Epidemiological and Genomic Characterization of Campylobacter jejuni Isolates from a Foodborne Outbreak at Hangzhou, China

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Causative Pathogen Scanning
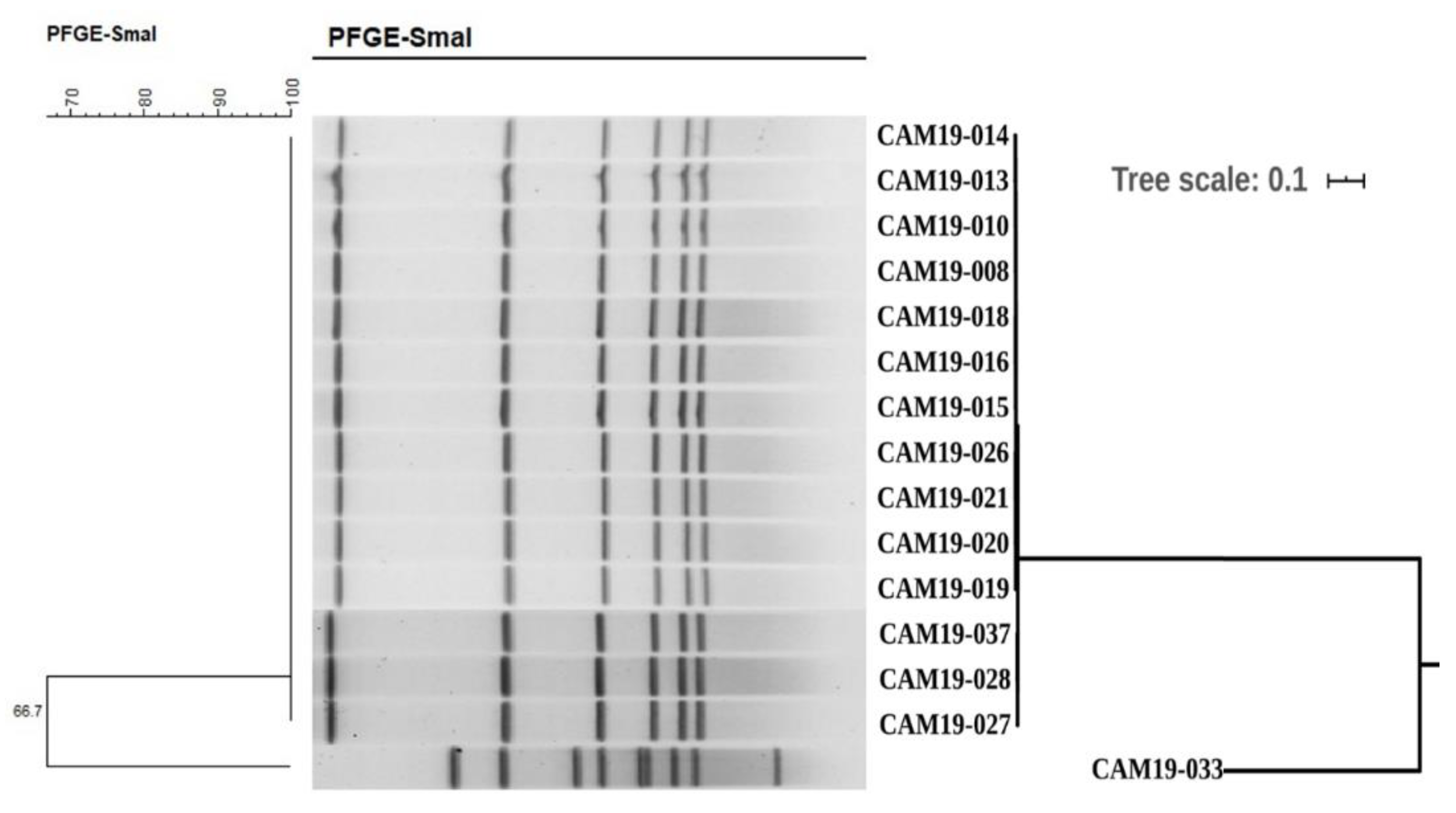
2.2. PFGE Profiling Studies
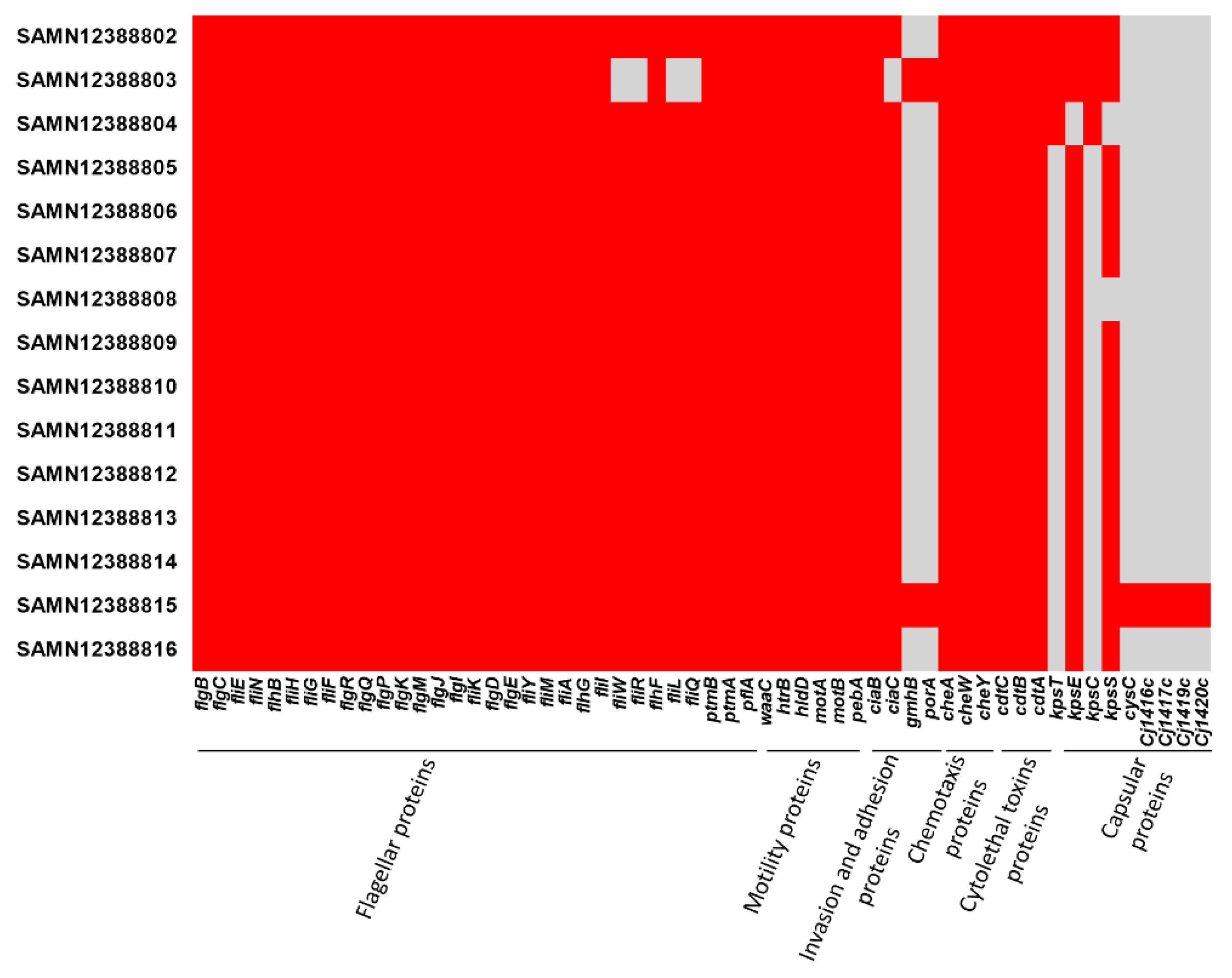
2.3. Genomic Sequencing
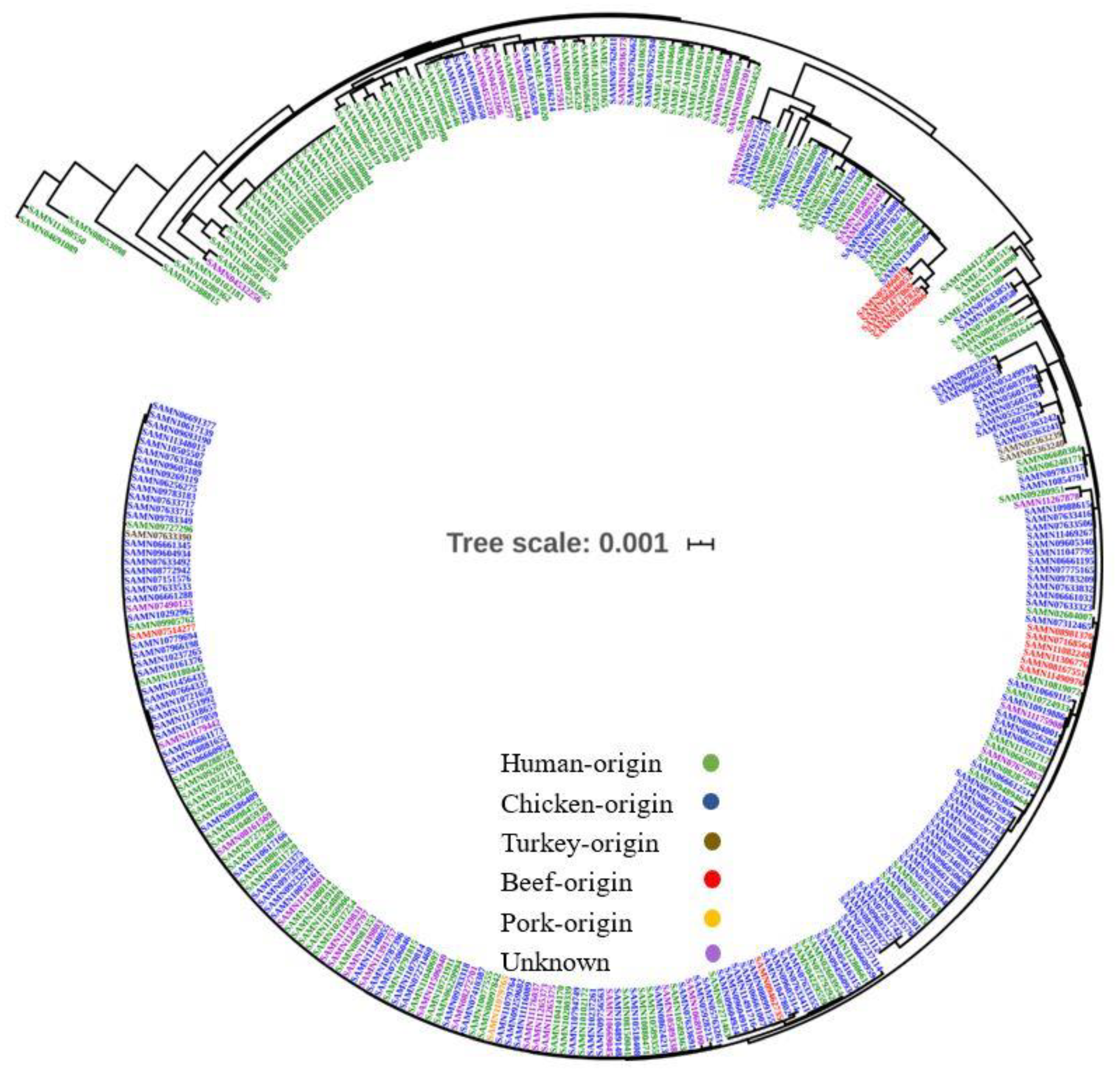
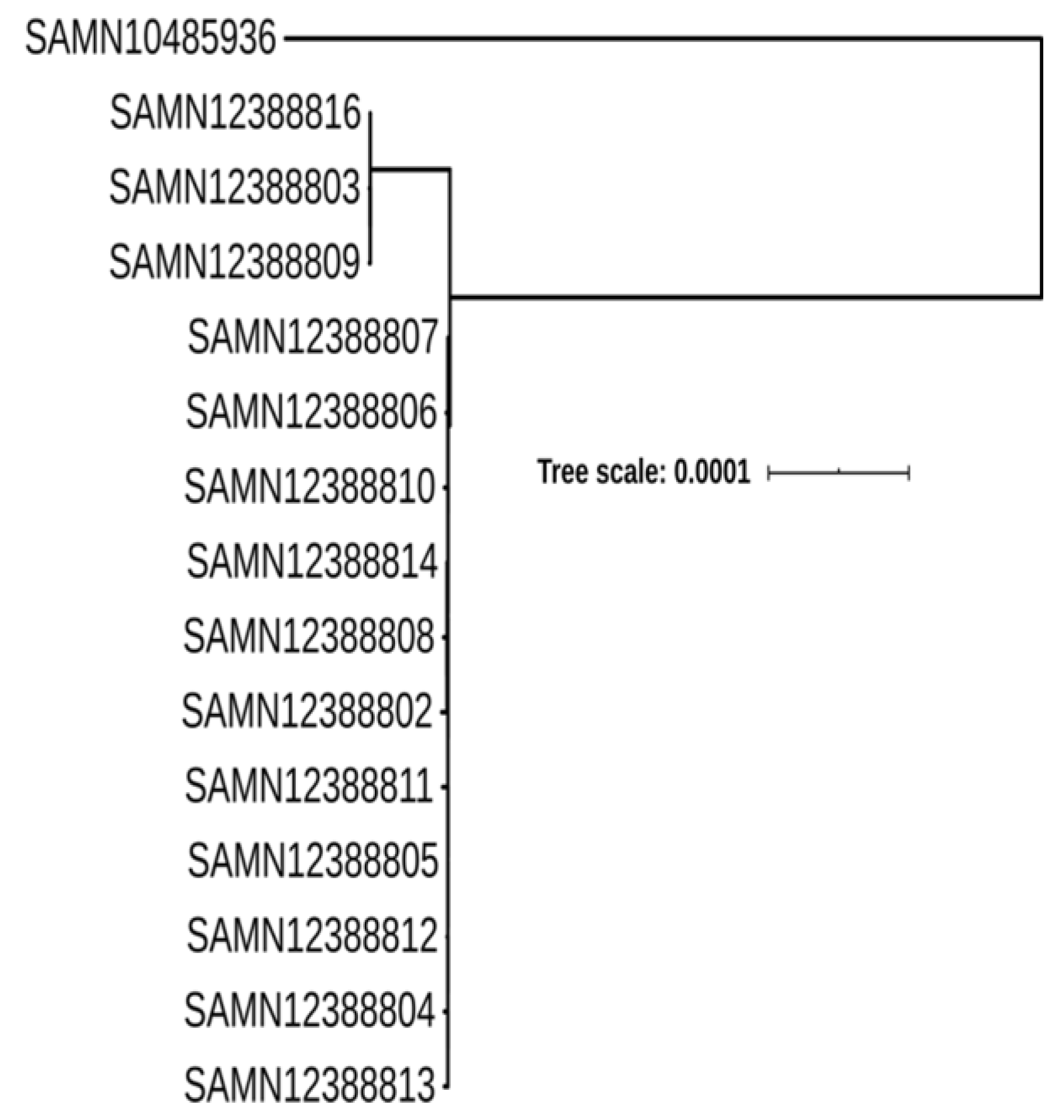
2.4. Genome Comparison and Phylogenomic Analysis
3. Discussion
4. Material and Methods
4.1. Epidemiological Investigation
4.2. Samples Collection
4.3. Pathogen Detection
4.4. Isolation and Identification of Campylobacter spp.
4.5. Pulsed Field Gel Electrophoresis (PFGE) Testing
4.6. Genomic Sequencing and Bioinformatic Analysis
4.7. Ethical Approval
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, M.; Li, Q.; He, L.; Meng, F.; Gu, Y.; Zheng, M.; Gong, Y.; Wang, P.; Ruan, F.; Zhou, L.; et al. Association Study Between an Outbreak of Guillain-Barre Syndrome in Jilin, China, and Preceding Campylobacter jejuni Infection. Foodborne Pathog. Dis. 2010, 7, 913–919. [Google Scholar] [CrossRef]
- Silva, J.; Leite, D.; Fernandes, M.; Mena, C.; Gibbs, P.A.; Teixeira, P. Campylobacter spp. as a Foodborne Pathogen: A Review. Front. Microbiol. 2011, 2, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paudyal, N.; Pan, H.; Liao, X.; Zhang, X.; Li, X.; Fang, W.; Yue, M. A Meta-Analysis of Major Foodborne Pathogens in Chinese Food Commodities Between 2006 and 2016. Foodborne Pathog. Dis. 2018, 15, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Wang, Y.; Zhang, Q.; Shen, J. Antimicrobial Resistance in Campylobacter spp. Microbiol. Spectr. 2018, 6, 317–330. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and foodborne outbreaks in 2017. EFSA J. 2018, 16, 5500. [Google Scholar]
- Tack, D.M.; Marder, E.P.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Hurd, S.; Scallan, E.; Lathrop, S.; Muse, A.; Ryan, P.; et al. Preliminary Incidence and Trends of Infections with Pathogens Transmitted Commonly Through Food—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2015–2018. MMWR 2019, 68, 369–373. [Google Scholar]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [Green Version]
- Lahti, E.; Löfdahl, M.; Skarin, J.; Hansson, I.; Engvall, E. Confirmation of a Campylobacteriosis Outbreak Associated with Chicken Liver Pâté Using PFGE and WGS. Zoonses Public Health 2016, 64, 14–20. [Google Scholar] [CrossRef]
- Duarte, A.; Seliwiorstow, T.; Miller, W.G.; De Zutter, L.; Uyttendaele, M.; Dierick, K.; Botteldoorn, N. Discriminative power of Campylobacter phenotypic and genotypic typing methods. J. Microbiol. Methods 2016, 125, 33–39. [Google Scholar] [CrossRef]
- Oakeson, K.F.; Wagner, J.M.; Rohrwasser, A.; Atkinson-Dunn, R. Whole-Genome Sequencing and Bioinformatic Analysis of Isolates from Foodborne Illness Outbreaks of Campylobacter jejuni and Salmonella enterica. J. Clin. Microbiol. 2018, 56, e00161-18. [Google Scholar] [CrossRef] [Green Version]
- Carleton, H.; Gerner-Smidt, P. Whole-Genome Sequencing Is Taking over Foodborne Disease Surveillance: Public health microbiology is undergoing its biggest change in a generation, replacing traditional methods with whole-genome sequencing. Microbe 2016, 11, 311–317. [Google Scholar]
- Acheson, D.; Allos, B.M. Campylobacter jejuni Infections: Update on Emerging Issues and Trends. Clin. Infect. Dis. 2001, 32, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Zhang, X.; Xue, F.; Wang, Y.; Jiang, L.; Jiang, Y. Phenotypic Characters and Molecular Epidemiology of Campylobacter jejuni in East China. J. Food Sci. 2016, 81, M106–M113. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.; Zhang, M.; Zhang, X.; Jia, L.; Xu, J.; Chu, Y.; Liang, Z.; Lv, B.; Liang, H.; Huang, Y.; et al. Molecular and epidemiologyical analysis of a Campylobacter jejuni outbreak in China, 2018. J. Infect. Dev. Ctries. 2019, 13, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, S.K.; Dallas, J.F.; Strachan, N.J.C.; MacRae, M.; McCarthy, N.D.; Wilson, D.J.; Gormley, F.J.; Falush, D.; Ogden, I.D.; Maiden, M.C.J.; et al. Campylobacter Genotyping to Determine the Source of Human Infection. Clin. Infect. Dis. 2009, 48, 1072–1078. [Google Scholar] [CrossRef] [Green Version]
- Scott, A.E.; Timms, A.R.; Connerton, P.L.; Loc Carrillo, C.; Adzfa Radzum, K.; Connerton, I.F. Genome dynamics of Campylobacter jejuni in response to bacteriophage predation. PLoS Pathog. 2007, 3, e119. [Google Scholar] [CrossRef] [Green Version]
- Hänninen, M.L.; Hakkinen, M.; Rautelin, H. Stability of related human and chicken Campylobacter jejuni genotypes after passage through chick intestine studied by pulsed-field gel electrophoresis. Appl. Environ. Microbiol. 1999, 65, 2272–2275. [Google Scholar] [CrossRef] [Green Version]
- Clark, C.G.; Price, L.; Ahmed, R.; Woodward, D.L.; Melito, P.L.; Rodgers, F.G.; Jamieson, F.; Ciebin, B.; Li, A.; Ellis, A. Characterization of waterborne outbreak-associated Campylobacter jejuni, Walkerton, Ontario. Emerg. Infect. Dis. 2003, 9, 1232–1241. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yin, T.; Du, X.; Yang, W.; Huang, J.; Jiao, X. Occurrence and genotypes of Campylobacter species in broilers during the rearing period. Avian Pathol. 2017, 46, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Iglesias-Torrens, Y.; Miró, E.; Guirado, P.; Llovet, T.; Muñoz, C.; Cerdà-Cuéllar, M.; Madrid, C.; Balsalobre, C.; Navarro, F. Population Structure, Antimicrobial Resistance, and Virulence-Associated Genes in Campylobacter jejuni Isolated from Three Ecological Niches: Gastroenteritis Patients, Broilers, and Wild Birds. Front. Microbiol. 2018, 9, 1676. [Google Scholar] [CrossRef] [Green Version]
- Wei, B.; Cha, S.-Y.; Kang, M.; Roh, J.-H.; Seo, H.-S.; Yoon, R.-H.; Jang, H.-K. Antimicrobial susceptibility profiles and molecular typing of Campylobacter jejuni and Campylobacter coli isolates from ducks in South Korea. Appl. Environ. Microbiol. 2014, 80, 7604–7610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieczorek, K.; Osek, J. Antimicrobial Resistance and Genotypes of Campylobacter jejuni from Pig and Cattle Carcasses Isolated in Poland During 2009–2016. Microb. Drug Resist. 2017, 24, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, L.; Gatica, M.A.; Riquelme, V.; Vergara, C.; Yanez, J.M.; San Martin, B.; Saenz, L.; Vidal, M.; Martinez, M.C.; Araya, P.; et al. Characterization of Antimicrobial Susceptibility and Its Association with Virulence Genes Related to Adherence, Invasion, and Cytotoxicity in Campylobacter jejuni and Campylobacter coli Isolates from Animals, Meat, and Humans. Microb. Drug Resist. 2016, 22, 432–444. [Google Scholar] [CrossRef]
- Humphrey, T.; O’Brien, S.; Madsen, M. Campylobacters as zoonotic pathogens: A food production perspective. Int. J. Food Microbiol. 2007, 117, 237–257. [Google Scholar] [CrossRef]
- Tresse, O.; Alvarez-Ordonez, A.; Connerton, I.F. Editorial: About the Foodborne Pathogen Campylobacter. Front. Microbiol. 2017, 8, 1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koolman, L.; Whyte, P.; Burgess, C.; Bolton, D. Distribution of virulence-associated genes in a selection of Campylobacter isolates. Foodborne Pathog Dis. 2015, 12, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Luangtongkum, T.; Jeon, B.; Han, J.; Plummer, P.; Logue, C.M.; Zhang, Q. Antibiotic resistance in Campylobacter: Emergence, transmission and persistence. Future Microbiol. 2009, 4, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Paudyal, N.; Yue, M. Antimicrobial Resistance in the “Dark Matter”. Clin. Infect. Dis. 2019, 69, 379–380. [Google Scholar] [CrossRef]
- Engberg, J.; Aarestrup, F.M.; Taylor, D.E.; Gerner-Smidt, P.; Nachamkin, I. Quinolone and macrolide resistance in Campylobacter jejuni and C. coli: Resistance mechanisms and trends in human isolates. Emerg. Infect. Dis. 2001, 7, 24–34. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, S.; He, M.; Zhang, Y.; Fu, Y.; Liang, H.; Jing, H.; Li, Y.; Ma, H.; Zhang, M. Prevalence and Molecular Characterization of Campylobacter spp. Isolated from Patients with Diarrhea in Shunyi, Beijing. Front. Microbiol. 2018, 9, 52. [Google Scholar] [CrossRef]
- Zhang, M.; Gu, Y.; He, L.; Ran, L.; Xia, S.; Han, X.; Li, H.; Zhou, H.; Cui, Z.; Zhang, J. Molecular typing and antimicrobial susceptibility profiles of Campylobacter jejuni isolates from north China. Appl. Environ. Microbiol. 2010, 59 Pt 10, 1171–1177. [Google Scholar] [CrossRef]
- Zhang, A.; Song, L.; Liang, H.; Gu, Y.; Zhang, C.; Liu, X.; Zhang, J.; Zhang, M. Molecular subtyping and erythromycin resistance of Campylobacter in China. Appl. Environ. Microbiol. 2016, 121, 287–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Ma, L.; Li, Y.; Jia, H.; Wei, J.; Shao, D.; Liu, K.; Shi, Y.; Qiu, Y.; Ma, Z. Antimicrobial Resistance of Campylobacter Species Isolated from Broilers in Live Bird Markets in Shanghai, China. Foodborne Pathog. Dis. 2017, 14, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Luo, Q.; Chen, Y.; Li, T.; Wen, G.; Zhang, R.; Luo, L.; Lu, Q.; Ai, D.; Wang, H.; et al. Molecular epidemiology, virulence determinants and antimicrobial resistance of Campylobacter spreading in retail chicken meat in Central China. Gut Pathog. 2016, 8, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbediwi, M.; Li, Y.; Paudyal, N.; Pan, H.; Li, X.; Xie, S.; Rajkovic, A.; Feng, Y.; Fang, W.; Rankin, S.C.; et al. Global Burden of Colistin-Resistant Bacteria: Mobilized Colistin Resistance Genes Study (1980–2018). Microorganisms 2019, 7, 461. [Google Scholar] [CrossRef] [Green Version]
- Law, J.W.-F.; Ab Mutalib, N.-S.; Chan, K.-G.; Lee, L.-H. Rapid methods for the detection of foodborne bacterial pathogens: Principles, applications, advantages and limitations. Front. Microbiol. 2015, 5, 770. [Google Scholar] [CrossRef] [Green Version]
- National Health and Family Planning Commission of the People’s Republic of China. Diagnostic Criteria for Infectious Diarrhea (WS 271–2007); People’s Medical Publishing House Co., Ltd.: Beijing, China, 2007. (In Chinese)
- Jiang, Z.; Paudyal, N.; Xu, Y.; Deng, T.; Li, F.; Pan, H.; Peng, X.; He, Q.; Yue, M. Antibiotic Resistance Profiles of Salmonella Recovered from Finishing Pigs and Slaughter Facilities in Henan, China. Front. Microbiol. 2019, 10, 1513. [Google Scholar] [CrossRef] [Green Version]
- National Health and Family Planning Commission of the People’s Republic of China. Diagnostic Criteria for Infectious Diarrhea (WS 287–2008); People’s Medical Publishing House Co., Ltd.: Beijing, China, 2008. (In Chinese)
- National Health and Family Planning Commission of the People’s Republic of China. Diagnostic Criteria for Infectious Diarrhea (WS 289–2008); People’s Medical Publishing House Co., Ltd.: Beijing, China, 2008. (In Chinese)
- Nachamkin, I.; Nguyen, P. Isolation of Campylobacter Species from Stool Samples by Use of a Filtration Method: Assessment from a United States-Based Population. J. Clin. Microbiol. 2017, 55, 2204. [Google Scholar] [CrossRef] [Green Version]
- Ribot, E.M.; Fitzgerald, C.; Kubota, K.; Swaminathan, B.; Barrett, T.J. Rapid pulsed-field gel electrophoresis protocol for subtyping of Campylobacter jejuni. J. Clin. Microbiol. 2001, 39, 1889–1894. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control (CDC). Standard Operating Procedure for PulseNet PGFE of Campylobacter Jejuni; 2011. Available online: http://www.cdc.gov/pulsenet/PDF/campylobacter-pfge-protocol-508c.pdf (accessed on 15 October 2019).
- Paudyal, N.; Pan, H.; Elbediwi, M.; Zhou, X.; Peng, X.; Li, X.; Fang, W.; Yue, M. Characterization of Salmonella Dublin isolated from bovine and human hosts. BMC Microbiol. 2019, 19, 226. [Google Scholar] [CrossRef]
- Elbediwi, M.; Pan, H.; Biswas, S.; Li, Y.; Yue, M. Emerging colistin resistance in Salmonella enterica serovar Newport isolates from human infections. Emerg. Microbes Infect. 2020, 9, 535–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.; Elbediwi, M.; Gu, G.; Yue, M. Genomic Characterization of New Variant of Hydrogen Sulfide (H2S)-Producing Escherichia coli with Multidrug Resistance Properties Carrying the mcr-1 Gene in China †. Antibiotics 2020, 9, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Parker, C.T.; Quinones, B.; Miller, W.G.; Horn, S.T.; Mandrell, R.E. Comparative genomic analysis of Campylobacter jejuni strains reveals diversity due to genomic elements similar to those present in C. jejuni strain RM1221. J. Clin. Microbiol. 2006, 44, 4125–4135. [Google Scholar] [CrossRef] [Green Version]
- Page, A.J.; Taylor, B.; Delaney, A.J.; Soares, J.; Seemann, T.; Keane, J.A.; Harris, S.R. SNP-sites: Rapid efficient extraction of SNPs from multi-FASTA alignments. Microb. Genom. 2016, 2, e000056. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Strains | Samples | ST* | PFGE | Genome Size | GC% | Contigs | N50 | Accession |
|---|---|---|---|---|---|---|---|---|---|
| 1 | CAM19-008 | Patient8 | 2988 | PA1 | 1,655,630 | 30.39 | 19 | 242,158 | SAMN12388802 |
| 2 | CAM19-010 | Patient10 | 2988 | PA1 | 1,652,624 | 30.39 | 13 | 332,935 | SAMN12388803 |
| 3 | CAM19-013 | Patient13 | 2988 | PA1 | 1,645,702 | 30.36 | 20 | 188,661 | SAMN12388804 |
| 4 | CAM19-014 | Patient14 | 2988 | PA1 | 1,649,319 | 30.35 | 48 | 69,212 | SAMN12388805 |
| 5 | CAM19-015 | Patient15 | 2988 | PA1 | 1,645,746 | 30.36 | 18 | 242,282 | SAMN12388806 |
| 6 | CAM19-016 | Patient16 | 2988 | PA1 | 1,644,870 | 30.35 | 19 | 255,151 | SAMN12388807 |
| 7 | CAM19-018 | Patient18 | 2988 | PA1 | 1,648,275 | 30.34 | 18 | 255,143 | SAMN12388808 |
| 8 | CAM19-019 | Patient19 | 2988 | PA1 | 1,647,897 | 30.34 | 20 | 255,112 | SAMN12388809 |
| 9 | CAM19-020 | Patient20 | 2988 | PA1 | 1,654,258 | 30.39 | 22 | 183,324 | SAMN12388810 |
| 10 | CAM19-021 | Patient21 | 2988 | PA1 | 1,645,811 | 30.36 | 18 | 179,689 | SAMN12388811 |
| 11 | CAM19-026 | Patient26 | 2988 | PA1 | 1,645,644 | 30.35 | 22 | 207,949 | SAMN12388812 |
| 12 | CAM19-027 | Patient27 | 2988 | PA1 | 1,650,856 | 30.34 | 42 | 112,156 | SAMN12388813 |
| 13 | CAM19-028 | Patient28 | 2988 | PA1 | 1,651,306 | 30.40 | 12 | 333,007 | SAMN12388814 |
| 14 | CAM19-033 | Patient33 | 8149 | PA2 | 1,678,861 | 30.49 | 21 | 120,767 | SAMN12388815 |
| 15 | CAM19-037 | Patient37 | 2988 | PA1 | 1,647,931 | 30.34 | 17 | 255,111 | SAMN12388816 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.; Elbediwi, M.; Zhou, X.; Shuai, H.; Lou, X.; Wang, H.; Li, Y.; Yue, M. Epidemiological and Genomic Characterization of Campylobacter jejuni Isolates from a Foodborne Outbreak at Hangzhou, China. Int. J. Mol. Sci. 2020, 21, 3001. https://doi.org/10.3390/ijms21083001
Yu H, Elbediwi M, Zhou X, Shuai H, Lou X, Wang H, Li Y, Yue M. Epidemiological and Genomic Characterization of Campylobacter jejuni Isolates from a Foodborne Outbreak at Hangzhou, China. International Journal of Molecular Sciences. 2020; 21(8):3001. https://doi.org/10.3390/ijms21083001
Chicago/Turabian StyleYu, Hua, Mohammed Elbediwi, Xiaohong Zhou, Huiqun Shuai, Xiuqin Lou, Haoqiu Wang, Yan Li, and Min Yue. 2020. "Epidemiological and Genomic Characterization of Campylobacter jejuni Isolates from a Foodborne Outbreak at Hangzhou, China" International Journal of Molecular Sciences 21, no. 8: 3001. https://doi.org/10.3390/ijms21083001