Exploring the Applications of the Photoprotective Properties of Anthocyanins in Biological Systems

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Chemical Equilibria and Stability of Anthocyanins
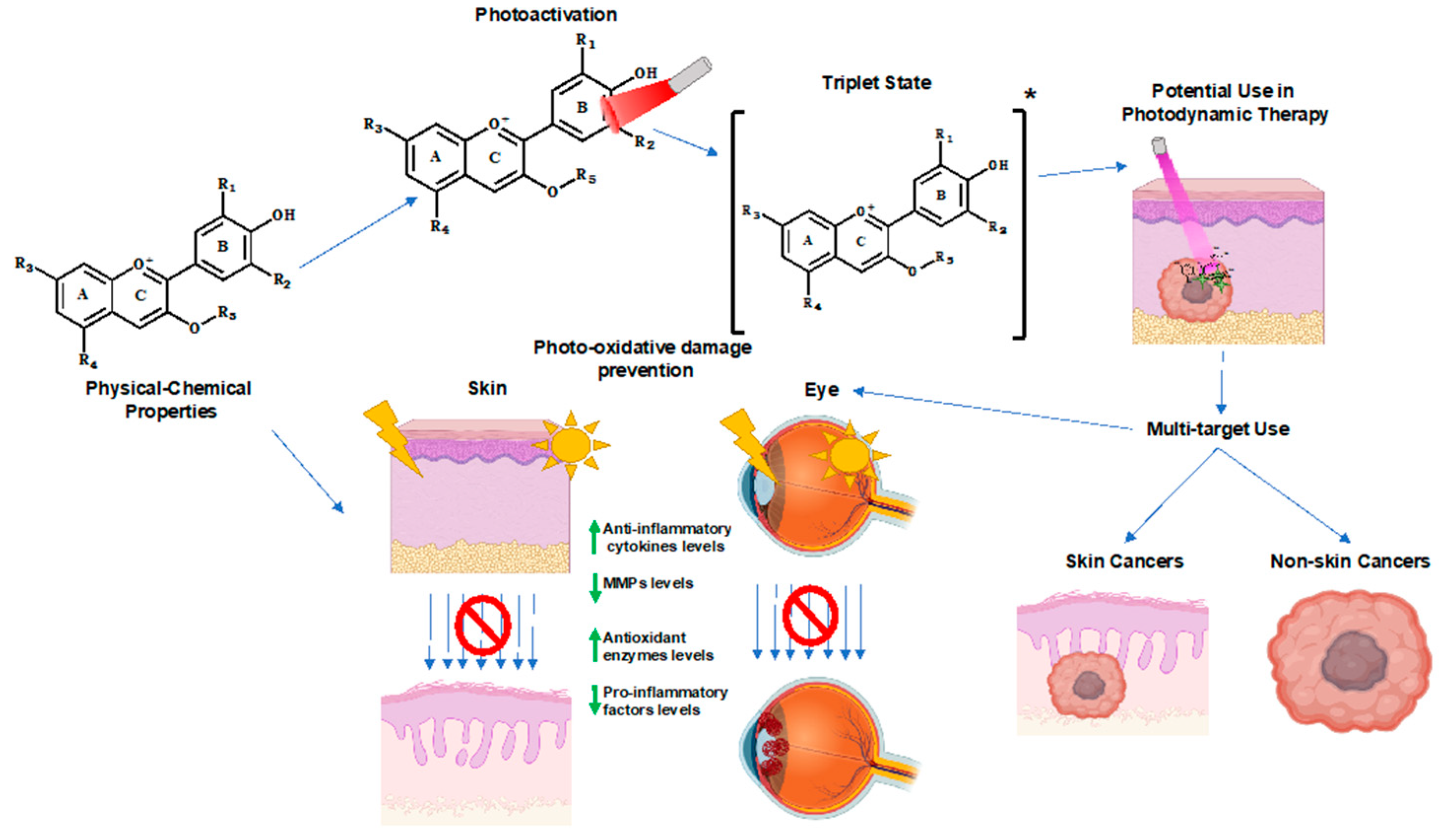
3. Photochemical Properties of Anthocyanins
4. Photo-Oxidative Damage and the Role of Anthocyanins
4.1. Skin Aging
4.2. Eye Degenerative Disease
4.2.1. Light-Induced Damage to the Retina
4.2.2. Endogenous Photosensitizers
4.2.3. Anthocyanins as Retinal Chromophores
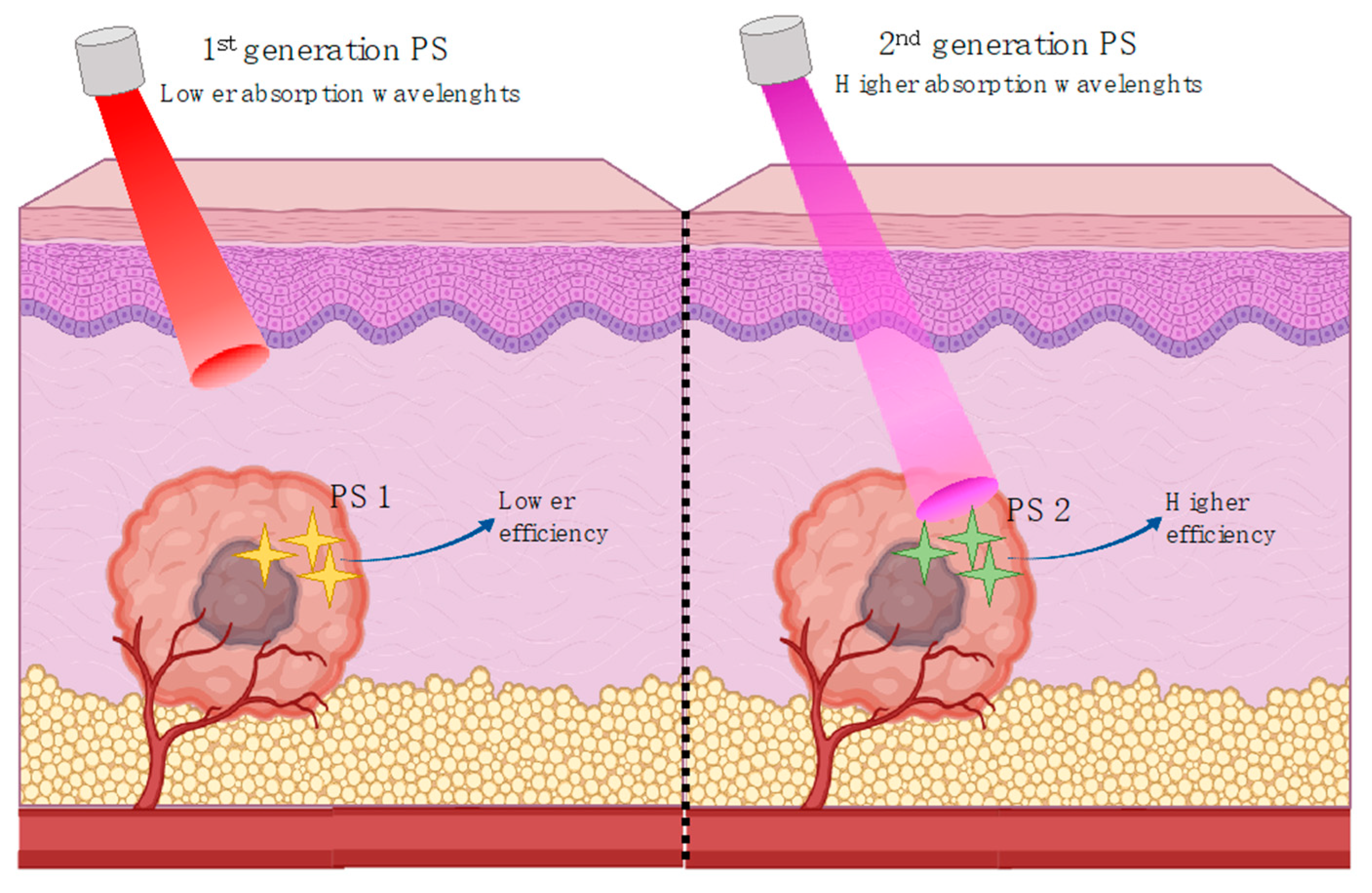
5. Photodynamic Therapy
5.1. Photodynamic Therapy Mechanism
5.2. Photodynamic Therapy Use in Cancer
5.3. The Use in Eye Degenerative Diseases
5.4. Anthocyanins and Their Derivatives, A Potential PDT Ally?
6. The Role of Bioavailability for Anthocyanins Photoprotective Properties
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Glover, B.J.; Martin, C. Anthocyanins. Curr. Biol. 2012, 22, 147–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pina, F.; Oliveira, J.; de Freitas, V. Anthocyanins and derivatives are more than flavylium cations. Tetrahedron 2015, 71, 14. [Google Scholar] [CrossRef]
- Wallace, T.; Giusti, M. Anthocyanins in Health and Disease; CRC Press: New York, NY, USA, 2014; ISBN 978-1-4398-9471-2. [Google Scholar]
- Oliveira, H.; Fernandes, I.; Bras, N.F.; Faria, A.; De Freitas, V.; Calhau, C.; Mateus, N. Experimental and Theoretical Data on the Mechanism by Which Red Wine Anthocyanins Are Transported through a Human MKN-28 Gastric Cell Model. J. Agric. Food Chem. 2015, 63, 7685–7692. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; Perez-Gregorio, R.; De Freitas, V.; Mateus, N.; Fernandes, I. Comparison of the in vitro gastrointestinal bioavailability of acylated and non-acylated anthocyanins: Purple-fleshed sweet potato vs red wine. Food Chem. 2019, 276, 410–418. [Google Scholar] [CrossRef]
- Mendoza, J.; Basílio, N.; Pina, F.; Kondo, T.; Yoshida, K. Rationalizing the Color in Heavenly Blue Anthocyanin: A Complete Kinetic and Thermodynamic Study. J. Phys. Chem. B 2018, 122, 4982–4992. [Google Scholar] [CrossRef]
- Yang, L.; Ling, W.; Du, Z.; Chen, Y.; Li, D.; Deng, S.; Liu, Z.; Yang, L. Effects of Anthocyanins on Cardiometabolic Health: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Adv. Nutr. 2017, 8, 684–693. [Google Scholar] [CrossRef]
- Fernandes, I.; Perez-Gregorio, R.; Soares, S.; Mateus, N.; de Freitas, V. Wine Flavonoids in Health and Disease Prevention. Molecules 2017, 22, 292. [Google Scholar] [CrossRef]
- Gowd, V.; Jia, Z.; Chen, W. Anthocyanins as promising molecules and dietary bioactive components against diabetes—A review of recent advances. Trends Food Sci. Technol. 2017, 68, 1–13. [Google Scholar] [CrossRef]
- MA, Y.; Ding, S.; Fei, Y.; Liu, G.; Jang, H.; Fang, J. Antimicrobial activity of anthocyanins and catechins against foodborne pathogens Escherichia coli and Salmonella. Food Control 2019, 106, 106712. [Google Scholar] [CrossRef]
- Krga, I.; Milenkovic, D. Anthocyanins: From sources and bioavailability to cardiovascular-health benefits and molecular mechanisms of action. J. Agric. Food Chem. 2019, 67, 1771–1783. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.; Liu, Y.; Wang, D.; Liu, J.; Ji, B. The protective effects of berry-derived anthocyanins against visible light-induced damage in human retinal pigment epithelial cells. J. Sci. Food Agric. 2015, 95, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Miyake, S.; Takahashi, N.; Sasaki, M.; Kobayashi, S.; Tsubota, K.; Ozawa, Y. Vision preservation during retinal inflammation by anthocyanin-rich bilberry extract: Cellular and molecular mechanism. Lab. Investig. 2012, 92, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Hara, H. The Involvement of Anthocyanin-Rich Foods in Retinal Damage. In Recent Advances in Polyphenol Research; Yoshida, K., Cheynier, V., Quideau, S., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2016. [Google Scholar] [CrossRef]
- Lyu, S.-Y.; Park, W.-B. Photoprotective Potential of Anthocyanins Isolated from Acanthopanax divaricatus Var. albeofructus Fruits against UV Irradiation in Human Dermal Fibroblast Cells. Biomol. Ther. 2012, 20, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Brouillard, R.; Lang, J. The hemiacetal-cis-chalcone equilibrium of malvidin, a natural anthocyanin. Can. J. Chem. Rev. Can. De Chim. 1990, 68, 755–761. [Google Scholar] [CrossRef]
- Oliveira, H.; Basílio, N.; Pina, F.; Fernandes, I.; De Freitas, V.; Mateus, N. Purple-Fleshed sweet potato acylated anthocyanins: Equilibrium network and photophysical properties. Food Chem. 2019, 288, 386–394. [Google Scholar] [CrossRef]
- Basílio, N.; Pina, F. Chemistry and Photochemistry of Anthocyanins and Related Compounds: A thermodynamic and kinetic approach. Molecules 2016, 21, 1502. [Google Scholar] [CrossRef]
- Pina, F. Chemical applications of anthocyanins and related compounds. A source of bioinspiration. J. Agric. Food Chem. 2014, 62, 6885–6897. [Google Scholar] [CrossRef]
- Pina, F. Anthocyanins and Related Compounds. Detecting the Change of Regime between Rate Control by Hydration or Tautomerization. Dye. Pigment. 2014, 102, 308–314. [Google Scholar] [CrossRef]
- Leydet, Y.; Gavara, R.; Petrov, V.; Diniz, A.; Parola, A.J.; Lima, J.; Pina, F. The effect of self-aggreagtion on the determination of the kinetic and thermodynamic constants of the network of chemical reactions in 3-glucoside anthocyanins. Phytochemistry 2012, 83, 125–135. [Google Scholar] [CrossRef]
- Maçanita, A.L.; Pina, F.; Parola, A.J. Proton transfer in anthocyanins and related flavylium salts. Determination of ground-state rate constants with nanosecond laser flash-photolysis. J. Phys. Chem. A 2002, 106, 1248–1255. [Google Scholar] [CrossRef]
- Dangles, O.; Saito, N.; Brouillard, R. Anthocyanin intramolecular copigment effect. Phytochemistry 1993, 34, 119–124. [Google Scholar] [CrossRef]
- Rein, M. Copigmentation reactions and color stability of berry anthocyanins. Hels. Uniservity Hels. 2005, 10–14. [Google Scholar]
- Liu, Y.; Liu, Y.; Tao, C.; Liu, M.; Pan, Y.; Lv, Z. Effect of temperature and pH on stability of anthocyanin obtained from blueberry. J. Food Meas. Charact. 2018, 12, 1744–1753. [Google Scholar] [CrossRef]
- Chen, C.-C.; Lin, C.L.; Chen, M.-H.; Chiang, P.-Y. Stability and Quality of Anthocyanin in Purple Sweet Potato Extracts. Foods 2019, 8, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.L.; Yu, Y.Q.; Chen, Z.J.; Wen, G.S.; Wei, F.G.; Zheng, Q.; Wang, C.D.; Xiao, X.L. Stability-increasing effects of anthocyanin glycosyl acylation. Food Chem. 2017, 214, 119–128. [Google Scholar] [CrossRef]
- Furtado, P.; Figueiredo, P.; Chaves das Neves, H.; Pina, F. Photochemical and thermal degradation of anthocyanidins. J. Phytochem. Photobiol. A Chem. 1993, 75, 113–118. [Google Scholar] [CrossRef]
- Markakis, P. Anthocyanins as Food Colours. Acad. Pressnew York 1982. [Google Scholar]
- Pang, X.Q.; Zhang, Z.Q.; Duan, X.W.; Ji, Z.L. Influence of pH and active oxygen on the stability of anthocyanins from litchi pericarp. Acta Hortic. 2001, 558, 339–342. [Google Scholar] [CrossRef]
- Mendoza, J.; Basílio, N.; de Freitas, V.; Pina, F. New Procedure To Calculate All Equilibrium Constants in Flavylium Compounds: Application to the Copigmentation of Anthocyanins. Acs Omega 2019, 4, 12058–12070. [Google Scholar] [CrossRef] [Green Version]
- Takeda, K. Blue metal complex pigments involved in blue flower color. Proc. Jpn. Acad. Ser. B Physicial Biol. Sci. 2006, 82, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Tachibana, N.; Kimura, Y.; Ohno, T. Examination of molecular mechanism for the enhanced thermal stability of anthocyanins by metal cations and polysaccharides. Food Chem. 2014, 143, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Yuan, C.; Wang, H.; Han, F.L.; Liu, Y.; Wang, L.; Liu, Y. Stability of Anthocyanins and Their Degradation Products from Cabernet Sauvignon Red Wine under Gastrointestinal pH and Temperature Conditions. Molecules 2018, 23, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, F.; Liu, Y.; Liu, J.; Wang, E. Stability of blueberry anthocyanin, anthocyanidin and pyranoanthocyanidin pigments and their inhibitory effects and mechanisms in human cervical cancer HeLa cells. RSC Adv. 2019, 9, 10842–10853. [Google Scholar] [CrossRef] [Green Version]
- Mateus, N.; Oliveira, J.; Haettich-Motta, M.; De Freitas, V. New Family of Bluish Pyranoanthocyanins. J. Biomed. Biotechnol. 2014, 5, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Castañeda, A.; Pacheco Hernandez, L.; Páez, E.; Rodriguez, J.; Galán-Vidal, C. Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Lima, J.C.; Vautier-Giongo, C.; Lopes, A.; Melo, E.; Quina, F.H.; Maçanita, A.L. Color Stabilization of Anthocyanins: Effect of SDS Micelles on the Acid−Base and Hydration Kinetics of Malvidin 3-Glucoside (Oenin). J. Phys. Chem. A 2002, 106, 5851–5859. [Google Scholar] [CrossRef]
- Pina, F.; Melo, M.J.; Laia, C.A.T.; Parola, A.J.; Lima, J.C. Chemistry and applications of flavylium compounds: A handful of colours. Chem. Soc. Rev. 2012, 41, 869–908. [Google Scholar] [CrossRef]
- Bridle, P.; Timberlake, C.F. Anthocyanins as natural food colours—selected aspects. Food Chem. 1997, 58, 103–109. [Google Scholar] [CrossRef]
- Brouillard, R.; Figueiredo, P.; Elhabiri, M.; Dangles, O. Molecular Interactions of Phenolic Compounds in Relation to the Colour of Fruit and Vegetables; Clarendon Press: Oxford, UK, 1997. [Google Scholar]
- Quina, F.H.; Freitas, A.A.; Maçanita, A.L.; Ferreira da Silva, P.; Lima, J.C. The (photo)chemistry of anthocyanins. In The Spectrum 19; Center for Photochemical Sciences: Bowling Green, OH, USA, 2006; pp. 12–17. [Google Scholar]
- Lima, J.C.; Abreu, I.; Brouillard, R.; Maçanita, A.L. Kinetics of ultra-fast excited state proton transfer from 7-hydroxy-4-methylflavylium chloride to water. Chem. Phys. Lett. 1998, 298, 189–195. [Google Scholar] [CrossRef]
- Freitas, A.A.; Quina, F.H.; Maçanita, A.A.L. Picosecond Dynamics of Proton Transfer of a 7-Hydroxyflavylium Salt in Aqueous–Organic Solvent Mixtures. J. Phys. Chem. A 2011, 115, 10988–10995. [Google Scholar] [CrossRef]
- Freitas, A.A.; Quina, F.H.; Maçanita, A.A.L. Femtosecond and Temperature-Dependent Picosecond Dynamics of Ultrafast Excited-State Proton Transfer in Water–Dioxane Mixtures. J. Phys. Chem. A 2014, 118, 10448–10455. [Google Scholar] [CrossRef] [PubMed]
- Giestas, L.; Yihwa, C.; Lima, J.C.; Vautier-Giongo, C.; Lopes, A.; Maçanita, A.L.; Quina, F.H. The Dynamics of Ultrafast Excited State Proton Transfer in Anionic Micelles. J. Phys. Chem. A 2003, 107, 3263–3269. [Google Scholar] [CrossRef]
- Moreira, P.F.; Giestas, L.; Yihwa, C.; Vautier-Giongo, C.; Quina, F.H.; Maçanita, A.L.; Lima, J.C. Ground- and Excited-State Proton Transfer in Anthocyanins: From Weak Acids to Superphotoacids. J. Phys. Chem. A 2003, 107, 4203–4210. [Google Scholar] [CrossRef]
- Paulo, L.; Freitas, A.A.; Ferreira da Silva, P.; Shimizu, K.; Quina, F.H.; Maçanita, A.L. Novel Ground- and Excited-State Prototropic Reactivity of a Hydroxycarboxyflavylium Salt. J. Phys. Chem. A 2006, 110, 2089–2096. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.O.; Freitas, A.A.; Maçanita, A.L.; Quina, F.H. Chemistry and photochemistry of natural plant pigments: The anthocyanins. J. Phys. Org. Chem. 2016, 29, 594–599. [Google Scholar] [CrossRef]
- Quina, F.H. Chemistry and Photochemistry Inspired by the Colors of Grapes and Red Wines. Available online: https://www.ciencia-e-vinho.com/2018/08/05/chemistry-and-photochemistry-inspired-by-the-colors-of-grapes-and-red-wines/ (accessed on 13 June 2020).
- Pina, F.; Maestri, M.; Balzani, V. Photochromic flavylium compounds as multistate/multifunction molecular-level systems. Chem. Commun. 1999, 2, 107–114. [Google Scholar] [CrossRef]
- Araújo, P.; Mendoza, J.; Pina, F.; Pereira, A.R.; Fernandes, I.; De Freitas, V.; Oliveira, J. Photochemistry of 5-Hydroxy-4’-Dimethylaminoflavylium in the presence of SDS micelles. The role of metastable states of flavylium cation-quinoidal base and trans-chalcones. J. Photochem. Photobiol. A Chem. 2020, 402, 112827. [Google Scholar] [CrossRef]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Characteristics of the Aging Skin. Adv Wound Care (New Rochelle) 2013, 2, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Rodero, S.; Fernández-Morera, J.L.; Menéndez-Torre, E.; Calvanese, V.; Fernández, A.F.; Fraga, M.F. Aging genetics and aging. Aging Dis. 2011, 2, 186–195. [Google Scholar]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Intrinsic and extrinsic factors in skin ageing: A review. Int. J. Cosmet. Sci. 2008, 30, 87–95. [Google Scholar] [CrossRef]
- Shin, J.-W.; Kwon, S.-H.; Choi, J.-Y.; Na, J.-I.; Huh, C.-H.; Choi, H.-R.; Park, K.-C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas-Rodríguez, S.; Folgueras, A.R.; López-Otín, C. The role of matrix metalloproteinases in aging: Tissue remodeling and beyond. Biochim. Et Biophys. Acta (BBA) Mol. Cell Res. 2017, 1864, 2015–2025. [Google Scholar] [CrossRef]
- Parrado, C.; Mercado-Saenz, S.; Perez-Davo, A.; Gilaberte, Y.; Gonzalez, S.; Juarranz, A. Environmental Stressors on Skin Aging. Mechanistic Insights. Front. Pharmacol. 2019, 10, 759. [Google Scholar] [CrossRef] [PubMed]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [Green Version]
- Rinnerthaler, M.; Bischof, J.; Streubel, M.; Trost, A.; Richter, K. Oxidative Stress in Aging Human Skin. Biomolecules 2015, 5, 545–589. [Google Scholar] [CrossRef] [Green Version]
- Rittié, L.; Fisher, G. Natural and Sun-Induced Aging of Human Skin. Cold Spring Harb. Perspect. Med. 2015, 5, a015370. [Google Scholar] [CrossRef]
- Varani, J.; Schuger, L.; Dame, M.K.; Leonard, C.; Fligiel, S.E.G.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Reduced Fibroblast Interaction with Intact Collagen as a Mechanism for Depressed Collagen Synthesis in Photodamaged Skin. J. Investig. Dermatol. 2004, 122, 1471–1479. [Google Scholar] [CrossRef] [Green Version]
- Tewari, A.; Grys, K.; Kollet, J.; Sarkany, R.; Young, A.R. Upregulation of MMP12 and Its Activity by UVA1 in Human Skin: Potential Implications for Photoaging. J. Investig. Dermatol. 2014, 134, 2598–2609. [Google Scholar] [CrossRef] [Green Version]
- Rabe, J.H.; Mamelak, A.J.; McElgunn, P.J.S.; Morison, W.L.; Sauder, D.N. Photoaging: Mechanisms and repair. J. Am. Acad. Dermatol. 2006, 55, 1–19. [Google Scholar] [CrossRef]
- Xu, H.; Zheng, Y.-W.; Liu, Q.; Liu, L.-P.; Luo, F.-L.; Zhou, H.-C.; Isoda, H.; Ohkohchi, N.; Li, Y.-M. Reactive Oxygen Species in Skin Repair, Regeneration, Aging, and Inflammation. In Reactive Oxygen Species (ROS) in Living Cells; Filip, C., Albu, E., Eds.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; He, Y.-Y. Ultraviolet radiation-induced non-melanoma skin cancer: Regulation of DNA damage repair and inflammation. Genes Dis. 2014, 1, 188–198. [Google Scholar] [CrossRef] [Green Version]
- Bosch, R.; Philips, N.; Suárez-Pérez, J.A.; Juarranz, A.; Devmurari, A.; Chalensouk-Khaosaat, J.; González, S. Mechanisms of Photoaging and Cutaneous Photocarcinogenesis, and Photoprotective Strategies with Phytochemicals. Antioxid. 2015, 4, 248–268. [Google Scholar] [CrossRef] [Green Version]
- Fuller, B.B. Antioxidants and Anti-inflammatories. In Cosmetic Dermatology: Products and Procedures, Second; Draelos, Z.D., Ed.; John Wiley & Sons: Hoboken, NJ, USA. [CrossRef]
- Fuller, B. Role of PGE-2 and Other Inflammatory Mediators in Skin Aging and Their Inhibition by Topical Natural Anti-Inflammatories. Cosmetics 2019, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Borg, M.; Brincat, S.; Camilleri, G.; Schembri-Wismayer, P.; Brincat, M.; Calleja-Agius, J. The role of cytokines in skin aging. Climacteric 2013, 16, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Fisher, D.E. MITF and UV responses in skin: From pigmentation to addiction. Pigment Cell Melanoma Res. 2019, 32, 224–236. [Google Scholar] [CrossRef] [Green Version]
- d’Ischia, M.; Wakamatsu, K.; Cicoira, F.; Di Mauro, E.; Garcia-Borron, J.; Commo, S.; Galván, I.; Ghanem, G.; Kenzo, K.; Meredith, P.; et al. Melanin and melanogenesis: From pigment cells to human health and technological applications. Pigment Cell Melanoma Res. 2015, 28, 520–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, J.L. The Genetics of Sun Sensitivity in Humans. Am. J. Hum. Genet. 2004, 75, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Nahhas, A.F.; Abdel-Malek, Z.A.; Kohli, I.; Braunberger, T.L.; Lim, H.W.; Hamzavi, I.H. The potential role of antioxidants in mitigating skin hyperpigmentation resulting from ultraviolet and visible light-induced oxidative stress. Photodermatol. Photoimmunol. Photomed. 2019, 35, 420–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passeron, T.; Picardo, M. Melasma, a photoaging disorder. Pigment Cell Melanoma Res. 2018, 31, 461–465. [Google Scholar] [CrossRef] [Green Version]
- Kovacs, D.; Cardinali, G.; Aspite, N.; Cota, C.; Luzi, F.; Bellei, B.; Briganti, S.; Amantea, A.; Torrisi, M.R.; Picardo, M. Role of fibroblast-derived growth factors in regulating hyperpigmentation of solar lentigo. Br. J. Dermatol. 2010, 163, 1020–1027. [Google Scholar] [CrossRef]
- Rojo, L.; Roopchand, D.; Graf, B.; Cheng, D.; Ribnicky, D.; Fridlender, B.; Raskin, I. Role of Anthocyanins in Skin Aging and UV Induced Skin Damage, and colorectal cancer. In Anthocyanins in Health and Disease; CRC Press: New York, NY, USA, 2013; pp. 307–316. [Google Scholar] [CrossRef]
- Afaq, F.; Syed, D.; Malik, A.; Hadi, N.; Sarfaraz, S.; Kweon, M.-H.; Khan, N.; Zaid, M.; Mukhtar, H. Delphinidin, an Anthocyanidin in Pigmented Fruits and Vegetables, Protects Human HaCaT Keratinocytes and Mouse Skin Against UVB-Mediated Oxidative Stress and Apoptosis. J. Investig. Dermatol. 2007, 127, 222–232. [Google Scholar] [CrossRef] [Green Version]
- Lim, T.-G.; Jung, S.K.; Kim, J.-e.; Kim, Y.; Lee, H.J.; Jang, T.S.; Lee, K.W. NADPH oxidase is a novel target of delphinidin for the inhibition of UVB-induced MMP-1 expression in human dermal fibroblasts. Exp. Dermatol. 2013, 22, 428–430. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.Y.; Lee, K.W.; Kim, J.-E.; Jung, S.K.; Kang, N.J.; Hwang, M.K.; Heo, Y.-S.; Bode, A.M.; Dong, Z.; Lee, H.J. Delphinidin suppresses ultraviolet B-induced cyclooxygenases-2 expression through inhibition of MAPKK4 and PI-3 kinase. Carcinogenesis 2009, 30, 1932–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-E.; Kwon, J.Y.; Seo, S.K.; Son, J.E.; Jung, S.K.; Min, S.Y.; Hwang, M.K.; Heo, Y.-S.; Lee, K.W.; Lee, H.J. Cyanidin suppresses ultraviolet B-induced COX-2 expression in epidermal cells by targeting MKK4, MEK1, and Raf-1. Biochem. Pharmacol. 2010, 79, 1473–1482. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Hu, Y.; Jiang, X.; Chen, T.; Ma, Y.; Wu, S.; Sun, J.; Jiao, R.; Li, X.; Deng, L.; et al. Cyanidin-3-O-glucoside inhibits the UVB-induced ROS/COX-2 pathway in HaCaT cells. J. Photochem. Photobiol. B Biol. 2017, 177, 24–31. [Google Scholar] [CrossRef]
- Rundhaug, J.; Fischer, S. Cyclo-oxygenase-2 Plays a Critical Role in UV-induced Skin Carcinogenesis. Photochem. Photobiol. 2008, 84, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Bucci, P.; Prieto, M.J.; Milla, L.; Calienni, M.N.; Mertinez, L.; Rivarola, V.; Alonso, S.; Montarani, J. Skin penetration and UV-damage prevention by nanoberries. J. Cosmet. Dermatol. 2018, 17, 889–899. [Google Scholar] [CrossRef]
- Bae, J.-Y.; Lim, S.S.; Kim, S.J.; Choi, J.-S.; Park, J.; Ju, S.M.; Han, S.J.; Kang, I.-J.; Kang, Y.-H. Bog blueberry anthocyanins alleviate photoaging in ultraviolet-B irradiation-induced human dermal fibroblasts. Mol. Nutr. Food Res. 2009, 53, 726–738. [Google Scholar] [CrossRef]
- Zhi, Q.; Lei, L.; Li, F.; Zhao, J.; Yin, R.; Ming, J. The anthocyanin extracts from purple-fleshed sweet potato exhibited anti-photoaging effects on ultraviolent B-irradiated BALB/c-nu mouse skin. J. Funct. Foods 2020, 64, 103640. [Google Scholar] [CrossRef]
- Li, A.; Xiao, R.; He, S.; An, X.; He, Y.; Wang, C.; Yin, S.; Wang, B.; Shi, X.; He, J. Research Advances of Purple Sweet Potato Anthocyanins: Extraction, Identification, Stability, Bioactivity, Application, and Biotransformation. Molecules 2019, 24, 3816. [Google Scholar] [CrossRef] [Green Version]
- Harborne, J.B. Spectral methods of characterizing anthocyanins. Biochem. J. 1958, 70, 22–28. [Google Scholar] [CrossRef]
- Silva, P.; Paulo, L.; Barbafina, A.; Eisei, F.; Quina, F.; Macanita, A. Photoprotection and the Photophysics of Acylated Anthocyanins. Chem. Eur. J. 2012, 18, 3736–3744. [Google Scholar] [CrossRef]
- Chan, C.-F.; Lien, C.-Y.; Lai, Y.-C.; Huang, C.-L.; Liao, W. Influence of purple sweet potato extracts on the UV absorption properties of a cosmetic cream. J. Cosmet. Sci. 2010, 61, 333–341. [Google Scholar] [PubMed]
- Cefali, L.C.; Franco, J.G.; Nicolini, G.F.; Ataide, J.A.; Mazzola, P.G. In vitro antioxidant activity and solar protection factor of blackberry and raspberry extracts in topical formulation. J. Cosmet. Dermatol. 2019, 18, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Gasparrini, M.; Forbes-Hernandez, T.Y.; Afrin, S.; Alvarez-Suarez, J.M.; Gonzàlez-Paramàs, A.M.; Santos-Buelga, C.; Bompadre, S.; Quiles, J.L.; Mezzetti, B.; Giampieri, F. A Pilot Study of the Photoprotective Effects of Strawberry-Based Cosmetic Formulations on Human Dermal Fibroblasts. Int. J. Mol. Sci. 2015, 16, 17870–17884. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, T.; Osawa, T. Inhibition of Tyrosinase Activity by the Anthocyanin Pigments Isolated from Phaseolus vulgaris L. Food Sci. Technol. Int. Tokyo 1997, 3, 82–83. [Google Scholar] [CrossRef]
- Xue, Y.-L.; Miyakawa, T.; Hayashi, Y.; Okamoto, K.; Hu, F.; Mitani, N.; Furihata, K.; Sawano, Y.; Tanokura, M. Isolation and Tyrosinase Inhibitory Effects of Polyphenols from the Leaves of Persimmon, Diospyros kaki. J. Agric. Food Chem. 2011, 59, 6011–6017. [Google Scholar] [CrossRef]
- Jhan, J.K.; Chung, Y.C.; Chen, G.H.; Chang, C.H.; Lu, Y.C.; Hsu, C.K. Anthocyanin contents in the seed coat of black soya bean and their anti-human tyrosinase activity and antioxidative activity. Int. J. Cosmet. Sci. 2016, 38, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Karunarathne, W.; Molagoda, N.; Park, S.; Kim, J.; Lee, O.-K.; Kwon, H.; Oren, M.; Choi, Y.; Ryu, H.W.; Oh, S.-R.; et al. Anthocyanins from Hibiscus syriacus L. Inhibit Melanogenesis by Activating the ERK Signaling Pathway. Biomolecules 2019, 9, 645. [Google Scholar] [CrossRef] [Green Version]
- Yasumoto, K.; Yokoyama, K.; Shibata, K.; Tomita, Y.; Shibahara, S. Microphthalmia-associated transcription factor as a regulator for melanocyte-specific transcription of the human tyrosinase gene. Mol. Cell. Biol. 1994, 14, 8058. [Google Scholar] [CrossRef]
- Santiago, A.R.; Boia, R.; Aires, I.D.; Ambrósio, A.F.; Fernandes, R. Sweet Stress: Coping With Vascular Dysfunction in Diabetic Retinopathy. Front. Physiol. 2018, 9, 820. [Google Scholar] [CrossRef]
- Aires, I.D.; Boia, R.; Rodrigues-Neves, A.C.; Madeira, M.H.; Marques, C.; Ambrósio, A.F.; Santiago, A.R. Blockade of microglial adenosine A2A receptor suppresses elevated pressure-induced inflammation, oxidative stress, and cell death in retinal cells. Glia 2019, 67, 896–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Organisciak, D.T.; Vaughan, D.K. Retinal light damage: Mechanisms and protection. Prog. Retin. Eye Res. 2010, 29, 113–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, J.; Morgan, J.; Merigan, W.; Sliney, D.; Sparrow, J.; Williams, D. The susceptibility of the retina to photochemical damage from visible light. Prog. Retin. Eye Res. 2011, 31, 28–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belleoud, L.; Leluan, D.; Boyer, Y. Study on the effects of anthocyanin glycosides on the nocturnal vision of air traffic controllers. Rev. Med. Aeronaut Spat. 1966, 18, 3–7. [Google Scholar]
- Jang, Y.; Zhou, J.; Nakanishi, K.; Sparrow, J. Anthocyanins Protect Against A2E Photooxidation and Membrane Permeabilization in Retinal Pigment Epithelial Cells. Photochem. Photobiol. 2005, 81, 529–536. [Google Scholar] [CrossRef]
- Milbury, P.E.; Graf, B.; Curran-Celentano, J.M.; Blumberg, J.B. Bilberry (Vaccinium myrtillus) anthocyanins modulate heme oxygenase-1 and glutathione S-transferase-pi expression in ARPE-19 cells. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2343–2349. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, J.; Kadekaru, T.; Ogawa, K.; Hitoe, S.; Shimoda, H.; Hara, H. Maqui berry (Aristotelia chilensis) and the constituent delphinidin glycoside inhibit photoreceptor cell death induced by visible light. Food Chem. 2013, 139, 129–137. [Google Scholar] [CrossRef]
- Huang, W.; Yan, Z.; Li, D.; Ma, Y.; Zhou, J.; Sui, Z. Antioxidant and Anti-Inflammatory Effects of Blueberry Anthocyanins on High Glucose-Induced Human Retinal Capillary Endothelial Cells. Oxidative Med. Cell. Longev. 2018, 2018, 1862462. [Google Scholar] [CrossRef]
- Song, Y.; Huang, L.; Yu, J. Effects of blueberry anthocyanins on retinal oxidative stress and inflammation in diabetes through Nrf2/HO-1 signaling. J. Neuroimmunol. 2016, 301, 1–6. [Google Scholar] [CrossRef]
- Kim, J.; Kim, C.S.; Lee, Y.M.; Sohn, E.; Jo, K.; Kim, J.S. Vaccinium myrtillus extract prevents or delays the onset of diabetes--induced blood-retinal barrier breakdown. Int. J. Food Sci. Nutr. 2015, 66, 236–242. [Google Scholar] [CrossRef]
- Paik, S.S.; Jeong, E.; Jung, S.W.; Ha, T.J.; Kang, S.; Sim, S.; Jeon, J.H.; Chun, M.H.; Kim, I.B. Anthocyanins from the seed coat of black soybean reduce retinal degeneration induced by N-methyl-N-nitrosourea. Exp. Eye Res. 2012, 97, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Kalt, W.; Hanneken, A.; Milbury, P.; Tremblay, F. Recent Research on Polyphenolics in Vision and Eye Health. J. Agric. Food Chem. 2010, 58, 4001–4007. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, F.; Waterhouse, J.; Nason, J.; Kalt, W. Prophylactic neuroprotection by blueberry-enriched diet in a rat model of light-induced retinopathy. J. Nutr. Biochem. 2013, 24, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.; Bras, N.F.; Mateus, N.; de Freitas, V. Understanding the molecular mechanism of anthocyanin binding to pectin. Langmuir 2014, 30, 8516–8527. [Google Scholar] [CrossRef]
- Marques, C.; Fernandes, I.; Norberto, S.; Sa, C.; Teixeira, D.; de Freitas, V.; Mateus, N.; Calhau, C.; Faria, A. Pharmacokinetics of blackberry anthocyanins consumed with or without ethanol: A randomized and crossover trial. Mol. Nutr. Food Res. 2016, 60, 2319–2330. [Google Scholar] [CrossRef]
- Czank, C.; Cassidy, A.; Zhang, Q.; Morrison, D.J.; Preston, T.; Kroon, P.A.; Botting, N.P.; Kay, C.D. Human metabolism and elimination of the anthocyanin, cyanidin-3-glucoside: A (13)C-tracer study. Am. J. Clin. Nutr. 2013, 97, 995–1003. [Google Scholar] [CrossRef] [Green Version]
- Aura, A.M.; Martin-Lopez, P.; O’Leary, K.A.; Williamson, G.; Oksman-Caldentey, K.M.; Poutanen, K.; Santos-Buelga, C. In vitro metabolism of anthocyanins by human gut microflora. Eur. J. Nutr. 2005, 44, 133–142. [Google Scholar] [CrossRef]
- Keppler, K.; Humpf, H.U. Metabolism of anthocyanins and their phenolic degradation products by the intestinal microflora. Bioorganic Med. Chem. 2005, 13, 5195–5205. [Google Scholar] [CrossRef]
- Nomi, Y.; Iwasaki-Kurashige, K.; Matsumoto, H. Therapeutic Effects of Anthocyanins for Vision and Eye Health. Molecules 2019, 24, 3311. [Google Scholar] [CrossRef] [Green Version]
- Kalt, W.; Blumberg, J.B.; McDonald, J.E.; Vinqvist-Tymchuk, M.R.; Fillmore, S.A.; Graf, B.A.; O’Leary, J.M.; Milbury, P.E. Identification of anthocyanins in the liver, eye, and brain of blueberry-fed pigs. J. Agric. Food Chem. 2008, 56, 705–712. [Google Scholar] [CrossRef]
- Oliveira, H.; Roma-Rodrigues, C.; Santos, A.; Veigas, B.; Brás, N.; Faria, A.; Calhau, C.; de Freitas, V.; Baptista, P.V.; Mateus, N.; et al. GLUT1 and GLUT3 involvement in anthocyanin gastric transport- Nanobased targeted approach. Sci. Rep. 2019, 9, 789. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, I.; Marques, C.; Évora, A.; Cruz, L.; de Freitas, V.; Calhau, C.; Faria, A.; Mateus, N. Pharmacokinetics of table and Port red wine anthocyanins: A crossover trial in healthy men. Food Funct. 2017, 8, 2030–2037. [Google Scholar] [CrossRef]
- Han, F.; Oliveira, H.; Brás, N.F.; Fernandes, I.; Cruz, L.; De Freitas, V.; Mateus, N. In vitro gastrointestinal absorption of red wine anthocyanins—Impact of structural complexity and phase II metabolization. Food Chem. 2020, 317, 126398. [Google Scholar] [CrossRef] [PubMed]
- Robertson, C.A.; Evans, D.H.; Abrahamse, H. Photodynamic therapy (PDT): A short review on cellular mechanisms and cancer research applications for PDT. J. Photochem. Photobiol. B Biol. 2009, 96, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Stepniowska, A.; Pergol, P.; Padzik-Graczyk, A. Photodynamic method of cancer diagnosis and therapy--mechanisms and applications. Postepy Biochem. 2015, 59, 53–63. [Google Scholar]
- Luksiene, Z. Photodynamic therapy: Mechanism of action and ways to improve the efficiency of treatment. Med. Kaunas, Lith. 2003, 39, 1137–1150. [Google Scholar]
- Li, X.; Lee, S.; Yoon, J. Supramolecular photosensitizers rejuvenate photodynamic therapy. Chem. Soc. Rev. 2018, 47, 1174–1188. [Google Scholar] [CrossRef]
- Cruz, P.M.; Mo, H.; McConathy, W.J.; Sabnis, N.; Lacko, A.G. The role of cholesterol metabolism and cholesterol transport in carcinogenesis: A review of scientific findings, relevant to future cancer therapeutics. Front. Pharmacol. 2013, 25, 119. [Google Scholar] [CrossRef] [Green Version]
- Castano, A.P.; Demidova, T.N.; Hamblin, M.R. Mechanisms in photodynamic therapy: Part three-Photosensitizer pharmacokinetics, biodistribution, tumor localization and modes of tumor destruction. Photodiagnosis Photodyn. Ther. 2005, 2, 91–106. [Google Scholar] [CrossRef] [Green Version]
- Bacellar, I.O.L.; Oliveira, M.C.; Dantas, L.S.; Costa, E.B.; Junqueira, H.C.; Martins, W.L.; Durantini, A.M.; Cosa, G.; Di Mascio, P.; Wainwright, M.; et al. Photosensitized Membrane Permeabilization Requires ContactDependent Reactions between Photosensitizer and Lipids. J. Am. Chem. Soc. 2018, 140, 9606–9615. [Google Scholar] [CrossRef]
- Jensen, T.J.; Vicente, M.G.; Luguya, R.; Norton, J.; Fronczek, F.R.; Smith, K.M. Effect of overall charge and charge distribution on cellular uptake, distribution and phototoxicity of cationic porphyrins in HEp2 cells. J. Photochem. Photobiol. B Biol. 2010, 100, 100–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrahamse, H.; Hamblin, M.R. New photosensitizers for photodynamic therapy. Biochem. J. 2016, 473, 347–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magda, D.; Miller, R.A. Motexafin gadolinium: A novel redox active drug for cancer therapy. Semin. Cancer Biol. 2006, 16, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Trachtenberg, J.; Weersink, R.A.; Davidson, S.R.; Haider, M.A.; Bogaards, A.; Gertner, M.R.; Evans, A.; Scherz, A.; Savrad, J.; Chin, J.L.; et al. Vascular-targeted photodynamic therapy (padoporfin, WST09) for recurrent prostate cancer after failure of external beam radiotherapy: A study of escalating light doses. Bju Int. 2008, 102, 556–562. [Google Scholar] [CrossRef] [PubMed]
- de Moraes, M.; de Vasconcelos, R.C.; Longo, J.P.; Muehlmann, L.A.; de Azevedo, R.B.; Lemos, T.M.; Costa Ade, L. Effects of photodynamic therapy mediated by nanoemulsion containing chloro-aluminum phthalocyanine: A histologic and immunohistochemical study in human gingiva. Photodiagnosis Photodyn. Ther. 2015, 12, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D.; Baron, E.D.; Scull, H.; Hsia, A.; Berlin, J.C.; McCormick, T.; Colussi, V.; Kenney, M.E.; Cooper, K.D.; Oleinick, N.L. Photodynamic therapy with the phthalocyanine photosensitizer Pc 4: The case experience with preclinical mechanistic and early clinical-translational studies. Toxicol. Appl. Pharmacol. 2007, 224, 290–299. [Google Scholar] [CrossRef] [Green Version]
- Graciano, T.B.; Coutinho, T.S.; Cressoni, C.B.; Freitas, C.P.; Pierre, M.B.; Pereira, S.A.; Shimano, M.M.; Frange, R.C.; Garcia, M.T. Using chitosan gels as a toluidine blue O delivery system for photodynamic therapy of buccal cancer: In vitro and in vivo studies. Photodiagnosis Photodyn. Ther. 2015, 12, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Delaey, E.; Van Laar, F.; De Vos, D.; Kamuhabwa, A.; Jacobs, P.; De Witte, P. A comparative study of the photosensitizing characteristics of some cyanine dyes. J. Photochem. Photobiol. B Biol. 2000, 55, 27–36. [Google Scholar] [CrossRef]
- Lim, S.H.; Thivierge, C.; Nowak-Sliwinska, P.; Han, J.; Van den Bergh, H.; Wagnières, G.; Burgess, K.; Lee, H.B. In Vitro and In Vivo Photocytotoxicity of Boron Dipyrromethene Derivatives for Photodynamic Therapy. J. Med. Chem. 2010, 53, 2865–2874. [Google Scholar] [CrossRef]
- Garg, A.D.; Krysko, D.V.; Vandenabeele, P.; Agostinis, P. Hypericin-based photodynamic therapy induces surface exposure of damage-associated molecular patterns like HSP70 and calreticulin. Cancer Immunol. Immunother. 2012, 61, 215–221. [Google Scholar] [CrossRef]
- Panzarini, E.; Inguscio, V.; Fimia, G.M.; Dini, L. Rose bengal acetate photodynamic therapy (RBAc-PDT) induces exposure and release of damage-associated molecular patterns (DAMPs) in human HeLa cells. PLoS ONE 2014, 9, e105778. [Google Scholar] [CrossRef] [PubMed]
- Cantisani, C.; Paolino, G.; Faina, V.; Frascani, F.; Cantoresi, F.; Bianchini, D.; Fazia, G.; Calvieri, S. Overview on Topical 5-ALA Photodynamic Therapy Use for Non Melanoma Skin Cancers. Int. J. Photoenergy 2014, 304862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, R.; Lange, N.; Guy, R.; Bentley, M. Photodynamic therapy of skin cancer: Controlled drug delivery of 5-ALA and its esters. Adv. Drug Deliv. Rev. 2004, 56, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Chudy, M.; Tokarska, K.; Jastrzebska, E.; Bulka, M.; Drozdek, S.; Lamch, L.; Wilk, K.; Brzózka, Z. Lab-on-a-chip systems for photodynamic therapy investigations. Biosens. Bioelectron. 2018, 101, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Ujiie, H.; Ding, L.; Fan, R.; Kato, T.; Lee, D.; Fujino, K.; Kinoshita, T.; Lee, C.; Waddell, T.; Keshavjee, S.; et al. Porphyrin–High-Density Lipoprotein: A Novel Photosensitizing Nanoparticle for Lung Cancer Therapy. Ann. Thorac. Surg. 2019, 107, 369–377. [Google Scholar] [CrossRef]
- Dougherty, T.J.; Grindey, G.B.; Fiel, R.; Wieshaupt, K.R.; Boyle, D.G. Photoradiation therapy. II. Cure of animal tumors with hematoporphyrin and light. J. Natl. Cancer Inst. 1975, 55, 115–121. [Google Scholar] [CrossRef]
- Banerjee, S.; El-Sheikh, S.; Malhotra, A.; Mosse, C.; Parker, S.; Williams, N.; MacRobert, A.; Hamoudi, R.; Bown, S.; Keshtgar, M. Photodynamic Therapy in Primary Breast Cancer. J. Clin. Med. 2020, 9, 483. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Li, J.; Chen, W.; Liu, L.; Yu, F. Light and sound to trigger the Pandora’s box against breast cancer: A combination strategy of sonodynamic, photodynamic and photothermal therapies. Biomaterials 2020, 232, 119685. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, X.; Li, H.; Li, C.X.; Ding, H.; Zhang, M.; Guo, Y.; Sun, M. Near-infrared light triggered liposomes combining photodynamic and chemotherapy for synergistic breast tumor therapy. Colloids Surf. B Biointerfaces 2019, 173, 564–570. [Google Scholar] [CrossRef]
- Phuong, P.; Lee, S.; Lee, C.; Seo, B.; Park, S.; Oh, K.; Lee, E.S.; Choi, H.-G.; Shin, B.S.; Youn, Y.S. Beta-carotene-bound albumin nanoparticles modified with chlorin e6 for breast tumor ablation based on photodynamic therapy. Colloids Surf. B Biointerfaces 2018, 171, 123–133. [Google Scholar] [CrossRef]
- Usuda, J.; Kato, H.; Okunaka, T.; Furukawa, K.; Tsutsui, H.; Yamada, K.; Suga, Y.; Honda, H.; Nagatsuka, Y.; Ohira, T.; et al. Photodynamic Therapy (PDT) for Lung Cancers. J. Thorac. Oncol. 2006, 1, 489–493. [Google Scholar] [CrossRef] [Green Version]
- Maziak, D.; Markman, B.; MacKay, J.; Evans, W. Photodynamic therapy in nonsmall cell lung cancer: A systematic review. Ann. Thorac. Surg. 2004, 77, 1484–1491. [Google Scholar] [CrossRef] [PubMed]
- Mokwena; Kruger, C.; Ivan, M.-T.; Heidi, A. A review of nanoparticle photosensitizer drug delivery uptake systems for photodynamic treatment of lung cancer. Photodiagnosis Photodyn. Ther. 2018, 22, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Pramual, S.; Lirdprapamongkol, K.; Juouan-Hureaux, V.; Barberi-Heyob, M.; Frochot, C.; Svasti, J.; Niamsiri, N. Overcoming the diverse mechanisms of multidrug resistance in lung cancer cells by photodynamic therapy using pTHPP-loaded PLGA-lipid hybrid nanoparticles. Eur. J. Pharm. Biopharm. 2020, 149, 218–228. [Google Scholar] [CrossRef]
- Usuda, J.; Inoue, T.; Tsuchida, T.; Ohtani, K.; Maehara, S.; Ikeda, N.; Ohsaki, Y.; Sasaki, T.; Oka, K. Clinical trial of photodynamic therapy for peripheral-type lung cancers using a new laser device in a pilot study. Photodiagnosis Photodyn. Ther. 2020, 30. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, J.; Zhang, Y.; Ke, L.; Lin, X.; Li, Z.; Chen, H.; Gao, Y. Indocyanine green-encapsulated erlotinib modified chitosan nanoparticles for targeted chemo photodynamic therapy of lung cancer cells. Dye. Pigment. 2019, 170, 107588. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Q.; Zhang, A.; Pan, S.; Cheng, J.; Zhi, X.; Ding, X.; Hong, L.; Zi, M.; Cui, D.; et al. Multifunctional co-loaded magnetic nanocapsules for enhancing targeted MR imaging and in vivo photodynamic therapy. Nanomed. Nanotechnol. Biol. Med. 2019, 21, 102047. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.-Y.; Yu, X.-H.; Wang, K.; Yin, Y.-J.; Tang, Y.-J.; Tang, Y.-L.; Liang, X.-H. Graphene quantum dots (GQDs)-based nanomaterials for improving photodynamic therapy in cancer treatment. Eur. J. Med. Chem. 2019, 182, 111620. [Google Scholar] [CrossRef]
- da Silva, L.; Núnez-Montenegro, A.; Magalhães, C.; Ferreira, P.; Duarte, D.; González-Berdullas, P.; Rodríguez-Borges, J.; Vale, N.; da Silva, J. Single-molecule chemiluminescent photosensitizer for a self-activating and tumor-selective photodynamic therapy of cancer. Eur. J. Med. Chem. 2019, 183, 111683. [Google Scholar] [CrossRef] [PubMed]
- Shah, Z.; Nazir, S.; Mazhar, K.; Abbasi, R.; Samokhvalov, I. PEGylated doped- and undoped-TiO2 nanoparticles for photodynamic Therapy of cancers. Photodiagnosis Photodyn. Ther. 2019, 27, 173–183. [Google Scholar] [CrossRef]
- Hirao, M.; Hosui, A.; Mimura, A.; Tanimoto, T.; Ohnishi, K.; Kusumoto, Y.; Yamaguchi, T.; Yamada, T.; Miwa, H.; Hiramatsu, N. Significance of in vitro photodynamic cytodiagnosis using 5-aminolevulinic acid in solid pancreatic tumors extracted by endoscopic ultrasound-guided fine-needle aspiration. Photodiagnosis Photodyn. Ther. 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Liu, L.; Liang, R.; Zhou, H.; Pan, H.; Zhang, S.; Cai, L. Tumor-targeted nanoplatform for in situ oxygenation-boosted immunogenic phototherapy of colorectal cancer. Acta Biomater. 2020, 104, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Bretin, L.; Pinon, A.; Bouramtane, S.; Ouk, C.; Richard, L.; Chaunavel, A.; Carrion, C.; Bregier, F.; Sol, V.; Chaleix, V.; et al. Photodynamic Therapy Activity of New Porphyrin-Xylan-Coated Silica Nanoparticles in Human Colorectal Cancer. Cancers 2019, 11, 1474. [Google Scholar] [CrossRef] [Green Version]
- Osman, H.; Elsahy, D.; Saadatzadeh, M.R.; Pollok, K.E.; Yocom, S.; Hattab, E.M.; Georges, J.; Cohen-Gadol, A.A. Acridine Orange as a Novel Photosensitizer for Photodynamic Therapy in Glioblastoma. World Neurosurg. 2018, 114, e1310–e1315. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Cho, H.; Sun, Q.; He, Y.; Ma, G.; Kim, Y.; Kim, B.; Kim, O. Acanthopanacis Cortex extract: A novel photosensitizer for head and neck squamous cell carcinoma therapy. Photodiagnosis Photodyn. Ther. 2019, 26, 142–149. [Google Scholar] [CrossRef]
- Zheng, B.-Y.; Wang, L.; Hu, Q.-Y.; Shi, J.; Ke, M.-R.; Huang, J.-D. Novel unsymmetrical silicon(IV) phthalocyanines as highly potent anticancer photosensitizers. Synthesis, characterization, and in vitro photodynamic activities. Dye. Pigment. 2020, 177, 108286. [Google Scholar] [CrossRef]
- Su, W.; Luo, Z.; Dong, S.; Chen, X.; Xiao, J.; Peng, B.; Li, P. Novel half-sandwich rhodium(III) and iridium(III) photosensitizers for dual chemo- and photodynamic therapy. Photodiagnosis Photodyn. Ther. 2019, 26, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.-H.; Zhu, X.-X.; Zhu, W.; Wu, D.; Chen, D.-Y.; Yan, Y.-J.; Wu, X.-F.; O’Shea, D.; Chen, Z.-L. Synthesis and evaluation of novel chlorophyll a derivatives as potent photosensitizers for photodynamic therapy. Eur. J. Med. Chem. 2020, 187, 111959. [Google Scholar] [CrossRef]
- Jalde, S.; Chauhan, A.; Lee, J.; Chaturvedi, P.; Park, J.; Kim, Y.-W. Synthesis of novel Chlorin e6-curcumin conjugates as photosensitizers for photodynamic therapy against pancreatic carcinoma. Eur. J. Med. Chem. 2018, 147, 66–76. [Google Scholar] [CrossRef]
- Warowicka, A.; Popenda, L.; Bartkowiak, G.; Musidlak, O.; Litowczenko-Cybulska, J.; Kuzma, D.; Nawrot, R.; Jurga, S.; Gozdzicka-Józefiak, A. Protoberberine compounds extracted from Chelidonium majus L. as novel natural photosensitizers for cancer therapy. Phytomedicine 2019, 64, 152919. [Google Scholar] [CrossRef]
- Swamy, P.; Sivaraman, G.; Priyanka, R.; Raja, S.; Ponnuvel, K.; Shanmugpriya, J.; Gulyani, A. Near Infrared (NIR) absorbing dyes as promising photosensitizer for photo dynamic therapy. Coord. Chem. Rev. 2020, 411, 213233. [Google Scholar] [CrossRef]
- Boyar, C.; Çamur, M. Novel water soluble 7-oxy-4-(pyridine-3-yl)coumarin substituted phthalocyanines as potential photosensitizers for photodynamic therapy. Inorg. Chim. Acta 2019, 494, 30–41. [Google Scholar] [CrossRef]
- Xiao, Q.; Wu, J.; Jiang, Y.; Wang, P.; Leung, A.W.; Gao, L.; Jiang, S.; Xu, C. Discovery and Development of Natural Products and their Derivatives as Photosensitizers for Photodynamic Therapy. Curr. Med. Chem. 2018, 25, 839–860. [Google Scholar] [CrossRef] [PubMed]
- Yadav, N.K.; Jayadev, C.; Rajendran, A.; Nagpal, M. Recent developments in retinal lasers and delivery systems. Indian J. Ophthalmol. 2014, 62, 50–54. [Google Scholar] [CrossRef]
- Park, J.J.; Pavesio, C. Prophylactic laser photocoagulation for acute retinal necrosis. Does it raise more questions than answers? Br. J. Ophthalmol. 2008, 92, 1161–1162. [Google Scholar] [CrossRef]
- Palanker, D.; Blumenkranz, M.S. Chapter 39—Retinal Laser Therapy: Biophysical Basis and Applications. In Retina, 5th ed.; Ryan, S.J., Sadda, S.R., Hinton, D.R., Schachat, A.P., Sadda, S.R., Wilkinson, C.P., Wiedemann, P., Schachat, A.P., Eds.; W.B. Saunders: London, UK, 2013; pp. 746–760. [Google Scholar] [CrossRef]
- Ammar, D.A.; Kahook, M.Y. In vitro effects of verteporfin on ocular cells. Mol. Vis. 2013, 19, 424–429. [Google Scholar]
- Yoshida, K.; Mori, M.; Kondo, T. Blue flower color development by anthocyanins: From chemical structure to cell physiology. Nat. Prod. Rep. 2009, 26, 884–915. [Google Scholar] [CrossRef]
- Fernando, R.; Senadeera, G.K.R. Natural anthocyanins as photosensitizers for dye-sensitized solar devices. Curr. Sci. 2008, 95, 663–666. [Google Scholar]
- Chien, C.-Y.; Hsu, B.-D. Optimization of the dye-sensitized solar cell with anthocyanin as photosensitizer. Sol. Energy 2013, 98, 203–211. [Google Scholar] [CrossRef]
- Dhafina, W.; Daud, M.; Salleh, H. The sensitization effect of anthocyanin and chlorophyll dyes on optical and photovoltaic properties of zinc oxide based dye-sensitized solar cells. Optik 2020, 207, 163808. [Google Scholar] [CrossRef]
- Pinto, A.L.; Cruz, H.; Oliveira, J.; Araújo, P.; Cruz, L.; Gomes, V.; Silva, C.P.; Silva, G.T.M.; Mateus, T.; Calogero, G.; et al. Dye-sensitized solar cells based on dimethylamino-π-bridge-pyranoanthocyanin dyes. Sol. Energy 2020, 206, 188–199. [Google Scholar] [CrossRef]
- Silva, G.T.M.; Thomas, S.S.; Silva, C.P.; Schlothauer, J.C.; Baptista, M.S.; Freitas, A.A.; Bohne, C.; Quina, F.H. Triplet Excited States and Singlet Oxygen Production by Analogs of Red Wine Pyranoanthocyanins. Photochem. Photobiol. 2019, 95, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Teerakapong, A.; Damrongrungruang, T.; Sattayut, S.; Morales, N.; Tantananugool, S. Efficacy of erythrosine and cyanidin-3-glucoside mediated photodynamic therapy on Porphyromonas gingivalis biofilms using green light laser. Photodiagnosis Photodyn. Ther. 2017, 20, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Laszló, I.; Laszló, M.; Toma, V.; Baldea, I.; Olteanu, D.; David, L.; Moldovan, B.; Ion, R.; Moldovan, R.; Filip, G.; et al. The in vivo modulatory effects of Cornus mas extract on photodynamic therapy in experimental tumors. Photodiagnosis Photodyn. Ther. 2020, 30, 101656. [Google Scholar] [CrossRef] [PubMed]
- Fatima, K.; Masood, N.; Luqman, S. Quenching of singlet oxygen by natural and synthetic antioxidants and assessment of electronic UV/Visible absorption spectra for alleviating or enhancing the efficacy of photodynamic therapy. Biomed. Reserach Ther. 2016, 3, 514–527. [Google Scholar] [CrossRef] [Green Version]
- De Rosso, V.V.; Vieyra, F.E.M.; Mercadante, A.Z.; Borsarelli, C.D. Singlet oxygen quenching by anthocyanin’s flavylium cations. Free Radic. Res. 2008, 42, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Yoon, S.H.; Jung, M.; Choe, E. Stability of meoru (Vitis coignetiea) anthocyanins under photochemically produced singlet oxygen by riboflavin. New Biotechnol. 2010, 27, 435–439. [Google Scholar] [CrossRef]
- Braga, A.; Murador, D.; de Souza, L.; De Rosso, V.V. Bioavailability of anthocyanins: Gaps in knowledge, challenges and future research. J. Food Compos. Anal. 2018, 68, 31–40. [Google Scholar] [CrossRef]
- Rodriguez-Mateos, A.; Istas, G.; Boschek, L.; Feliciano, R.; Mills, C.; Boby, C.; Gomez-Alonso, S.; Milenkovic, D.; Heiss, C. Circulating Anthocyanin Metabolites Mediate Vascular Benefits of Blueberries: Insights From Randomized Controlled Trials, Metabolomics, and Nutrigenomics. J. Gerontol. Biol. Sci. 2019, 74, 967–976. [Google Scholar] [CrossRef]
- Istas, G.; Feliciano, R.; Weber, T.; Garcia-Villalba, R.; Tomas-Barberan, F.; Heiss, C.; Rodriguez-Mateos, A. Plasma urolithin metabolites correlate with improvements in endothelial function after red raspberry consumption: A double-blind randomized controlled trial. Arch. Biochem. Biophys. 2018, 651, 43–51. [Google Scholar] [CrossRef]
- Passamonti, S.; Vrhovsek, U.; Mattivi, F. The interaction of anthocyanins with bilitranslocase. Biochem. Biophys. Res. Commun. 2002, 296, 631–636. [Google Scholar] [CrossRef]
- Passamonti, S.; Vanzo, A. Hepatic uptake of grape anthocyanins and the role of bilitranslocase. Food Chem. Toxicolgy 2005, 38, 953–960. [Google Scholar] [CrossRef]
- Faria, A.; Pestana, D.; Azevedo, J.; Martel, F.; de Freitas, V.; Azevedo, I.; Mateus, N.; Calhau, C. Absorption of anthocyanins through intestinal epithelial cells—Putative involvement of GLUT2. Mol. Nutr. Food Res. 2009, 53, 7. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; Fernandes, A.; Natércia, B.; Mateus, N.; De Freitas, V.; Fernandes, I. Anthocyanins as Antidiabetic Agents—In Vitro and In Silico Approaches of Preventive and Therapeutic Effects. Molecules 2020, 25, 3813. [Google Scholar] [CrossRef] [PubMed]
- Priprem, A.; Damrongrungruang, T.; Limsitthichaikoon, S.; Khampaenjiraroch, B.; Nukulkit, C.; Thapphasaraphong, S.; Limphirat, W. Topical Niosome Gel Containing an Anthocyanin Complex: A Potential Oral Wound Healing in Rats. AAPS PharmSciTech 2018, 19, 1681–1692. [Google Scholar] [CrossRef] [PubMed]
- Bojanowski, K. Hypodermal delivery of cosmetic actives for improved facial skin morphology and functionality. Int. J. Cosmet. Sci. 2013, 35, 562–567. [Google Scholar] [CrossRef]







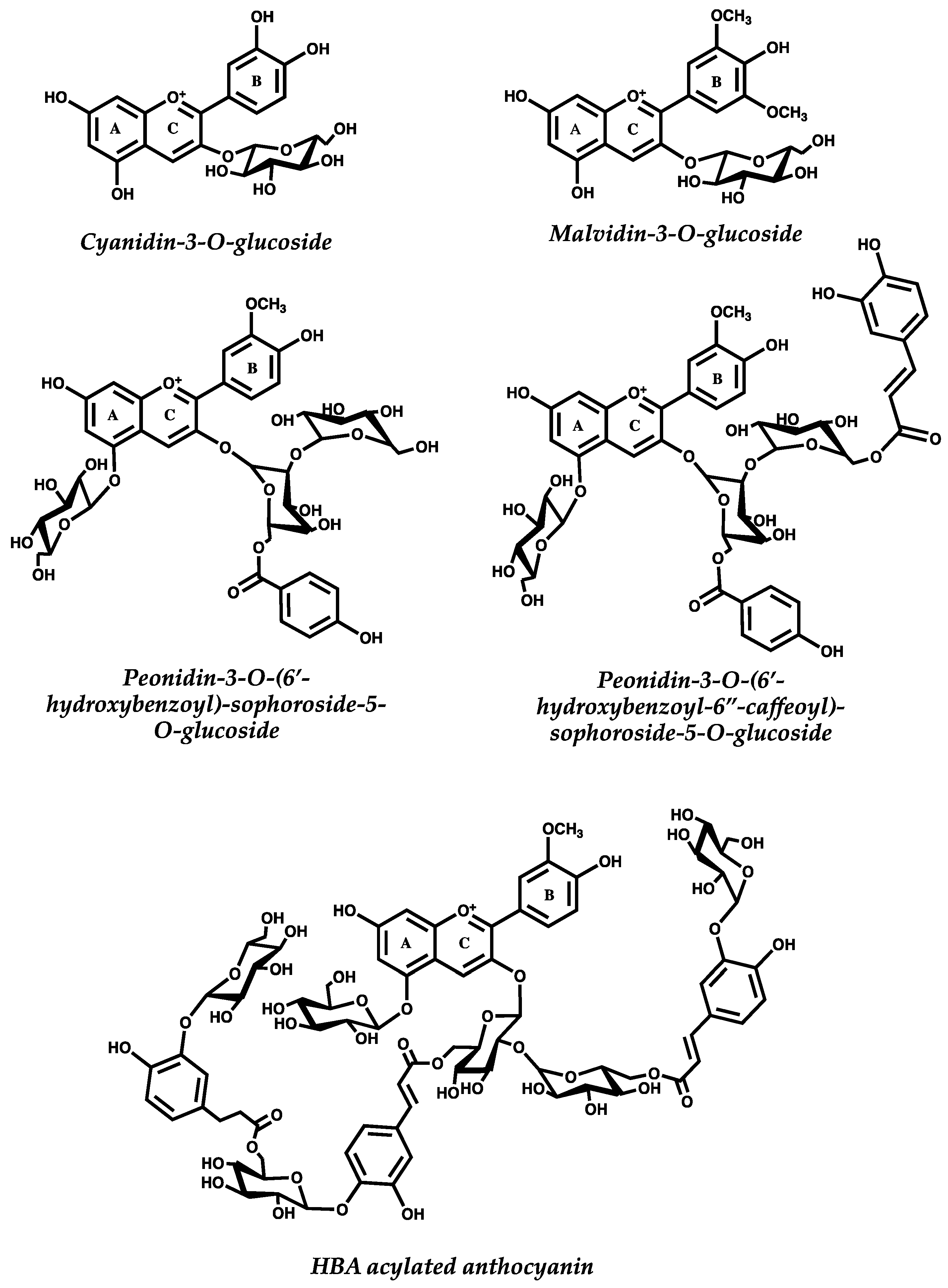{kind=link}
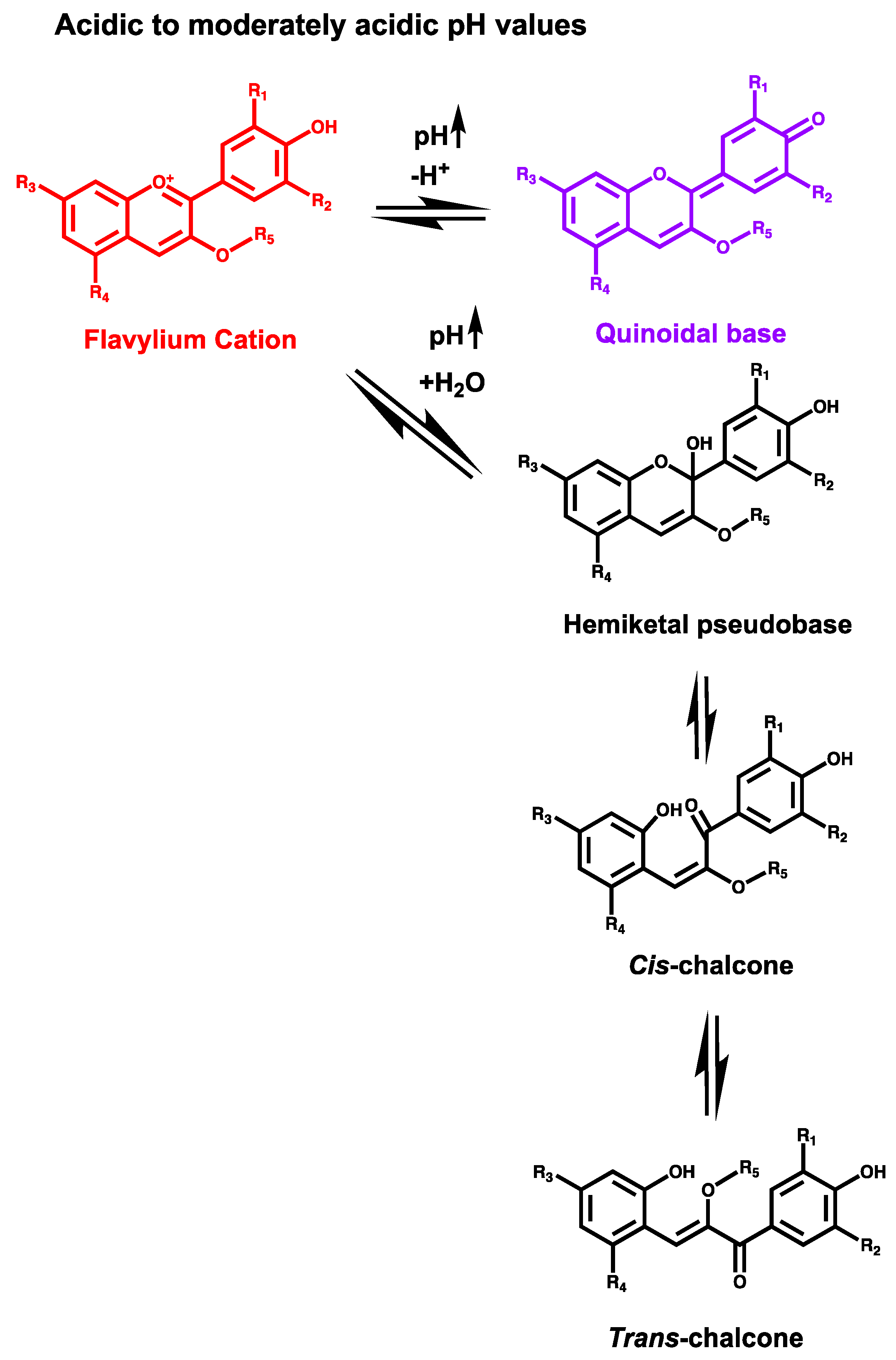{kind=link}
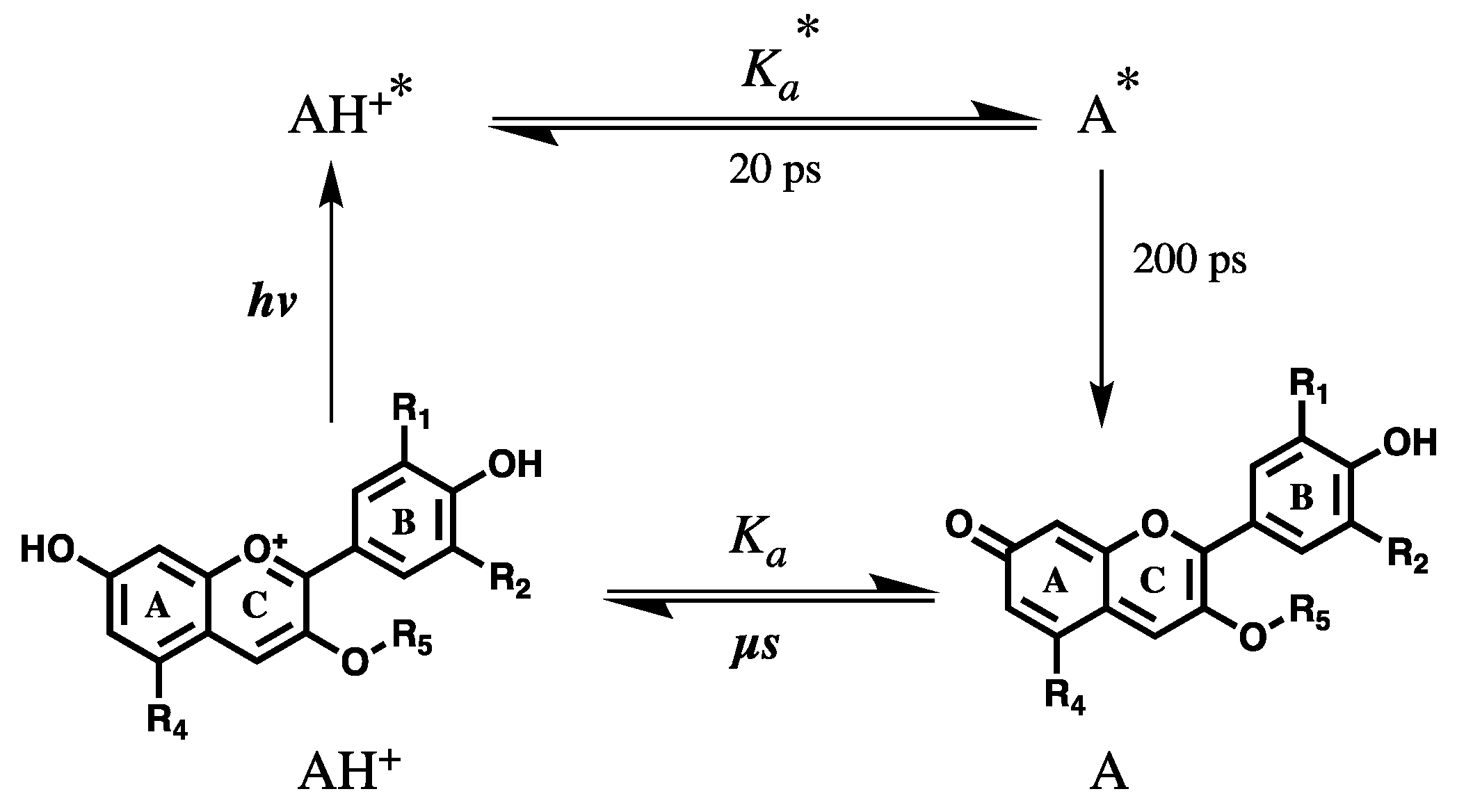{kind=link}
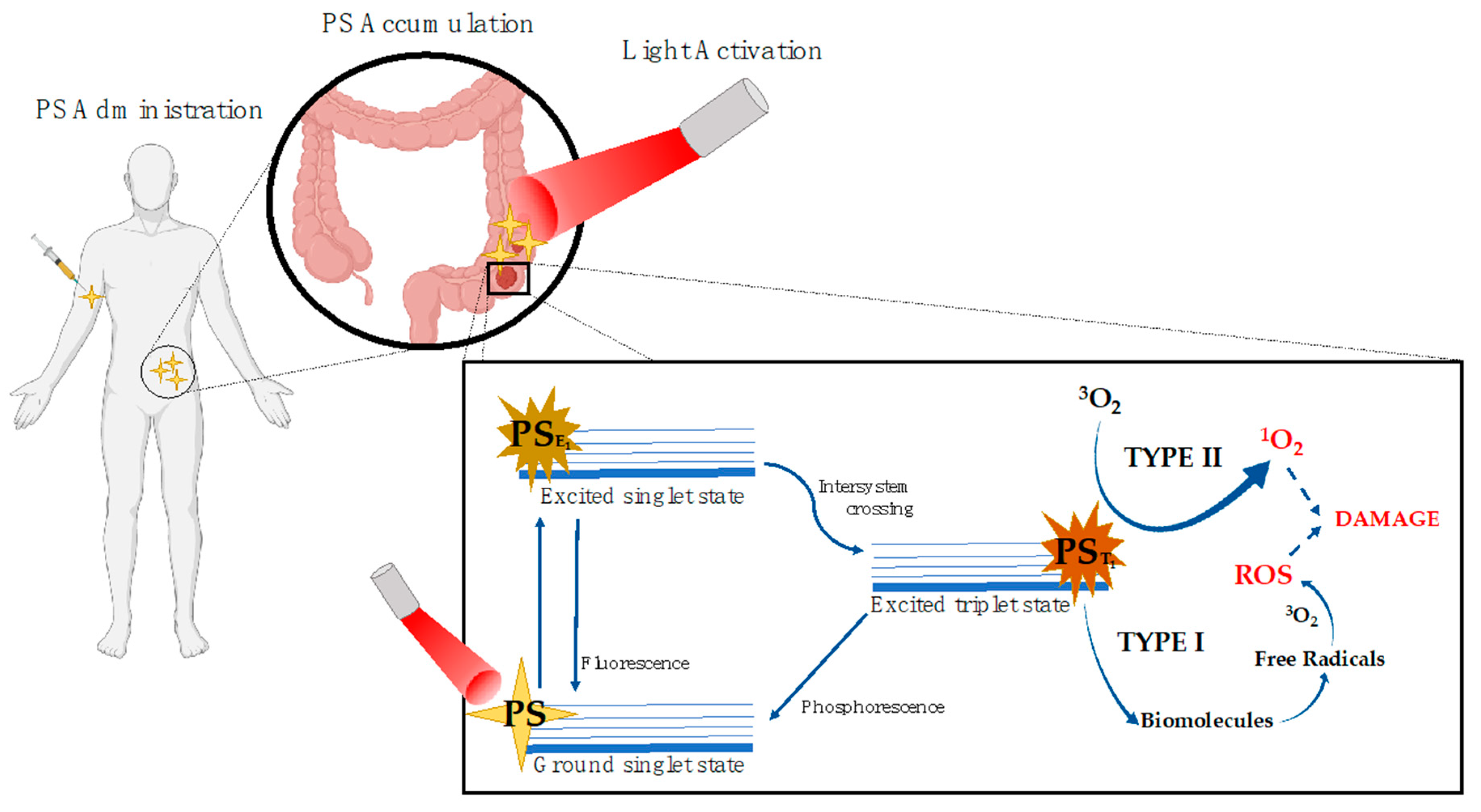{kind=link}
{kind=link}
{kind=link}
| Sources | Anthocyanin Components | Study Models | Biological Effects | Reference |
|---|---|---|---|---|
| — | Delphinidin-3-O-glucoside | HaCat human keratinocytes; SKH1-hairless mouse skin; Human dermal fibroblasts; JB6 P+ mouse keratinocytes | UVB-mediated oxidative stress and DNA damage; UVB-induced MMP-1 expression; UVB-induced COX-2 expression (via MAPKK4 and PI-3K targeting) and PGE2 production | [78,79,80] |
| — | Cyanidin-3-O-glucoside | JB6 P+ mouse keratinocytes | UVB-induced COX-2 expression (via MKK4, MEK1, and Raf-1 targeting) and PGE2 production | [81] |
| — | Cyanidin-3-O-glucoside | Human keratinocytes | UVB-induced cellular morphology change and toxicity UVB-induced ROS generation UVB-induced COX-2 expression | [82] |
| Topical formulation with blueberry extract (Vaccinium myrtillus) | n.s. | HaCat human keratinocytes; Zebrafish; Human skin explants | penetration of the stratum corneum (liposomal formulation) UVC and UVA-mediated cytotoxicity wound repair | [84] |
| Bog blueberry extract (Vaccinium uliginosum L.) | Cyanidin, Petunidin, Malvidin and Delphinidin-3-O-glucosides; Delphinidin-3-O-arabinose | Human dermal fibroblasts (HFF-1) | UVB-induced MMP-1, MMP-8, and MMP-13; UVB-induced production of pro-inflammatory cytokines TNF- α, IL-8, IL-6, and IL-1β | [85] |
| Purple-fleshed sweet potato extract | Cyanidin and Peonidin-3-(p-hydroxybenzoyl)-O-sophoroside-5-O-glucosides; Cyanidin and Peonidin 3-(caffeoyl)-O-sophoroside-5-O-glucosides; Cyanidin and Peonidin 3-(feruloyl)-O-sophoroside-5-O-glucosides | BALB/c-nu mouse skin | UVB-induced oxidative stress Prevention of UVB-induced water loss, collagen degradation, epidermal hyperplasia and wrinkle formation; UVB-induced production and pro-inflammatory cytokines TNF-α and IL-6; | [86] |
| Topical formulation with strawberry extract enriched with coenzyme Q10 | Pelargonidin and Cyanidin-3-O-glucosides; Pelargonidin-3-(malonyl)-O-glucoside; Pelargonidin-3-O-rutinoside | Human dermal fibroblasts (HuDe) | UVA-mediated cytotoxicity | [92] |
| Seed coat of black soya bean extract | Cyanidin, Peonidin and Delphinidin-3-O-glucoside | — | Human and mushroom tyrosinase | [95] |
| Rose of Sharon (Hibiscus syriacus L.) | Cyanidin-3-O-glucoside, Cyanidin-3-O-galactoside, Cyanidin-3,5-O-diglucoside | Mouse melanocytes (B16F10); Zebrafish | Melanogenesis by activation of ERK signaling pathway | [96] |
| Bilberry extract | Delphinidin 3-galactoside delphinidin 3-glucoside, cyanidin 3-galactoside, delphinidin 3-arabinoside, cyanidin 3-glucoside, cyanidin 3-arabinoside, petunidin 3-glucoside, malvidin 3-glucoside malvidin 3-arabinoside | Human adult RPE cells (ARPE-19) | Photooxidation of pyridinium bisretinoid A2E | [103] |
| Bilberry (Vaccinium myrtillus) | - | Human adult RPE cells (ARPE-19) | Upregulate the oxidative stress defense enzymes HO-1 and GST-pi | [104] |
| Maqui berry extract (Aristotelia chilensis) | Delphinidin 3,5-O-diglucoside and delphinidin 3-O-sambubioside-5-O-glucoside | Murine photoreceptor cells (661W) | Light-induced photoreceptor degeneration by inhibiting phosphorylation of p38 | [105] |
| Blueberry extract | Malvidin, malvidin-3-glucoside and malvidin-3-galactoside | human retinal capillary endothelial cells (HRCECs) | SOD and catalase Inhibition of ICAM-1 and NF-κβ | [106] |
| Blueberry anthocyanins bilberries (Vaccinium myrtillus) | Rats | Diabetes-induced retinal abnormalities through Nrf2/HO-1 signaling, Decreased retinal vascular endothelial growth factor (VEGF) expression Degradation of zonula occludens-1, occludin and claudin-5 | [107,108] | |
| Black soybean seeds | Rat retinal neurons | damage induced by N-methyl-N-nitrosourea | [109] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, H.; Correia, P.; Pereira, A.R.; Araújo, P.; Mateus, N.; de Freitas, V.; Oliveira, J.; Fernandes, I. Exploring the Applications of the Photoprotective Properties of Anthocyanins in Biological Systems. Int. J. Mol. Sci. 2020, 21, 7464. https://doi.org/10.3390/ijms21207464
Oliveira H, Correia P, Pereira AR, Araújo P, Mateus N, de Freitas V, Oliveira J, Fernandes I. Exploring the Applications of the Photoprotective Properties of Anthocyanins in Biological Systems. International Journal of Molecular Sciences. 2020; 21(20):7464. https://doi.org/10.3390/ijms21207464
Chicago/Turabian StyleOliveira, Hélder, Patrícia Correia, Ana Rita Pereira, Paula Araújo, Nuno Mateus, Victor de Freitas, Joana Oliveira, and Iva Fernandes. 2020. "Exploring the Applications of the Photoprotective Properties of Anthocyanins in Biological Systems" International Journal of Molecular Sciences 21, no. 20: 7464. https://doi.org/10.3390/ijms21207464