Gga-miR-3525 Targets PDLIM3 through the MAPK Signaling Pathway to Regulate the Proliferation and Differentiation of Skeletal Muscle Satellite Cells


Abstract
:1. Introduction
2. Results
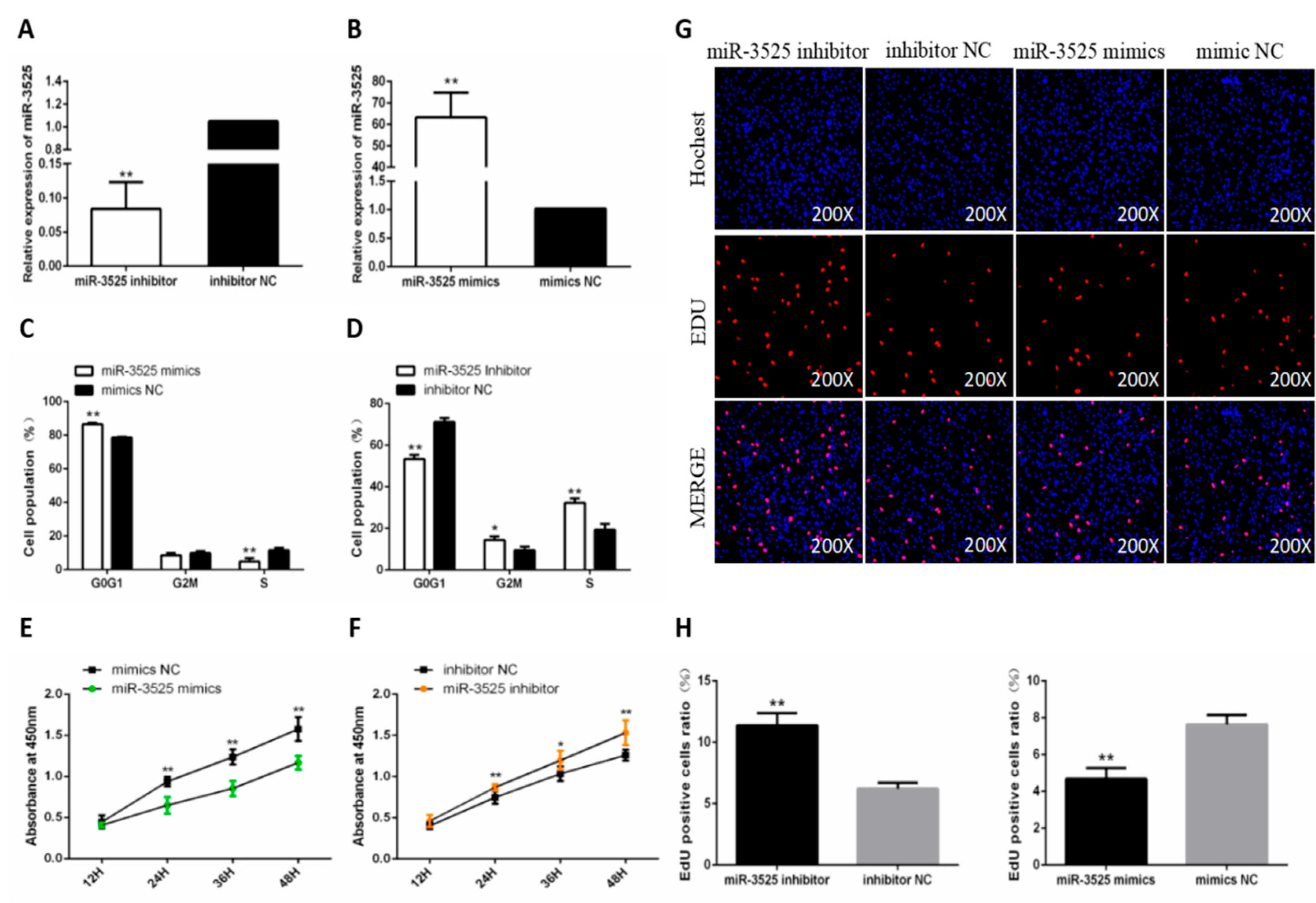
2.1. gga-miR-3525 Reduced the Proliferation of Chicken SMSCs
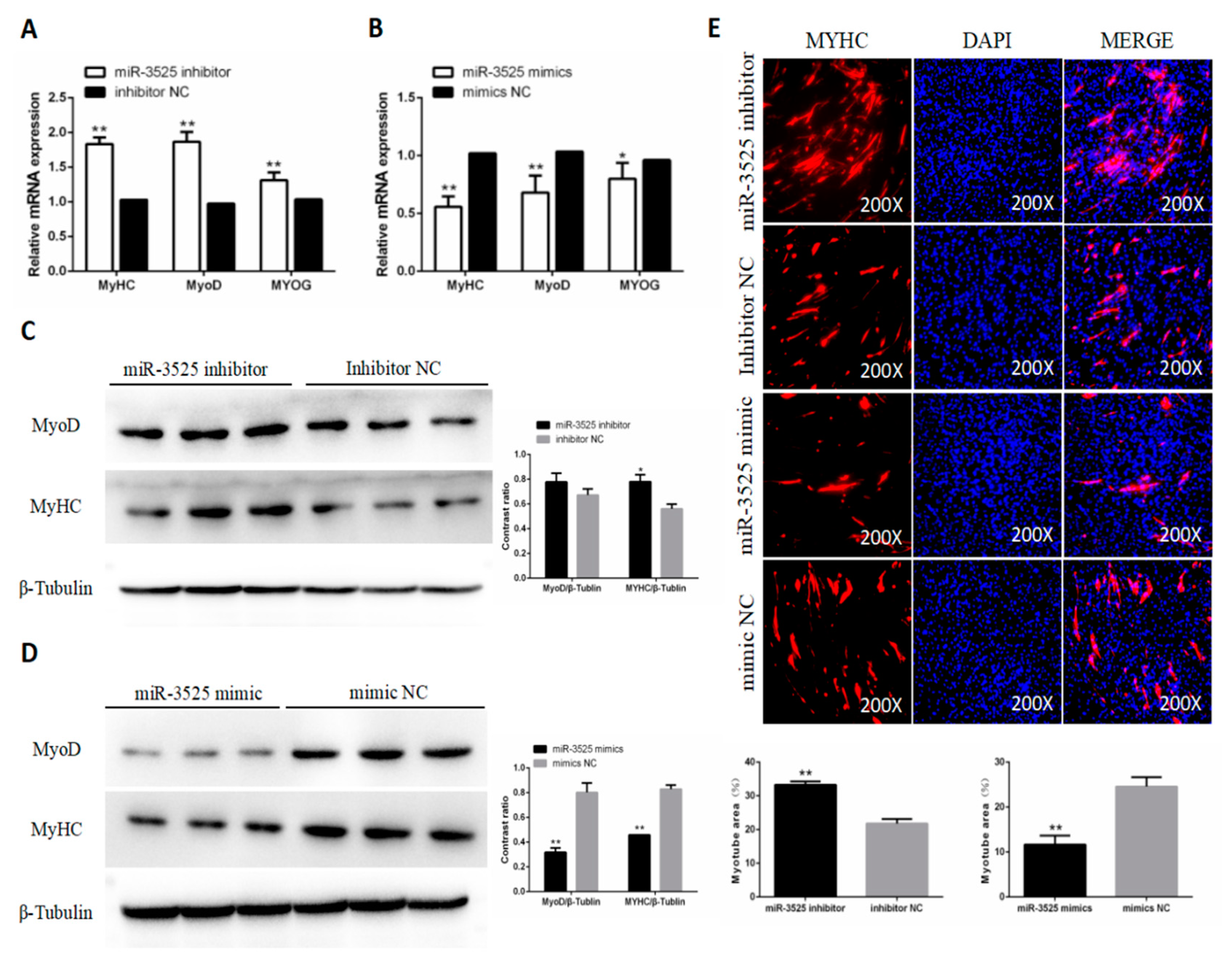
2.2. gga-miR-3525 Reduced the Differentiation of Chicken SMSCs
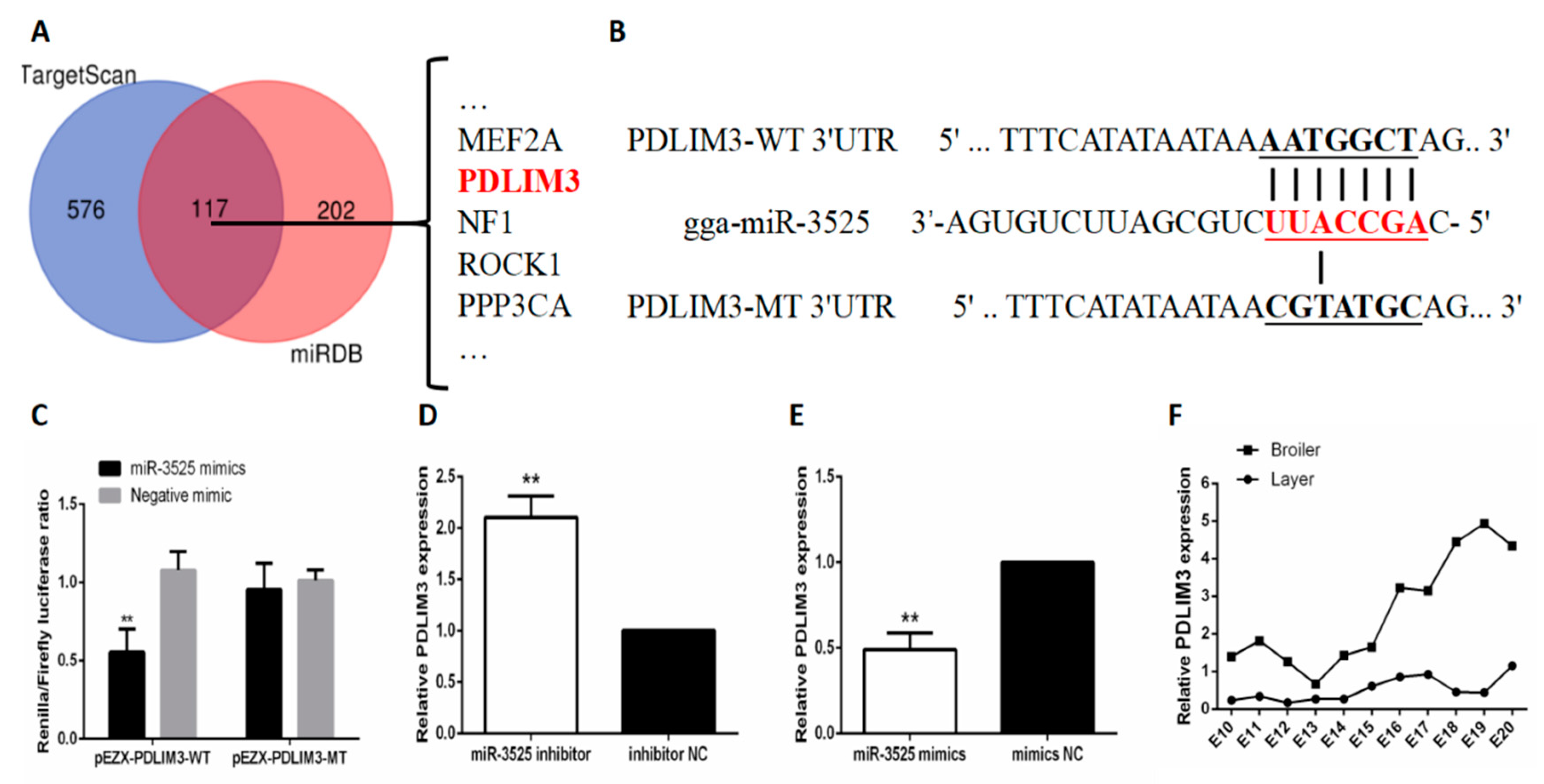
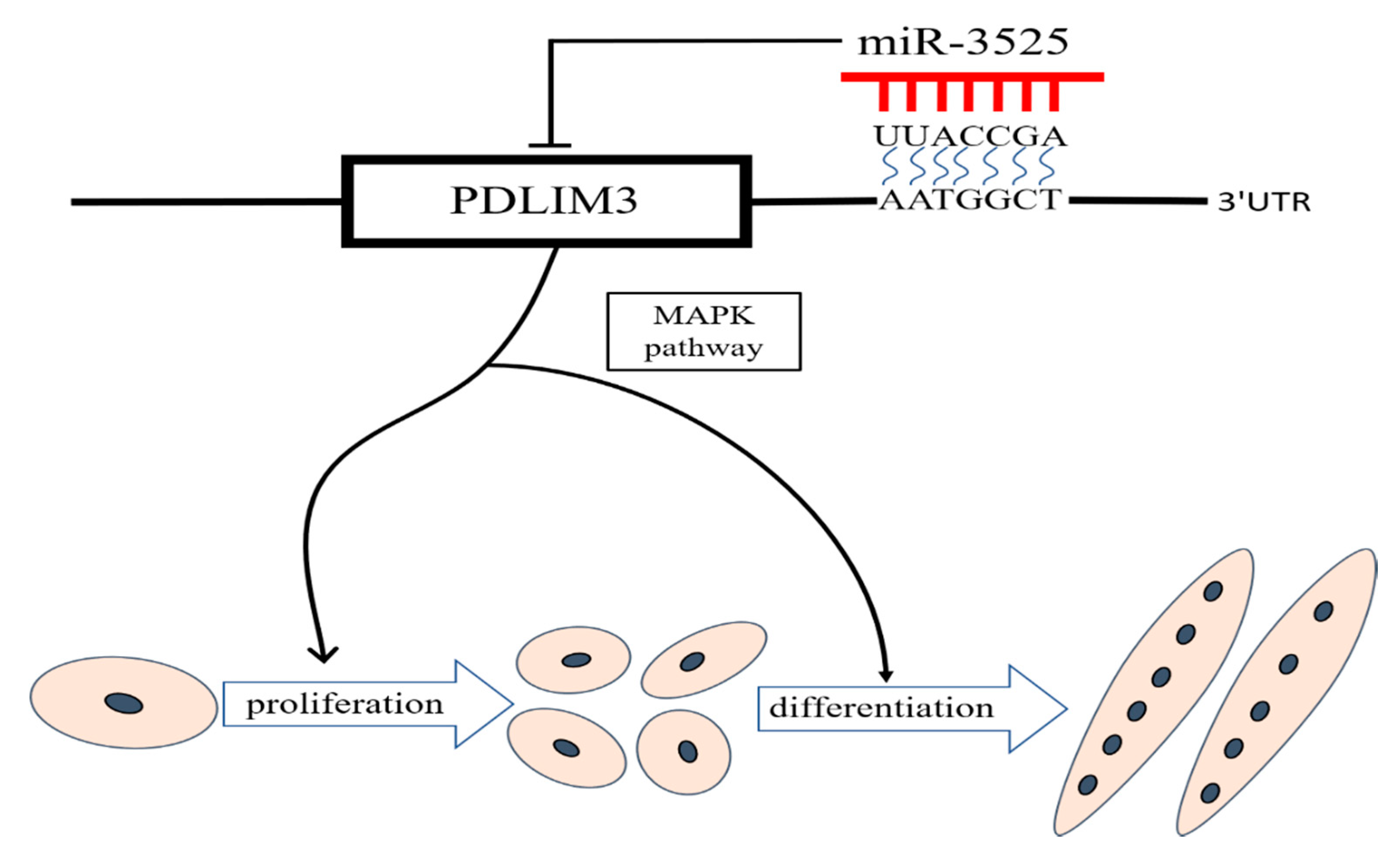
2.3. gga-miR-3525 Can Target PDLIM3 and Inhibit its Expression
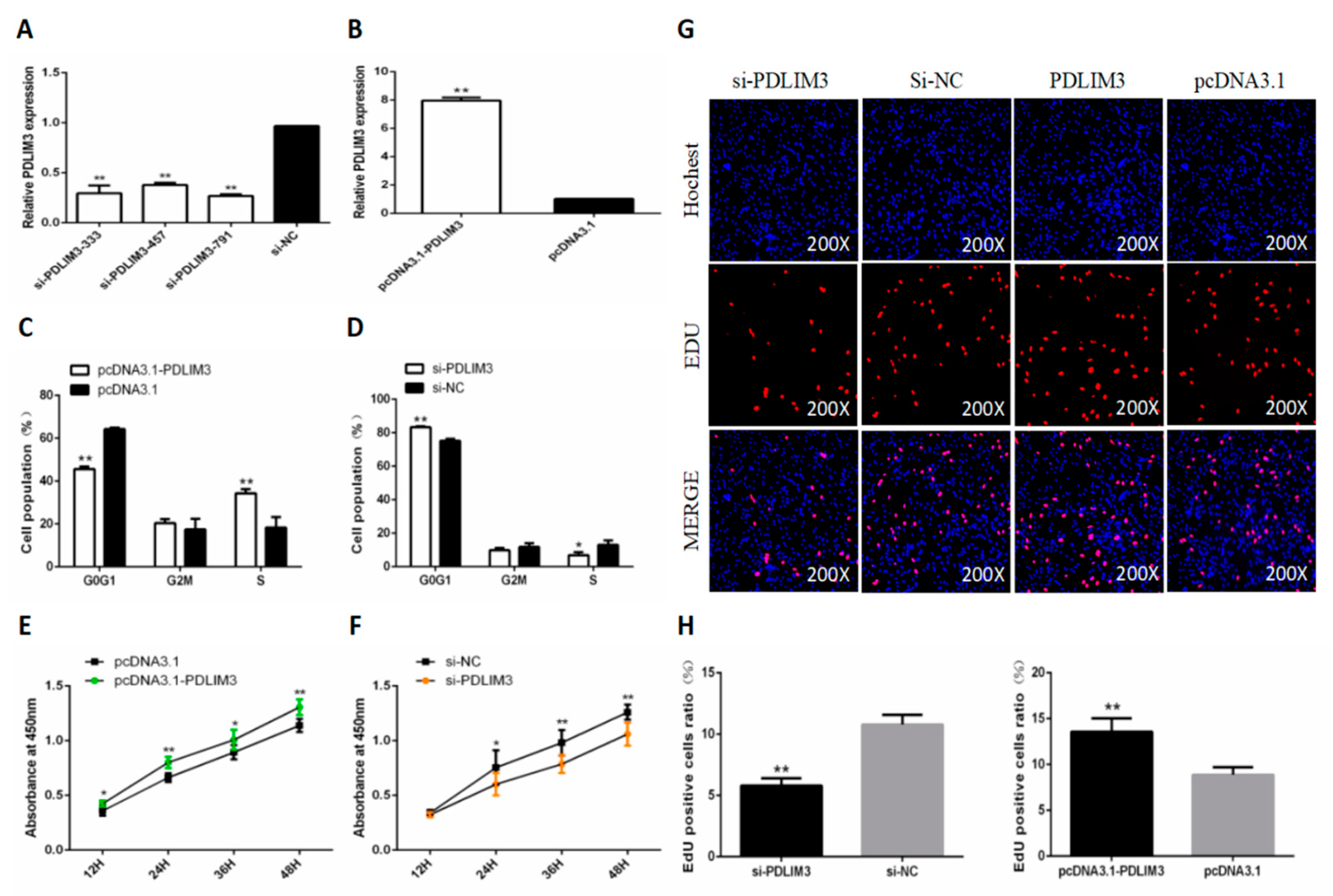
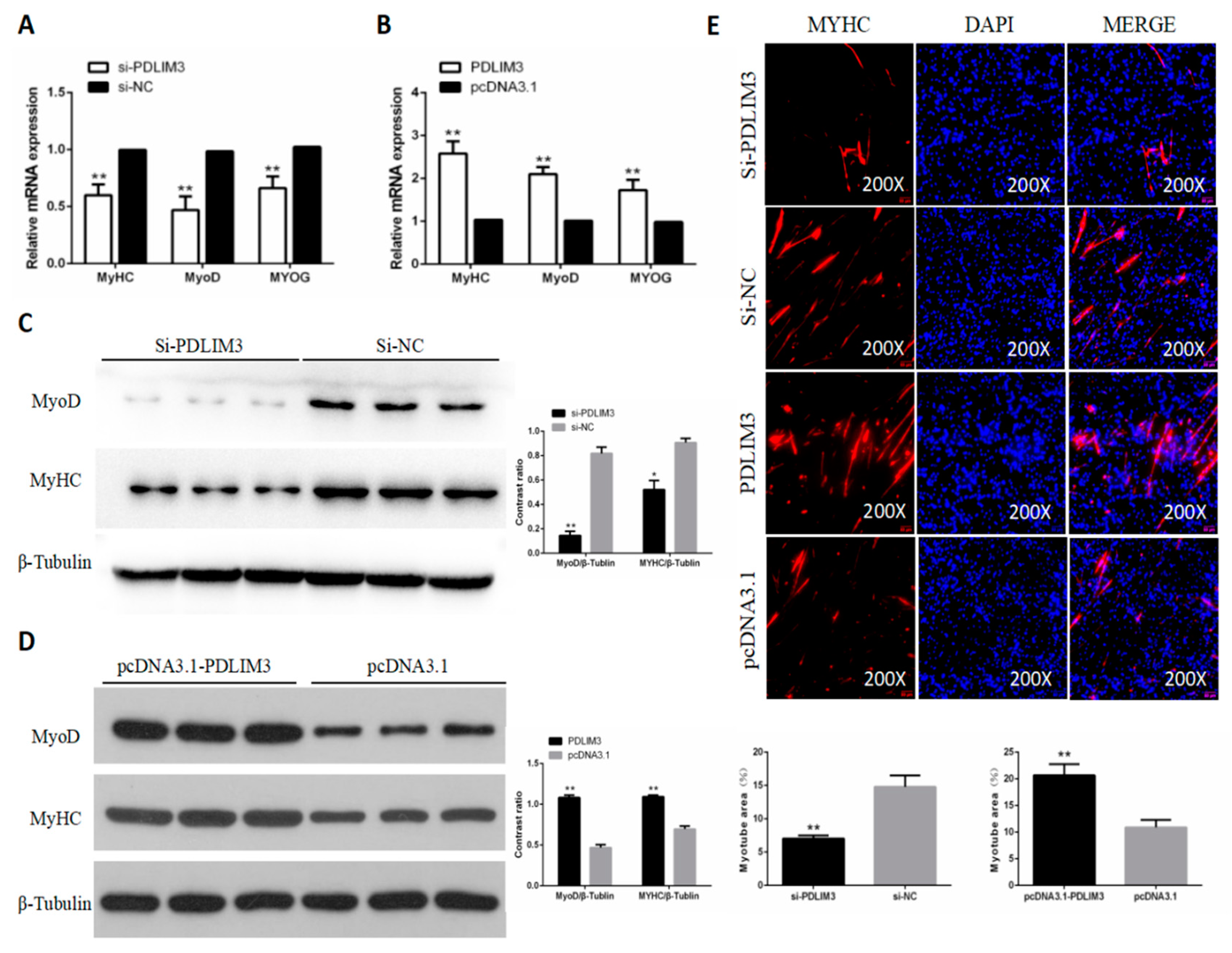
2.4. PDLIM3 Promotes the Proliferation of Chicken SMSCs
2.5. PDLIM3 Promotes the Differentiation of Chicken SMSCs
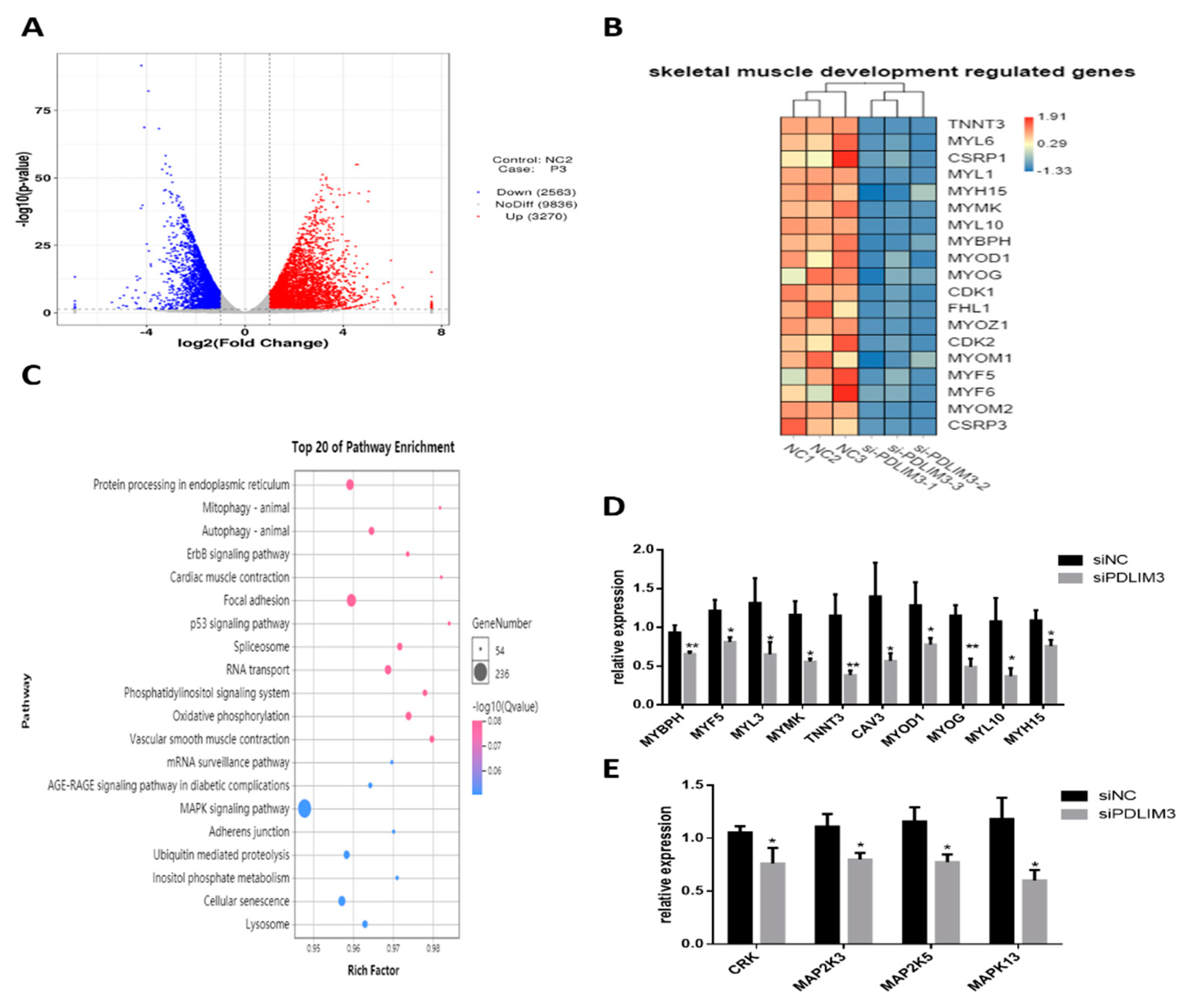
2.6. Transcriptomic Changes Induced by PDLIM3
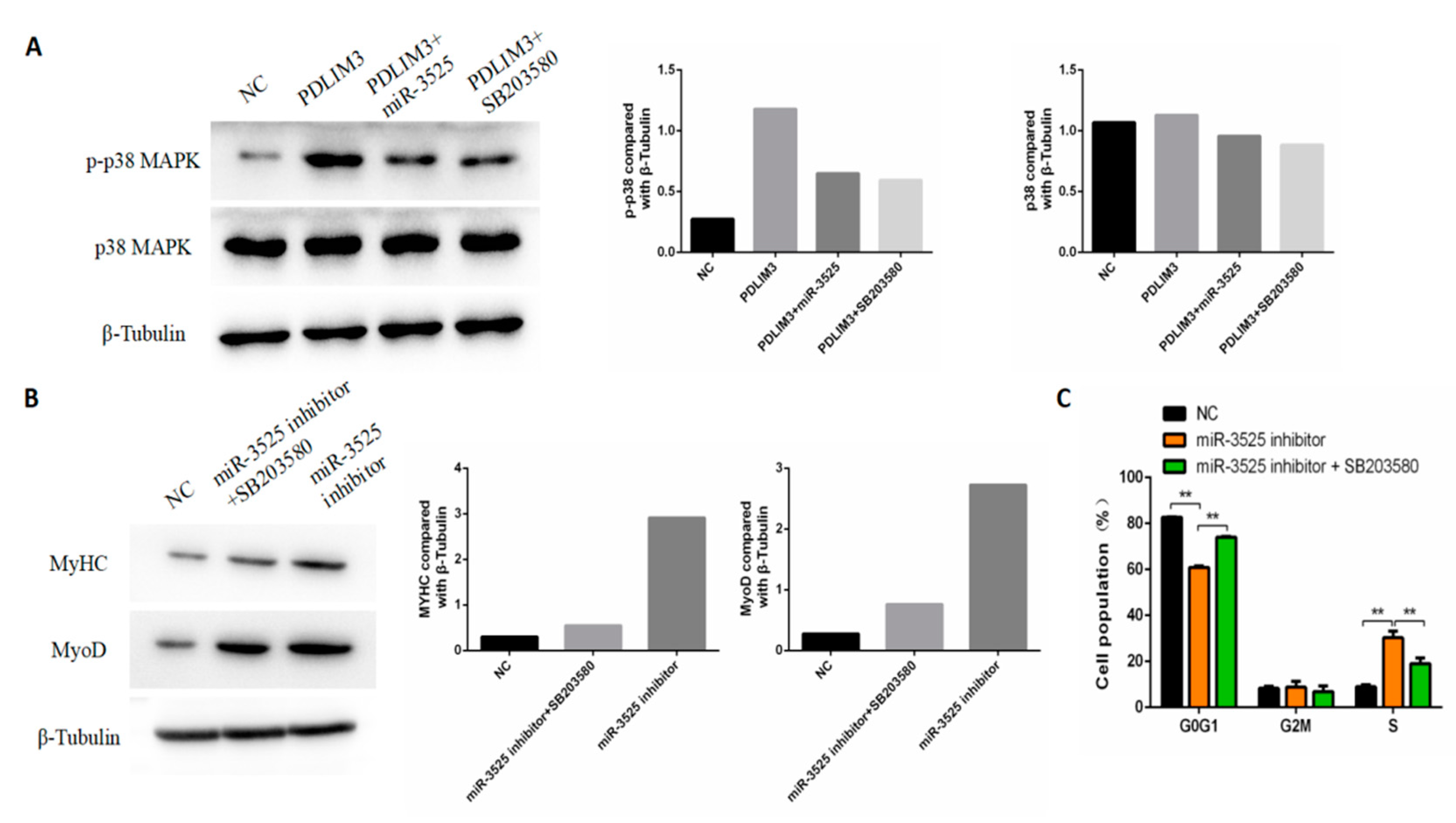
2.7. gga-miR-3525 Regulates Proliferation and Differentiation through the p38/MAPK Pathway
3. Discussion
4. Materials and Methods
4.1. Ethics Standards
4.2. Animals and Samples
4.3. Cell Culture
4.4. Construction and Transfection of Plasmid and RNA Oligonucleotides
4.5. Extraction of RNA, Synthesis of cDNA, and Real-Time Quantitative PCR
4.6. Cell Proliferation
4.7. Cell Differentiation
4.8. Luciferase Reporter Assay
4.9. Transcriptome Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Horak, M.; Novak, J.; Bienertova-Vasku, J. Muscle-specific microRNAs in skeletal muscle development. Dev. Biol. 2016, 410, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, D.; Tang, S.; Li, D.; Han, R.; Tian, Y.; Li, H.; Li, G.; Liu, X.; Kang, X.; et al. A multiallelic indel in the promoter region of the Cyclin-dependent kinase inhibitor 3 gene is significantly associated with body weight and carcass traits in chickens. Poult. Sci. 2019, 98, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.D.; Li, D.Y.; Zhang, L.; Yang, M.Y.; Zhao, X.L.; Wang, Y.; Liu, Y.P.; Zhu, Q. Housing system influences abundance of Pax3 and Pax7 in postnatal chicken skeletal muscles. Poult. Sci. 2014, 93, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.H.; Grote, E.; Mohler, W.; Vignery, A. Cell-cell fusion. FEBS Lett. 2007, 581, 2181–2193. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z. Regulation of skeletal myogenesis by microRNAs. J. Cell. Physiol. 2019, 235, 87–104. [Google Scholar]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Shukla, G.C.; Singh, J.; Barik, S. MicroRNAs: Processing, maturation, target recognition and regulatory functions. Mol. Cell. Pharmacol. 2011, 3, 83. [Google Scholar]
- Tong, H.L.; Jiang, R.Y.; Zhang, W.W.; Yan, Y.Q. MiR-2425-5p targets RAD9A and MYOG to regulate the proliferation and differentiation of bovine skeletal muscle-derived satellite cells. Sci. Rep. 2017, 7, 418. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Tang, S.; Du, F.; Li, H.; Shen, X.; Li, D.; Wang, Y.; Zhang, Z.; Xia, L.; Zhu, Q.; et al. miR-99a-5p Regulates the Proliferation and Differentiation of Skeletal Muscle Satellite Cells by Targeting MTMR3 in Chicken. Genes 2020, 11, 369. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; He, H.; Shen, X.; Tang, S.; Zhao, J.; Cao, X.; Han, S.; Cui, C.; Chen, Y.; Wei, Y.; et al. MicroRNA Profiling Reveals an Abundant miR-200a-3p Promotes Skeletal Muscle Satellite Cell Development by Targeting TGF-β2 and Regulating the TGF-β2/SMAD Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 3274. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Brahmakshatriya, V.; Zhu, H.; Lupiani, B.; Reddy, S.M.; Yoon, B.-J.; Gunaratne, P.H.; Kim, J.H.; Chen, R.; Wang, J.; et al. Identification of differentially expressed miRNAs in chicken lung and trachea with avian influenza virus infection by a deep sequencing approach. BMC Genom. 2009, 10, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Tong, C.; Ma, S.; Zhou, L.; Zhao, L.; Zhao, X. Involvement of MicroRNAs in Probiotics-Induced Reduction of the Cecal Inflammation by Salmonella Typhimurium. Front. Immunol. 2017, 8, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Liao, R.; Wu, N.; Zhu, G.; Tu, Y.; Yang, C. Integrating miRNA and mRNA expression profiles in plasma of laying hens associated with heat stress. Mol. Biol. Rep. 2019, 46, 2779–2789. [Google Scholar] [CrossRef]
- Scaal, M.; Marcelle, C. Chick muscle development. Int. J. Dev. Biol. 2018, 62, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Davey, M.G.; Tickle, C. The chicken as a model for embryonic development. Cytogenet. Genome Res. 2007, 117, 231–239. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using david bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Han, S.; Cui, C.; Wang, Y.; He, H.; Liu, Z.; Shen, X.; Chen, Y.; Li, D.; Zhu, Q.; Yin, H. Knockdown of CSRP3 inhibits differentiation of chicken satellite cells by promoting TGF-β/Smad3 signaling. Gene 2019, 707, 36–43. [Google Scholar] [CrossRef]
- Xia, H.; Winokur, S.T.; Kuo, W.L.; Altherr, M.R.; Bredt, D.S. Actinin-associated LIM protein: Identification of a domain interaction between PDZ and spectrin-like repeat motifs. J. Cell Biol. 1997, 139, 507–515. [Google Scholar] [CrossRef]
- Bouju, S.; Piétu, G.; Le Cunff, M.; Cros, N.; Malzac, P.; Pellissier, J.-F.; Pons, F.; Léger, J.-J.; Auffray, C.; Dechesne, C.A. Exclusion of muscle specific actinin-associated LIM protein (ALP) gene from 4q35 facioscapulohumeral muscular dystrophy (FSHD) candidate genes. Neuromuscul. Disord. 1999, 9, 3–10. [Google Scholar] [CrossRef]
- Passier, R.; Richardson, J.A.; Olson, E.N. Oracle, a novel PDZ-LIM domain protein expressed in heart and skeletal muscle. Mech. Dev. 2000, 92, 277–284. [Google Scholar] [CrossRef]
- Ohsawa, N.; Koebis, M.; Suo, S.; Nishino, I.; Ishiura, S. Alternative splicing of PDLIM3/ALP, for α-actinin-associated LIM protein 3, is aberrant in persons with myotonic dystrophy. Biochem. Biophys. Res. Commun. 2011, 409, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Pomiès, P.; Pashmforoush, M.; Vegezzi, C.; Chien, K.R.; Auffray, C.; Beckerle, M.C. The cytoskeleton-associated PDZ-LIM protein, ALP, acts on serum response factor activity to regulate muscle differentiation. Mol. Biol. Cell 2007, 18, 1723–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cambier, L.; Pomiès, P. Nuclear translocation of the cytoskeleton-associated protein, smALP, upon induction of skeletal muscle differentiation. Biochem. Biophys. Res. Commun. 2011, 409, 628–633. [Google Scholar] [CrossRef]
- Xue, K.; Wang, Y.; Hou, Y.; Wang, Y.; Zhong, T.; Li, L.; Zhang, H.; Wang, L. Molecular characterization and expression patterns of the actinin-associated LIM protein (ALP) subfamily genes in porcine skeletal muscle. Gene 2014, 539, 111–116. [Google Scholar] [CrossRef]
- Piórkowska, K.; Żukowski, K.; Ropka-Molik, K.; Tyra, M.; Gurgul, A. A comprehensive transcriptome analysis of skeletal muscles in two Polish pig breeds differing in fat and meat quality traits. Genet. Mol. Biol. 2018, 41, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.-J.; Li, J.-H.; Chen, H.-F.; Tan, Y.-Y.; Liu, S.-R.; Zhang, Y.; Xu, H.; Yang, J.-H.; Liu, S.; Zheng, L.-L.; et al. Inhibition of the JNK/MAPK signaling pathway by myogenesis-associated miRNAs is required for skeletal muscle development. Cell Death Differ. 2018, 25, 1581–1597. [Google Scholar] [CrossRef] [Green Version]
- Robert-Gangneux, F.; Creuzet, C.; Dupouy-Camet, J.; Roisin, M.P. Involvement of the mitogen-activated protein (MAP) kinase signalling pathway in host cell invasion by Toxoplasma gondii. Parasite 2000, 7, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Jones, N.C.; Tyner, K.J.; Nibarger, L.; Stanley, H.M.; Cornelison, D.D.; Fedorov, Y.V.; Olwin, B.B. The p38alpha/beta MAPK functions as a molecular switch to activate the quiescent satellite cell. J. Cell Biol. 2005, 169, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Bernet, J.D.; Doles, J.D.; Hall, J.K.; Kelly Tanaka, K.; Carter, T.A.; Olwin, B.B. p38 MAPK signaling underlies a cell-autonomous loss of stem cell self-renewal in skeletal muscle of aged mice. Nat. Med. 2014, 20, 265–271. [Google Scholar] [CrossRef]
- Keren, A.; Tamir, Y.; Bengal, E. The p38 MAPK signaling pathway: A major regulator of skeletal muscle development. Mol. Cell. Endocrinol. 2006, 252, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tan, J.; Qi, Q.; Yang, L.; Wang, Y.; Zhang, C.; Hu, L.; Chen, H.; Fang, X. miR-487b-3p Suppresses the Proliferation and Differentiation of Myoblasts by Targeting IRS1 in Skeletal Muscle Myogenesis. Int. J. Biol. Sci. 2018, 14, 760–774. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Lin, S.; Li, G.; Nie, Q.; Zhang, X. Integrative Analyses of miRNA-mRNA Interactions Reveal let-7b, miR-128 and MAPK Pathway Involvement in Muscle Mass Loss in Sex-Linked Dwarf Chickens. Int. J. Mol. Sci. 2016, 17, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; He, H.; Shen, X.; Zhao, J.; Cao, X.; Han, S.; Cui, C.; Chen, Y.; Wei, Y.; Xia, L.; et al. miR-9-5p Inhibits Skeletal Muscle Satellite Cell Proliferation and Differentiation by Targeting IGF2BP3 through the IGF2-PI3K/Akt Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Shen, X.; Cao, X.; He, H.; Han, S.; Chen, Y.; Cui, C.; Wei, Y.; Wang, Y.; Li, D.; et al. HDAC4 Regulates the Proliferation, Differentiation and Apoptosis of Chicken Skeletal Muscle Satellite Cells. Animals 2020, 10, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) |
|---|---|
| miR-3525 mimic | CAGCCAUUCUGCGAUUCUGUGA ACAGAAUCGCAGAAUGGCUGUU |
| Negative mimic | UUGUACUACACAAAAGUACUG |
| miR-3525 inhibitor | UCACAGAAUCGCAGAAUGGCUG |
| Negative inhibitor | CAGUACUUUUGUGUAGUACAA |
| si-PDLIM3-333 | GCAAAGCAAAUCCCUACAATT UUGUAGGGAUUUGCUUUGCTT |
| si-PDLIM3-457 | GGUAGUGAGCUCCUCCUAUTT AUAGGAGGAGCUCACUACCTT |
| si-PDLIM3-791 | GGGAGUGGAAUUCUAGGAATT UUCCUAGAAUUCCACUCCCTT |
| si-NC | UUCUCCGAACGUGUCACGUTT ACGUGACACGUUCGGAGAATT |
| pcDNA3.1-PDLIM3 | GACACAAGCTTGCCACCATGCCACA GTGTCCTCGAGCTAGCATTTTGGGTA |
| Gene | Primer Sequences (5′-3′) | Product Size (bp) | TM (°C) |
|---|---|---|---|
| β-actin | F: GTCCACCGCAAATGCTTCTAA R: TGCGCATTTATGGGTTTTGTT | 78 | 58 |
| MyoG | F: CGTGTGCCACAGCCAATG R: CCGCCGGAGAGAGACCTT | 63 | 60 |
| MyoD1 | F: GCCGCCGATGACTTCTATGA R: CAGGTCCTCGAAGAAGTGCAT | 66 | 60 |
| MYHC | F: GAAGGAGACCTCAACGAGATGG R: ATTCAGGTGTCCCAAGTCATCC | 138 | 60 |
| PDLIM3 | F: AAGCACCTGTAACAAAGATA R: GCCCTCCACAAAGAAGTAGC | 188 | 55 |
| MYBPH | F: ATCCGCCTACCTCGTCAG R: GGCTGGTTGTCCTTGGTC | 122 | 58 |
| MYL10 | F: TGTTGCTTAACCTCTTGCTTT R: TACCAAATGCTCTTCCCAGT | 89 | 56 |
| MYL3 | F: GAAGAACCCAAACCAGCA R: CCCAAAGCCCTCAAGAC | 200 | 55 |
| TNNT3 | F: GGCTGAGAAGGAGAAGGAG R: GCTGTATGAGGCACCCA | 136 | 56 |
| MYMK | F: TCCCCACCATCAGCATC R: GCATGAAACATAGCACCGA | 129 | 56 |
| MAPK13 | F: TTCCTGAGTCGTGTTTGGT R: GGGGCATGGCTGTAGTAA | 145 | 56 |
| CAV3 | F: GTGCCCTGCATCAAGAG R: CGCAGCATAACCCTGAC | 134 | 55 |
| CRK | F: CACTCCGCTCCCTAACC R: CCTTCCCACTGACCACTC | 150 | 56 |
| MAP2K3 | F: GCCTATGGTGTGGTGGAGA R: AAGCAGTCAACCGTCCTCA | 143 | 58 |
| MAP2K5 | F: GGCCAGATGAATGAACAAG R: GCCAGGATTTTCCCACTA | 104 | 54 |
| miR-3525 | F: CAGCCATTCTGCGATTCTGTGA R: CAGGTCCAGTTTTTTTTTTTTTT | – | 56 |
| U6 | F: GGGCCATGCTAATCTTCTCTGTA R: CAGGTCCAGTTTTTTTTTTTTTT | – | 56 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, H.; Zhao, J.; He, H.; Chen, Y.; Wang, Y.; Li, D.; Zhu, Q. Gga-miR-3525 Targets PDLIM3 through the MAPK Signaling Pathway to Regulate the Proliferation and Differentiation of Skeletal Muscle Satellite Cells. Int. J. Mol. Sci. 2020, 21, 5573. https://doi.org/10.3390/ijms21155573
Yin H, Zhao J, He H, Chen Y, Wang Y, Li D, Zhu Q. Gga-miR-3525 Targets PDLIM3 through the MAPK Signaling Pathway to Regulate the Proliferation and Differentiation of Skeletal Muscle Satellite Cells. International Journal of Molecular Sciences. 2020; 21(15):5573. https://doi.org/10.3390/ijms21155573
Chicago/Turabian StyleYin, Huadong, Jing Zhao, Haorong He, Yuqi Chen, Yan Wang, Diyan Li, and Qing Zhu. 2020. "Gga-miR-3525 Targets PDLIM3 through the MAPK Signaling Pathway to Regulate the Proliferation and Differentiation of Skeletal Muscle Satellite Cells" International Journal of Molecular Sciences 21, no. 15: 5573. https://doi.org/10.3390/ijms21155573