognition by SNX-BARs mediates retromer-independent transport of CI-MPR. J. Cell Biol. 2017, 216, 3695–3712. [Google Scholar] [CrossRef] [Green Version]
- Yong, X.; Zhao, L.; Deng, W.; Sun, H.; Zhou, X.; Mao, L.; Hu, W.; Shen, X.; Sun, Q.; Billadeau, D.D.; et al. Mechanism of cargo recognition by retromer-linked SNX-BAR proteins. PLoS. Biol. 2020, 18, e3000631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuho, I.; Balaji, S.; Muntean, B.S.; Skamangas, N.K.; Chavali, S.; Tesmer, J.J.G.; Babu, M.M.; Martemyanov, K.A. A global map of G protein signaling regulation by RGS proteins. Cell 2020, 183, 503–521. [Google Scholar] [CrossRef]
- Ma, M.; Burd, C.G. Retrograde trafficking and plasma membrane recycling pathways of the budding yeast Saccharomyces cerevisiae. Traffic 2020, 21, 45–59. [Google Scholar] [CrossRef]
- van Weering, J.R.; Sessions, R.B.; Traer, C.J.; Kloer, D.P.; Bhatia, V.K.; Stamou, D.; Carlsson, S.R.; Hurley, J.H.; Cullen, P.J. Molecular basis for SNX-BAR-mediated assembly of distinct endosomal sorting tubules. EMBO J. 2012, 31, 4466–4480. [Google Scholar] [CrossRef]
- Seaman, M.N.J. Retromer and the cation-independent mannose 6-phosphate receptor-time for a trial separation? Traffic 2018, 19, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Norris, A.; Tammineni, P.; Wang, S.; Gerdes, J.; Murr, A.; Kwan, K.Y.; Cai, Q.; Grant, B.D. SNX-1 and RME-8 oppose the assembly of HGRS-1/ESCRT-0 degradative microdomains on endosomes. Proc. Natl. Acad. Sci. USA 2017, 114, E307–E316. [Google Scholar] [CrossRef] [Green Version]
- Norris, A.; Grant, B.D. Endosomal microdomains: Formation and function. Curr. Opin. Cell Biol. 2020, 65, 86–95. [Google Scholar] [CrossRef]
- Simonetti, B.; Paul, B.; Chaudhari, K.; Weeratunga, S.; Steinberg, F.; Gorla, M.; Heesom, K.J.; Bashaw, G.J.; Collins, B.M.; Cullen, P.J. Molecular identification of a BAR domain-containing coat complex for endosomal recycling of transmembrane proteins. Nat. Cell Biol. 2019, 21, 1219–1233. [Google Scholar] [CrossRef] [PubMed]
- Dores, M.R.; Trejo, J. Endo-lysosomal sorting of G-protein-coupled receptors by ubiquitin: Diverse pathways for G-protein-coupled receptor destruction and beyond. Traffic 2019, 20, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Schwihla, M.; Korbei, B. The Beginning of the end: Initial steps in the degradation of plasma membrane proteins. Front Plant Sci. 2020, 11, 680. [Google Scholar] [CrossRef] [PubMed]
- Greaves, J.; Prescott, G.R.; Gorleku, O.A.; Chamberlain, L.H. The fat controller: Roles of palmitoylation in intracellular protein trafficking and targeting to membrane microdomains (Review). Mol. Membr. Biol. 2009, 26, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, E.; Hoogenraad, C.C. Polarized trafficking: The palmitoylation cycle distributes cytoplasmic proteins to distinct neuronal compartments. Curr. Opin. Cell Biol. 2018, 50, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Jean-Charles, P.Y.; Snyder, J.C.; Shenoy, S.K. Chapter One—Ubiquitination and deubiquitination of G protein-coupled receptors. Prog. Mol. Biol. Transl. Sci. 2016, 141, 1–55. [Google Scholar]
- Williams, R.L.; Urbe, S. The emerging shape of the ESCRT machinery. Nat. Rev. Mol. Cell Biol. 2007, 8, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Lucas, M.; Gershlick, D.C.; Vidaurrazaga, A.; Rojas, A.L.; Bonifacino, J.S.; Hierro, A. Structural mechanism for cargo recognition by the retromer complex. Cell 2016, 167, 1623–1635.e1614. [Google Scholar] [CrossRef] [Green Version]
- Bottcher, R.T.; Stremmel, C.; Meves, A.; Meyer, H.; Widmaier, M.; Tseng, H.Y.; Fassler, R. Sorting nexin 17 prevents lysosomal degradation of beta1 integrins by binding to the β1- integrin tail. Nat. Cell Biol. 2012, 14, 584–592. [Google Scholar] [CrossRef]
- McNally, K.E.; Faulkner, R.; Steinberg, F.; Gallon, M.; Ghai, R.; Pim, D.; Langton, P.; Pearson, N.; Danson, C.M.; Nagele, H.; et al. Retriever is a multiprotein complex for retromer-independent endosomal cargo recycling. Nat. Cell Biol. 2017, 19, 1214–1225. [Google Scholar] [CrossRef]
- Clairfeuille, T.; Mas, C.; Chan, A.S.M.; Yang, Z.; Tello-Lafoz, M.; Chandra, M.; Widagdo, J.; Kerr, M.C.; Paul, B.; Mérida, I.; et al. A molecular code for endosomal recycling of phosphorylated cargos by the SNX27-retromer complex. Nat. Struct. Mol. Biol. 2016, 23, 921. [Google Scholar] [CrossRef] [PubMed]
- Zechi-Ceide, R.M.; Rodrigues, M.G.; Jehee, F.S.; Kokitsu-Nakata, N.M.; Passos-Bueno, M.R.; Guion-Almeida, M.L. Saethre-Chotzen phenotype with learning disability and hyper IgE phenotype in a patient due to complex chromosomal rearrangement involving chromosomes 3 and 7. Am. J. Med. Genet. A 2012, 158, 1680–1685. [Google Scholar] [CrossRef]
- Huang, H.S.; Yoon, B.J.; Brooks, S.; Bakal, R.; Berrios, J.; Larsen, R.S.; Wallace, M.L.; Han, J.E.; Chung, E.H.; Zylka, M.J.; et al. Snx14 regulates neuronal excitability, promotes synaptic transmission, and is imprinted in the brain of mice. PLoS ONE 2014, 9, e98383. [Google Scholar] [CrossRef]
- Bryant, D.; Seda, M.; Peskett, E.; Maurer, C.; Pomeranz, G.; Ghosh, M.; Hawkins, T.A.; Cleak, J.; Datta, S.; Hariri, H.; et al. Diverse species-specific phenotypic consequences of loss of function sorting nexin 14 mutations. Sci. Rep. 2020, 10, 13763. [Google Scholar] [CrossRef]
- Hu, Y.F.; Zhang, H.L.; Cai, T.; Harashima, S.; Notkins, A.L. The IA-2 interactome. Diabetologia 2005, 48, 2576–2581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bard, M.P.; Hegmans, J.P.; Hemmes, A.; Luider, T.M.; Willemsen, R.; Severijnen, L.A.; van Meerbeeck, J.P.; Burgers, S.A.; Hoogsteden, H.C.; Lambrecht, B.N. Proteomic analysis of exosomes isolated from human malignant pleural effusions. Am J Respir Cell Mol Biol. 2004, 31, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Muglia, M.; Magariello, A.; Citrigno, L.; Passamonti, L.; Sprovieri, T.; Conforti, F.L.; Mazzei, R.; Patitucci, A.; Gabriele, A.L.; Ungaro, C.; et al. A novel locus for dHMN with pyramidal features maps to chromosome 4q34.3–q35.2. Clin. Genet. 2008, 73, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.K.; Gudmundsdottir, V.; Brunak, S. Pancreatic islet protein complexes and their dysregulation in type 2 diabetes. Front Genet. 2017, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Mulindwa, J.; Noyes, H.; Ilboudo, H.; Pagani, L.; Nyangiri, O.; Kimuda, M.P.; Ahouty, B.; Asina, O.F.; Ofon, E.; Kamoto, K.; et al. High levels of genetic diversity within Nilo-Saharan populations: Implications for human adaptation. Am. J. Hum. Genet. 2020, 107, 473–486. [Google Scholar] [CrossRef]
- Ebrahimi-Fakhari, D. Congenital disorders of autophagy: What a pediatric neurologist should know. Neuropediatrics 2018, 49, 18–25. [Google Scholar] [CrossRef]
- Ma, L.; Semick, S.A.; Chen, Q.; Li, C.; Tao, R.; Price, A.J.; Shin, J.H.; Jia, Y. BrainSeq Consortium, Brandon, N.J.; Cross, A.J.; et al. Schizophrenia risk variants influence multiple classes of transcripts of sorting nexin 19 (SNX19). Mol. Psychiatry 2020, 25, 831–843. [Google Scholar] [CrossRef]
- Martin, A.R.; Lin, M.; Granka, J.M.; Myrick, J.W.; Liu, X.; Sockell, A.; Atkinson, E.G.; Werely, C.J.; Möller, M.; Sandhu, M.S.; et al. An unexpectedly complex architecture for skin pigmentation in Africans. Cell 2017, 171, 1340–1353.e14. [Google Scholar] [CrossRef] [Green Version]
- Ugrankar, R.; Bowerman, J.; Hariri, H.; Chandra, M.; Chen, K.; Bossanyi, M.F.; Datta, S.; Rogers, S.; Eckert, K.M.; Vale, G.; et al. Drosophila snazarus regulates a lipid droplet population at plasma membrane-droplet contacts in adipocytes. Dev. Cell 2019, 50, 557–572.e5. [Google Scholar] [CrossRef] [PubMed]
- Hugenroth, M.; Bohnert, M. Come a little bit closer! Lipid droplet-ER contact sites are getting crowded. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118603. [Google Scholar] [CrossRef] [PubMed]
- Hines, R.M.; Kang, R.; Goytain, A.; Quamme, G.A. Golgi-specific DHHC zinc finger protein GODZ mediates membrane Ca2+ transport. J. Biol. Chem. 2010, 285, 4621–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felder, R.A.; Sanada, H.; Xu, J.; Yu, P.Y.; Wang, Z.; Watanabe, H.; Asico, L.D.; Wang, W.; Zheng, S.; Yamaguchi, I.; et al. G protein-coupled receptor kinase 4 gene variants in human essential hypertension. Proc. Natl. Acad. Sci. USA 2002, 99, 3872–3877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Han, W.; Villar, V.A.; Keever, L.B.; Lu, Q.; Hopfer, U.; Quinn, M.T.; Felder, R.A.; Jose, P.A.; Yu, P. D1-like receptors regulate NADPH oxidase activity and subunit expression in lipid raft microdomains of renal proximal tubule cells. Hypertension 2009, 53, 1054–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, W.; Li, H.; Villar, V.A.; Pascua, A.M.; Dajani, M.I.; Wang, X.; Natarajan, A.; Quinn, M.T.; Felder, R.A.; Jose, P.A.; et al. Lipid rafts keep NADPH oxidase in the inactive state in human renal proximal tubule cells. Hypertension 2008, 51, 481–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Li, X.X.; Albrecht, F.E.; Hopfer, U.; Carey, R.M.; Jose, P.A. Dopamine(1) receptor, Gsα, and Na(+)-H(+) exchanger interactions in the kidney in hypertension. Hypertension 2000, 36, 395–399. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Subfamily * | Members | Major Domain Architecture ** | Roles in Signaling, Trafficking, and Degradation |
|---|---|---|---|
| SNX-PXA-RGS-PXC (4) | SNX13, SNX14, SNX19, SNX25 |  | Plays important roles in receptor signaling and membrane trafficking, see text for details. |
| SNX-FERM (3) | SNX17, SNX27, SNX31 |  | Involved in cargo loading and binding to membrane structures and endosome to plasma membrane trafficking or lysosomal degradation. |
| SNX-BAR (12) | SNX1, SNX2, SNX4, SNX5, SNX6, SNX7, SNX8, SNX9, SNX18, SNX30, SNX32, SNX33 |  | Recognizes and targets to a wide range of cargoes, in coordination with retromers or other retrieval machineries to regulate receptor signaling and trafficking in retromer-dependent and -independent manners. |
| SNX-PX (10) | SNX3, SNX10, SNX11, SNX12, SNX16, SNX20, SNX21, SNX22, SNX24, SNX29 |  | Forms endosome transport carriers in retromer-dependent or -independent manners on a diversity of cargo sorting, retrograde protein trafficking, and lysosomal degradation. |
| Unclassified SNXs (4) | SNX15, SNX23 SNX26, SNX28 |  | Binds to endosomes in Ca2+-dependent or -independent manners; regulates cargoes, such as amyloid-β precursor protein recycling to cell surface and processing for amyloid-β generation. |
| SNX | Chromosomal Locus | Major Cellular Distribution | Major Tissue Distribution | Phosphoinositide Binding Preferences | References |
|---|---|---|---|---|---|
| SNX13 | 7p21(human) 12(mouse) | Endosome ER | Pancreas Heart CNS Adipose Spleen | PI(3)P PI(3,4)P2 PI(3,5)P2 PI(4,5)P2 PI(3,4,5)P3 | [5,6,7,9,10,11,12,13] |
| SNX14 | 6q14(human) 9(mouse) | Lysosomes ER Lipid droplets | CNS Adipose Lung Heart Testis | PI(3,5)P2 | [5,6,7,9,10,11,13,14,15,16,17] |
| SNX19 | 11q24.3-q25(human) 9(mouse) | Early endosomes Plasma membrane Mitochondria | Kidney CNS Bone marrow Heart Pancreas | PI(3)P PI(4,5)P2 PI(3,4,5)P3 | [5,6,7,9,10,11,18,19,20,21,22,23] |
| SNX25 | 4q35(human) 8A4(mouse) | Endosomes Lysosomes Nucleus | Lung Kidney CNS | PI(3,4)P2 PI(3,5)P2 PI(4,5)P2 PI(3,4,5)P3 PI(3)P | [5,6,7,9,10,11,23,24,25,26,27] |
| SNX | RGS Domain | Gαs Interaction | GAP Activity | Gαs Signaling | GPCR Cargo Example | Reference(s) |
|---|---|---|---|---|---|---|
| SNX13 | + | + | + | inhibition | β2-AR EGFR | [33,35] |
| SNX14 | + | + | - | inhibition | 5-HT6R | [13] |
| SNX19 | - | NA | NA | NA | D1R | [20] |
| SNX25 | + | ND | ND | ND | D1R D2R TGF-β1 | [23,27,36] |
| Subfamily | Signaling | Trafficking Function | Disease Links | References |
|---|---|---|---|---|
| SNX13 | Gαs inhibition | Lysosomal degradation | Saethre-Chotzen syndrome phenotype Type 2 diabetes Skin pigmentation | [33,35,89,95,96] |
| SNX14 | cAMP/PKA inhibition | Lysosomal degradation | SCAR20 Neuron development and differentiation Microcephaly Down syndrome Cerebellar ataxia Intellectual disability Congenital disorders of autophagy Squamous cell carcinoma | [17,90,91,97] |
| SNX19 | Palmitoylation Akt/PKB | Lipid raft targeting Lysosomal degradation Recycling | Hypertension Type I diabetes Atherosclerosis Schizophrenia | [18,19,20,23,92,98] |
| SNX25 | TGFβ-SMAD phosphorylation? | Lysosomal degradation | Temporal lobe epilepsy dHMN LOAD EOAD Hypertension | [25,26,27,94] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amatya, B.; Lee, H.; Asico, L.D.; Konkalmatt, P.; Armando, I.; Felder, R.A.; Jose, P.A. SNX-PXA-RGS-PXC Subfamily of SNXs in the Regulation of Receptor-Mediated Signaling and Membrane Trafficking. Int. J. Mol. Sci. 2021, 22, 2319. https://doi.org/10.3390/ijms22052319
Amatya B, Lee H, Asico LD, Konkalmatt P, Armando I, Felder RA, Jose PA. SNX-PXA-RGS-PXC Subfamily of SNXs in the Regulation of Receptor-Mediated Signaling and Membrane Trafficking. International Journal of Molecular Sciences. 2021; 22(5):2319. https://doi.org/10.3390/ijms22052319
Chicago/Turabian StyleAmatya, Bibhas, Hewang Lee, Laureano D. Asico, Prasad Konkalmatt, Ines Armando, Robin A. Felder, and Pedro A. Jose. 2021. "SNX-PXA-RGS-PXC Subfamily of SNXs in the Regulation of Receptor-Mediated Signaling and Membrane Trafficking" International Journal of Molecular Sciences 22, no. 5: 2319. https://doi.org/10.3390/ijms22052319