Double Gamers—Can Modified Natural Regulators of Higher Plants Act as Antagonists against Phytopathogens? The Case of Jasmonic Acid Derivatives

 ,
,  ,
,  , , , ,
, , , ,  ,
,  ,
,
Abstract
:
1. Introduction
2. Results
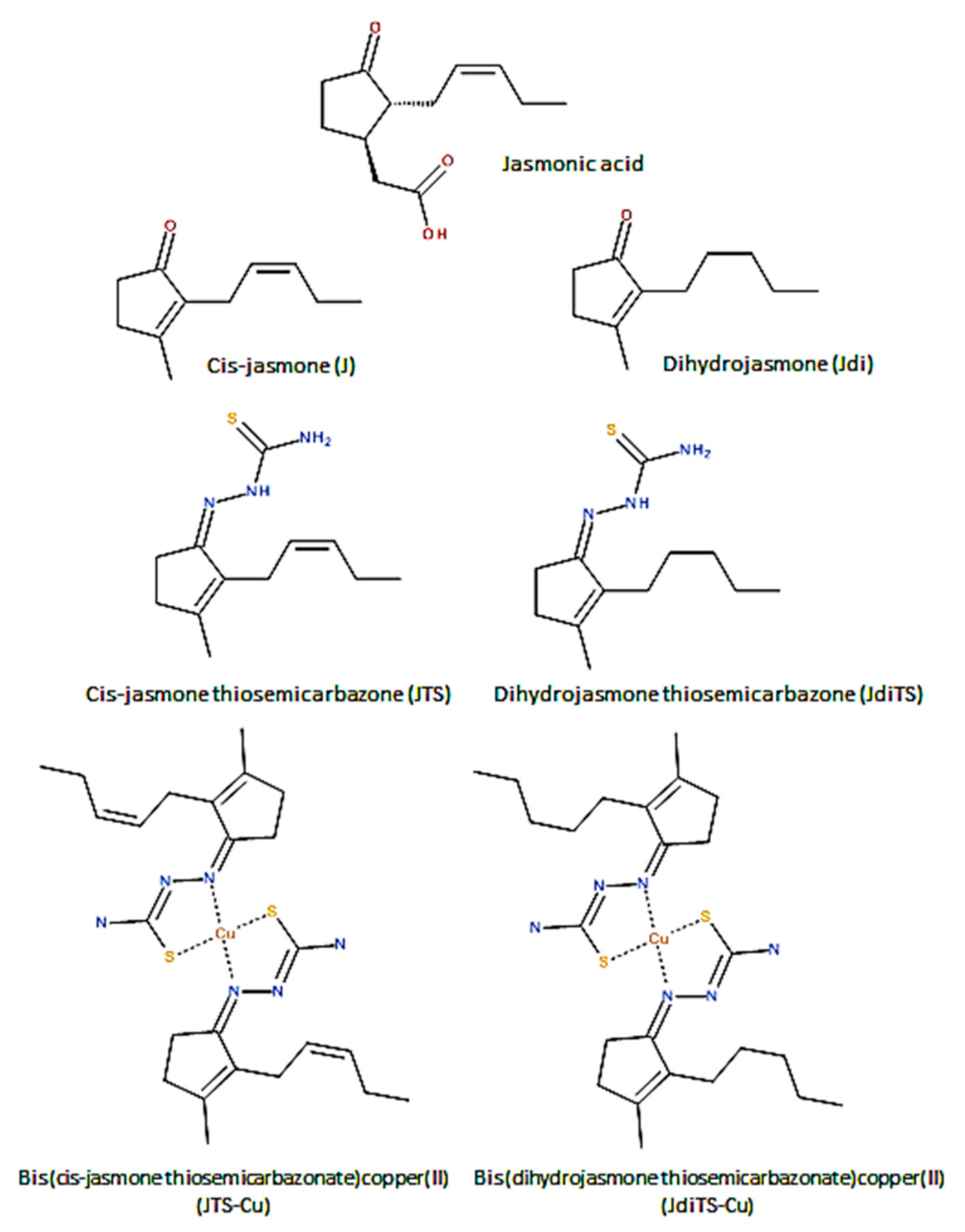
2.1. Chemistry
2.2. Scavenging Potential Determination of Compounds
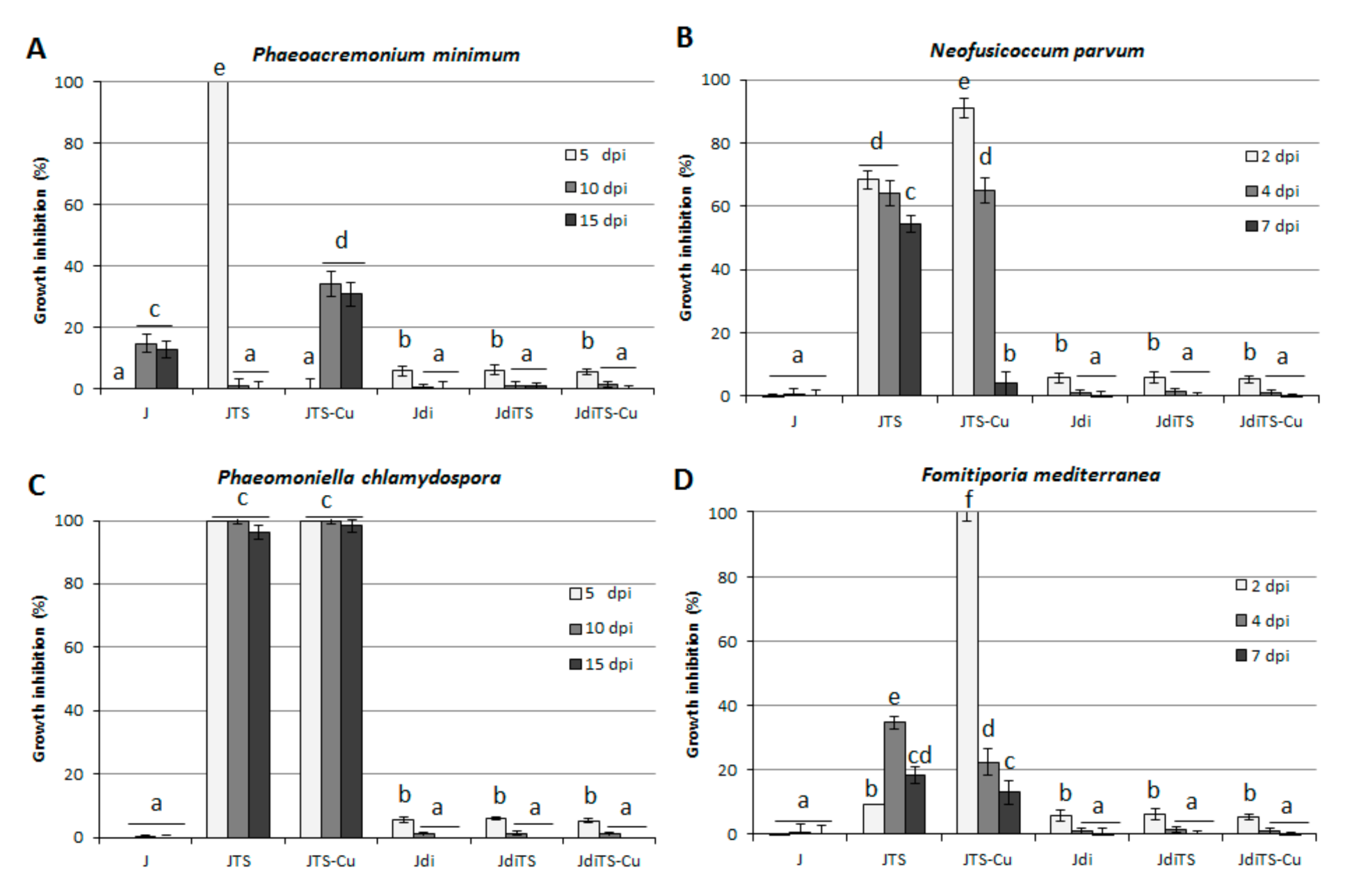
2.3. Antifungal Activity on Grapevine Esca-Associated Pathogens
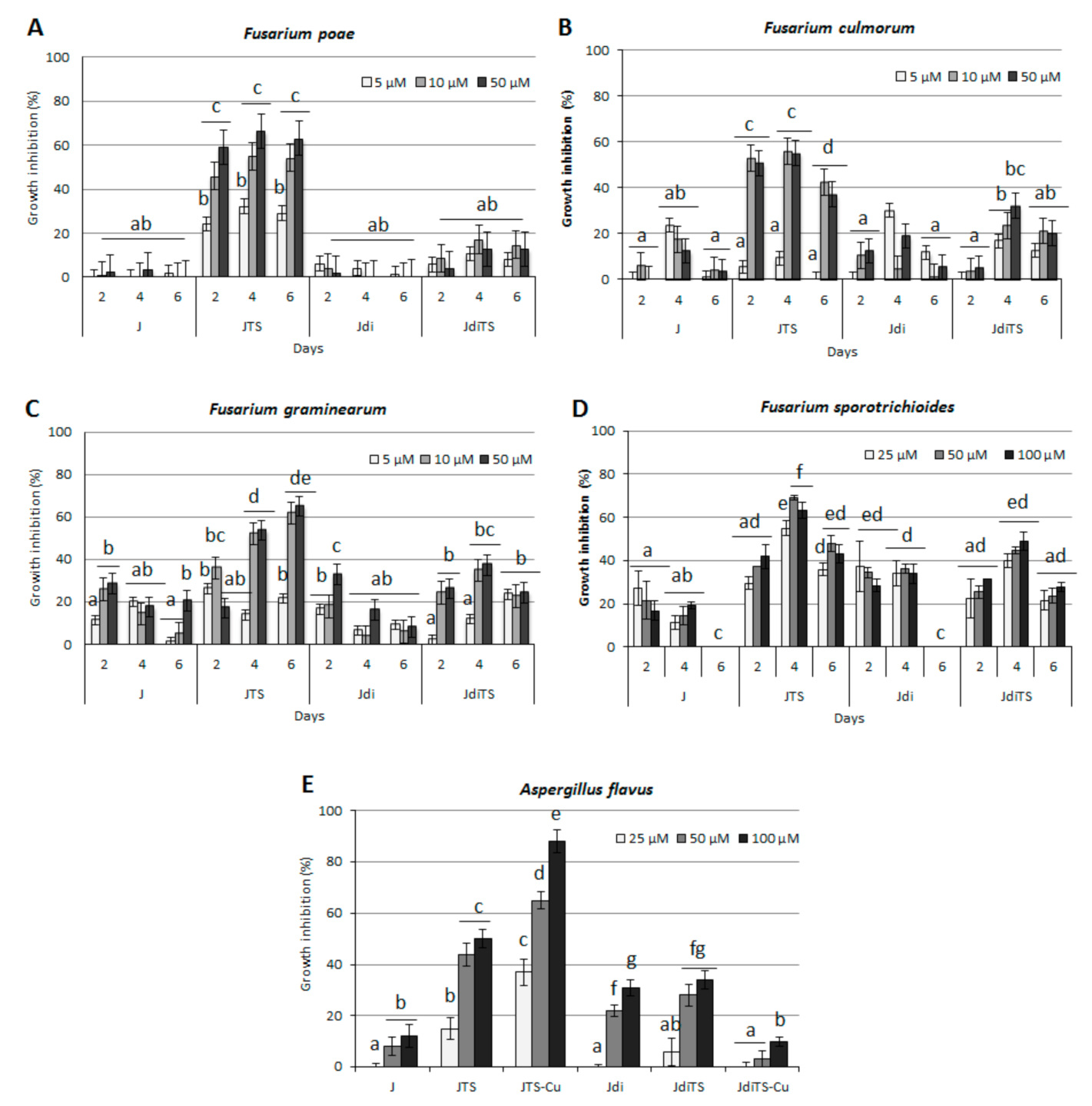
2.4. Antifungal Activity on Cereals Pathogens
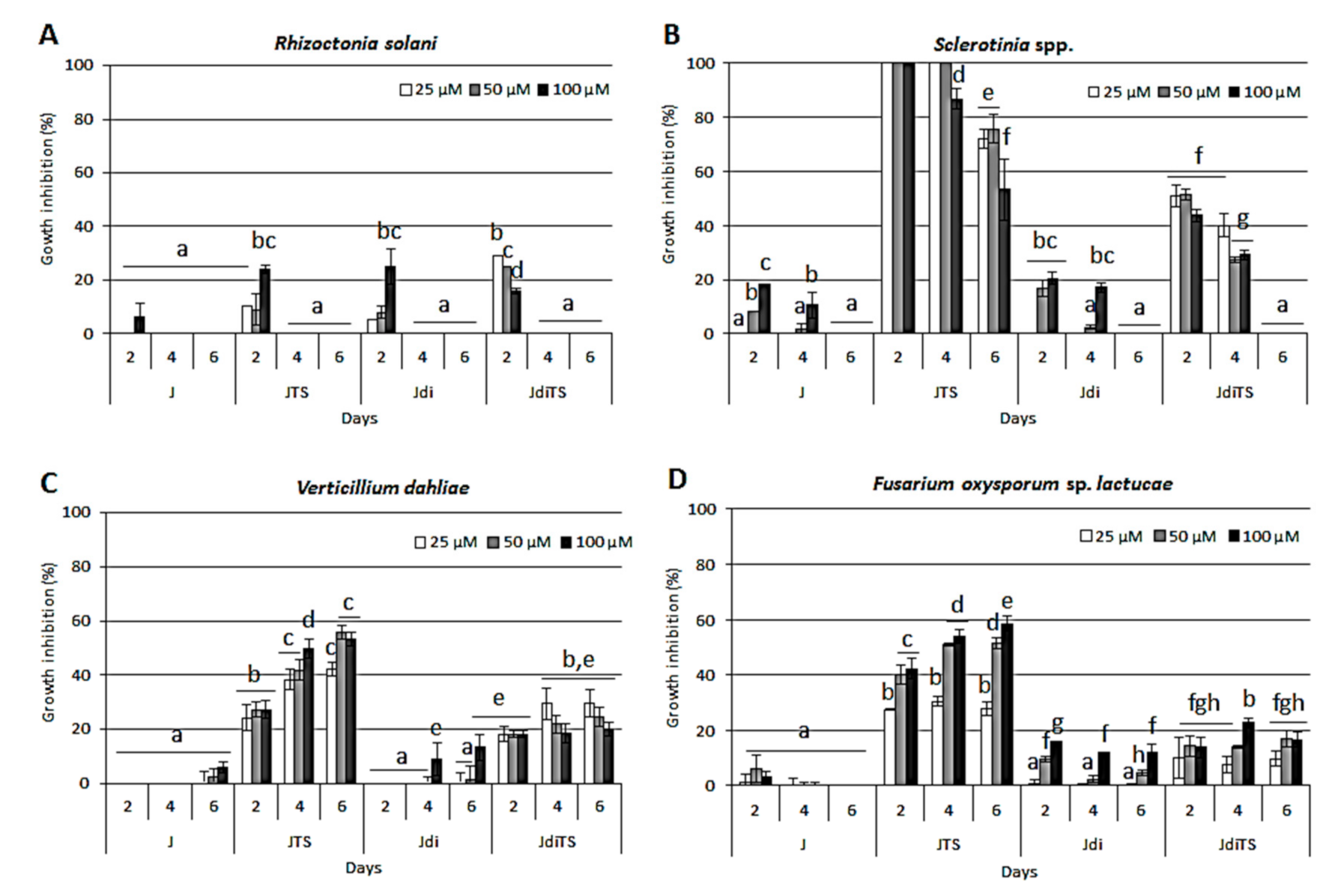
2.5. Antifungal Activity on Horticultural Crop Pathogens—Sclerotinia spp., V. dahliae, Rhizoctonia solani and F. oxysporum
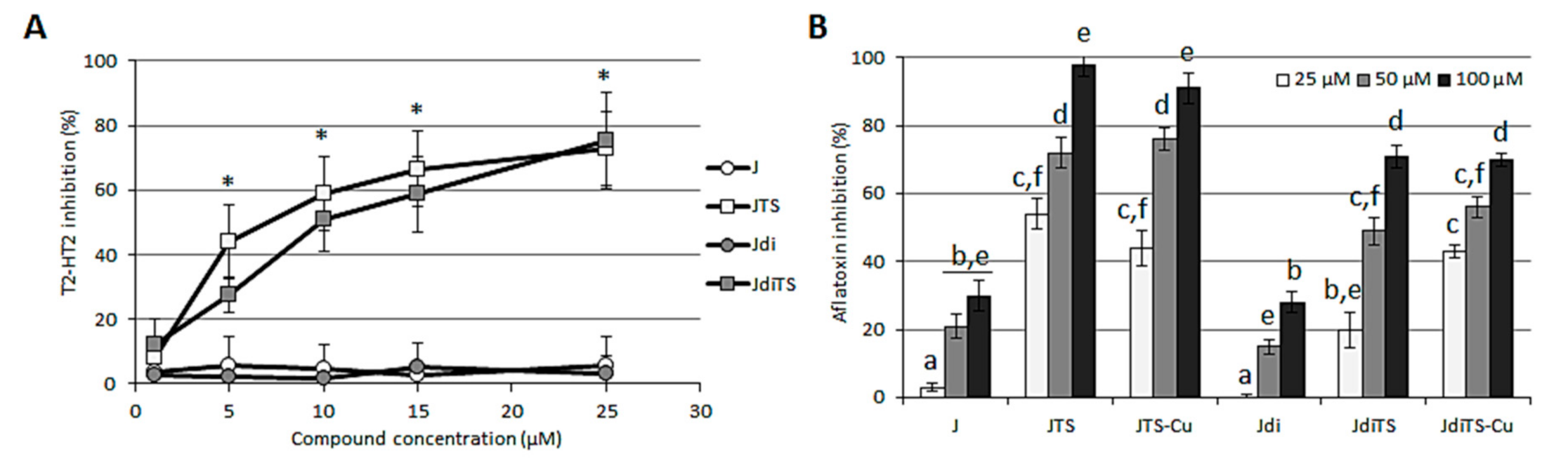
2.6. Interference with Fungal Secondary Metabolism—Effect on Mycotoxin Production and Sclerotia Development
3. Discussion
4. Materials and Methods
4.1. Chemical Synthesis
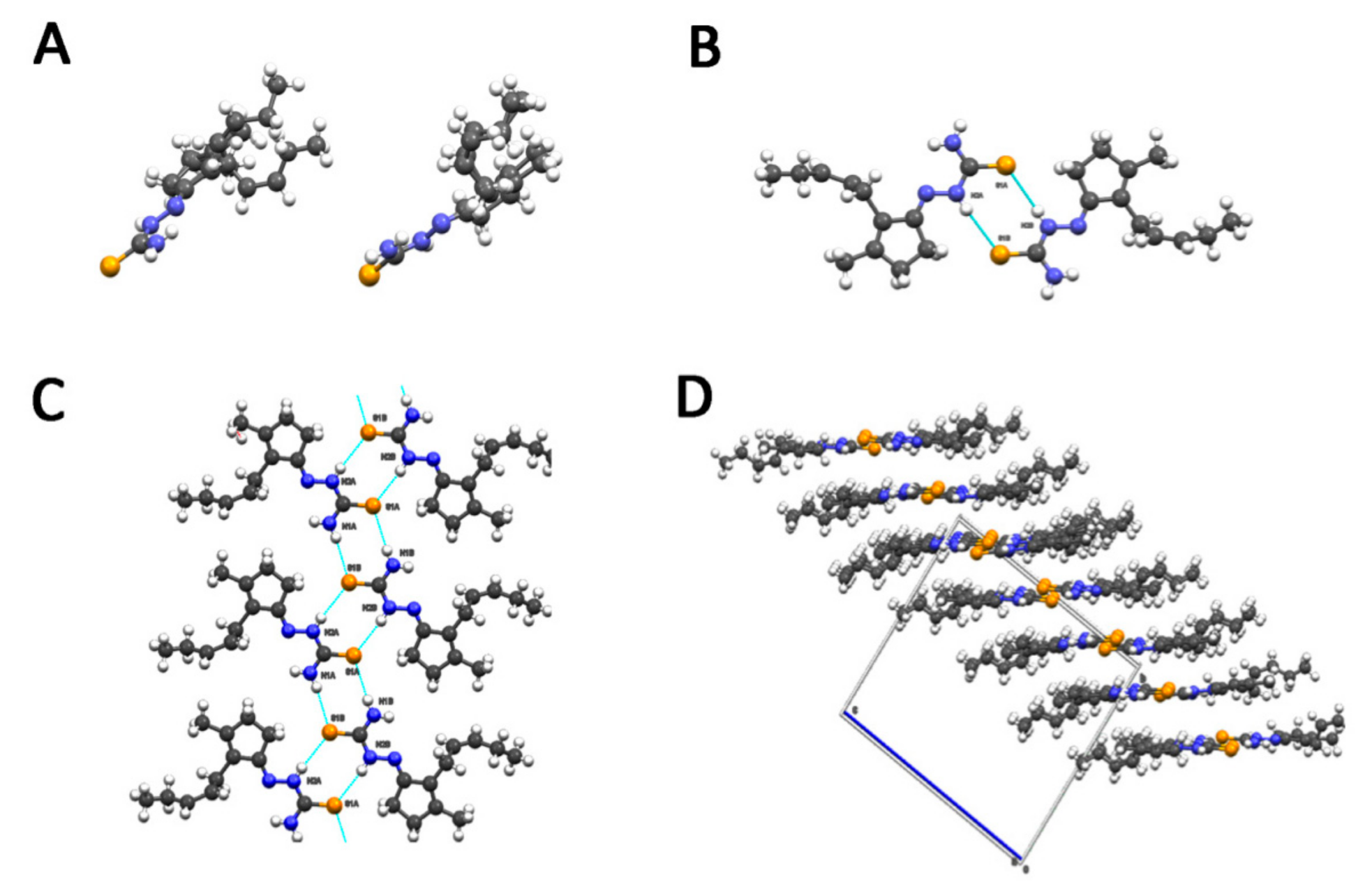
4.2. X-ray Analysis
4.3. Scavenging Activity Assay (DPPH)
4.4. Biological Assays
4.4.1. Microorganisms
4.4.2. Grapewine Diseases-Associated Fungal Species Growth Inhibition Assay
4.4.3. Fusarium spp. and Soil-Borne Pathogens Growth Inhibition Assay
4.4.4. F. sporotrichioides T2-HT2 Toxins Determination
4.4.5. Aflatoxin, Biomass and Sclerotia Inhibition Assays in A. flavus
4.4.6. Sclerotia Inhibition Assays in A. flavus and Sclerotinia spp.
4.4.7. Statistical Analysis of Biological Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, H.; Liu, B.; Liu, L.; Song, S. Jasmonate action in plant growth and development. J. Exp. Bot. 2017, 68, 1349–1359. [Google Scholar] [CrossRef] [Green Version]
- Irankhah, S.; Chitarra, W.; Nerva, L.; Antoniou, C.; Lumini, E.; Volpe, V.; Ganjeali, A.; Cheniany, M.; Mashregh, M.; Fotopoulos, V.; et al. Impact of an arbuscular mycorrhizal fungal inoculum and exogenous MeJA on fenugreek secondary metabolite production under water deficit. Environ. Exp. Bot. 2020, 176, 104096. [Google Scholar] [CrossRef]
- Chanclud, E.; Morel, J.B. Plant hormones: A fungal point of view. Mol. Plant Pathol. 2016, 17, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Kamisaka, S.; Yanagishima, N.; Masuda, Y. Effect of auxin and gibberellin on sporulation in yeast. Physiol. Plant. 1967, 20, 90–97. [Google Scholar] [CrossRef]
- Nakamura, T.; Mukai, C.; Ozaki, Y.; Saotome, M.; Murayama, T. Effects of auxin and gibberellin on conidial germination and elongation of young hyphae in a cyclic 3′:5′ adenosine monophosphate-dependent protein kinase mutant of Neurospora crassa. Plant Growth Regul. 1988, 7, 201–207. [Google Scholar]
- Degani, O.; Drori, R.; Goldblat, Y. Plant growth hormones suppress the development of Harpophora maydis, the cause of late wilt in maize. Physiol. Mol. Biol. Plants 2015, 21, 137–149. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Qasim, M.; Hussain, M.; Akutse, K.S.; Avery, P.B.; Dash, C.K.; Wang, L. The herbivore-induced plant volatiles methyl salicylate and menthol positively affect growth and pathogenicity of entomopathogenic fungi. Sci. Rep. 2017, 7, 40494. [Google Scholar] [CrossRef] [PubMed]
- Morishita, Y.; Okazaki, Y.; Luo, Y.Y.; Nunoki, J.; Taniguchi, T.; Oshima, Y.; Asai, T. Use of plant hormones to activate silent polyketide biosynthetic pathways in Arthrinium sacchari, a fungus isolated from a spider. Org. Biomol. Chem. 2019, 17, 780. [Google Scholar] [CrossRef]
- Norman, S.M.; Bennett, R.D.; Maier, V.P.; Poling, S.M. Cytokinins inhibit abscisic acid biosynthesis in Cercospora rosicola. Plant Sci. Lett. 1983, 28, 255–263. [Google Scholar] [CrossRef]
- Tudzynski, B.; Hölter, K. Gibberellin biosynthetic pathway in Gibberella fujikuroi: Evidence for a gene cluster. Fungal Genet. Biol. 1998, 25, 157–170. [Google Scholar] [CrossRef]
- Takino, J.; Kozaki, T.; Sato, Y.; Liu, C.; Ozaki, T.; Minami, A.; Oikawa, H. Unveiling biosynthesis of the phytohormone abscisic acid in fungi: Unprecedented mechanism of core scaffold formation catalyzed by an unusual sesquiterpene synthase. J. Am. Chem. Soc. 2018, 140, 12392–12395. [Google Scholar] [CrossRef]
- Nerva, L.; Zanzotto, A.; Gardiman, M.; Gaiotti, F.; Chitarra, W. Soil microbiome analysis in an ESCA diseased vineyard. Soil Biol. Biochem. 2019, 135, 60–70. [Google Scholar] [CrossRef]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Juan, C.; Ferrer, E.; Mañes, J. Fusarium species, chemotype characterisation and trichothecene contamination of durum and soft wheat in an area of central Italy. J. Sci. Food Agric. 2015, 95, 540–551. [Google Scholar] [CrossRef] [PubMed]
- Salgado, J.D.; Madden, L.V.; Paul, P.A. Efficacy and economics of integrating in-field and harvesting strategies to manage Fusarium head blight of wheat. Plant Dis. 2014, 98, 1407–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [Green Version]
- Mielniczuk, E.; Skwaryło-Bednarz, B. Fusarium Head Blight, mycotoxins and strategies for their reduction. Agronomy 2020, 10, 509. [Google Scholar] [CrossRef] [Green Version]
- Tournas, V.H. Spoilage of vegetable crops by bacteria and fungi and related health hazards. Crit. Rev. Microbiol. 2005, 31, 33–44. [Google Scholar] [CrossRef]
- Gross-Steinmeyer, K.; Eaton, D.L. Dietary modulation of the biotransformation and genotoxicity of aflatoxin B(1). Toxicology 2012, 299, 69–79. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Streit, E. Global occurrence of mycotoxins in the food and feed chain: Facts and figures. World Mycotoxin J. 2013, 6, 213–222. [Google Scholar] [CrossRef]
- Garibaldi, A.; Gullino, M.L. Emerging soilborne diseases of horticultural crops and new trends in their management. Acta Hortic. 2010, 883, 37–48. [Google Scholar] [CrossRef]
- Pelosi, G. Thiosemicarbazone metal complexes: From structure to activity. Open Crystall. J. 2010, 3, 16–28. [Google Scholar] [CrossRef]
- Gowda, N.K.S.; Malathi, V.; Suganthi, R.U. Effect of some chemical and herbal compounds on growth of Aspergillus parasiticus and aflatoxin production. Anim. Feed Sci. Technol. 2004, 116, 281–291. [Google Scholar] [CrossRef]
- Rogolino, D.; Gatti, A.; Carcelli, M.; Pelosi, G.; Bisceglie, F.; Restivo, F.M.; Degola, F.; Buschini, A.; Montalbano, S.; Feretti, D.; et al. Thiosemicarbazone scaffold for the design of antifungal and antiaflatoxigenic agents: Evaluation of ligands and related copper complexes. Sci. Rep. 2017, 7, 11214. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.W.; Kim, S.C.; Hwang, S.S.; Choi, B.K.; Ahn, H.J.; Lee, M.Y.; Park, S.H.; Kim, S.K. Antioxidant activity and free radical scavenging capacity between Korean medicinal plants and flavonoids by assay-guided comparison. Plant Sci. 2002, 163, 1161–1168. [Google Scholar] [CrossRef]
- Nerva, L.; Turina, M.; Zanzotto, A.; Gardiman, M.; Gaiotti, F.; Gambino, G.; Chitarra, W. Isolation, molecular characterization and virome analysis of culturable wood fungal endophytes in esca symptomatic and asymptomatic grapevine plants. Environ. Microbiol. 2019, 21, 2886–2904. [Google Scholar] [CrossRef] [PubMed]
- Carvalhais, L.C.; Schenk, P.M.; Dennis, P.G. Jasmonic acid signalling and the plant holobiont. Curr. Opin. Microbiol. 2017, 37, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Prost, I.; Dhondt, S.; Rothe, G.; Vicente, J.; Rodriguez, M.J.; Kift, N.; Carbonne, F.; Griffiths, G.; Esquerré-Tugayé, M.-T.; Rosahl, S.; et al. Evaluation of the antimicrobial activities of plant oxylipins supports their involvement in defense against pathogens. Plant Physiol. 2005, 139, 1902–1913. [Google Scholar] [CrossRef] [Green Version]
- Goodrich-Tanrikulu, M.; Mahoney, N.E.; Rodriguezt, S.B. The plant growth regulator methyl jasmonate inhibits aflatoxin production by Aspergillus flavus. Microbiology 1995, 141, 2831–2837. [Google Scholar] [CrossRef] [Green Version]
- Fischer, M.; Peighami-Ashnaei, S. Grapevine, esca complex and environment: The disease triangle. Phytopathol. Mediterr. 2019, 58, 17–37. [Google Scholar]
- Fischer, J.; Beckers, S.J.; Yiamsawas, D.; Thines, E.; Landfester, K.; Wurm, F.R. Targeted Drug delivery in plants: Enzyme-responsive lignin nanocarriers for the curative treatment of the worldwide grapevine trunk disease Esca. Adv. Sci. 2019, 6, 1802315. [Google Scholar] [CrossRef] [Green Version]
- Paiva, R.; Kneipp, L.F.; Goular, C.M.; Albuquerque, M.A.; Echevarria, A. Antifungal activities of thiosemicarbazones and semicarbazones against mycotoxigenic fungi. Ciên. Agrotec. 2014, 38, 531–537. [Google Scholar] [CrossRef] [Green Version]
- Degola, F.; Morcia, C.; Bisceglie, F.; Mussi, F.; Tumino, G.; Ghizzoni, R.; Pelosi, G.; Terzi, V.; Buschini, A.; Restivo, F.M.; et al. In vitro evaluation of the activity of thiosemicarbazone derivates against mycotoxigenic fungi affecting cereals. Int. J. Food Microbiol. 2015, 200, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, C.D.; Patsoukis, N.; Papapostolou, I.; Zervoudakis, G. Sclerotial metamorphosis in filamentous fungi is induced by oxidative stress. Integr. Comp. Biol. 2006, 46, 691–712. [Google Scholar] [CrossRef] [PubMed]
- Patsoukis, N.; Georgiou, C.D. Thiol redox state and oxidative stress affect sclerotial differentiation of the phytopathogenic fungi Sclerotium rolfsii and Sclerotinia sclerotiorum. J. Appl. Microbiol. 2007, 104, 42–50. [Google Scholar] [CrossRef]
- Papapostolou, I.; Georgiou, C.D. Superoxide radical induces sclerotial differentiation in filamentous phytopathogenic fungi: A superoxide dismutase mimetics study. Microbiology 2010, 156, 960–966. [Google Scholar] [CrossRef] [Green Version]
- Degola, F.; Bisceglie, F.; Pioli, M.; Palmano, S.; Elviri, L.; Pelosi, G.; Lodi, T.; Restivo, F.M. Structural modification of cuminaldehyde thiosemicarbazone increases inhibition specificity toward aflatoxin biosynthesis and sclerotia development in Aspergillus flavus. Appl. Microbiol. Biotechnol. 2017, 101, 6683–6696. [Google Scholar] [CrossRef] [PubMed]
- Sheldrick, G.M. A short history of SHELX. Acta Cryst. 2008, A64, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolomanov, L.J.; Bourhis, R.J.; Gildea, J.A.; Howard, K.; Puschmann, H. OLEX2: A complete structure solution refinement and analysis program. J. Appl. Cryst. 2009, 42, 339–341. [Google Scholar] [CrossRef]
- Macrae, C.F.; Edington, P.R.; McCabe, P.; Pidcock, E.; Shields, G.P.; Taylor, R.; Towler, M.; van de Streek, J. Mercury: Visualization and analysis of crystal structures. J. Appl. Cryst. 2006, 39, 453–457. [Google Scholar] [CrossRef] [Green Version]
- Bartoli, J.; Montalbano, S.; Spadola, G.; Rogolino, D.; Pelosi, G.; Bisceglie, F.; Restivo, F.M.; Degola, F.; Serra, O.; Buschini, A.; et al. Antiaflatoxigenic thiosemicarbazones as cropprotective agents: A cytotoxic and genotoxic study. J. Agric. Food Chem. 2019, 67, 10947–10953. [Google Scholar] [CrossRef]
- Bisceglie, F.; Degola, F.; Rogolino, D.; Giannelli, G.; Orsoni, N.; Spadola, G.; Pioli, M.; Restivo, F.M.; Carcelli, M.; Pelosi, G. Sisters in structure but different in character, some benzaldehyde and cinnamaldehyde derivatives differentially tune Aspergillus flavus secondary metabolism. Sci. Rep. 2020, 10, 17686. [Google Scholar] [CrossRef] [PubMed]
- Nerva, L.; Forgia, M.; Ciuffo, M.; Chitarra, W.; Chiapello, M.; Vallino, M.; Varese, G.C.; Turina, M. The mycovirome of a fungal collection from the sea cucumber Holothuria polii. Virus Res. 2019, 273, 197737. [Google Scholar] [CrossRef] [PubMed]
- Thanassoulopoulos, C.C.; Giannopolitis, C.N.; Vitsos, G.T. Evaluation of sensitiveness and development of resistance of Fusarium oxysporum f. sp. lycopersici to benomyl. Phytopathol. Zeitsch. 1971, 70, 114–120. [Google Scholar]
- Jamiołkowska, A.; Kowalski, R. Laboratory effect of Silphium perfoliatum L. on the growth of tested fungi. Acta Sci. Pol. Hortorum Cultus 2012, 11, 43–55. [Google Scholar]
- Degola, F.; Dall’Asta, C.; Restivo, F.M. Development of a simple and high-throughput method for detecting aflatoxins production in culture media. Lett. Appl. Microbiol. 2012, 55, 82–89. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Osdaghi, E.; Behlau, F.; Köhl, J.; Jones, J.B.; Aubertot, J.-N. Thirteen decades of antimicrobial copper compounds applied in agriculture. A review. Agron. Sustain. Dev. 2018, 38, 28. [Google Scholar] [CrossRef] [Green Version]
- La Torre, A.; Iovino, V.; Caradonia, F. Copper in plant protection: Current situation and prospects. Phytopathol. Mediterr. 2018, 57, 201–236. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | A | B | C | |
| C7–C8–C9–C10 | 138.9 (8)° | −125.1 (8)° | −130.9 (6)° | |
| C8–C9–C10–C11 | −4.3 (16)° | −2.0 (15)° | −2.0 (15)° | |
| C9–C10–C11–C12 | −153.6 (13)° | 134.3 (11)° | 123.9 (8)° |
 | Atoms | Average Length (Å) |
| C-N4 | 1.320 | |
| C-S | 1.695 | |
| C-N2 | 1.346 | |
| N2-N1 | 1.389 |
| Aspergillus flavus | Sclerotinia spp. | |
|---|---|---|
| Compound | Inhibition (%) mg/Colony Area (cm2) | Inhibition (%) nr/Colony Area (cm2) |
| JTS | 71.9 ± 3.0 * | 55.3 ± 7.1 * |
| JTS-Cu | 96.9 ± 1.6 * | n.d. |
| JdiTS | 62.5 ± 5.7 * | 52.6 ± 3.5 * |
| JdiTS-Cu | 0.0 ± 0.2 | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orsoni, N.; Degola, F.; Nerva, L.; Bisceglie, F.; Spadola, G.; Chitarra, W.; Terzi, V.; Delbono, S.; Ghizzoni, R.; Morcia, C.; et al. Double Gamers—Can Modified Natural Regulators of Higher Plants Act as Antagonists against Phytopathogens? The Case of Jasmonic Acid Derivatives. Int. J. Mol. Sci. 2020, 21, 8681. https://doi.org/10.3390/ijms21228681
Orsoni N, Degola F, Nerva L, Bisceglie F, Spadola G, Chitarra W, Terzi V, Delbono S, Ghizzoni R, Morcia C, et al. Double Gamers—Can Modified Natural Regulators of Higher Plants Act as Antagonists against Phytopathogens? The Case of Jasmonic Acid Derivatives. International Journal of Molecular Sciences. 2020; 21(22):8681. https://doi.org/10.3390/ijms21228681
Chicago/Turabian StyleOrsoni, Nicolò, Francesca Degola, Luca Nerva, Franco Bisceglie, Giorgio Spadola, Walter Chitarra, Valeria Terzi, Stefano Delbono, Roberta Ghizzoni, Caterina Morcia, and et al. 2020. "Double Gamers—Can Modified Natural Regulators of Higher Plants Act as Antagonists against Phytopathogens? The Case of Jasmonic Acid Derivatives" International Journal of Molecular Sciences 21, no. 22: 8681. https://doi.org/10.3390/ijms21228681